
Siegesbeckia orientalis Extract Inhibits TGFβ1-Induced Migration and Invasion of Endometrial Cancer Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
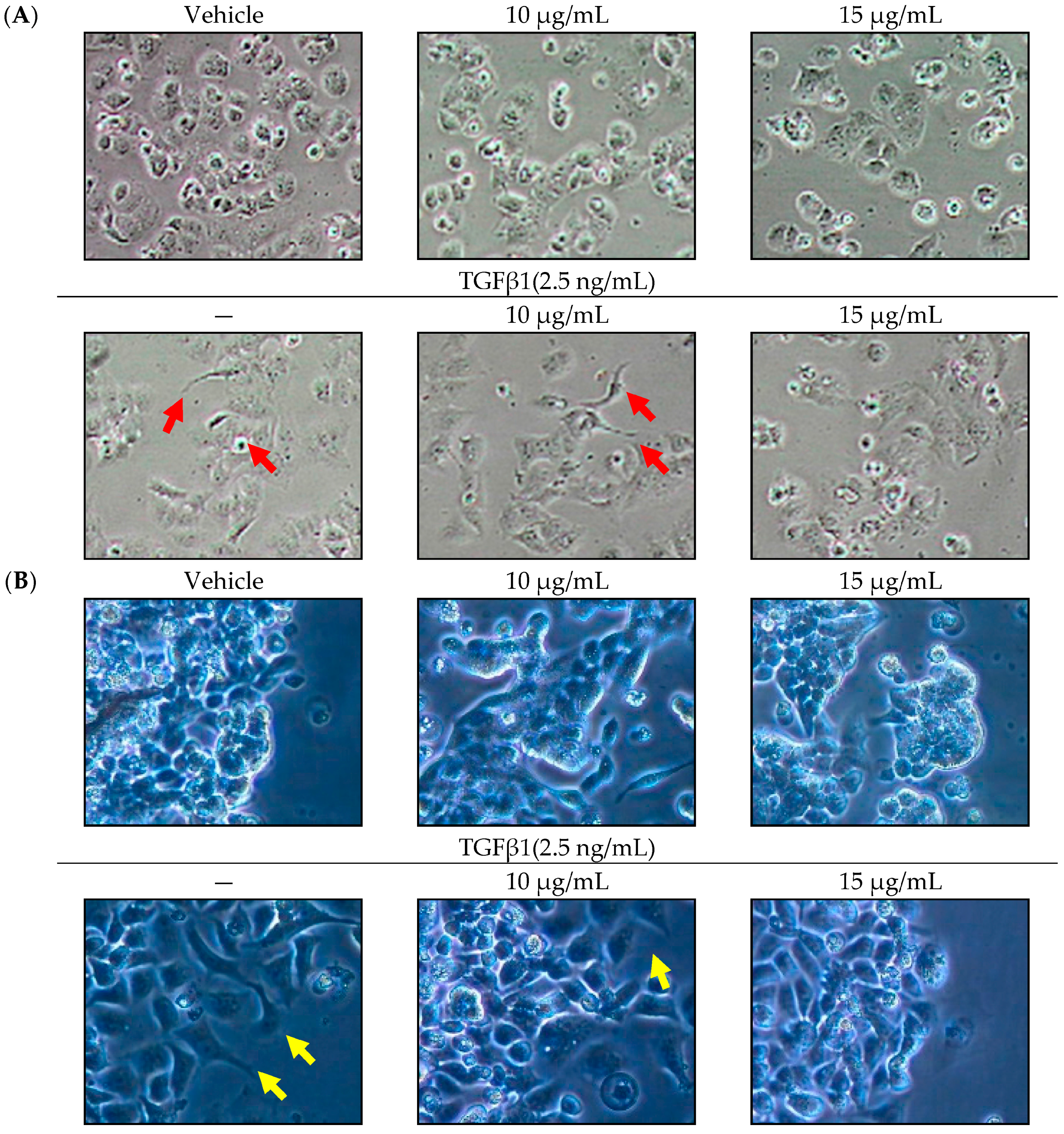
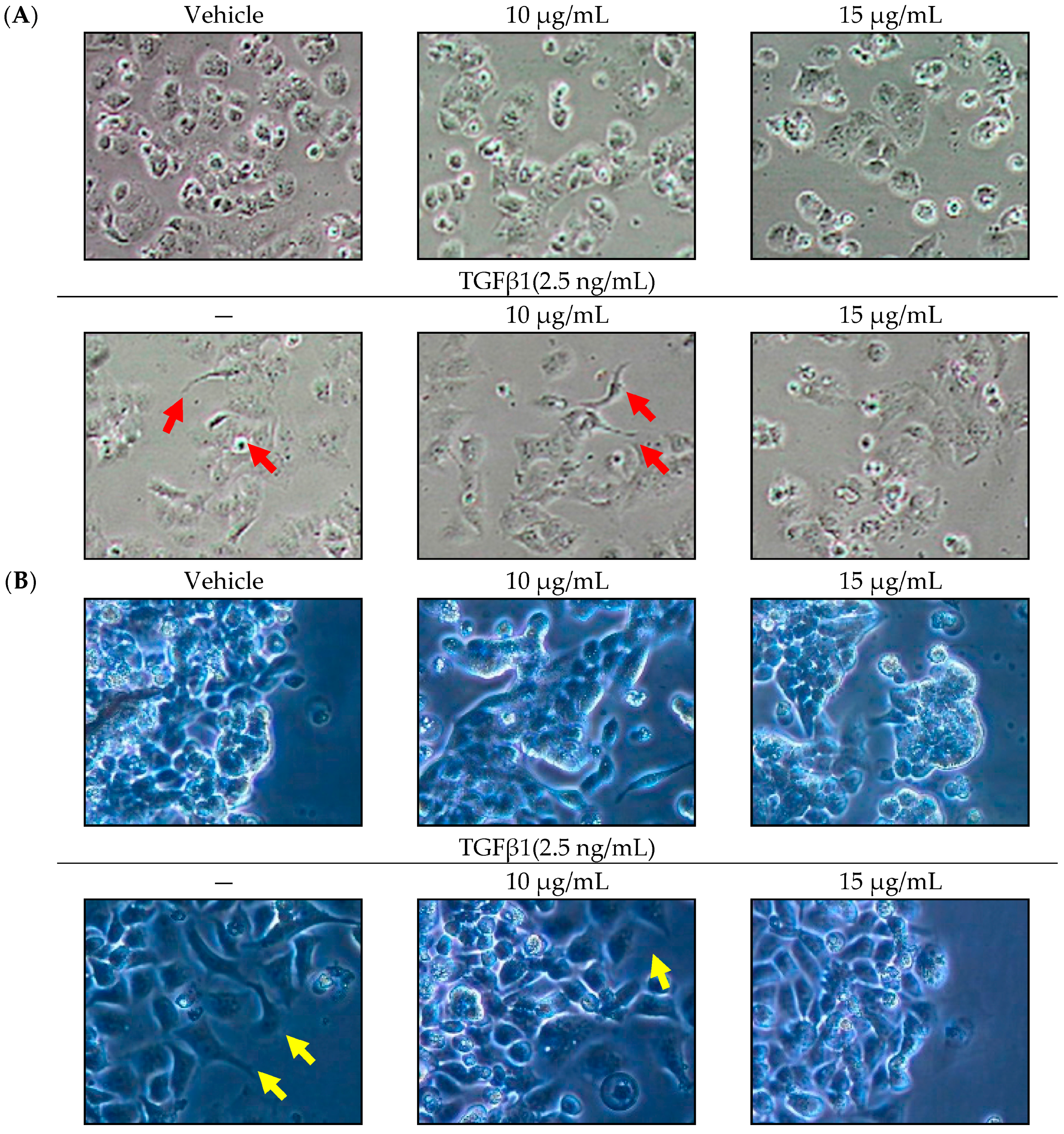
2.1. Effect of SOE on Morphology of Endometrial Cancer Cells
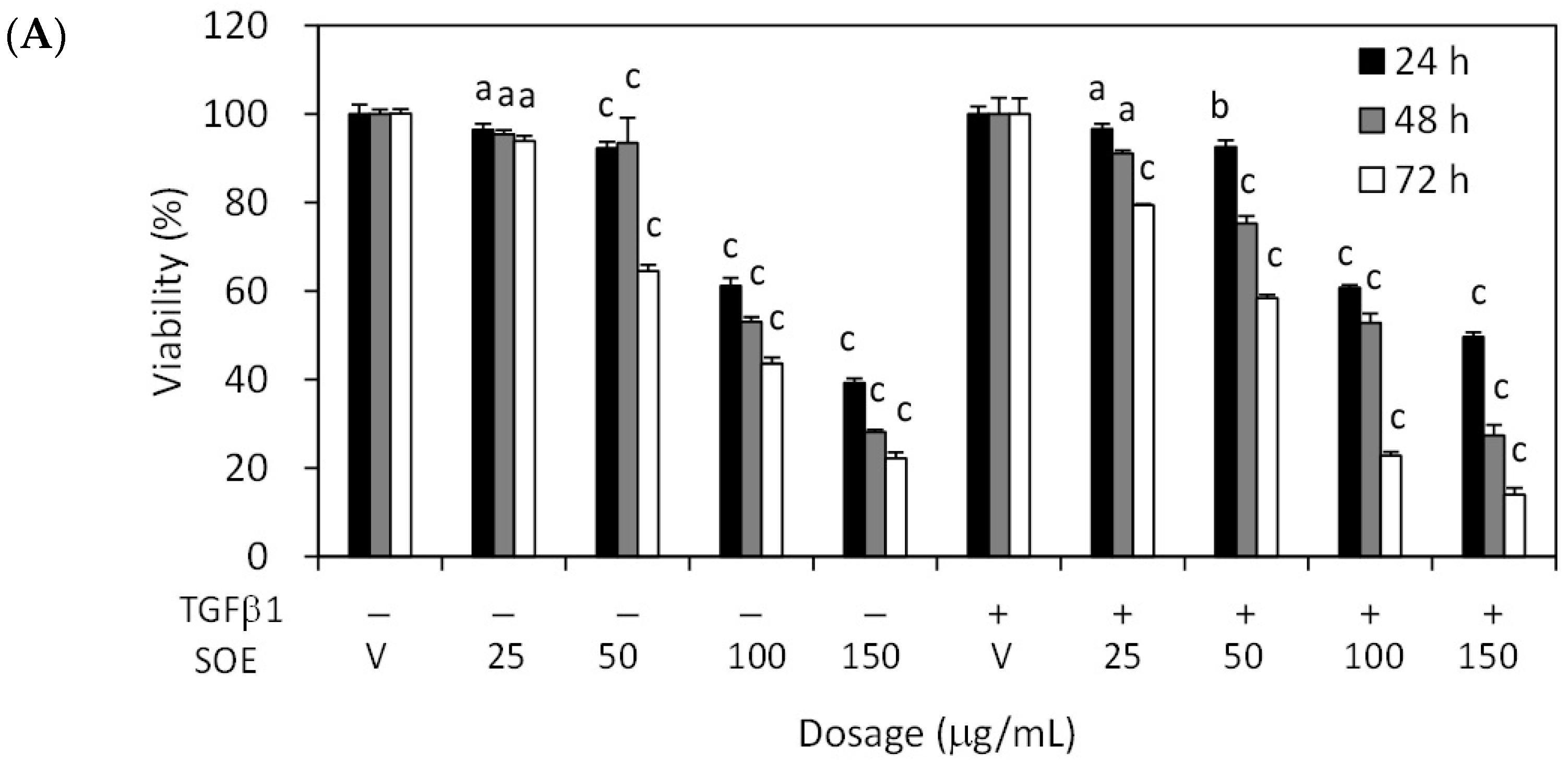
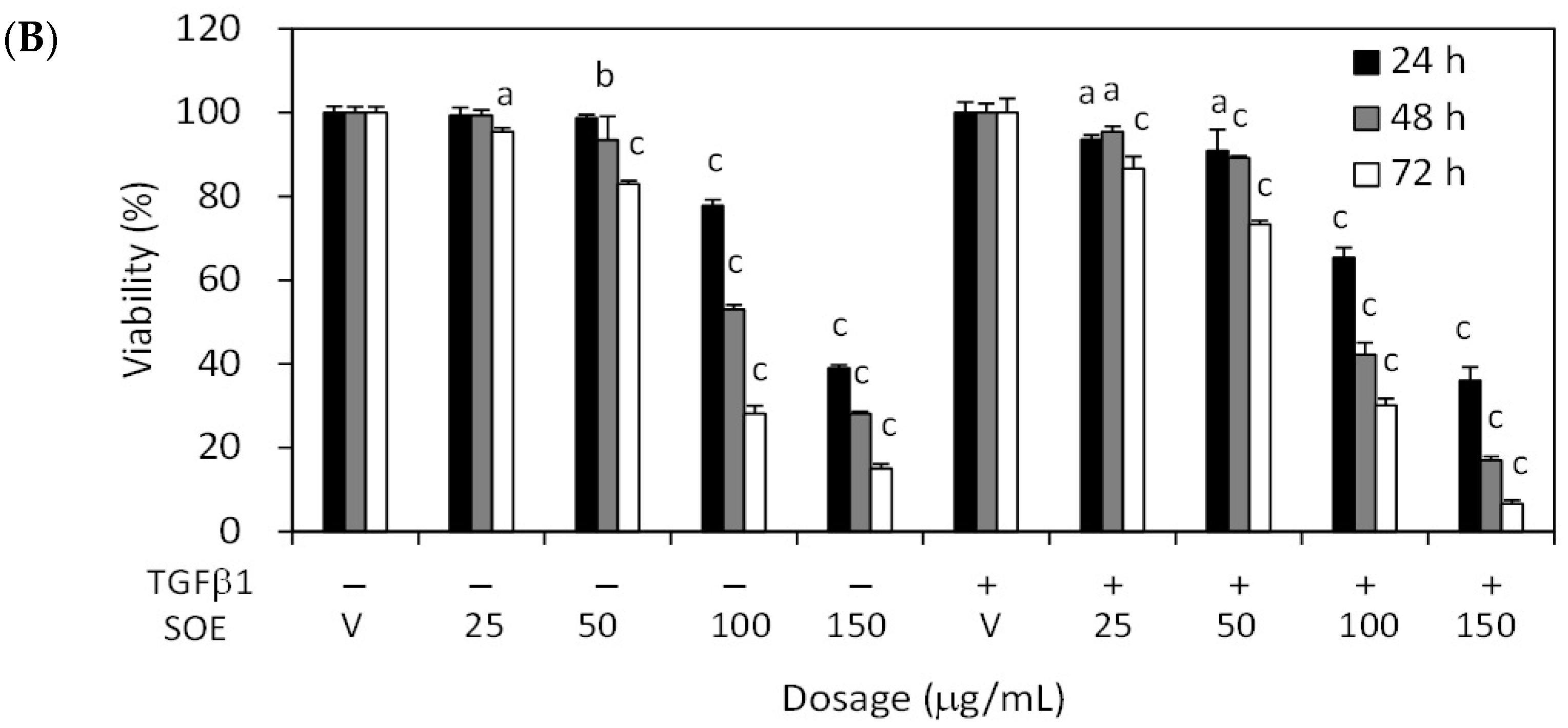
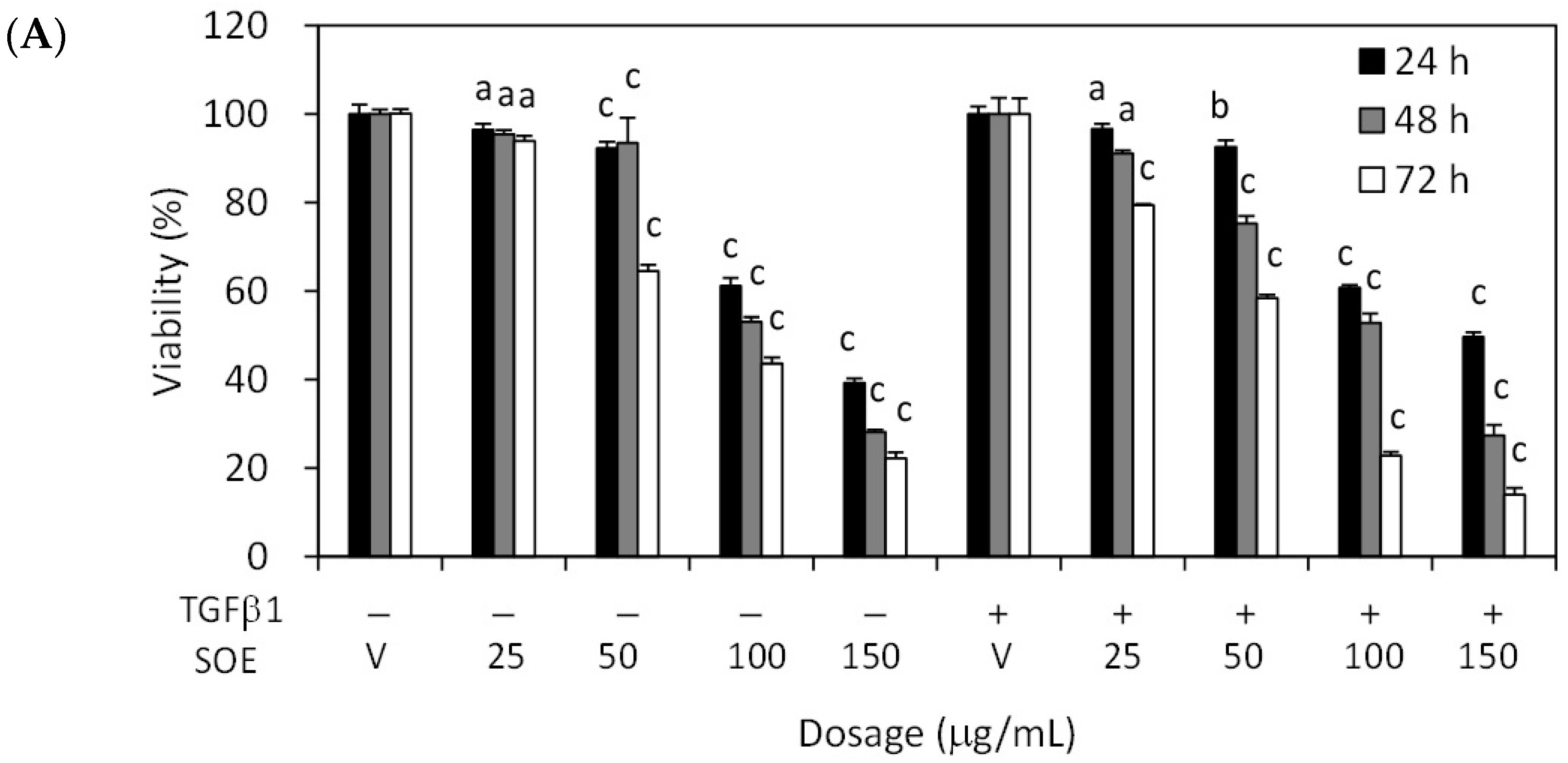
2.2. Inhibitory Effect of SOE on Proliferation of Endometrial Cancer Cells
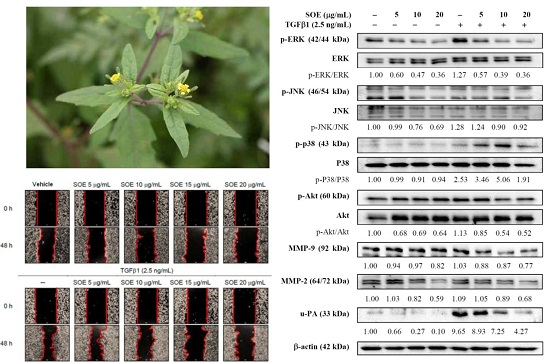
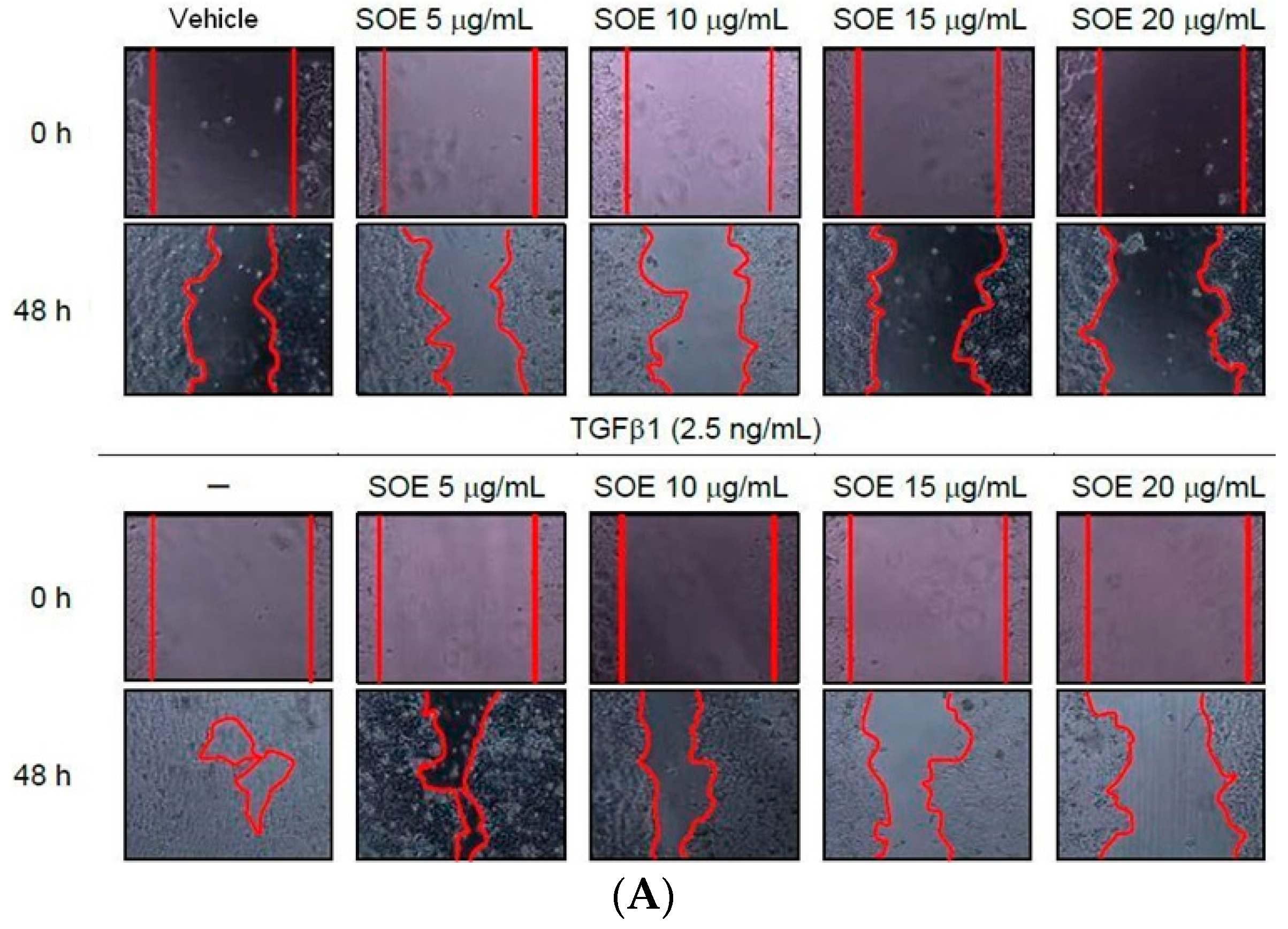
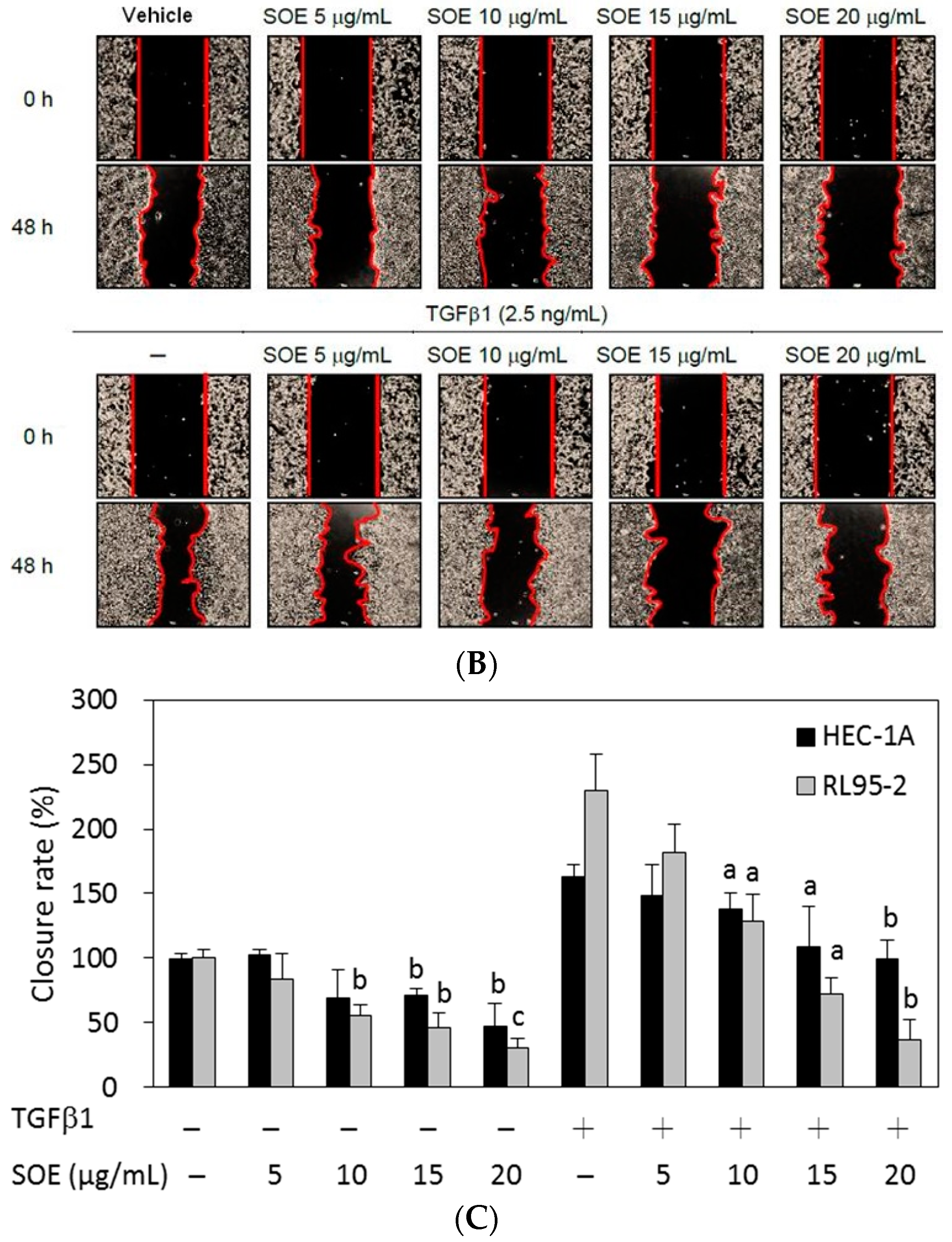
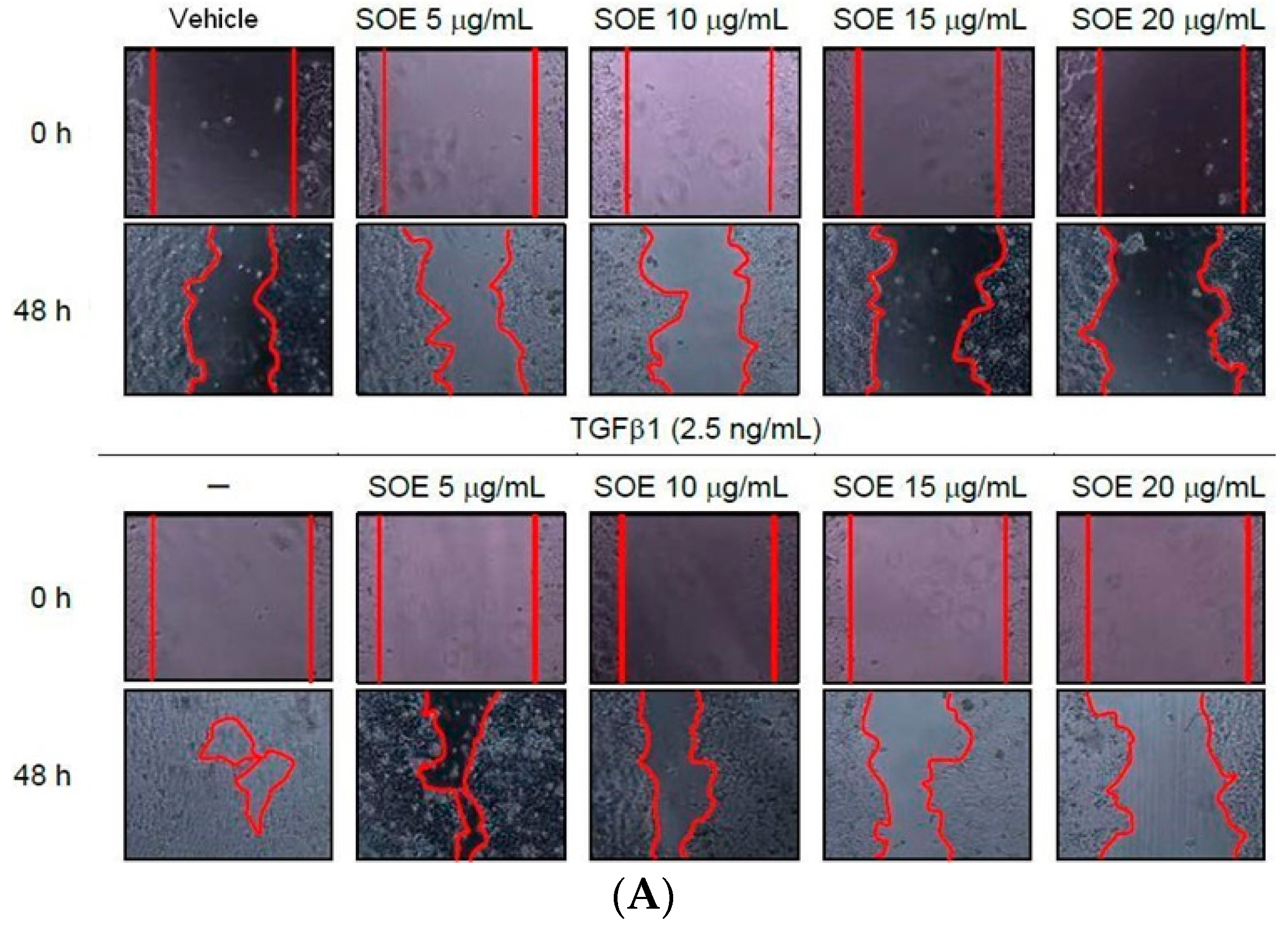
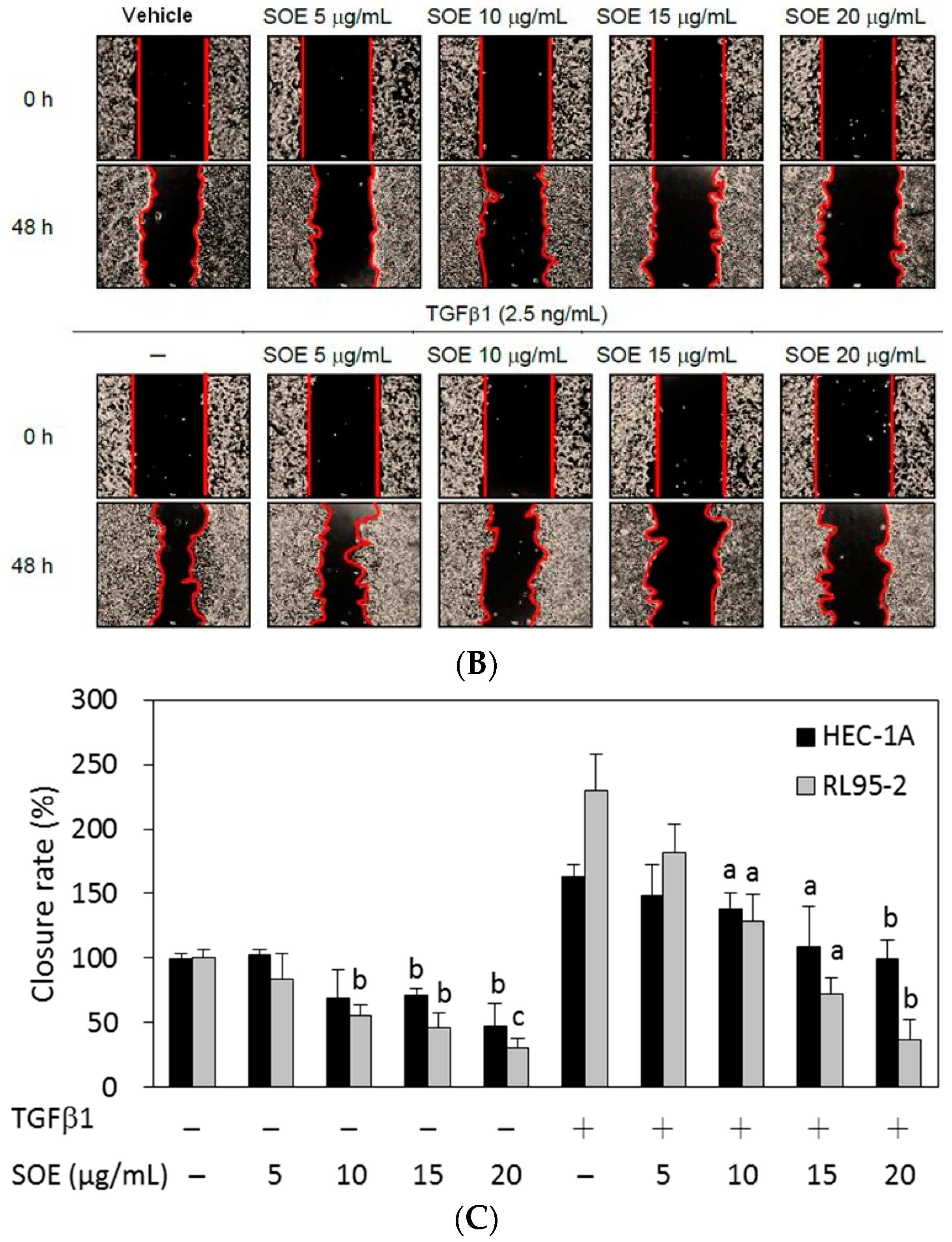
2.3. Inhibitory Effect of SOE on Migration of Endometrial Cancer Cells
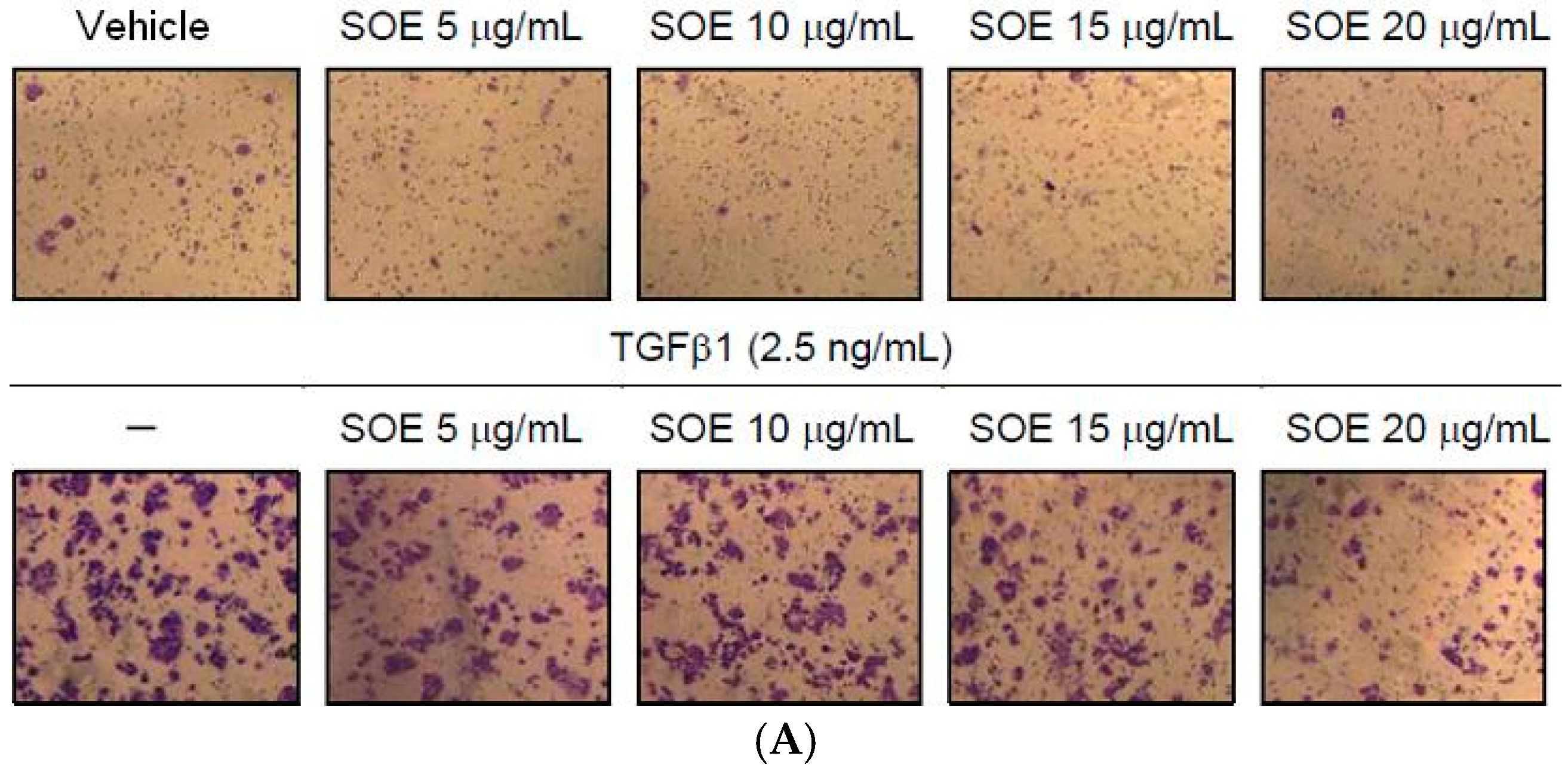
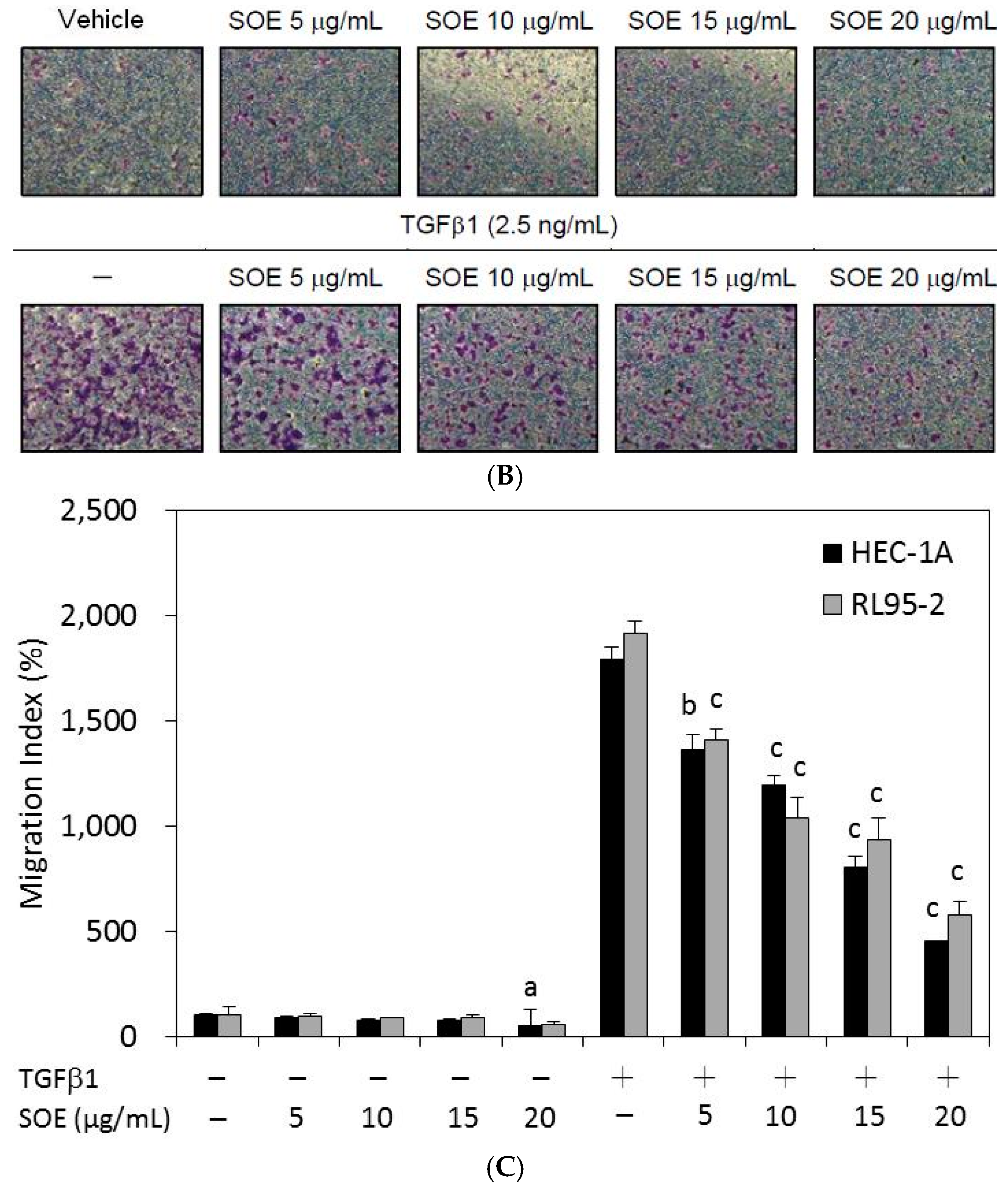
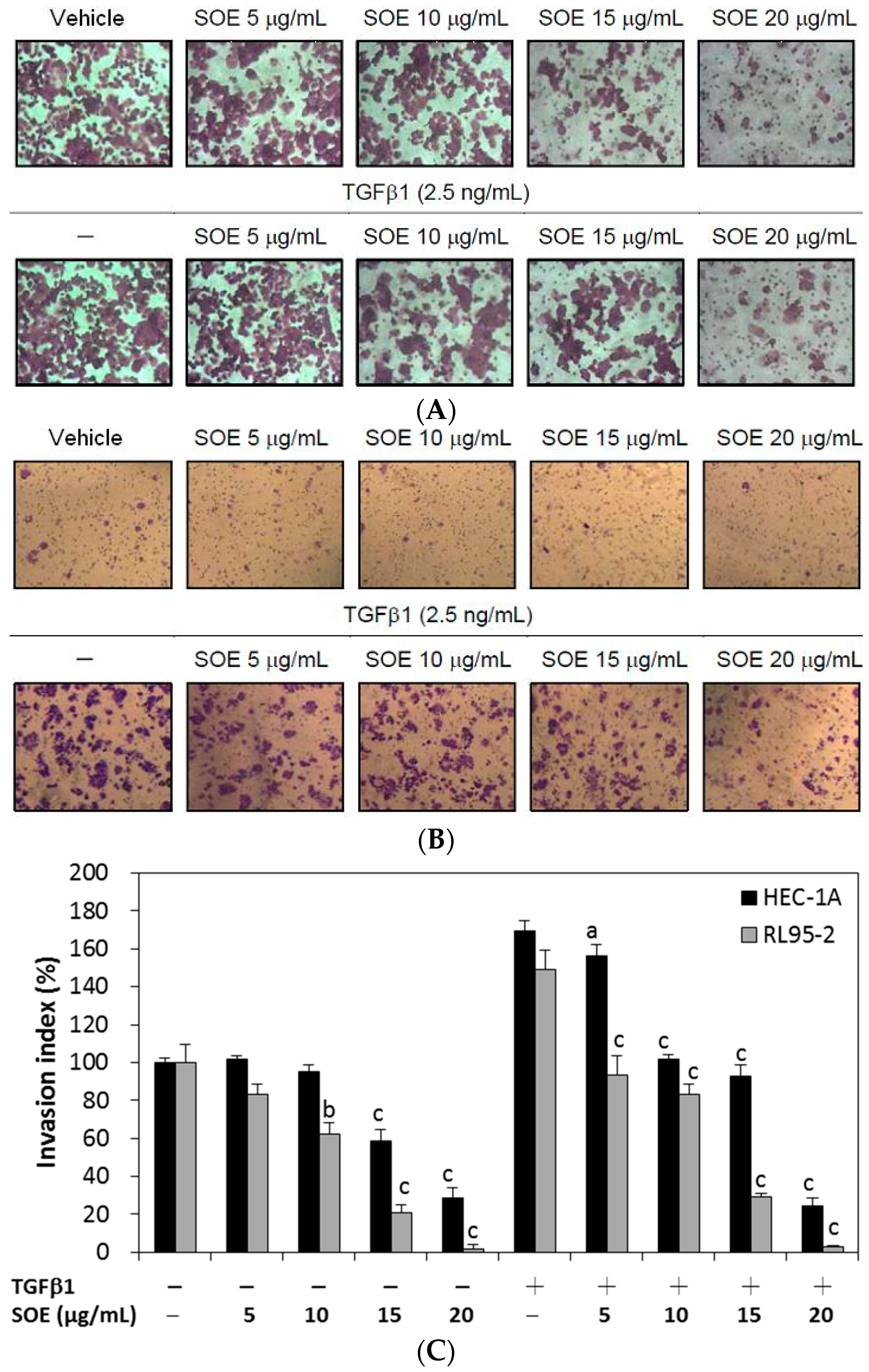
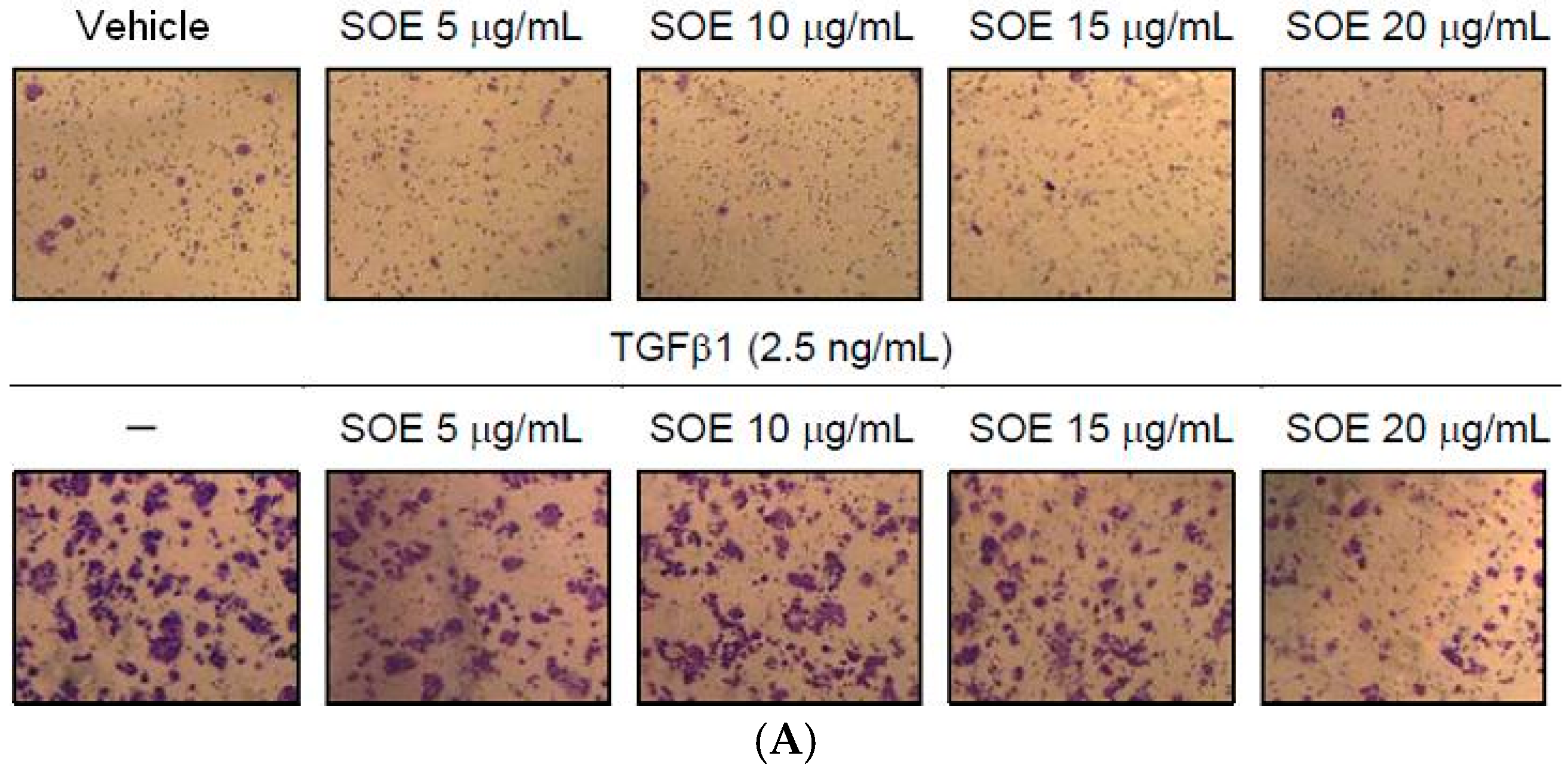
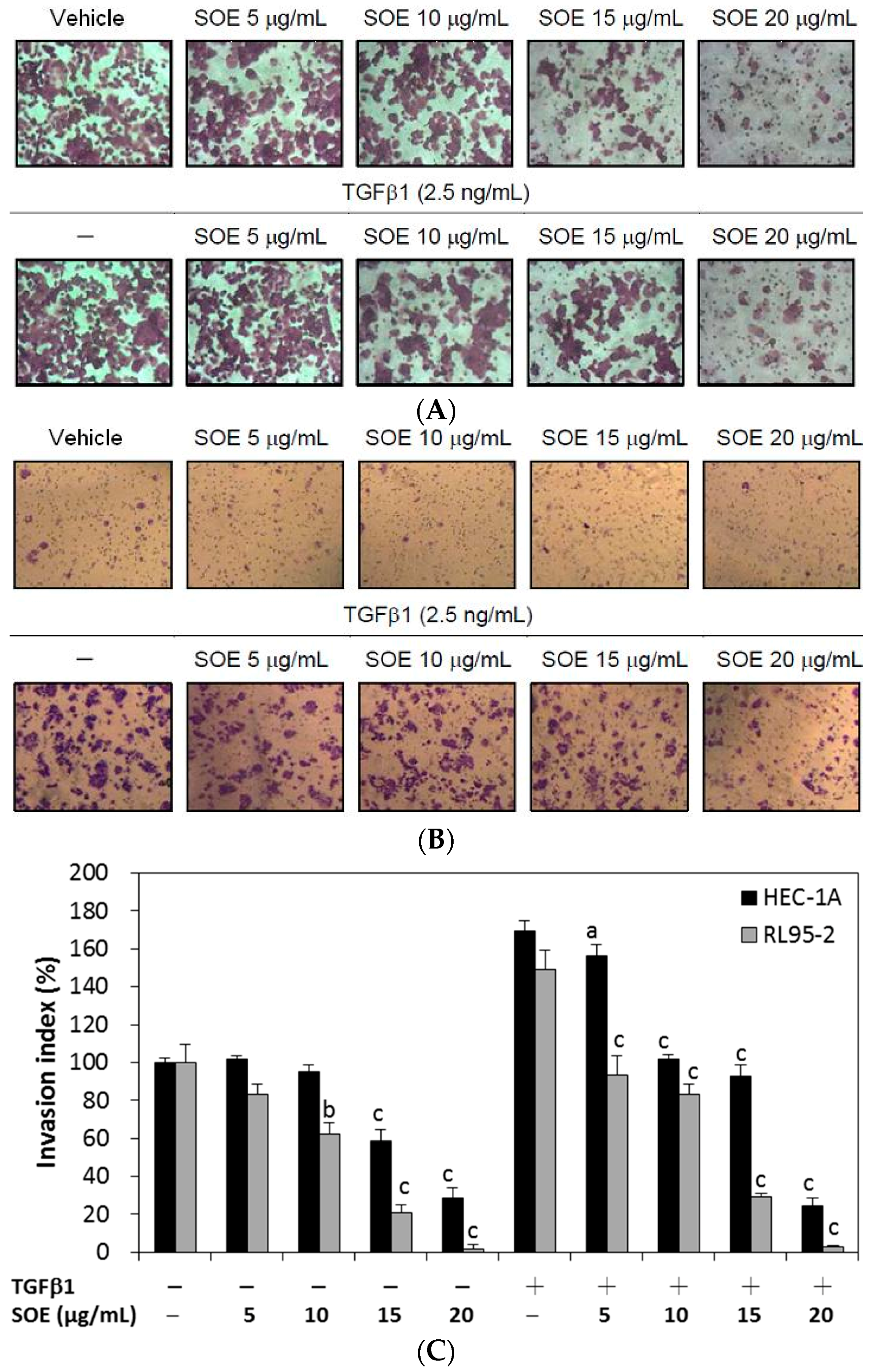
2.4. Inhibitory Effect of SOE on Invasion of Endometrial Cancer Cells
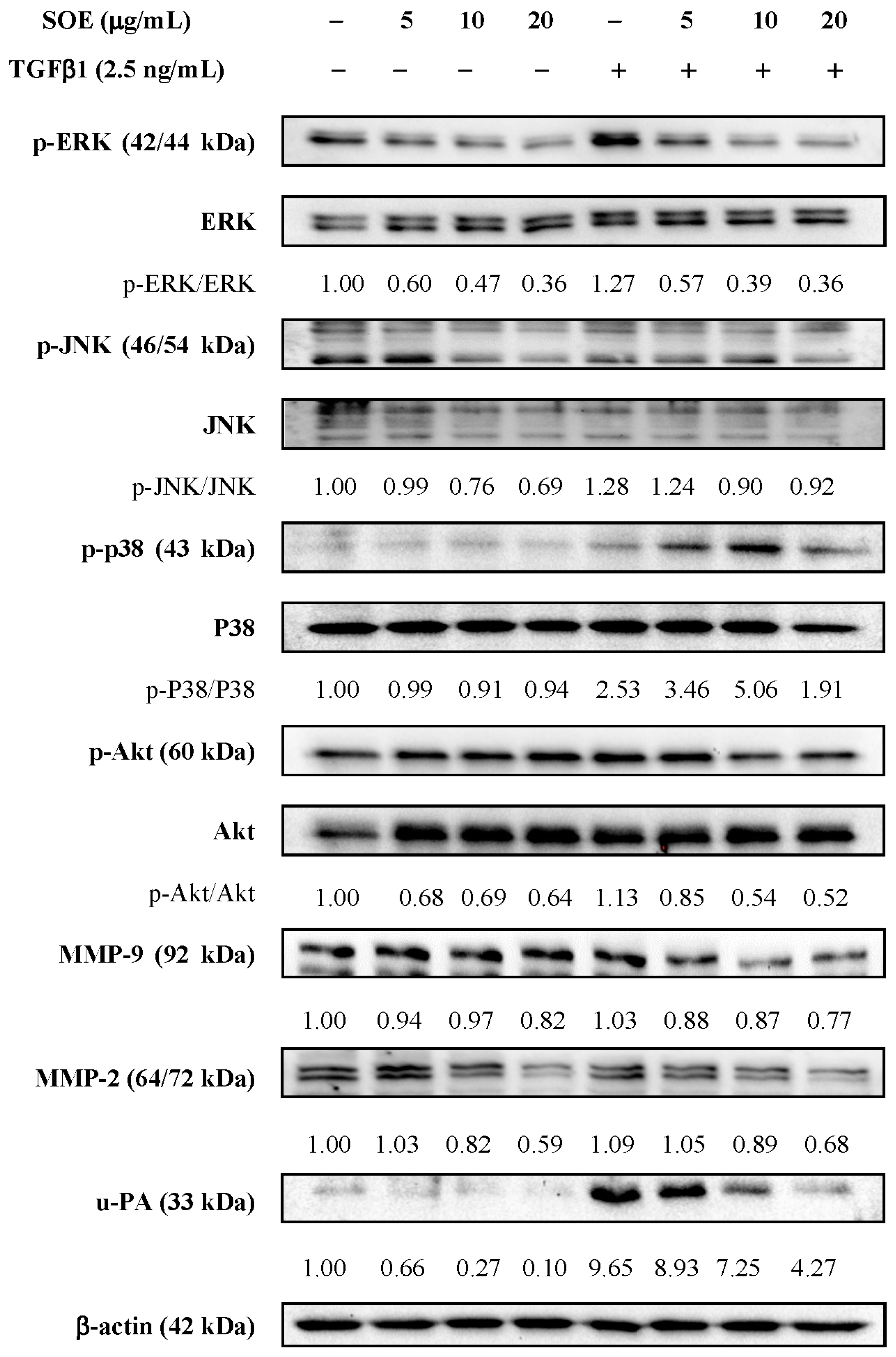
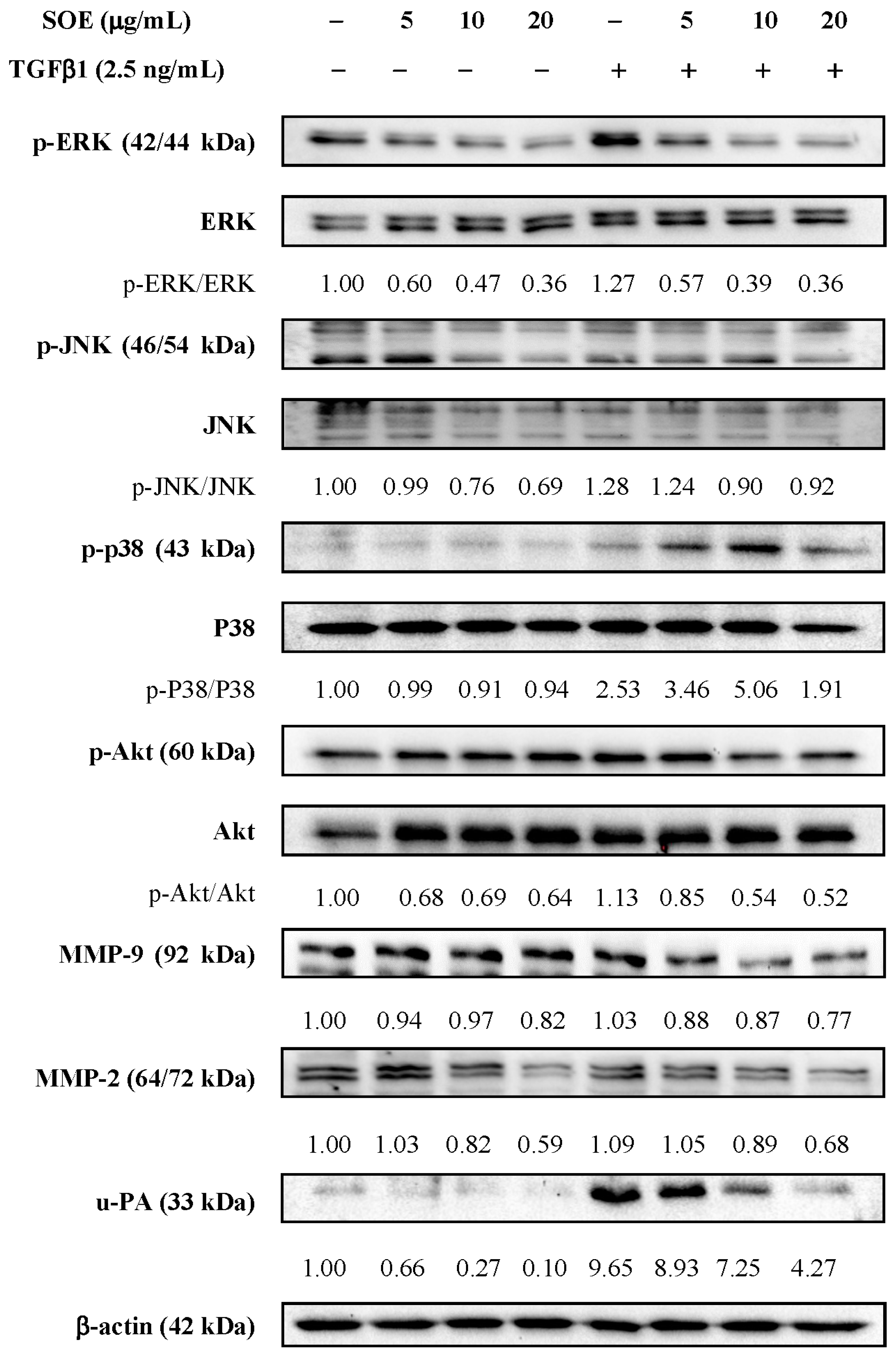
2.5. Effects of SOE on Related Proteins Expression in RL95-2 Cells
3. Materials and Methods
3.1. Materials
3.2. Preparation of S. orientalis Ethanol Extract
3.3. Cancer Cell Lines and Culture
3.4. Determination of Cytotoxicity for Cancer Cells
3.5. Cell Migration by Wound Healing Assay
3.6. Cell Migration and Invasion Assays by Boyden Chamber
