
Sorbicillinoids from Fungi and Their Bioactivities


 ,
,  and
and
Abstract
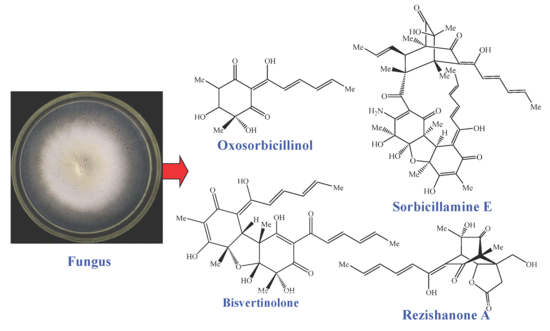
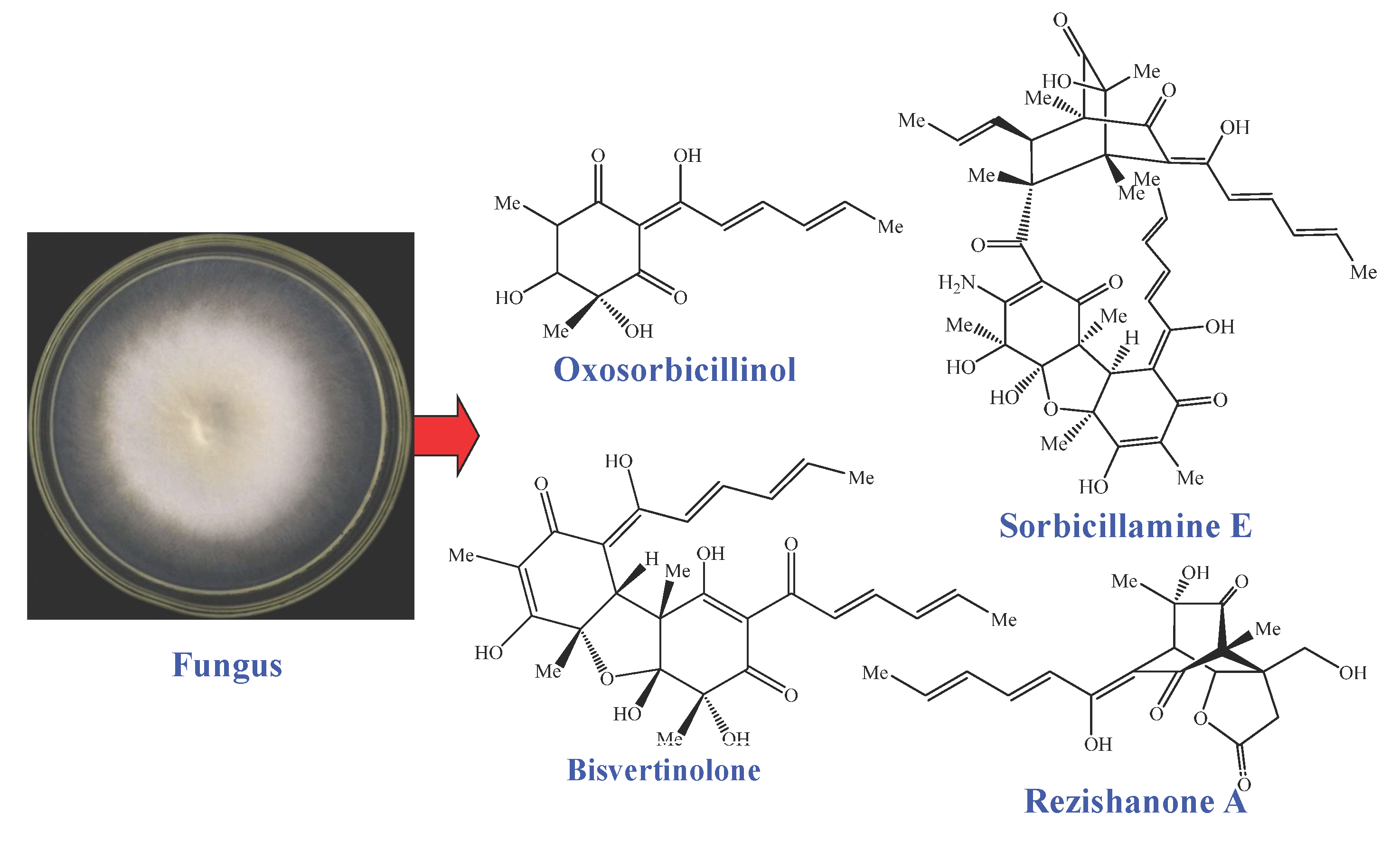
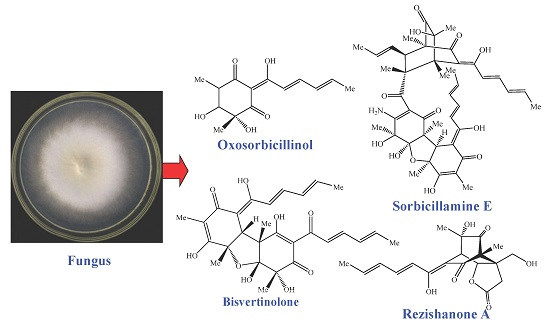
:
1. Introduction
2. Occurrence
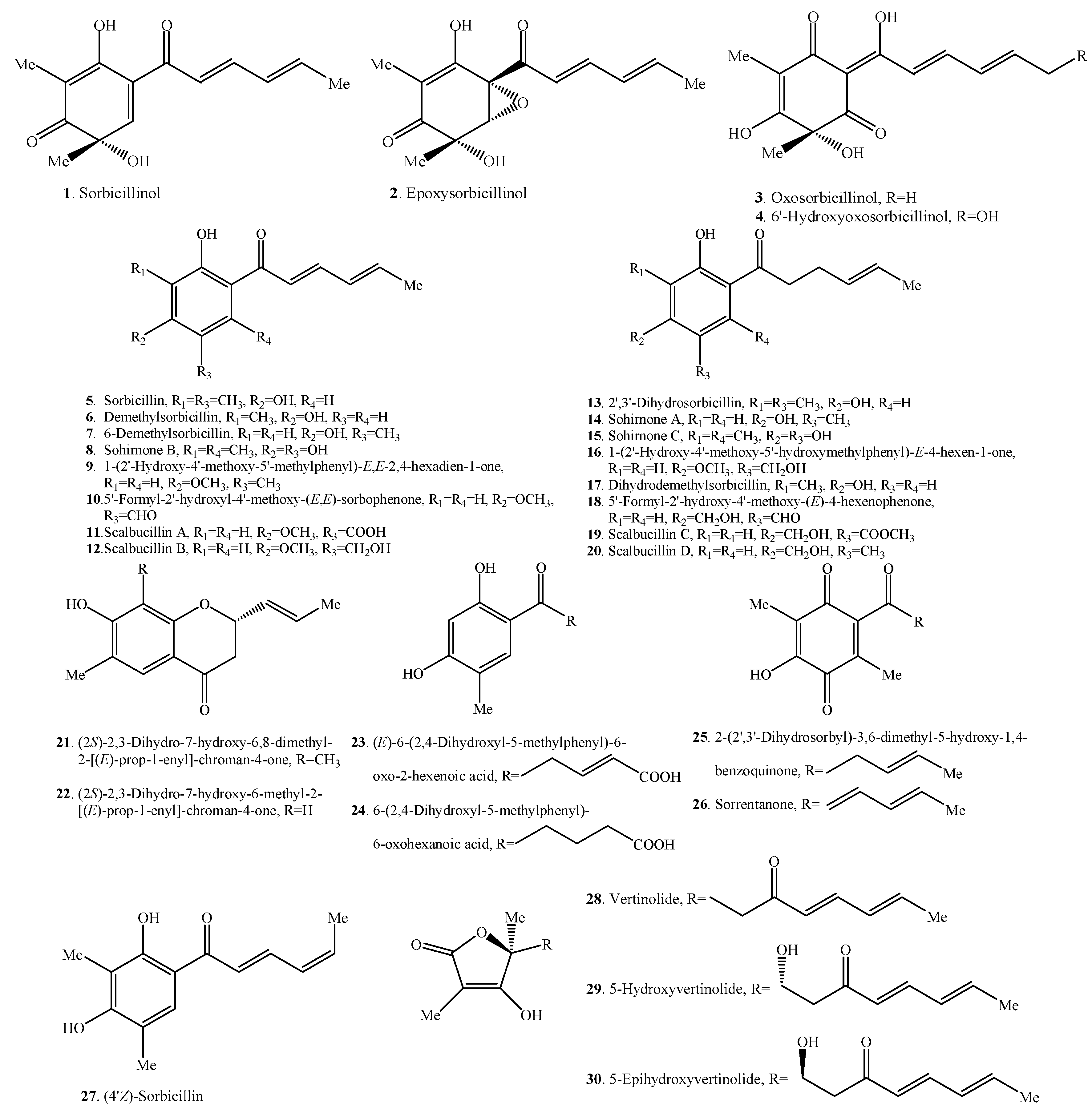
2.1. Monomeric Sorbicillinoids
2.2. Bisorbicillinoids
2.3. Trisorbicillinoids
2.4. Hybrid Sorbicillinoids
3. Biological Activities
3.1. Cytotoxic Activity
3.2. Antimicrobial Activity
3.3. Antiviral Activity
3.4. Antioxidant Activity
3.5. Other Biological Activities
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harned, A.M.; Volp, K.A. The sorbicillinoid family of natural products: Isolation, biosynthesis and synthetic studies. Nat. Prod. Rep. 2011, 28, 1790–1810. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, F.; Cai, S.; Zeng, X.; Xiao, X.; Gu, Q.; Zhu, W. Two new bisorbicillinoids isolated from a deep-sea fungus, Phialocephala sp. FL30r. J. Antibiot. 2007, 60, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Lateff, A.; Fisch, K.; Wright, A.D. Trichopyrone and other constituents from the marine sponge-derived fungus Trichoderma sp. Z. Naturforsch. 2009, 64c, 186–192. [Google Scholar] [CrossRef]
- Lan, W.-J.; Zhao, Y.; Xie, Z.-L.; Liang, L.-Z.; Shao, W.-Y.; Zhu, L.-P.; Yang, D.-P.; Zhu, X.-F.; Li, H.-J. Novel sorbicillin analogues from the marine fungus Trichoderma sp. associated with the seastar Acanthaster planci. Nat. Prod. Commun. 2012, 7, 1337–1340. [Google Scholar] [PubMed]
- Maskey, R.P.; Grün-Wollny, I.; Grün-Wollny, H. Sorbicillin analogues and related dimeric compounds from Penicillium notatum. J. Nat. Prod. 2005, 68, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, W.; Huang, Y.; Rong, X. Two acid sorbicillin analogues from saline lands-derived fungus Trichoderma sp. J. Antibiot. 2011, 64, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.-M.; Zhan, Z.-J.; Ding, Z.-S.; Shan, W.-G. Bioactive metabolites from Penicillium sp. P-1, a fungal endophyte in Huperzia serrata. Chem. Nat. Compd. 2011, 47, 541–544. [Google Scholar] [CrossRef]
- Cram, D.J.; Tishler, M. Mold metabolites. I. Isolation of several compounds from clinical penicillin. J. Am. Chem. Soc. 1948, 70, 4238–4239. [Google Scholar] [CrossRef] [PubMed]
- Cram, D.J. Mold metabolites. II. The structure of sorbicillin, a pigment produced by the mold Penicillium notatum. J. Am. Chem. Soc. 1948, 70, 4240–4243. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Peng, J.; Zhu, T.; Gu, Q.; Keyzers, R.A.; Li, D. Sorbicillamines A−E, nitrogen-containing sorbicillinoids from the deep-sea-derived fungus Penicillium sp. F23-2. J. Nat. Prod. 2013, 76, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Fahad, A.; Abood, A.; Fisch, K.M.; Osipow, A.; Davison, J.; Avramovic, M.; Butts, C.P.; Piel, J.; Simpson, T.J.; Cox, R.J. Oxidative dearomatisation: The key step of sorbicillinoid biosynthesis. Chem. Sci. 2014, 5, 523–527. [Google Scholar] [CrossRef] [PubMed]
- El-Elimat, T.; Raja, H.A.; Figueroa, M.; Swanson, S.M.; Falkinham, J.O., III; Lucas, D.M.; Grever, M.R.; Wani, M.C.; Pearce, C.J.; Oberlies, N.H. Sorbicillinoid analogs with cytotoxic and selective anti-Aspergillus activities from Scytalidium album. J. Antibiot. 2015, 68, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.-M.; Qi, F.-M.; Li, J.; Jiang, C.-X.; Hou, Y.; Shi, Y.-P.; Di, D.-L.; Zhang, J.-W.; Wu, Q.-X. Isolation of secondary metabolites from the soil-derived fungus Clonostachys rosea YRS-06, a biological control agent, and evaluation of antibacterial activity. J. Agric. Food Chem. 2016, 64, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Sugimoto, O.; Tanji, K.; Hirota, A. Identification of the quinol metabolite “Sorbicillinol”, a key intermediate postulated in bisorbicillinoid biosynthesis. J. Am. Chem. Soc. 2000, 122, 12606–12607. [Google Scholar] [CrossRef]
- Abe, N.; Yamamoto, K.; Arakawa, T.; Hirota, A. The biosynthesis of bisorbicillinoids: Evidence for a biosynthetic route from bisorbibutenolide and bisorbicillinolide. Chem. Commun. 2001, 2001, 23–24. [Google Scholar] [CrossRef]
- Abe, N.; Arakawa, T.; Yamamoto, K.; Hirota, A. Biosynthesis of bisorbicillinoid in Trichoderma sp. USF-2690; evidence for the biosynthetic pathway, via sorbicillinol, of sorbicillin, bisorbicillinol, bisorbibutenolide, and bisorbicillinolide. Bisosci. Biotechnol. Biochem. 2002, 66, 2090–2099. [Google Scholar] [CrossRef]
- Sugaya, K.; Koshino, H.; Hongo, Y.; Yasunaga, K.; Onose, J.; Yoshikawa, K.; Abe, N. The biosynthesis of sorbicillinoids in Trichoderma sp. USF-2690: Prospect for the existence of a common precursor to sorbicillinol and 5-epihydroxyvertinolide, a new sorbicillinoid member. Tetrahedron Lett. 2008, 49, 654–657. [Google Scholar] [CrossRef]
