
Ellagitannins from Raspberry (Rubus idaeus L.) Fruit as Natural Inhibitors of Geotrichum candidum


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
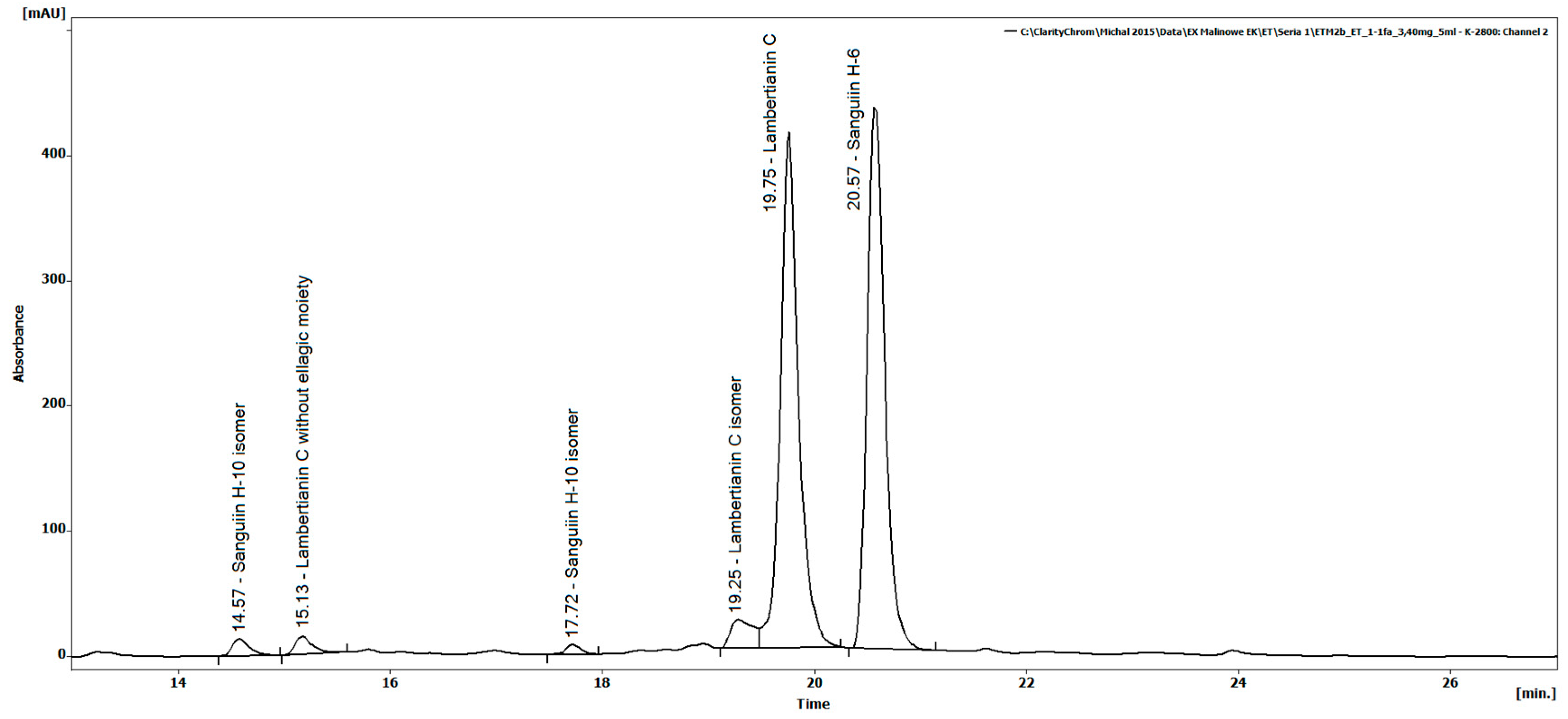
2.1. Ellagitannin Identification in REP by LC-MS
2.2. Quantitative Composition of Phenolic Compounds in REP
2.3. Inhibition of Geotrichum Candidum Growth by Ellagitannins
2.4. The Fungistatic Activity (FA) of Ellagitannins and the Linear Growth Rate (T) of Filamentous Fungi in Their Presence
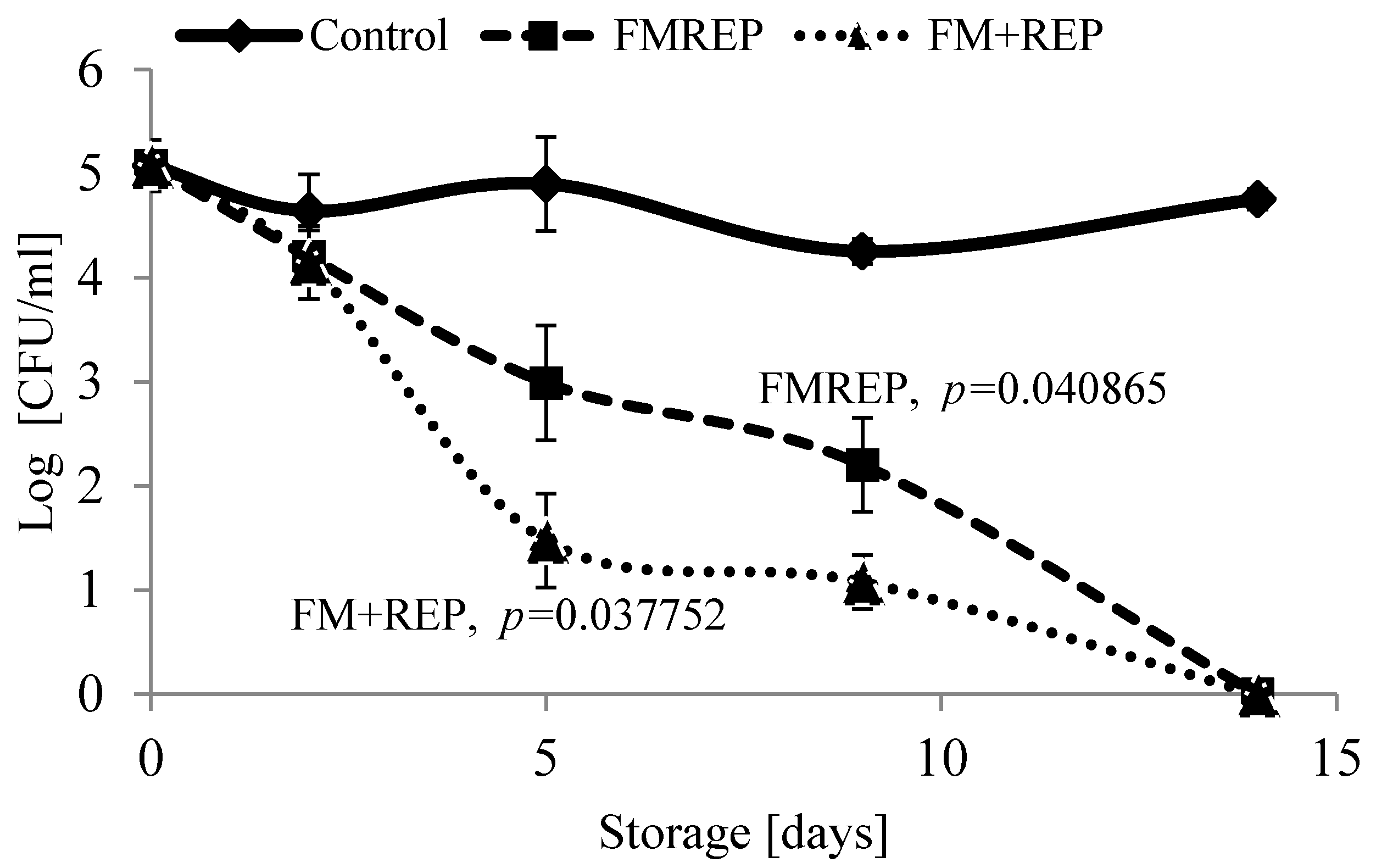
2.5. In Situ Effect of Ellagitannin Preparation
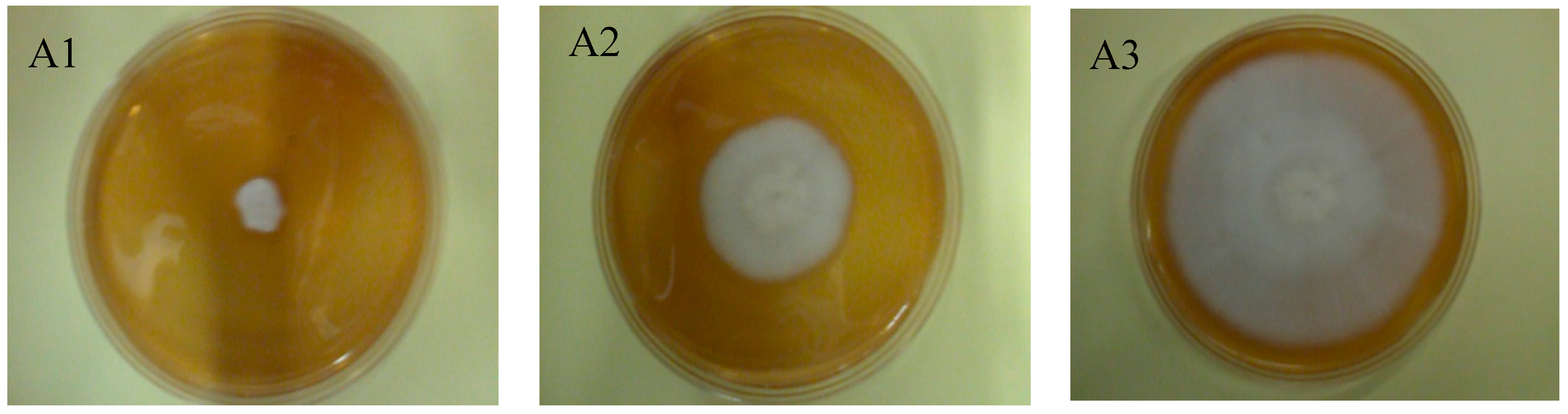
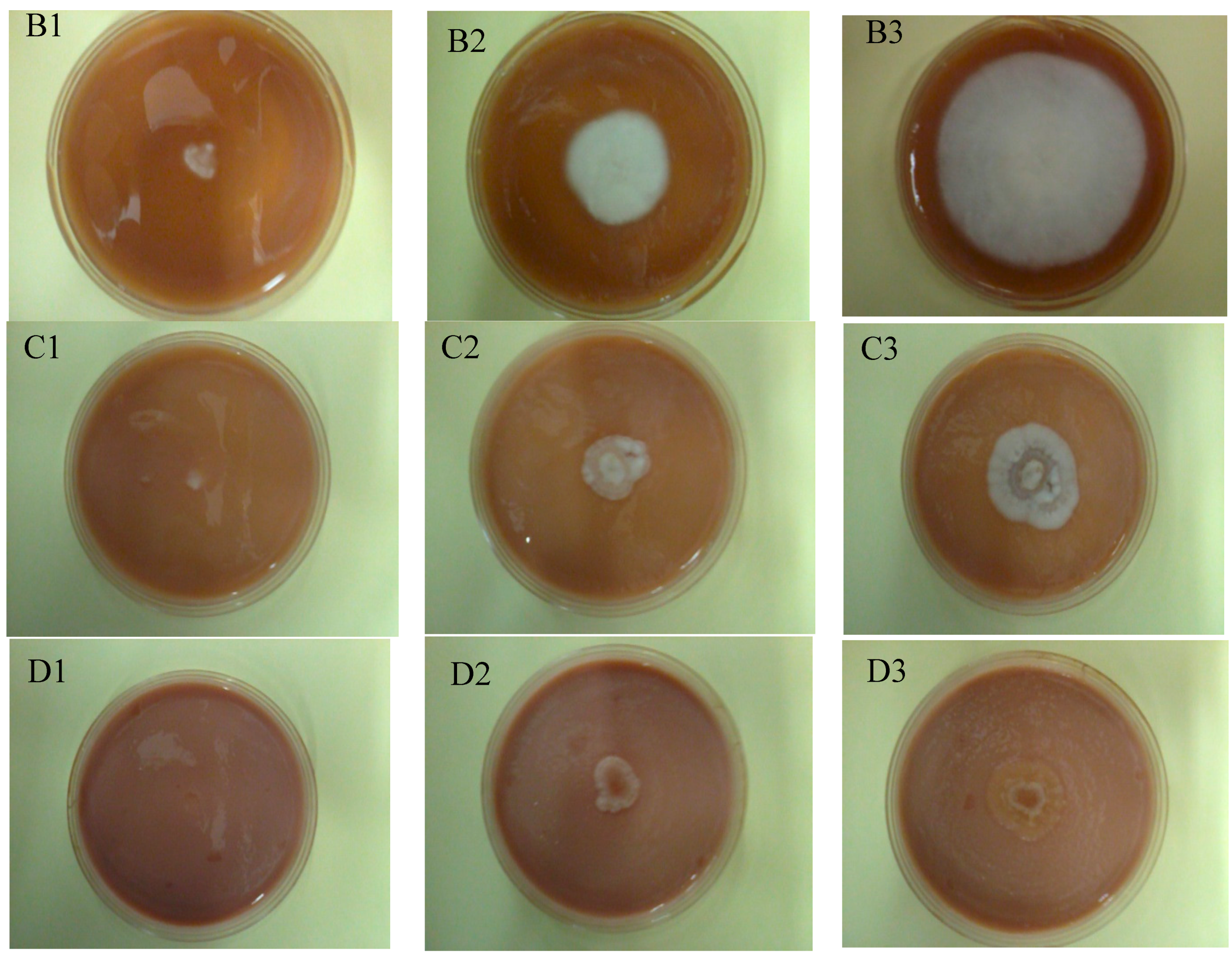
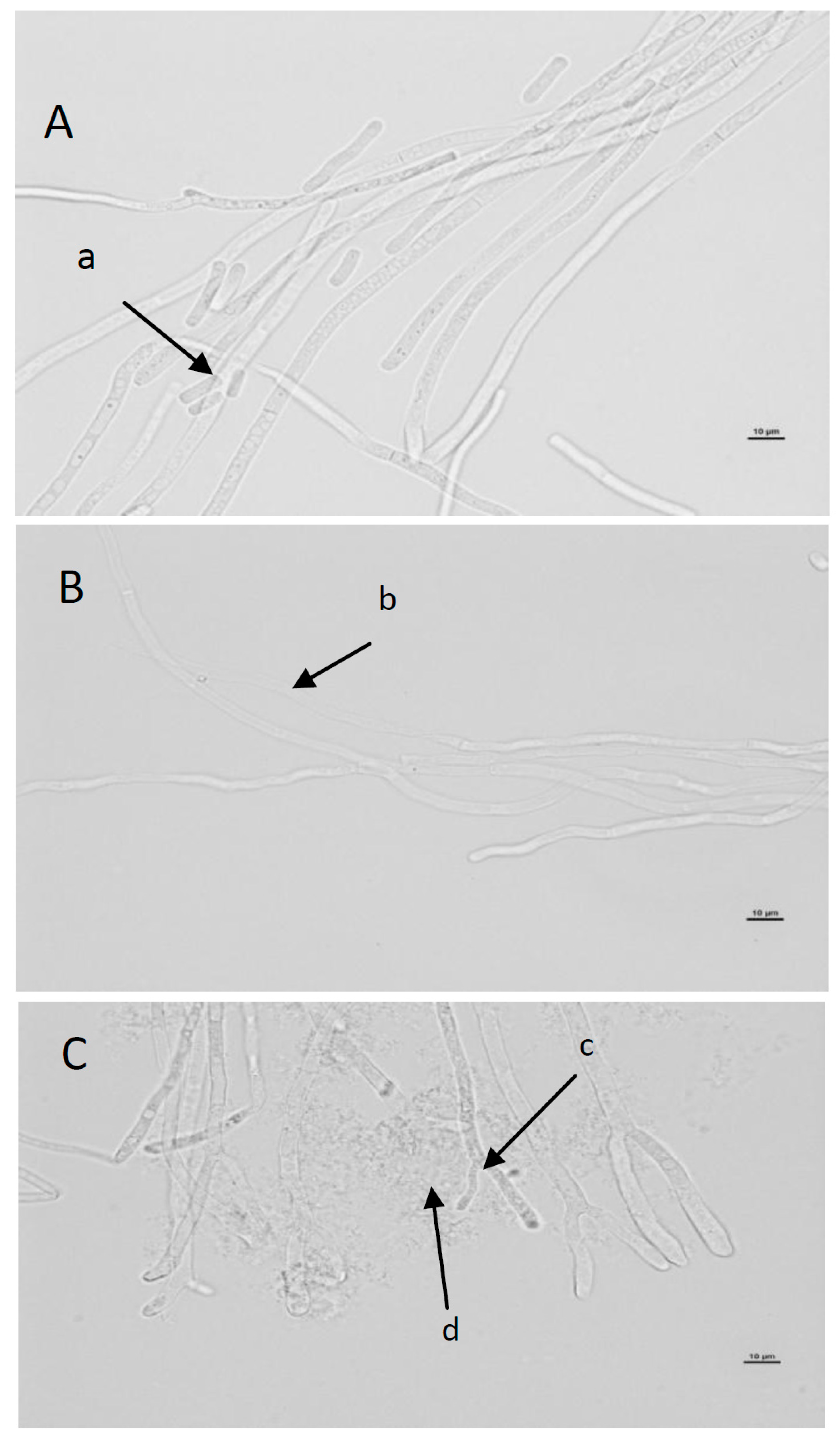
2.6. Evaluation of Macroscopic and Microscopic Morphology of Geotrichum Candidum ŁOCK 0511
3. Materials and Methods
3.1. Plant Material
3.2. Fruit Processing and Ellagitannin Extraction
3.3. Isolation of Sanguiin H-6 and Lambertianin C
3.4. Identification of Ellagitannins
3.5. Quantification of Ellagitannins
3.6. HPLC Analysis of Anthocyanins
3.7. HPLC Analysis of Flavanols (Sum of proAnthocyanidins and Catechins)
3.8. Microorganisms
3.9. Antagonistic Activity of Ellagitannins
3.10. Inhibition of Geotrichum Candidum Growth by Ellagitannins
3.11. Fungistatic Activity and Linear Growth Rate of Filamentous Fungi in the Presence of Ellagitannins
3.12. In Situ Effect of the Raspberry Ellagitannin Preparation
