
Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Inflammatory Bowel Disease (IBD)
1.2. Overview of Catechins Activity
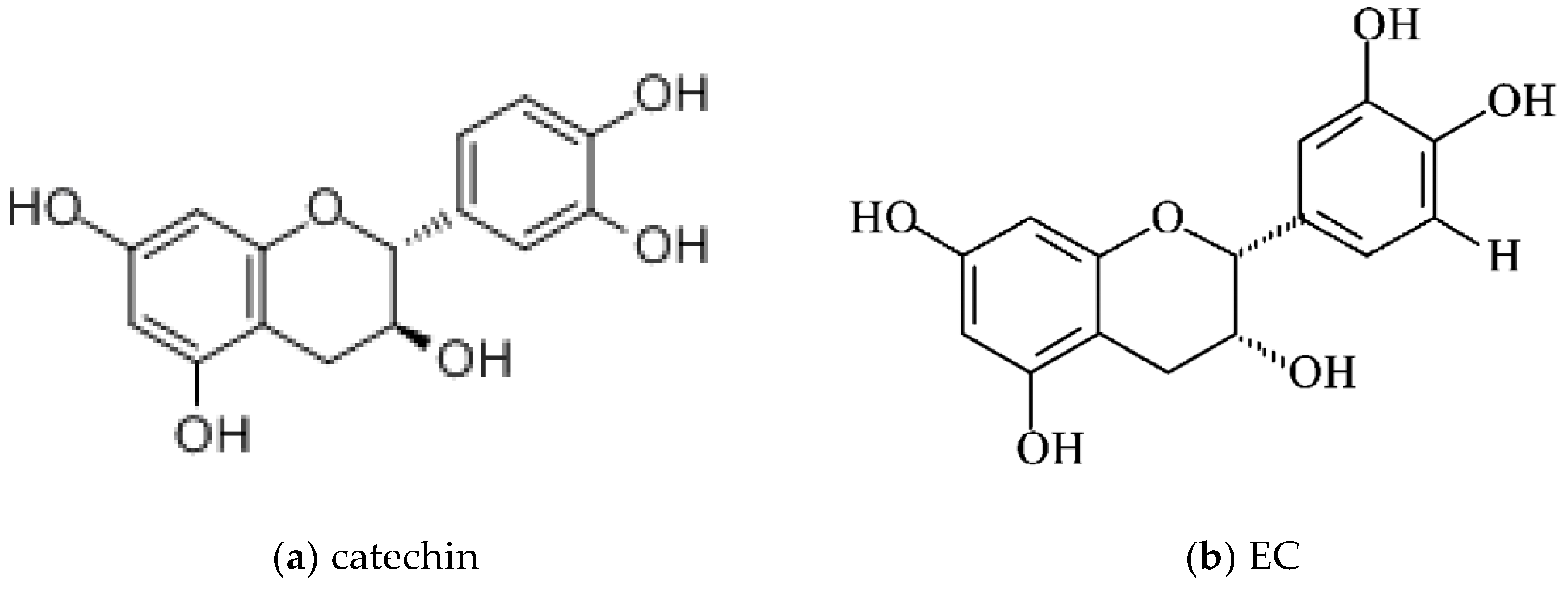
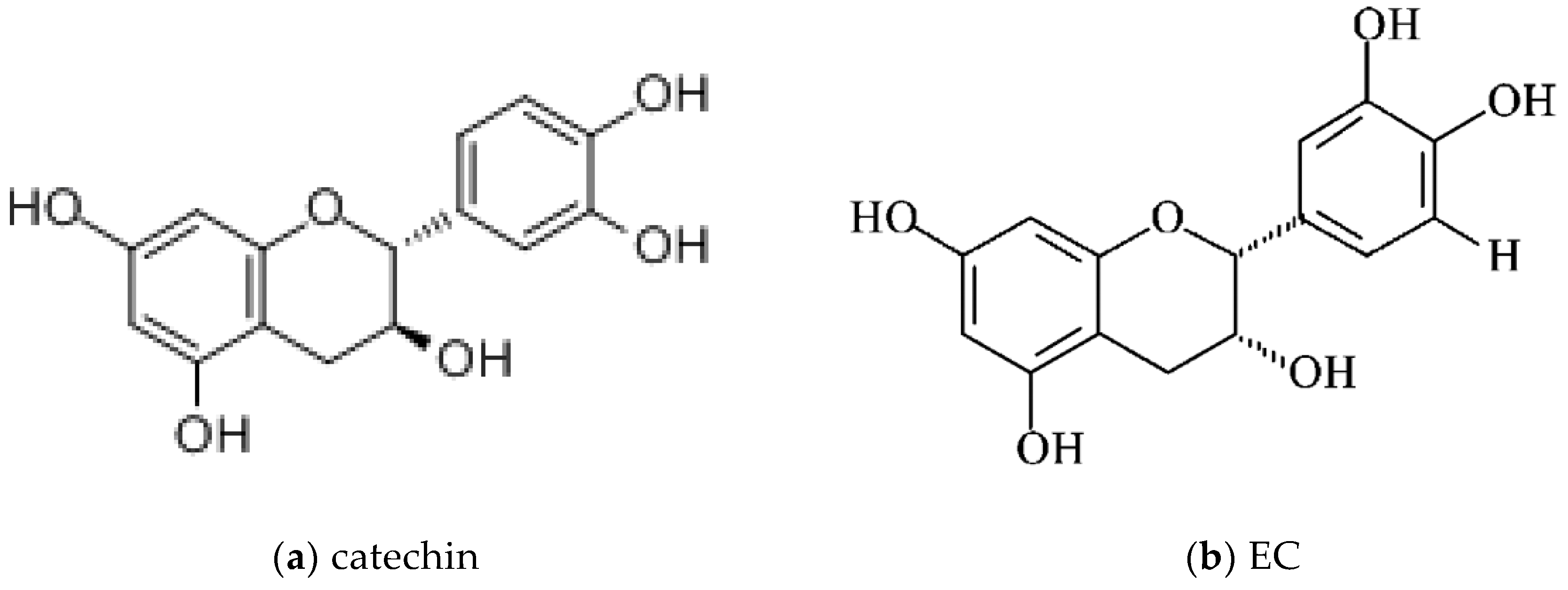