- Miller, R.F.; Huang, S. Isolation and structure of sorrentanone: A new tetrasubstituted quinone from Penicillium chrysogenum. J. Antibiot. 1995, 48, 520–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G. Two new benzoquinone derivatives and two new bisorbicillinoids were isolated from a marine-derived fungus Penicillium terrestre. J. Antibiot. 2005, 58, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Sperry, S.; Samuels, G.J.; Crews, P. Vertinoid polyketides from the saltwater culture of the fungus Trichoderma longibrachiatum separated from a Haliclona marine sponge. J. Org. Chem. 1998, 63, 10011–10014. [Google Scholar] [CrossRef]
- Bringmann, G.; Lang, G.; Gulder, T.A.M.; Tsuruta, H.; Mühlbacher, J.; Maksimenka, K.; Steffens, S.; Schaumann, K.; Stohr, R.; Wiese, J.; et al. The first sorbicillinoid alkaloids, the antileukemic sorbicillactones A and B, from a sponge-derived Penicillium chrysogenum strain. Tetrahedron 2005, 61, 7252–7265. [Google Scholar] [CrossRef]
- Komoda, T.; Nishikawa, M. 6′-Hydroxyoxosorbicillinol, a new lipoxygenase inhibitor and PGD2/LTB4 release suppressor from Penicillium sp. Biosci. Biotechnol. Biochem. 2012, 76, 1404–1406. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Yamamoto, K.; Hirota, A. Novel fungal metabolites, demethylsorbicillin and oxosorbicillinol, isolated from Trichoderma sp. USF-2690. Biosci. Biotechnol. Biochem. 2000, 64, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Itabashi, T.; Wakana, D.; Takeda, H.; Yaguchi, T.; Kawai, K.; Hosoe, T. Isolation and structure elucidation of new phthalide and phthalane derivatives, isolated as antimicrobial agents form Emericella sp. IFM57991. J. Antibiot. 2016, 69, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Arima, K.; Nakamura, H.; Komagata, K. Studies on variation of penicillin producing mold. Part II. Biochemical genetical studies on the yellow pigments losing mutation of chrysogenum Q 176 to pigmentless sultant Pen. chrysogenum Q 176. J. Agric. Chem. Soc. Jpn. 1953, 27, 345–348. [Google Scholar]
- Andrade, R.; Ayer, W.A.; Mebe, P.P. The metabolites of Trichoderma longibrachiatum. Part 1. Isolation of the metabolites and the structure of trichodimerol. Can. J. Chem. 1992, 70, 2526–2535. [Google Scholar] [CrossRef]
- Du, L.; Zhu, T.; Li, L.Y.; Cai, S.; Zhao, B.; Gu, Q. Cytotoxic sorbicillinoids and bisorbicillinoids from a marine-derived fungus Trichoderma sp. Chem. Pharm. Bull. 2009, 57, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Zhao, L.X.; Chen, Y.W.; Huang, R.; Miao, C.P.; Wang, J. Sesquiterpenoids from the endophytic fungus Trichoderma sp. PR-35 of Paeonia delavayi. Chem. Biodivers. 2011, 8, 1717–1723. [Google Scholar]
- Abe, N.; Murata, T.; Hirota, A. Novel oxidized sorbicillin dimers with 1,1-diphenyl-2-picrylhydrazyl-radial scavenging activity from a fungus. Biosci. Biotechnol. Biochem. 1998, 62, 2120–2126. [Google Scholar] [CrossRef]
- Yao, Y.; Li, J.; Jiang, C.-S.; Zhao, X.-X.; Miao, Z.-H.; Liu, H.-T.; Zheng, P.; Yao, W.-X.; Li, W.-Q. Trichodimerol and sorbicillin induced apoptosis of HL-60 cells is mediated by reactive oxygen species. Pharmazie 2015, 70, 394–398. [Google Scholar] [PubMed]
- Trifonov, L.S.; Dreiding, A.S.; Hoesch, L.; Rast, D.M. Isolation of four hexaketides from Verticillium intertextum. Helv. Chim. Acta 1981, 64, 1843–1846. [Google Scholar] [CrossRef]
- Trifonov, L.S.; Bieri, J.H.; Prewo, R.; Dreiding, A.S. Isolation and structure elucidation of three metabolites from Verticillium intertextum: Sorbicillin, dihydrosorbicillin and bisvertinoquinol. Tetrahedron 1983, 39, 4243–4256. [Google Scholar] [CrossRef]
- Reátegui, R.F.; Wicklow, D.T.; Gloer, J.B. Phaeofurans and sorbicillin analogues from a fungicolous Phaeoacremonium species (NRRL 32148). J. Nat. Prod. 2006, 69, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Geigert, J.; Stermitz, F.R.; Schroeder, H.A. Two new natural substituted hexenophenones from the fungus Scytalidium. Tetrahedron 1973, 29, 2343–2345. [Google Scholar] [CrossRef]
- Bringmann, G.; Lang, G.; Bruhn, T.; Schäffler, K.; Steffens, S.; Schmaljohann, R.; Wiese, J.; Imhoff, J.F. Sorbifuranones A-C, sorbicillinoid metabolites from Penicillium strains isolated from Mediterranean sponges. Tetrahedron 2010, 66, 9894–9901. [Google Scholar] [CrossRef]
- Li, D.; Cai, S.; Zhu, T.; Wang, F.; Xiao, X.; Gu, Q. New cytotoxic metabolites from a deep-sea-derived fungus, Phialocephala sp., strain FL30r. Chem. Biodivers. 2011, 8, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Neumann, K.; Abdel-Lateff, A.; Wright, A.D.; Kehraus, S.; Krick, A.; König, G.M. Novel sorbicillin derivatives with an unprecedented carbon skeleton from the sponge-derived fungus Trichoderma species. Eur. J. Org. Chem. 2007, 2007, 2268–2275. [Google Scholar] [CrossRef]
- Trifonov, L.S.; Bieri, J.H.; Prewo, R.; Dreiding, A.S.; Rast, D.M.; Hoesch, L. The constitution of vertinolide, a new derivative of tetronic acid, produced by Verticillium intertextum. Tetrahedron 1982, 38, 397–403. [Google Scholar] [CrossRef]
- Andrade, R.; Ayer, W.A.; Trifonov, L.S. The metabolites of Trichoderma longibrachiatum III. Two new tetronic acids: 5-hydroxyvertinolide and bislongiquinolide. Aust. J. Chem. 1997, 50, 255–257. [Google Scholar] [CrossRef]