3.13. Microscopic Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yoshida, T.; Amakura, Y.; Yoshimura, M. Structural features and biological properties of ellagitannins in some plant families of the order Myrtales. Int. J. Mol. Sci. 2010, 11, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Ellagitannins, ellagic acid and their derivative metabolites: A review about source, metabolism, function and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Vrhovsek, U.; Guella, G.; Mattivi, F. Profiling and accurate quantification of Rubus ellagitannins and ellagic acid conjugates using direct UPLC-Q-TOF HDMS and HPLC-DAD analysis. J. Agric. Food Chem. 2010, 58, 4602–4616. [Google Scholar] [CrossRef] [PubMed]
- Ascacio-Valdés, J.; Burboa, E.; Aguilera-Carbo, A.F.; Aparicio, M.; Perez-Schmidt, R.; Rodrígez, R.; Aguilar, C.N. Antifungal ellagitannin isolated from Euphorbia antisyphilitia Zucc. Asian Pac. J. Trop. Biomed. 2013, 3, 41–46. [Google Scholar] [CrossRef]
- Türkyılmaz, M.; Tağı, S.; Dereli, U.; Özkan, M. Effects of various pressing programs and yields on the antioxidant activity, antimicrobial activity, phenolic content and colour of pomegranate juices. Food Chem. 2013, 138, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Al-Zoreky, N.S. Antimicrobial activity of pomegranate (Punica granatum L.) fruit peels. Int. J. Food Microbiol. 2009, 134, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Navarro, V.; Villarreal, M.L.; Rojas, G.; Lozoya, X. Antimicrobial evaluation of some plants used in Mexican traditional medicine for the treatment of infectious diseases. J. Ethnopharmacol. 1996, 53, 143–147. [Google Scholar] [CrossRef]