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Colombo, N.; Preti, E.; Landoni, F.; Carinelli, S.; Colombo, A.; Marini, C.; Sessa, C.; ESMO Guidelines Working Group. Endometrial cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 6, vi33–vi38. [Google Scholar]
- Harkenrider, M.M.; Block, A.M.; Siddiqui, Z.A.; Small, W., Jr. The role of vaginal cuff brachytherapy in endometrial cancer. Gynecol. Oncol. 2015, 136, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.Y.; Chen, Y.J.; Yen, M.S.; Chao, K.C.; Chen, R.F.; Yeh, S.O.; Twu, N.F. Clinicopathological features and survival in young Taiwanese women with endometrial carcinoma. Int. J. Gynecol. Cancer 2014, 24, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Bokhman, J.V. Two pathogenetic types of endometrial carcinoma. Gynecol. Oncol. 1983, 15, 10–17. [Google Scholar] [CrossRef]
- Abal, M.; Llaurado, M.; Doll, A.; Monge, M.; Colas, E.; Gonzalez, M.; Rigau, M.; Alazzouzi, H.; Demajo, S.; Castellví, J.; et al. Molecular determinants of invasion in endometrial cancer. Clin. Transl. Oncol. 2007, 9, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Muinelo-Romay, L.; Colas, E.; Barbazan, J.; Alonso-Alconada, L.; Alonso-Nocelo, M.; Bouso, M.; Curiel, T.; Cueva, J.; Anido, U.; Forteza, J.; et al. High-risk endometrial carcinoma profiling identifies TGF-β1 as a key factor in the initiation of tumor invasion. Mol. Cancer Ther. 2011, 10, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Drabsch, Y.; ten Dijke, P. TGF-β