- Koyama, N.; Ohshiro, T.; Tomoda, H.; Ōmura, S. Fungal isobisvertinol, a new inhibitor of lipid droplet accumulation in mouse macrophages. Org. Lett. 2007, 9, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, L.S.; Hilpert, H.; Floersheim, P.; Dreiding, A.S.; Rast, D.M.; Skrivanova, R.; Hoesch, L. Bisvertinols: A new group of dimeric vertinoids from Verticillium intertextum. Tetrahedron 1986, 42, 3157–3179. [Google Scholar] [CrossRef]
- Kontani, M.; Sakagami, Y.; Marumo, S. Frist β-1,6-giucan biosynthesis inhibitor, bisvertinolone isolated from fungus, Acremonium stricturn and its absolute stereochemistry. Tetrahedron Lett. 1994, 35, 2577–2580. [Google Scholar] [CrossRef]
- Ueda, J.; Hashimoto, J.; Inaba, S.; Takagi, M.; Shin-ya, K. JBIR-59, a new sorbicillinoid, from a marine-derived fungus Penicillium citrinum SpI080624G1f01. J. Antibiot. 2010, 63, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Murata, T.; Hirota, A. Novel DPPH radical scavengers, bisorbicillinol and demethyltrichodimerol, from a fungus. Biosci. Biotechnol. Biochem. 1998, 62, 661–666. [Google Scholar] [CrossRef]
- Gao, Q.; Leet, J.E.; Thomas, S.T.; Matson, J.A. Crystal structure of trichodimerol. J. Nat. Prod. 1995, 58, 1817–1821. [Google Scholar] [CrossRef]
- Warr, G.A.; Veitch, J.A.; Walsh, A.W.; Hesler, G.A.; Pirnik, D.M.; Leet, J.E.; Lin, P.-F.M.; Medina, I.A.; McBrien, K.D.; Forenza, S.; et al. BMS-182123, a fungal metabolite that inhibits the production of TNF-α by macrophage and monocytes. J. Antibiot. 1996, 49, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G. Dihydrotrichodimerol and tetrahydrotrichodimerol, two new bisorbicillinoids, from a marine-derived Penicillium terrestre. J. Antibiot. 2005, 58, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Ganassi, S.; Altomare, C.; Favilla, M.; Cristofaro, A.D.; Vitagliano, S.; Sabatini, M.A. Bisorbicillinoids produced by the fungus Trichoderma citrinoviride affect feeding preference of the aphid Schizaphis graminum. J. Chem. Ecol. 2009, 35, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Shirota, O.; Pathak, V.; Hossain, C.F.; Sekita, S.; Takatori, K.; Satake, M. Structural elucidation of trichotetronines: Polyketides possessing a bicycle [2.2.2] octane skeleton with a tetronic acid moiety isolated from Trichoderma sp. J. Chem. Soc. Perk. Trans. 1 1997, 1997, 2961–2964. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.H.; Cai, X.F.; Shin, J.C.; Lee, K.; Hong, Y.-S.; Lee, J.J. Fungal metabolites, sorbicillinoid polyketides and their effects on the activation of peroxisome proliferator-activated receptor γ. J. Antibiot. 2005, 58, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Balde, E.S.; Andolfi, A.; Bruyère, C.; Cimmino, A.; Lamoral-Theys, D.; Vurro, M.; Damme, M.V.; Altomare, C.; Mathieu, V.; Kiss, R.; et al. Investigations of fungal secondary metabolites with potential anticancer activity. J. Nat. Prod. 2010, 73, 969–971. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Murata, T.; Yamamoto, K.; Hirota, A. Bisorbibetanone, a novel oxidized sorbicillin dimer, with 1,1-diphenyl-2-picrylhydrazyl radical scavenging activity from a fungus. Tetrahedron Lett. 1999, 40, 5203–5206. [Google Scholar] [CrossRef]
- Washida, K.; Abe, N.; Sugiyama, Y.; Hirota, A. Novel DPPH radical scavengers, demethylbisorbibutenolide and trichopyrone, from a fungus. Biosci. Biotech. Biochem. 2007, 71, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.; Ayer, W.A.; Trifonov, L.S. The metabolites of Trichoderma longibrachiatum Part II. The structures of trichodermolide and sorbiquinol. Can. J. Chem. 1996, 74, 371–379. [Google Scholar] [CrossRef]
- Li, D.; Wang, F.; Xiao, X.; Fang, Y.; Zhu, T.; Gu, Q.; Zhu, W. Trisorbicillinone A, a novel sorbicillin trimer, from a deep sea fungus, Phialocephala sp. FL30r. Tetrahedron Lett. 2007, 48, 5235–5238. [Google Scholar] [CrossRef]
- Li, D.; Cai, S.; Zhu, T.; Wang, F.; Xiao, X.; Gu, Q. Three new sorbicillin trimers, trisorbicillinones B, C, and D, from a deep ocean sediment derived fungus, Phialocephala sp. FL30r. Tetrahedron 2010, 66, 5101–5106. [Google Scholar] [CrossRef]
- Cabrera, G.M.; Butler, M.; Rodriguez, A.; Godeas, A.; Haddad, R.; Eberlin, M.N. A sorbicillinoid urea from an intertidal Paecilomyces marquandii. J. Nat. Prod. 2006, 69, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, L.; Zhu, T.; Kurtán, T.; Mándi, A.; Zhao, Z.; Li, J.; Gu, Q. Chloctanspirones A and B, novel chlorinated polyketides with an unprecedented skeleton, from marine sediment derived fungus Penicillium terrestre. Tetrahedron 2011, 67, 7913–7918. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, X.; Du, L.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Sorbicatechols A and B, antiviral sorbicillinoids from the marine- derived fungus Penicillium chrysogenum PJX-17. J. Nat. Prod. 2014, 77, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Washida, K.; Abe, N.; Sugiyama, Y.; Hirta, A. Novel secondary metabolites, spirosorbicillinols A, B, and C, from a fungus. Biosci. Biotech. Biochem. 2009, 73, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, T.; Ding, Y.; Khan, I.A.; Gu, Q.; Li, D. Sorbiterrin A, a novel sorbicillin derivative with cholinesterase inhibition activity from the marine-derived fungus Penicillium terrestre. Tetrahedron Lett. 2012, 53, 325–328. [Google Scholar] [CrossRef]