- O’Brien, M.; O’Keiely, P.; Fuller, H.T. Fungi isolated from contaminated baled grass silage on farms in the Irish Midlands. FEMS Microbiol. Lett. 2005, 247, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Skóra, M.; Witalis, J.; Krzysciak, P.; Macura, A.B. Fungal genus Geotrichum: An opportunistic pathogen of humans. Post. Mikrobiol. 2009, 48, 125–132. [Google Scholar]
- Pottier, I.; Gente, S.; Vernoux, J.P.; Guéguen, M. Safety assessment of dairy microorganisms: Geotrichum candidum. Int. J. Food Microbiol. 2008, 126, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.; Kylli, P.; Ollilainen, V.; Salminen, J.-P.; Heinonen, M. Antioxidant Activity of Isolated Ellagitannins from Red Raspberriesand Cloudberries. J. Agric. Food Chem. 2012, 60, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Hager, T.J.; Howard, L.R.; Liyanage, R.; Lay, J.O.; Prior, R.L. Ellagitannin composition of blackberry by HPLC-ESI-MS and MALDI-TOF-MS. J. Agric. Food Chem. 2008, 56, 661–669. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.; Martinussen, I.; Stewart, D. Towards fruitful metabolomics: High throughput analyses of polyphenol composition in berries using direct infusion mass spectrometry. J. Chromatogr. B 2008, 871, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Mertz, C.; Cheynier, V.; Günata, Z.; Brat, P. Analysis of phenolics compounds in two