signalling and its role in cancer progression and metastasis. Cancer Metastasis Rev. 2012, 31, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, P.J.; Ebner, R.; Lopez, A.R.; Derynck, R. TGF-β induced transdifferentiation of mammary epithelial cells to mesenchymal cells: Involvement of type I receptors. J. Cell Biol. 1994, 127, 2021–2036. [Google Scholar] [CrossRef] [PubMed]
- Perlino, E.; Loverro, G.; Maiorano, E.; Giannini, T.; Cazzolla, A.; Napoli, A.; Fiore, M.G.; Ricco, R.; Marra, E.; Selvaggi, L. Down-regulated expression of transforming growth factor β1 mRNA in endometrial carcinoma. Br. J. Cancer 1998, 77, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Yabushita, H.; Narumiya, H.; Hiratake, K.; Yamada, H.; Shimazu, M.; Sawaguchi, K.; Noguchi, M.; Nakanishi, M. The association of transforming growth factor-β1 with myometrial invasion of endometrial carcinomas through effects on matrix metalloproteinase. J. Obstet. Gynaecol. Res. 2000, 26, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Chopra, V.; Dinh, T.V.; Hannigan, E.V. Serum levels of interleukins, growth factors and angiogenin in patients with endometrial cancer. J. Cancer Res. Clin. Oncol. 1997, 123, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Teicher, B.A. Malignant cells, directors of the malignant process: Role of transforming growth factor-β. Cancer Metastasis Rev. 2001, 20, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.I.; Saxena, B.; Mittal, K.R.; Marmor, M.; Goswami, S.; Nactigal, L.; Korc, M.; Demopoulos, R.I. Increased