- Kawahara, T.; Takagi, M.; Shin-ya, K. JBIR-124: A novel antioxidative agent from a marine sponge-derived fungus Penicillium citrinum SpI080624G1f01. J. Antibiot. 2012, 65, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Acharya, A.B.; Subramaniyan, S.; Babu, S.; Kulkarni, S.; Narayanappa, R. Secondary metabolites extracted from marine sponge associated Comamonas testosterone and Citrobacter freundii as potential antimicrobials against MDR pathogens and hypothetical leads for VP40 matrix protein of Ebola virus: An in vitro and in silico investigation. J. Biomol. Struct. Dyn. 2016, 34. [Google Scholar]
- Finkel, T. Radical medicine: Treating ageing to cure disease. Nat. Rev. Mol. Cell Biol. 2005, 6, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Hirota, A. Chemical studies of the radical scavenging mechanism of bisorbicillinol using the 1,1-diphenyl-2-picrylhydrzyl radical. Chem. Commun. 2002, 2002, 662–663. [Google Scholar] [CrossRef]
- Mazzucco, C.E.; Warr, G. Trichodimerol (BMS-182123) inhibits lipopolysaccharide-induced eicosanoid secretion in THP-1 human monocytic cells. J. Leukocyte Biol. 1996, 60, 271–277. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sorbicillinoid | Fungus and its Origin | Ref. |
|---|---|---|
| Sorbicillinol (1) | Trichoderma sp. USF-2690 from a soil sample | [14] |
| Epoxysorbicillinol (2) | Trichoderma longibrachiatum from the sponge Haliclona sp. | [20] |
| Oxosorbicillinol (3) | Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [21] |
| Penicillium notatum from a benchtop contamination | [5] | |
| Penicillium sp. 06T121 from a soil sample | [22] | |
| Trichoderma sp. USF-2690 from a soil sample | [23] | |
| 6′-Hydroxyoxosorbicillinol (4) | Penicillium sp. 06T121 from a soil sample | [22] |
| Sorbicillin (5) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Emericella sp. IFM57991 and its origin was not clear | [24] | |
| Penicillium chrysogenum Q176 and its origin was not clear | [25] | |
| Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [11,21] | |
| Penicillium notatum and its origin was not clear | [8,9] | |
| Penicillium sp. P-1 as an endophyte from the stems of Huperzia serrata | [7] | |
| Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26] | |
| Trichoderma sp. from the seastar Acanthaster planci | [4] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Trichoderma sp. PR-35 as an endophyte from Paeonia delavayi | [28] | |
| Trichoderma sp. USF-2690 from a soil sample | [29] | |
| Trichothecium sp. from a marine sediment | [30] | |
| Verticillium intertextum and its origin was not clear | [31,32] | |
| Demethylsorbicillin (6) | Trichoderma sp. USF-2690 from a soil sample | [23] |
| 6-Demethylsorbicillin (7) | Trichoderma sp. f-13 from a marine sediment | [27] |
| Sohirnone B (8) | Penicillium notatum from a benchtop contamination | [5] |
| 1-(2′-Hydroxy-4′-methoxy-5′-methylphenyl)-E,E-2,4-hexadien-1-one (9) | Phaeoacremonium sp. NRRL32148 from the surface of stromata of Hypoxylon truncatum formed on a dead hardwood branch | [33] |
| Scytalidium album MSX51631 from a soil sample | [12] | |
| 5′-Formyl-2′-hydroxyl-4′-methoxy-(E,E)-sorbophenone (10) | Phaeoacremonium sp. NRRL32148 from the surface of stromata of Hypoxylon truncatum formed on a dead hardwood branch | [33] |
| Scytalidium album MSX51631 from a soil sample | [12] | |
| Scytalidium sp. FY as an immunizing commensal of Douglasfir utility poles | [34] | |
| Scalbucillin A (11) | Scytalidium album MSX51631 from a soil sample | [12] |
| Scalbucillin B (12) | Scytalidium album MSX51631 from a soil sample | [12] |
| 2′,3′-Dihydrosorbicillin (13) | Penicillium chrysogenum R03-8/4 from the sponge Tethya aurantium | [35] |
| Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [11] | |
| Penicillium notatum from a benchtop contamination | [5] | |
| Penicillium sp. P-1 as an endophyte from the stems of Huperzia serrata | [7] | |
| Trichoderma sp. from the seastar Acanthaster planci | [4] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Verticillium intertextum from a laboratory contaminant | [31,32] | |
| Sohirnone A (14) | Penicillium notatum from a benchtop contamination | [5] |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Sohirnone C (15) | Penicillium notatum from a benchtop contamination | [5] |
| 1-(2′-Hydroxy-4′-methoxy-5′-hydroxymethylphenyl)-E-4-hexen-1-one (16) | Phaeoacremonium sp. from the surface of stromata of Hypoxylon truncatum formed on a dead hardwood branch | [33] |