blackberry species (Rubus glaucus and Rubus adenotrichus) by high-performance liquid chromatography with diode array detection and electrospray ion trap mass spectrometry. J. Agric. Food Chem. 2007, 55, 8616–8624. [Google Scholar] [CrossRef] [PubMed]
- Foss, R.S.; Nakamura, C.V.; Udea-Nakamura, T.; Cortez, D.A.G.; Endo, E.H.; dias Filho, B.P. Antifungal activity of pomegranate peel extract and isolated compound punicalagin against dermatophytes. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Mehrabian, S.; Majd, A.; Majd, I. Antimicrobial effect of three plants (Rubia tinctorum, Carthamus tinctorius and Juglans regia) on some airborne microorganisms. Aerobiologia 2000, 16, 455–458. [Google Scholar] [CrossRef]
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; ait ben Aoumar, A. Antifungal activity of some Moroccan plants against Geotrichum candidum the casual agent of postharvest citrus sour rot. Crop Protect. 2012, 35, 41–46. [Google Scholar] [CrossRef]
- Webster, D.; Taschereau, P.; Belland, R.J.; Sand, C.; Rennie, R.P. Antifungal activity of medicinal plant extracts; preliminary screening studies. J. Ethnopharmacol. 2008, 115, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Thirach, S.; Tragoolpua, K.; Punjaisee, S.; Khamwan, C.; Jatisatienr, C.; Kunyanone, N. Antifungal activity of some medicinal plant extracts against Candida albicans and Cryptococcus neoformans. Acta Hortic. 2003, 597, 217–221. [Google Scholar] [CrossRef]