1.2.1. Catechins: Chemical Structures
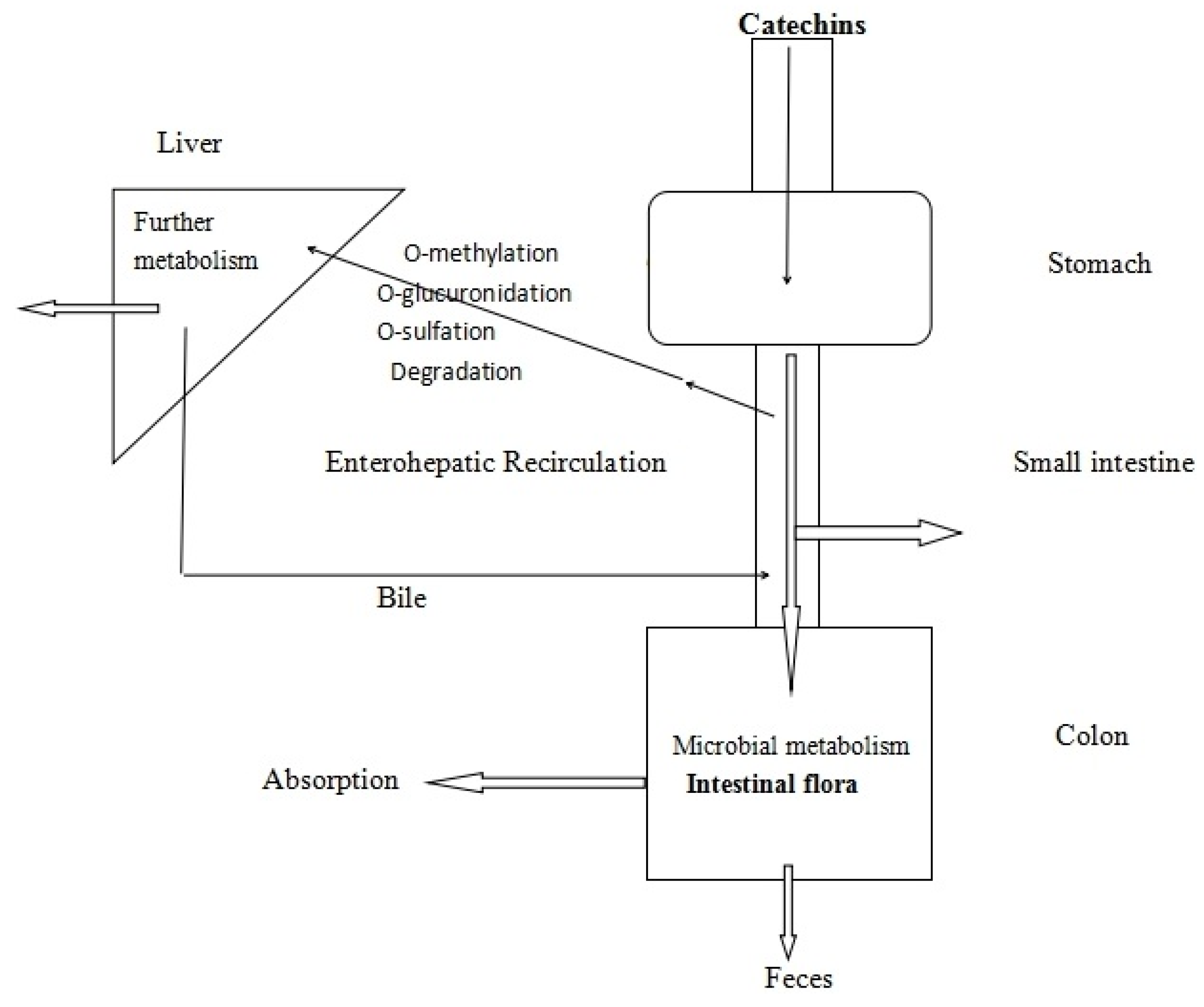