| Scytalidium album MSX51631 from a soil sample | [12] | |
| Dihydrodemethylsorbicillin (17) | Phialocephala sp. FL30r from a deep sea sediment | [36] |
| 5′-Formyl-2′-hydroxy-4′-methoxy-(E)-4-hexenophenone (18) | Scytalidium album MSX51631 from a soil sample | [12] |
| Scytalidium sp. FY as an immunizing commensal of Douglasfir utility poles | [34] | |
| Scalbucillin C (19) | Scytalidium album MSX51631 from a soil sample | [12] |
| Scalbucillin D (20) | Scytalidium album MSX51631 from a soil sample | [12] |
| (2S)-2,3-Dihydro-7-hydroxy-6,8-dimethyl-2-[(E)-prop-1-enyl]-chroman-4-one (21) | Trichoderma sp. from the seastar Acanthaster planci | [4] |
| Penicillium sp. P-1 as an endophyte from the stems of Huperzia serrata | [7] | |
| (2S)-2,3-Dihydro-7-hydroxy-6-methyl-2- [(E)-prop-1-enyl]-chroman-4-one (22) | Trichoderma sp. from the seastar Acanthaster planci | [4] |
| (E)-6-(2,4-Dihydroxyl-5-methylphenyl)-6-oxo-2-hexenoic acid (23) | Trichoderma sp. JH8 from the soil of saline lands | [6] |
| 6-(2,4-Dihydroxyl-5-methylphenyl)-6-oxohexanoic acid (24) | Trichoderma sp. JH8 from the soil of saline lands | [6] |
| 2-(2′,3′ -Dihydrosorbyl)-3,6-dimethyl-5-hydroxy-1,4-benzoquinone (25) | Penicillium terrestre from a marine sediment | [19] |
| Sorrentanone = 3-hydroxy-2,5-dimethyl-6-(1′-oxo-2′,4′-dienylhexyl)-1,4-benzoquione (26) | Penicillium chrysogenum SC13887 and its origin was not clear | [18] |
| (4’Z)-Sorbicillin (27) | Trichoderma sp. from the seastar Acanthaster planci | [4] |
| Vertinolide (28) | Trichoderma viride from the sponge Agelas dispar | [3] |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Verticillium intertextum from a laboratory contaminant | [31,38] | |
| 5-Hydroxyvertinolide (29) | Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [39] |
| 5-Epihydroxyvertinolide (30) | Trichoderma sp. USF-2690 from a soil sample | [17] |
| Sorbicillinoid | Fungus and Its Origin | Ref. |
|---|---|---|
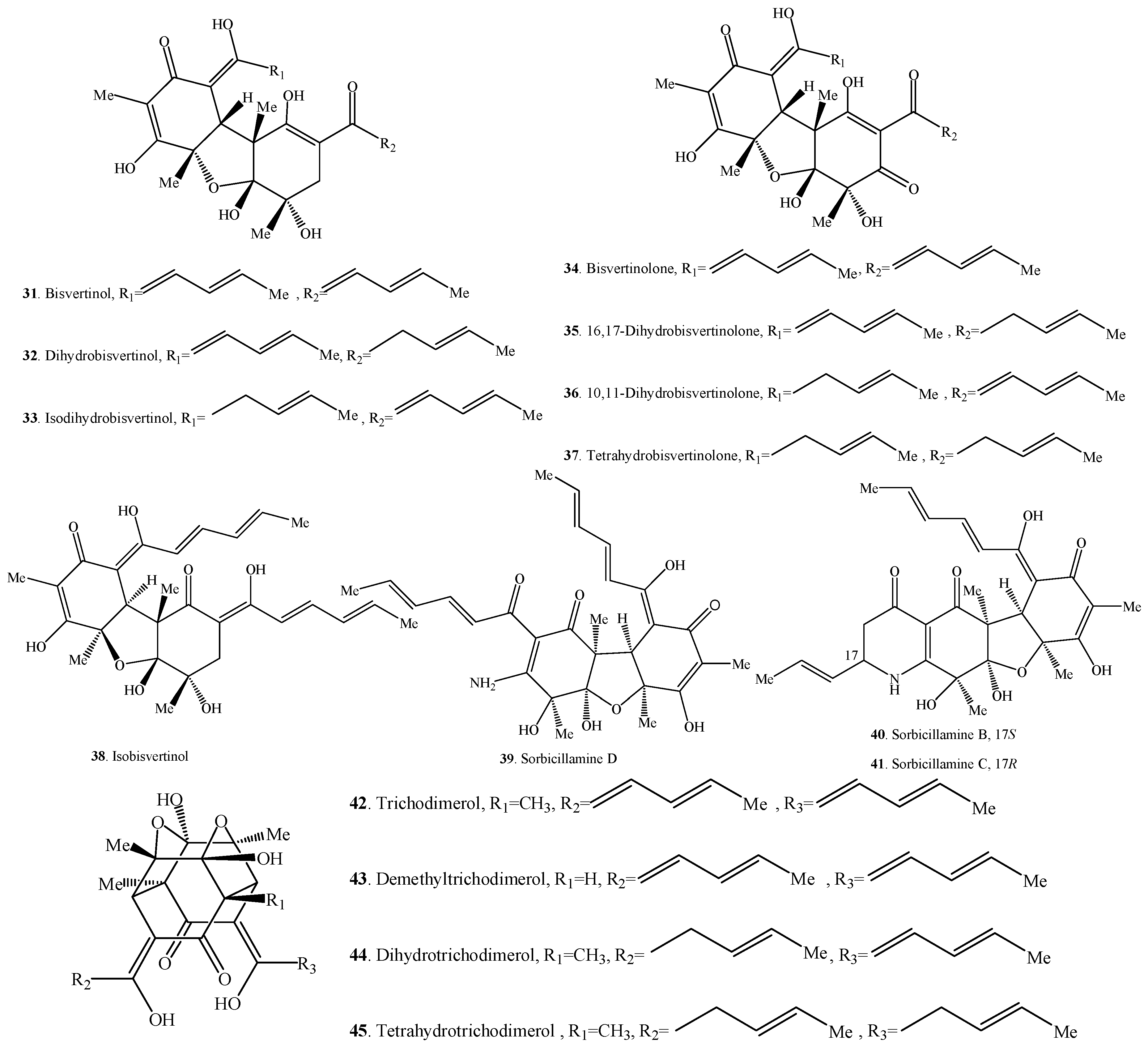
| Bisvertinol (31) | Aspergillus sp. FKI-1746 from a mangrove slurry sample | [40] |
| Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26] | |
| Trichoderma viride from the sponge Agelas dispar | [3] | |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Verticillium intertextum from a laboratory contaminant | [41] | |
| Dihydrobisvertinol (32) | Aspergillus sp. FKI-1746 from a mangrove slurry sample | [40] |
| Verticillium intertextum from a laboratory contaminant | [41] | |
| Isodihydrobisvertinol (33) Bisvertinolone (34) | Verticillium intertextum from a laboratory contaminant | [41] |
| Acremonium strictum and its origin was not clear | [42] | |
| Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [21] | |
| Penicillium citrinum SpI080624G1f01 from a marine sponge | [43] | |
| Penicillium notatum from a benchtop contamination | [5] | |
| Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Trichoderma sp. JH8 from the soil of saline lands | [6] | |
| Trichoderma sp. USF-2690 isolated from a soil sample | [44] | |
| Verticillium intertextum from a laboratory contaminant | [41] | |
| 16,17-Dihydrobisvertinolone (35) | Penicillium terrestre from a marine sediment | [19] |
| 10,11-Dihydrobisvertinolone (36) | Trichoderma sp. f-13 from a marine sediment | [27] |
| Tetrahydrobisvertinolone (37) | Penicillium terrestre from a marine sediment | [19] |
| Isobisvertinol (38) | Aspergillus sp. FKI-1746 from a mangrove slurry sample | [40] |
| Sorbicillamine D (39) | Penicillium sp. F23-2 from a deep-sea sediment | [10] |
| Sorbicillamine B (40) | Penicillium sp. F23-2 from a deep-sea sediment | [10] |
| Sorbicillamine C (41) | Penicillium sp. F23-2 from a deep-sea sediment | [10] |
| Trichodimerol = MS-182123 (42) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Penicillium chrysogenum V39673 and its origin was not clear | [45,46] | |
| Penicillium citrinum SpI080624G1f01 from a marine sponge | [43] | |
| Penicillium terrestre from a marine sediment | [47] | |
| Trichoderma citrinoviride ITEM 4484 from the soil under the tree Abies sp. | [48] | |
| Trichoderma viride from the sponge Agelas dispar | [3] | |
| Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26] | |
| Trichoderma sp. from the straws of rice | [49] | |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Trichoderma sp. JH8 from the soil of saline lands | [6] | |
| Trichoderma sp. USF-2690 from a soil sample | [44] | |
| Trichothecium sp. from a marine sediment | [30] | |
| Unidentified fungus B00853 from a soil sample | [50] | |
| Demethyltrichodimerol (43) | Trichoderma sp. USF-2690 isolated from a soil sample | [44] |
| Dihydrotrichodimerol (44) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Penicillium terrestre from a marine sediment | [47] | |
| Trichoderma citrinoviride ITEM 4484 from the soil under the tree Abies sp. | [48,51] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Unidentified fungus B00853 from a soil sample | [50] | |