expression of transforming growth factor β isoforms and basic fibroblast growth factor in complex hyperplasia and adenocarcinoma of the endometrium: Evidence for paracrine and autocrine action. Cancer Res. 1994, 54, 2347–2358. [Google Scholar] [PubMed]
- Bokhari, A.A.; Syed, V. Inhibition of Transforming Growth Factor-β (TGF-β) Signaling by Scutellaria baicalensis and Fritillaria cirrhosa Extracts in Endometrial Cancer. J. Cell. Biochem. 2015, 116, 1797–1805. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K. Adjuvant treatment of advanced-stage endometrial cancer. Clin. Obstet. Gynecol. 2011, 54, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Randall, M.E.; Filiaci, V.L.; Muss, H.; Spirtos, N.M.; Mannel, R.S.; Fowler, J.; Thigpen, J.T.; Benda, J.A.; Gynecologic Oncology Group Study. Randomized phase III trial of whole-abdominal irradiation versus doxorubicin and cisplatin chemotherapy in advanced endometrial carcinoma: A Gynecologic Oncology Group Study. J. Clin. Oncol. 2006, 24, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Wang, N.; Tan, H.Y.; Tsao, S.W.; Feng, Y. MicroRNAs and Chinese medicinal herbs: New possibilities in cancer therapy. Cancers 2015, 7, 1643–1657. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; An, H.M.; Wang, S.S.; Chen, J.J.; Xu, L. Preventive and therapeutic effects of Chinese herbal compounds against hepatocellular carcinoma. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Qian, R.; Zhang, C.; Fu, H. Study on therapeutic mechanism of anti-rheumatism action of Herba siegesbeckiae. Zhongguo Zhong Xi Yi Jie He Za Zhi 2000, 20, 192–195. [Google Scholar] [PubMed]