- Daglia, M. Polyphones as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.-M. Antimicrobial properties of phenolic compound from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Bialonska, D.; Kasimsetty, S.G.; Schrader, K.K.; Ferreira, D. The effect of pomegranate (Punica granatum L.) byproducts and ellagitannins on the growth of human gut bacteria. J. Agric. Food Chem. 2014, 57, 8344–8349. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hewes, D.; Salaheen, S.; Federman, C. Effects if blackberry juice on growth inhibition of foodborne pathogens and growth promotion of Lactobacillus. Food Control. 2014, 37, 15–20. [Google Scholar] [CrossRef]
- Boutrou, R.; Guéguen, M. Interest in Geotrichum candidum for cheese technology. Int. J. Food Microbiol. 2005, 102, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Osawa, R.; Kuroiso, K.; Goto, S.; Shimizu, A. Isolation of tannin-depending lactobacilli from humans and fermented food. Appl. Environ. Microbiol. 2000, 66, 3093–3097. [Google Scholar] [CrossRef] [PubMed]
- Li, M.S.; Yao, K.; He, Q.; Jia, D.Y. Biodegradation of gallotannins and ellagitannins. J. Basic. Microbiol. 2006, 1, 68–84. [Google Scholar]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; di Venere, D. Activity of extracts from wild edible herbs against postharvest fungal diseases of fruit and vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]