| Tetrahydrotrichodimerol (45) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Penicillium terrestre from a marine sediment | [47] | |
| Bisorbibetanone (46) | Trichoderma sp. USF-2690 isolated from a soil sample | [52] |
| Bisvertinoquinol (47) | Penicillium notatum from a benchtop contamination | [5] |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Verticillium intertextum from a laboratory contaminat | [31,32] | |
| Bisorbicillinol (48) | Penicillium notatum from a benchtop contamination | [5] |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Trichoderma sp. USF-2690 from a soil sample | [44] | |
| Bislongiquinolide = Bisorbibutenolide = Trichotetronine (49) | Penicillium citrinum SpI080624G1f01 from the sponge Demospongiae sp. | [43] |
| Trichoderma citrinoviride ITEM 4484 from the soil under the tree Abies sp. | [48,51] | |
| Trichoderma longibrachiatum from the sponge Haliclona sp. | [20] | |
| Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26,39] | |
| Trichoderma viride from the sponge Agelas dispar | [3] | |
| Trichoderma sp. from the straws of rice plant | [49] | |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Trichoderma sp. f-13 from a marine sediment | [27] | |
| Trichoderma sp. USF-2690 from a soil sample | [29] | |
| 24,25-Dihydrotrichotetronine = 16,17-Dihydrobislongiquinolide (50) | Trichoderma citrinoviride ITEM 4484 from the soil under the tree Abies sp. | [48,51] |
| Trichoderma sp. from the straws of rice plant | [49] | |
| Demethylbisorbibutenolide (51) | Trichoderma sp. USF-4860 from a soil sample | [53] |
| Sorbiquinol (52) | Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26,54] |
| Oxosorbiquinol (53) | Phialocephala sp. FL30r from a deep-sea sediment | [2] |
| Dihydrooxosorbiquinol (54) | Phialocephala sp. FL30r from a deep-sea sediment | [2] |
| Bisorbicillinolide (55) | Trichoderma sp. USF-2690 from a soil sample | [29] |
| Dihydrotrichodermolide (56) | Phialocephala sp. FL30r from a deep-sea sediment | [36] |
| Trichodermolide (57) | Trichoderma longibrachiatum UAMH 4159 and its origin was not clear | [26,54] |
| Tetrahydrotrichodimer ether (58) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Dihydrotrichodimer ether A (59) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Dihydrotrichodimer ether B (60) | Clonostachys rosea YRS-06 from a soil sample | [13] |
| Sorbicillinoid | Fungus and Its Origin | Ref. |
|---|---|---|
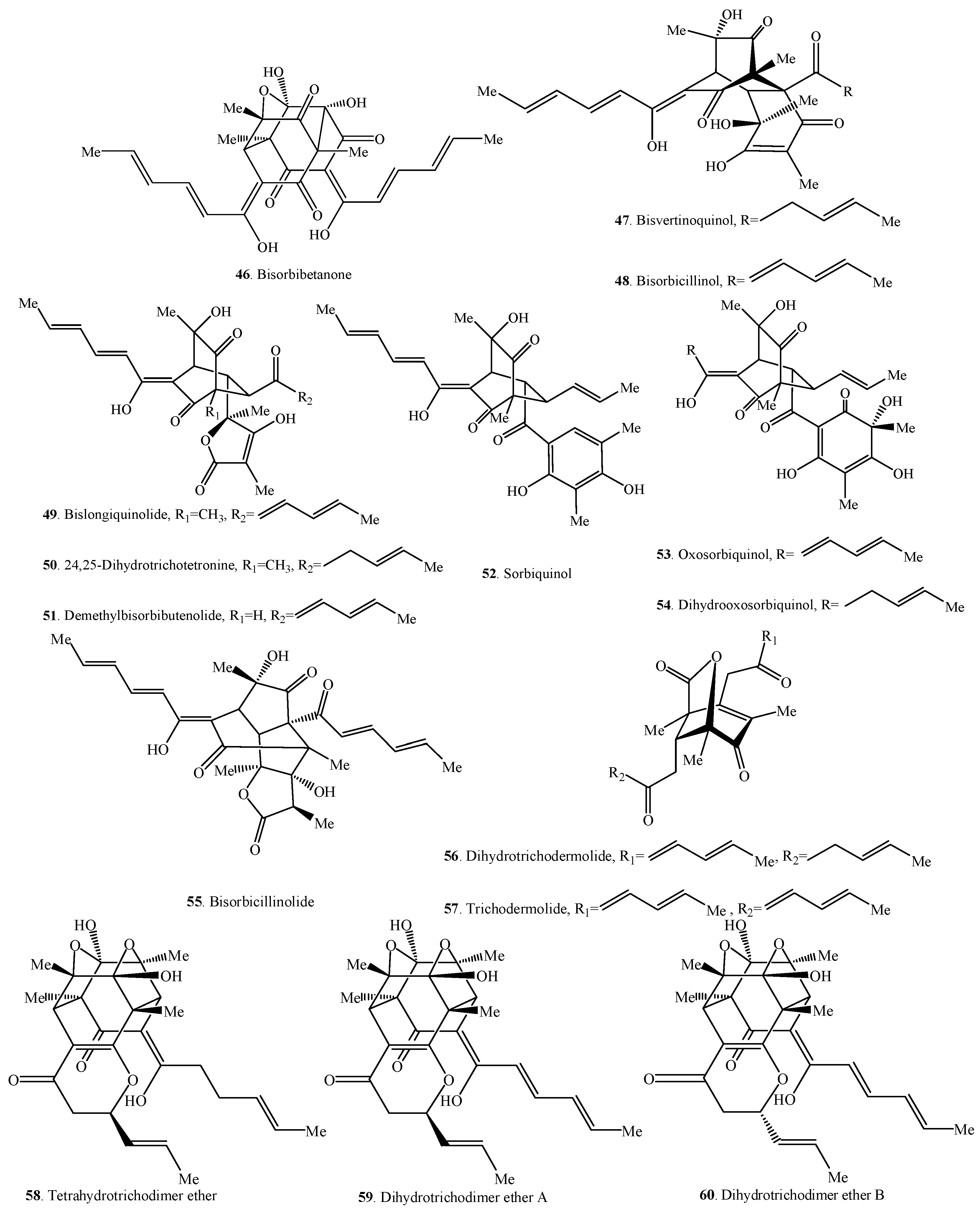
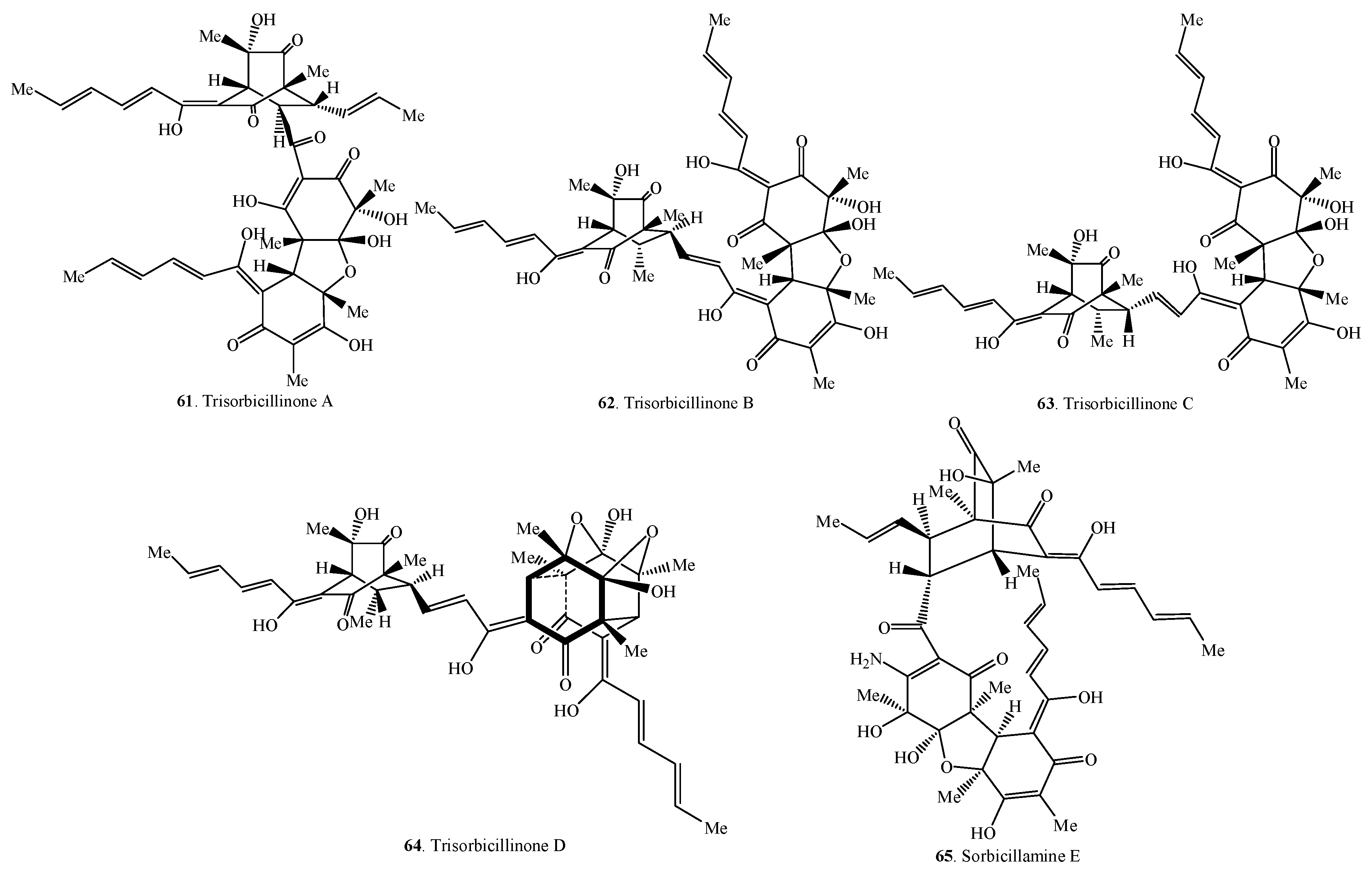
| Trisorbicillinone A (61) | Phialocephala sp. FL30r from a deep-sea sediment | [55] |
| Trisorbicillinone B (62) | Phialocephala sp. FL31r from a deep-sea sediment | [56] |
| Trisorbicillinone C (63) | Phialocephala sp. FL32r from a deep-sea sediment | [56] |
| Trisorbicillinone D (64) | Phialocephala sp. FL33r from a deep-sea sediment | [56] |
| Sorbicillamine E (65) | Penicillium sp. F23-2 from a deep-sea sediment | [10] |
| Sorbicillinoid | Fungus and Its Origin | Ref. |
|---|---|---|
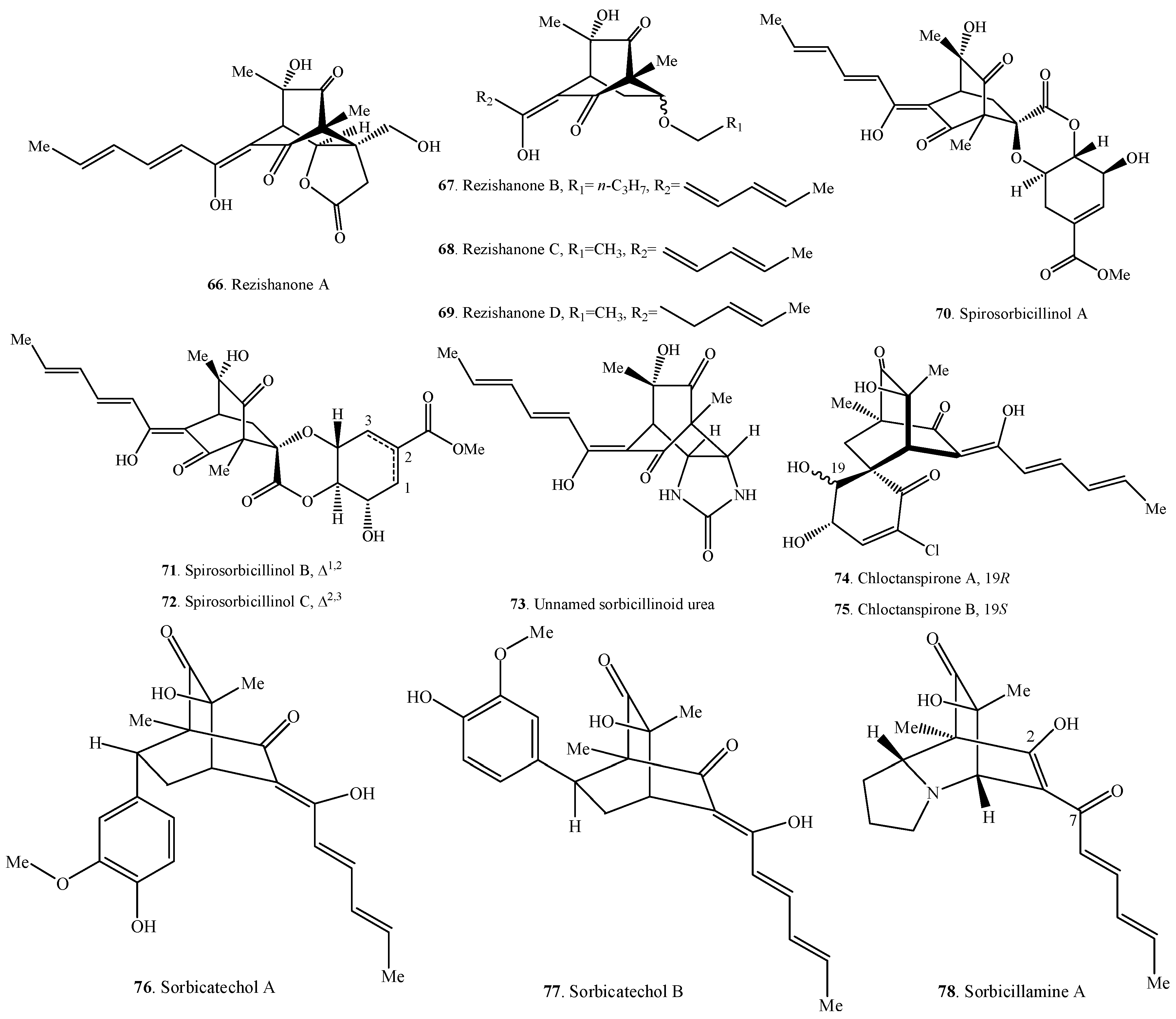
| Rezishanone A (66) | Penicillium notatum from a benchtop contamination | [5] |
| Rezishanone B (67) | Penicillium notatum from a benchtop contamination | [5] |
| Rezishanone C = Sorbivinetone (68) | Penicillium chrysogenum isolated from the sponge Ircinia fasciculata | [21] |
| Penicillium notatum from a benchtop contamination | [5] | |
| Trichoderma viride from the sponge Agelas dispar | [3] | |
| Trichoderma sp. isolated from the sponge Agelas dispar | [37] | |
| Unidentified fungus B00853 from a soil sample | [50] | |
| Rezishanone D (69) | Penicillium notatum from a benchtop contamination | [5] |
| Unidentified fungus B00853 collected from a soil sample | [50] | |
| Spirosorbicillinol A (70) | Trichoderma sp. USF-4860 from a soil sample | [60] |
| Spirosorbicillinol B (71) | Trichoderma sp. USF-4860 from a soil sample | [60] |
| Spirosorbicillinol C (72) | Trichoderma sp. USF-4860 from a soil sample | [60] |
| Unnamed sorbicillinoid urea (73) | Paecilomyces marquandii BAFC 486 from a marine sediment | [57] |
| Chloctanspirone A (74) | Penicillium terrestre from a marine sediment | [58] |
| Chloctanspirone B (75) | Penicillium terrestre from a marine sediment | [58] |
| Sorbicatechol A (76) | Penicillium chrysogenum PJX-17 from a marine sediment | [59] |
| Sorbicatechol B (77) | Penicillium chrysogenum PJX-17 from a marine sediment | [59] |
| Sorbicillamine A (78) | Penicillium sp. F23-2 from a deep-sea sediment | [10] |
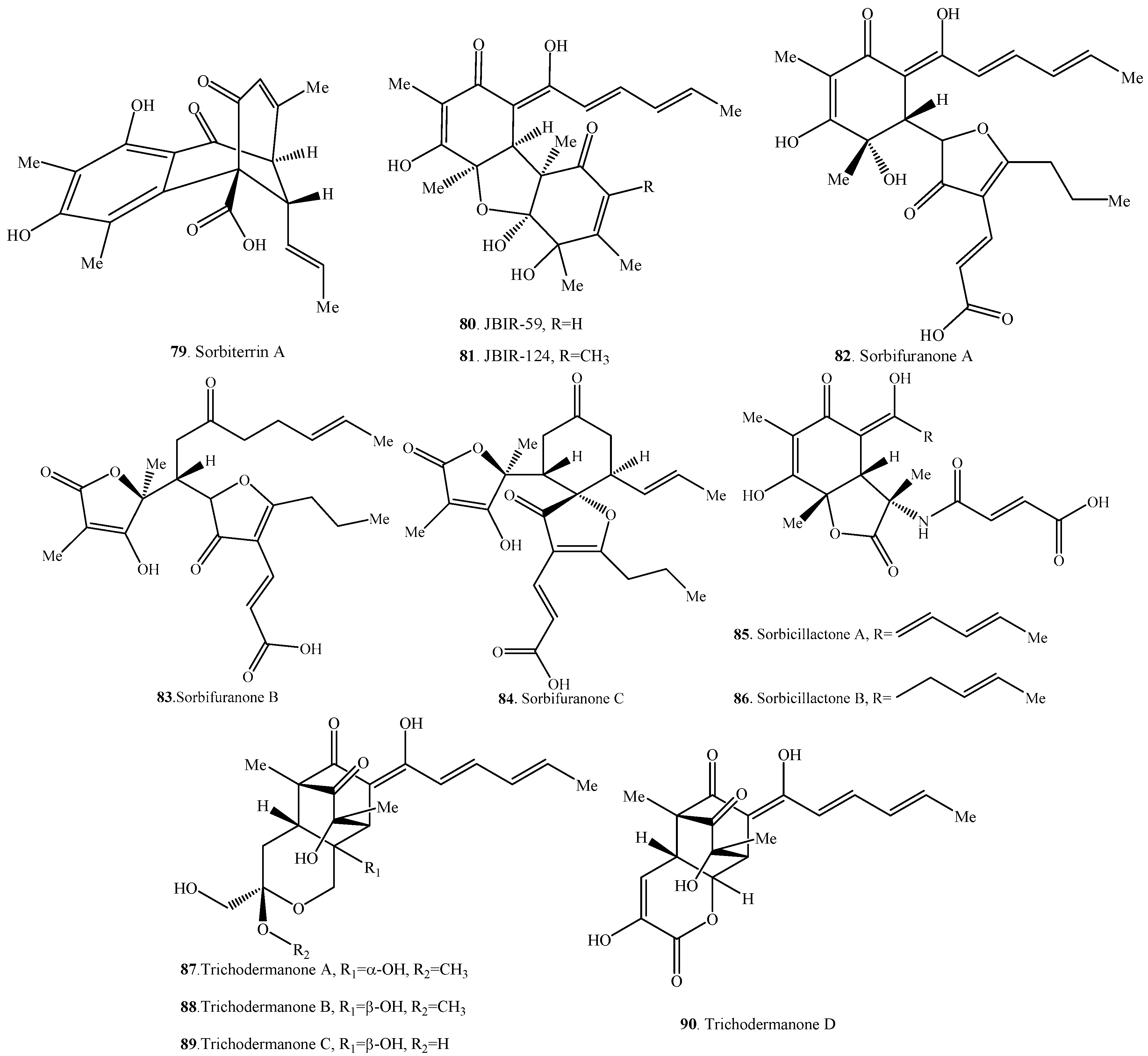
| Sorbiterrin A (79) | Penicillium terrestre from a marine sediment | [61] |
| JBIR-59 (80) | Penicillium citrinum SpI080624G1f01 from the sponge Demospongiae sp. | [43] |
| JBIR-124 (81) | Penicillium citrinum SpI080624G1f01 from the sponge Demospongiae sp. | [62] |
| Sorbifuranone A (82) | Penicillium chrysogenum E03-8/4 from the sponge Tethya aurantium | [35] |
| Sorbifuranone B (83) | Penicillium chrysogenum E03-8/4 from the sponge Tethya aurantium | [35] |