- Wang, J.P.; Zhou, Y.M.; Ye, Y.J.; Shang, X.M.; Cai, Y.L.; Xiong, C.M.; Wu, Y.X.; Xu, H.X. Topical anti-inflammatory and analgesic activity of kirenol isolated from Siegesbeckia orientalis. J. Ethnopharmacol. 2011, 137, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.J.; Park, E.J.; Jang, C.H.; Han, S.W.; Oh, G.J.; Kim, N.S.; Kim, H.M. Inhibitory effect of immunoglobulin E production by jin-deuk-chal (Siegesbeckia orientalis). Immunopharmacol. Immunotoxicol. 2001, 23, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Wang, H. Immunosuppressive activity of the ethanol extract of Siegesbeckia orientalis on the immune responses to ovalbumin in mice. Chem. Biodivers. 2006, 3, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cheng, X.L.; Li, Y.J.; Shi, S.; Liu, J.K. ent-Pimarane diterpenoids from Siegesbeckia orientalis and structure revision of a related compound. J. Nat. Prod. 2009, 72, 2005–2008. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Luo, Q.; Ruan, J.L.; Chen, Y.A.; Chen, M.X. Effect of Siegesbeckia orientalis L. on cervical cancer HeLa cell in vitro. Her. Med. 2009, 8, 45–46. (In Chinese) [Google Scholar]
- Hong, Y.H.; Weng, L.W.; Chang, C.C.; Hsu, H.F.; Wang, C.P.; Wang, S.W.; Houng, J.Y. Anti-inflammatory effects of Siegesbeckia orientalis ethanol extract in in vitro and in vivo models. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef]