- Yuvamoto, P.D.; Said, S. Germination, duplication cycle and septum formationare altered by caffeine, caffeic acid and cinnamic acid in Aspergillus nidulans. Microbiology 2007, 76, 735–738. [Google Scholar] [CrossRef]
- Kanaani, J.; Ginsburg, H. Effects of cinnamic acid derivatives on in vitro growth of Plasmodium falciparum and on the permeability of the membrane of malariainfected erythrocites. Antimicrob. Agents Chemother. 1992, 36, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Sójka, M.; Kołodziejczyk, K.; Milala, J.; Abadias, M.; Viñas, I.; Guyot, S.; Baron, A. Composition and properties of the polyphenolic extracts obtained from industrial plum pomaces. J. Funct. Foods 2015, 12, 168–178. [Google Scholar] [CrossRef]
- Sójka, M.; Klimczak, E.; Macierzyński, J.; Kołodziejczyk, K. Nutrient and polyphenolic composition of industrial strawberry press cake. Eur. Food Res. Technol. 2013, 237, 995–1007. [Google Scholar] [CrossRef]
- Yamaguchi, M.U.; Garcia, F.P.; Cortez, D.A.G; Ueda-Nakamura, T.; Filho, B.P.D.; Nakamura, C.V. Antifungal effects of ellagitannin isolated from leaves of Ocotea odorifera (Lauraceae). Atonie Van Leuw. 2011, 99, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Gleń, K.; Boligłowa, E. The effect of PRP SOL fertilizer on the dynamics of phytopathogenic and antagonistic fungi growth in vito. J. Res. Appl. Agric. Eng. 2011, 56, 97–101. [Google Scholar]
- Sample Availability: Samples of the REP preparation are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min) | MS Data | MS/MS Value | MS/MS Data | Tentative Structural Assignment |