| Sorbifuranone C (84) | Penicillium chrysogenum E03-8/4 from the sponge Tethya aurantium | [35] |
| Sorbicillactone A (85) | Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [21] |
| Penicillium chrysogenum R03-8/4 from the sponge Tethya aurantium | [35] | |
| Sorbicillactone B (86) | Penicillium chrysogenum E01-10/3 from the sponge Ircinia fasciculata | [21] |
| Trichodermanone A (87) | Trichoderma viride from the sponge Agelas dispar | [3] |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Trichodermanone B (88) | Trichoderma viride from the sponge Agelas dispar | [3] |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Trichodermanone C (89) | Trichoderma viride from the sponge Agelas dispar | [3] |
| Trichoderma sp. from the sponge Agelas dispar | [37] | |
| Trichodermanone D (90) | Trichoderma viride from the sponge Agelas dispar | [3] |
| Trichoderma sp. from the sponge Agelas dispar | [37] |
| Sorbicillinoid | Cytotoxic Activity | Ref. |
|---|---|---|
| Sorbicllin (5) | IC50 of 12.7 μM on HL-60 (Leukemia) cell line. | [27] |
| IC50s of 1.6 and 27.2 μM on HeLa and HepG2 cells, respectively. | [7] | |
| IC50s of 6.55 to 28.55 μM on HL-60, U937 and T47D cell lines. | [30] | |
| 6-Demethylsorbicillin (7) | IC50 of 23.9 μM on HL-60 cell line. | [27] |
| 1-(2′-Hydroxy-4′-methoxy-5′-methylphenyl)-E,E-2,4-hexadien-1-one (9) | IC50s of 65.2 and 15.1 µM on MDA-MB-435 and SW-620 cell lines at 72 h, respectively. | [12] |
| 5'-Formyl-2′-hydroxyl-4′-methoxy-(E,E)-sorbophenone (10) | IC50s of 1.5 and 0.5 µM on MDA-MB-435 (melanoma) and SW-620 (colon) cell lines at 72 h, respectively, IC50 of 3.1 µM on OSU-CLL (lymphocytic leukemia) cell line at 48 h. | [12] |
| Scalbucillin B (12) | IC50s of 67.9 and 16.0 µM on MDA-MB-435 and SW-620 cell lines at 72 h, respectively. | [12] |
| 2′,3′-Dihydrosorbicillin (13) | IC50s of 7.4 and 44.4 μM on HeLa and HepG2 cells, respectively. | [7] |
| IC50s of 9.19 to 21.93 μg/mL on various human cancer cell lines. | [4] | |
| Dihydrodemethylsorbicillin (17) | IC50s of 0.1 and 4.8 μM on P388 and K562 cell lines, respectively. | [36] |
| 5′-Formyl-2′-hydroxy-4′-methoxy-(E)-4-hexenophenone (18) | IC50s of 2.3 and 2.5 µM on MDA-MB-435 and SW-620 cell lines at 72 h, respectively. | [12] |
| (2S)-2,3-Dihydro-7-hydroxy-6,8-dimethyl-2-[(E)-prop-1-enyl]-chroman-4-one (21) | IC50 of 9.51 µg/mL on human breast cancer cell line MCF-7. | [4] |
| (2S)-2,3-Dihydro-7-hydroxy-6-methyl-2-[(E)-prop-1-enyl]-chroman-4-one (22) | IC50 of 7.82 µg/mL on human breast cancer cell line MCF-7. | [4] |
| (E)-6-(2,4-Dihydroxyl-5-methylphenyl)-6-oxo-2-hexenoic acid (23) | IC50s of 44.5 μM and 72.8 μM on HL-60 and P388 cell lines, respectively. | [6] |
| 6-(2,4-Dihydroxyl-5-methylphenyl)-6-oxohexanoic acid (24) | IC50s of 81.2 μM and 52.5 μM on HL-60 and P388 cell lines, respectively. | [6] |
| 2-(2′,3′-Dihydrosorbyl)-3,6-dimethyl-5-hydroxy-1,4-benzoquinone (25) | IC50s of 15.7 μM and 5.3 μM on P388 and A549 cell lines, respectively. | [19] |
| Bisvertinolone (34) | IC50 of 5.3 μM on HL-60 cell line. | [27] |
| 16,,17-Dihydrobisvertinolone (35) | IC50s of 1.7 μM and 0.52 μM on P388 and A549 cell lines, respectively. | [19] |
| 10,11-Dihydrobisvertinolone (36) | IC50 of 49 μM on HL-60 cell line. | [27] |
| Tetrahydrobisvertinolone (37) | IC50s of 16.7 μM on A549 cell line. | [19] |
| Trichodimerol = MS-182123 (42) | IC50 of 7.8 μM on HL-60 cell line. | [27] |
| IC50s of 0.33 and 4.7 μM on P388 and A549 cell lines, respectively. | [47] | |
| IC50s of 6.55 to 28.55 μM on HL-60, U937 and T47D cell lines. | [30] | |
| Dihydrotrichodimerol (44) | IC50 of 36.4 μM on HL-60 cell line. | [27] |
| IC50s of 2.8 and 2.1 μM on P388 and A549 cell lines, respectively. | [47] | |
| IC50s of 3-34 μM on U373, A549, SKMEL-28, OE21, Hs683, and B16F10 cell lines. | [51] | |
| Tetrahydrotrichodimerol (45) | IC50s of 8.8 and 4.3 μM on P388 and A549 cell lines, respectively. | [47] |
| Bislongiquinolide =Bisorbibutenolide = Trichotetronine (49) | IC50s of 4-22 μM on U373, A549, SKMEL-28, OE21, Hs683, and B16F10 cell lines. | [51] |
| Oxosorbiquinol (53) | IC50s of 8.9, 29.9, 103.5, 12.7 and 56.3 μM on HL-60, P388, A549, BEL7402 and K562 cell lines, respectively. | [2] |
| Dihydrooxosorbiquinol (54) | IC50s of 10.5, 40.3, 97.6, 31.8 and 68.2 μM on HL-60, P388, A549, BEL7402 and K562 cell lines, respectively. | [2] |
| Dihydrotrichodermolide (56) | IC50s of 11.5 and 22.9 μM on P388 and K562 cell lines, respevtively. | [36] |
| Trisorbicillinone A (61) | IC50s of 3.14, 9.10, 60.28 and 30.21 μM on HL-60, P388, BEL7402 and K562 cell lines, respectively. | [55] |
| Trisorbicillinone B (62) | IC50s of 77.1 and 88.2 μM on P388 and K562 cell lines, respectively. | [56] |
| Trisorbicillinone C (63) | IC50s of 78.3 and 54.3 μM on P388 and K562 cell lines, respectively. | [56] |
| Trisorbicillinone D (64) | IC50s of 65.7 and 51.2 μM on P388 and K562 cell lines, respectively. | [56] |