- Chang, C.C.; Hsu, H.F.; Huang, K.H.; Wu, J.M.; Kuo, S.M.; Ling, X.H.; Houng, J.Y. Anti-proliferative effects of Siegesbeckia orientalis ethanol extract on human endometrial RL-95 cancer cells. Molecules 2014, 19, 19980–19994. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.C.; Park, H.Y.; Hwang, H.S.; Kang, S.U.; Pyun, J.H.; Lee, M.H.; Choi, E.C.; Kim, C.H. (−)-Epigallocatechin-3-gallate (EGCG) inhibits HGF-induced invasion and metastasis in hypopharyngeal carcinoma cells. Cancer Lett. 2008, 271, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.N.; Hsieh, Y.S.; Chiou, H.L.; Chu, S.C. Silibinin inhibits cell invasion through inactivation of both PI3K-Akt and MAPK signaling pathways. Chem. Biol. Interact. 2005, 156, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, M.; Lund, L.R.; Romer, J.; Almholt, K.; Dano, K. Cancer invasion and tissue remodeling: Common themes in proteolytic matrix degradation. Curr. Opin. Cell Biol. 1998, 10, 667–671. [Google Scholar] [CrossRef]
- Chambers, A.F.; Matrisian, L.M. Changing views of the role of matrix metalloproteinases in metastasis. J. Natl. Cancer Inst. 1997, 89, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Kajanne, R.; Miettinen, P.; Mehlem, A.; Leivonen, S.K.; Birrer, M.; Foschi, M.; Kahari, V.M.; Leppa, S. EGF-R regulates MMP function in fibroblasts through MAPK and AP-1 pathways. J. Cell. Physiol. 