|---|---|---|---|---|
| 14.57 | [1567.15]−1 [783.07]−2 | 783 | [1235.07]−1 [935.08]−1 [633.07]−1 [469.01]−1 [301.00]−1 | Sanguiin H-10 isomer |
| 15.13 | [1250.60]−2 [833.40]−3 | 1250 | [2200.19]−1 [1867.14]−1 [1567.14]−1 [1235.07]−1 [933.06]−2 [633.07]−1 [301.00]−1 | Lambertianin C without ellagic moiety |
| 17.72 | [1567.15]−1 [783.07]−2 | 783 | [1265.14]−1 [1103.09]−1 [935.08]−1 [933.07]−1 [633.07]−1 [469.01]−1 [301.00]−1 | Sanguiin H-10 isomer |
| 19.25 | [1401.01]−2 [934.07]−3 | 1401 | [1869.14]−1 [1567.14]−1 [1235.07]−1 [935.08]−1 [633.07]−1 [301.00]−1 | Lambertianin C isomer |
| 19.75 | [1401.01]−2 [934.07]−3 | 1401 | [1869.14]−1 [1567.14]−1 [1235.07]−1 [935.08]−1 [633.07]−1 [301.00]−1 | Lambertianin C (standard) |
| 20.57 | [934.07]−2 [1869.14]−1 | 934 | [1567.14]−1 [1235.07]−1 [935.08]−1 [633.07]−1 [301.00]−1 | Sanguiin H-6 (standard) |
| Compound | Mean (mg/100 g) | SD |
|---|---|---|
| Ellagitannins | ||
| Sanguiin H-10 isomer 1 | 1065 | 57 |
| Lambertianin C without ellagic moiety | 1456 | 97 |
| Sanguiin H-10 isomer 2 | 731 | 47 |
| Lambertianin C isomer | 2961 | 332 |
| Lambertianin C | 44156 | 1705 |
| Sanguiin H-6 | 35917 | 1300 |
| Total ellagitannins | 86287 | 3537 |
| Other Phenolics | ||
| Total anthocyanins | 823 | 6 |
| Total flavanols (mDP) | 7003 (1.9) | 158 (0.1) |
| Test | REP | LC | SH6 |
|---|---|---|---|
| MIC (mg/mL) | 10.0 | 10.0 | 10.0 |
| MFC (mg/mL) | 30.0 | 30.0 | 30.0 |
| Zone of growth inhibition (mm) ± SD | 10.3 ± 0.47 a | 9.3± 0.47 a | 8.3± 1.69 a |