| Chloctansprirone A (74) | IC50s of 9.2 and 39.7 μM on HL-60 and A549 cell lines, respectively | [58] |
| Chloctansprirone B (75) | IC50 of 37.8 μM on HL-60 cell line. | [58] |
| Sorbicillactone A (85) | IC50 of 2.2 µg/mL on L5178y (murine leukemic lymphoblasts) cell line. | [21] |
| Sorbicillinoid | Antimicrobial activity | Ref. |
|---|---|---|
| Oxosorbicillinol (3) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Sohirnone B (8) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| 5′-Formyl-2′-hydroxyl-4′-methoxy-(E,E)-sorbophenone (10) | Showed potent activity against Aspergillus flavus (NRRL 6541) and moderate activity against Fusarium verticillioides (NRRL 25457). | [33] |
| Scalbucillin B (12) | MIC value of 0.60 μg/mL (2.42 μM) against Aspergillus niger. | [12] |
| 2′,3′-Dihydrosorbicillinol (13) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Sohirnone A (14) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| 1-(2′-Hydroxy-4′-methoxy-5′-hydroxymethylphenyl)-E-4-hexen-1-one (16) | Showed potent activity against Aspergillus flavus (NRRL 6541) and weak activity against Fusarium verticillioides (NRRL 25457). | [33] |
| 5′-Formyl-2′-hydroxy-4′-methoxy-(E)-4-hexenophenone (18) | Strong antifungal activity on Aspergillus niger with MIC values of 0.04 μg/mL (0.16 μM). | [12] |
| Sorrentanone [=3-hydroxy-2,5-dimethyl-6-(1′-oxo-2′,4′-dienylhexyl)-1,4-benzoquione, 26] | MIC values of 32, 16, 128, 32, 32 and 64 µg/mL on Staphylococcus pneumoniae A9585, S. pyogenes A9604, Enterococcus faecalis A20688, S. aureus/Hetero MR A27218, S. epidermidis A24548, and S. haemolytic A21638, respectively. | [18] |
| Dihydrotrichodimerol (44) | Strong antibacterial activity on Bacillus megaterium with MIC value of 25 μg/mL. | [13] |
| Tetrahydrotrichodimerol (45) | Strong antibacterial activity on Bacillus megaterium with MIC value of 12.5 μg/mL. | [13] |
| Bisvertinoquinol (47) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Bisorbicillinol (48) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Dihydrotrichodimer ether A (59) | Strong antibacterial activity on Escherichia coli with MIC value of 25 μg/mL. | [13] |
| Dihydrotrichodimer ether B (60) | Strong antibacterial activity on Escherichia coli and Ballus subtilis with MIC values of 50 μg/mL. | [13] |
| Rezishanones A (66) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Rezishanone B (67) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Rezishanone C = Sorbivinetone (68) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Rezishanone D (69) | Weak antibacterial activity on Staphylococcus aureus and Bacillus subtilis. | [5] |
| Strong antifungal activity on Aspergillus niger with MIC value of 0.05 μg/mL (0.20 μM) | [12] |
| Sorbicillinoid | Biological Activity | Ref. |
|---|---|---|
| 6′-Hydroxyoxosorbicillinol (4) | Inhibitory activity on soybean lipoxygenase; Prostaglandin D2 and leucotriene B4 release suppression activity. | [22] |
| Bisvertinolone (34) | Inhibitory effect on β-l,6-glucan biosynthesis | [42] |
| Isobisvertinol (38) | Inhibitory effect on lipid droplet accumulation in mouse macrophages | [40] |
| Trichodimerol (42) | Inhibitory effect on bacterial endotoxin-induced production of tumor necrosis factor (TNF-α) in murine macrophages and human peripheral blood monocytes | [46] |
| Inhibitory effect on lipopolysaccharide-induced eicosanoid secretion in THP-1 human monocytic cells | [66] | |
| Suppression of the production of tumor necrosis factor-α and nitric oxide in LPS-stimulate RAW264.7 cells | [50] | |
| Dihydrotrichodimerol (44) | Activation of peroxisome proliferator-activated recptor γ (PPAR γ) with an ED50 of 80 ng/mL | [50] |
| Suppression of the production of tumor necrosis factor-α and nitric oxide in LPS-stimulate RAW264.7 cells | [50] | |
| Effect on feeding perference of the aphid | [48] | |
| Bislongiquinolide (49) | Effect on feeding perference of the aphid | [48] |
| Sorbiterrin A (79) | Inhibitory effect on acetylcholinesterase activity with IC50 value of 25 μg/mL | [61] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, J.; Wang, X.; Xu, D.; Fu, X.; Zhang, X.; Lai, D.; Zhou, L.; Zhang, G. Sorbicillinoids from Fungi and Their Bioactivities. Molecules 2016, 21, 715. https://doi.org/10.3390/molecules21060715
Meng J, Wang X, Xu D, Fu X, Zhang X, Lai D, Zhou L, Zhang G. Sorbicillinoids from Fungi and Their Bioactivities. Molecules. 2016; 21(6):715. https://doi.org/10.3390/molecules21060715
Chicago/Turabian StyleMeng, Jiajia, Xiaohan Wang, Dan Xu, Xiaoxiang Fu, Xuping Zhang, Daowan Lai, Ligang Zhou, and Guozhen Zhang. 2016. "Sorbicillinoids from Fungi and Their Bioactivities" Molecules 21, no. 6: 715. https://doi.org/10.3390/molecules21060715
APA StyleMeng, J., Wang, X., Xu, D., Fu, X., Zhang, X., Lai, D., Zhou, L., & Zhang, G. (2016). Sorbicillinoids from Fungi and Their Bioactivities. Molecules, 21(6), 715. https://doi.org/10.3390/molecules21060715