2007, 212, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Nho, K.J.; Chun, J.M.; Lee, A.Y.; Kim, H.K. Anti-metastatic effects of Rheum Palmatum L. extract in human MDA-MB-231 breast cancer cells. Environ. Toxicol. Pharmacol. 2015, 40, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Geng, Y.Q.; An, L.L.; Wu, K.F. Expression of gelatinase A and B in cord blood CD34+ cells and leukemic cell lines. Zhonghua Xue Ye Xue Za Zhi 2003, 24, 78–81. [Google Scholar] [PubMed]
- Lee, Y.C.; Lin, H.H.; Hsu, C.H.; Wang, C.J.; Chiang, T.A.; Chen, J.H. Inhibitory effects of andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur. J. Pharmacol. 2010, 632, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Yang, H.W.; Su, C.W.; Lue, K.H.; Yang, S.F.; Hsieh, Y.S. Phyllanthus urinaria suppresses human osteosarcoma cell invasion and migration by transcriptionally inhibiting u-PA via ERK and Akt signaling pathways. Food Chem. Toxicol. 2013, 52, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gao, Q.; Chen, K.; Wang, Y.; Chen, L.; Li, X.U. Curcumin suppresses migration and invasion of human endometrial carcinoma cells. Oncol. Lett. 2015, 10, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, X.; Zhang, H.; Wang, Y.; Li, Y. MPA influences tumor cell proliferation, migration, and invasion induced by RANKL through PRB involving the MAPK pathway in endometrial cancer. Oncol. Rep. 2015, 33, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.F.; Chang, S.F.; Chen, Z.H.; Yuan, S.S.F.; Tsai, Y.D.; Wang, C.P.; Wang, S.W.; Fang, L.W.; Houng, J.Y. Cytotoxic effect of Anisomeles indica extract on human pharynx squamous cancer cells. J. Med. Plants Res. 2012, 6, 5002–5012. [Google Scholar] [CrossRef]
- Sample Availability: Samples are not available.


© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-C.; Ling, X.-H.; Hsu, H.-F.; Wu, J.-M.; Wang, C.-P.; Yang, J.-F.; Fang, L.-W.; Houng, J.-Y. Siegesbeckia orientalis Extract Inhibits TGFβ1-Induced Migration and Invasion of Endometrial Cancer Cells. Molecules 2016, 21, 1021. https://doi.org/10.3390/molecules21081021
Chang C-C, Ling X-H, Hsu H-F, Wu J-M, Wang C-P, Yang J-F, Fang L-W, Houng J-Y. Siegesbeckia orientalis Extract Inhibits TGFβ1-Induced Migration and Invasion of Endometrial Cancer Cells. Molecules. 2016; 21(8):1021. https://doi.org/10.3390/molecules21081021
Chicago/Turabian StyleChang, Chi-Chang, Xue-Hua Ling, Hsia-Fen Hsu, Jing-Mei Wu, Chao-Ping Wang, Jyh-Ferng Yang, Li-Wen Fang, and Jer-Yiing Houng. 2016. "Siegesbeckia orientalis Extract Inhibits TGFβ1-Induced Migration and Invasion of Endometrial Cancer Cells" Molecules 21, no. 8: 1021. https://doi.org/10.3390/molecules21081021
APA StyleChang, C.-C., Ling, X.-H., Hsu, H.-F., Wu, J.-M., Wang, C.-P., Yang, J.-F., Fang, L.-W., & Houng, J.-Y. (2016). Siegesbeckia orientalis Extract Inhibits TGFβ1-Induced Migration and Invasion of Endometrial Cancer Cells. Molecules, 21(8), 1021. https://doi.org/10.3390/molecules21081021