| Ellagitannins (mg/mL) | FA (%) | Growth Rate TEllagitannins (mm/day) | Growth Rate TControl (mm/day) | |||
|---|---|---|---|---|---|---|
| Time (days) | ||||||
| 1 | 2 | 3 | 6 | |||
| 1.0 | 24.7 | 27.5 | 16.7 | 13.8 | 51.9 * | 61.5 |
| 5.0 | 54.5 | 49.4 | 50.3 | 51.4 | 33.9 ** | 56.0 |
| 10.0 | 66.4 | 60.1 | 74.4 | 65.2 | 16.2 *** | 56.0 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klewicka, E.; Sójka, M.; Klewicki, R.; Kołodziejczyk, K.; Lipińska, L.; Nowak, A. Ellagitannins from Raspberry (Rubus idaeus L.) Fruit as Natural Inhibitors of Geotrichum candidum. Molecules 2016, 21, 908. https://doi.org/10.3390/molecules21070908
Klewicka E, Sójka M, Klewicki R, Kołodziejczyk K, Lipińska L, Nowak A. Ellagitannins from Raspberry (Rubus idaeus L.) Fruit as Natural Inhibitors of Geotrichum candidum. Molecules. 2016; 21(7):908. https://doi.org/10.3390/molecules21070908
Chicago/Turabian StyleKlewicka, Elżbieta, Michał Sójka, Robert Klewicki, Krzysztof Kołodziejczyk, Lidia Lipińska, and Adriana Nowak. 2016. "Ellagitannins from Raspberry (Rubus idaeus L.) Fruit as Natural Inhibitors of Geotrichum candidum" Molecules 21, no. 7: 908. https://doi.org/10.3390/molecules21070908
APA StyleKlewicka, E., Sójka, M., Klewicki, R., Kołodziejczyk, K., Lipińska, L., & Nowak, A. (2016). Ellagitannins from Raspberry (Rubus idaeus L.) Fruit as Natural Inhibitors of Geotrichum candidum. Molecules, 21(7), 908. https://doi.org/10.3390/molecules21070908