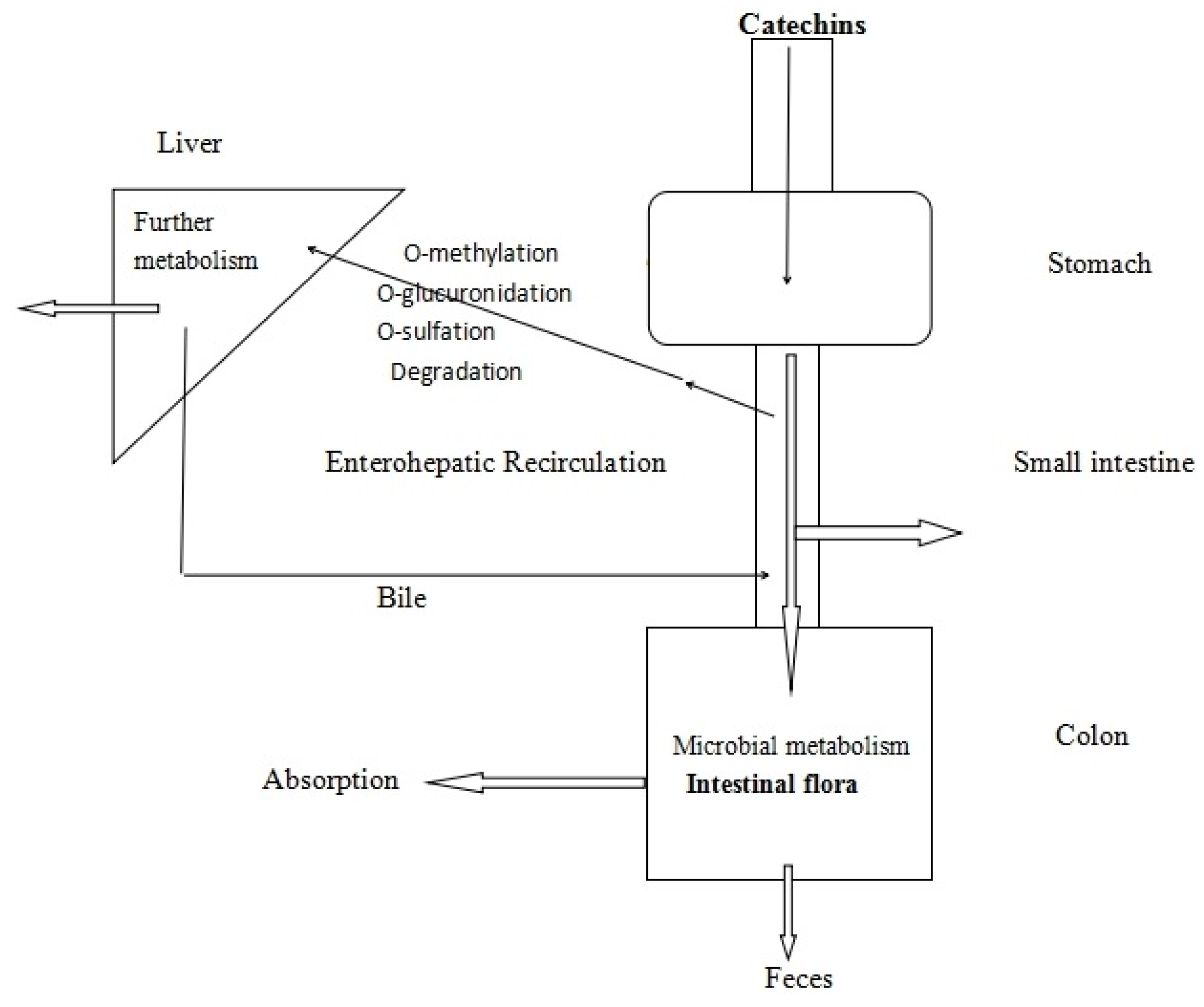
1.2.2. The Bioavailability of Catechins
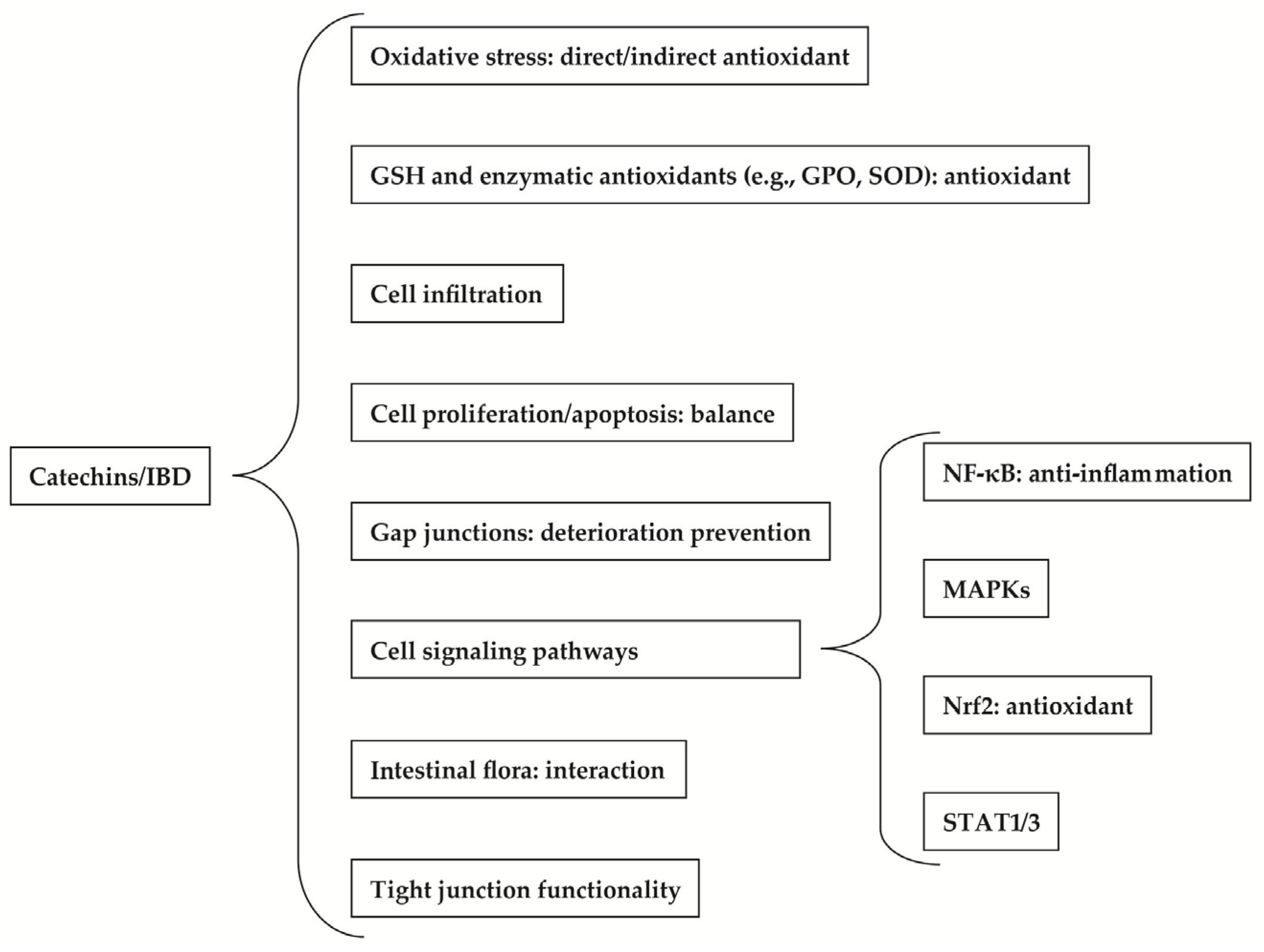
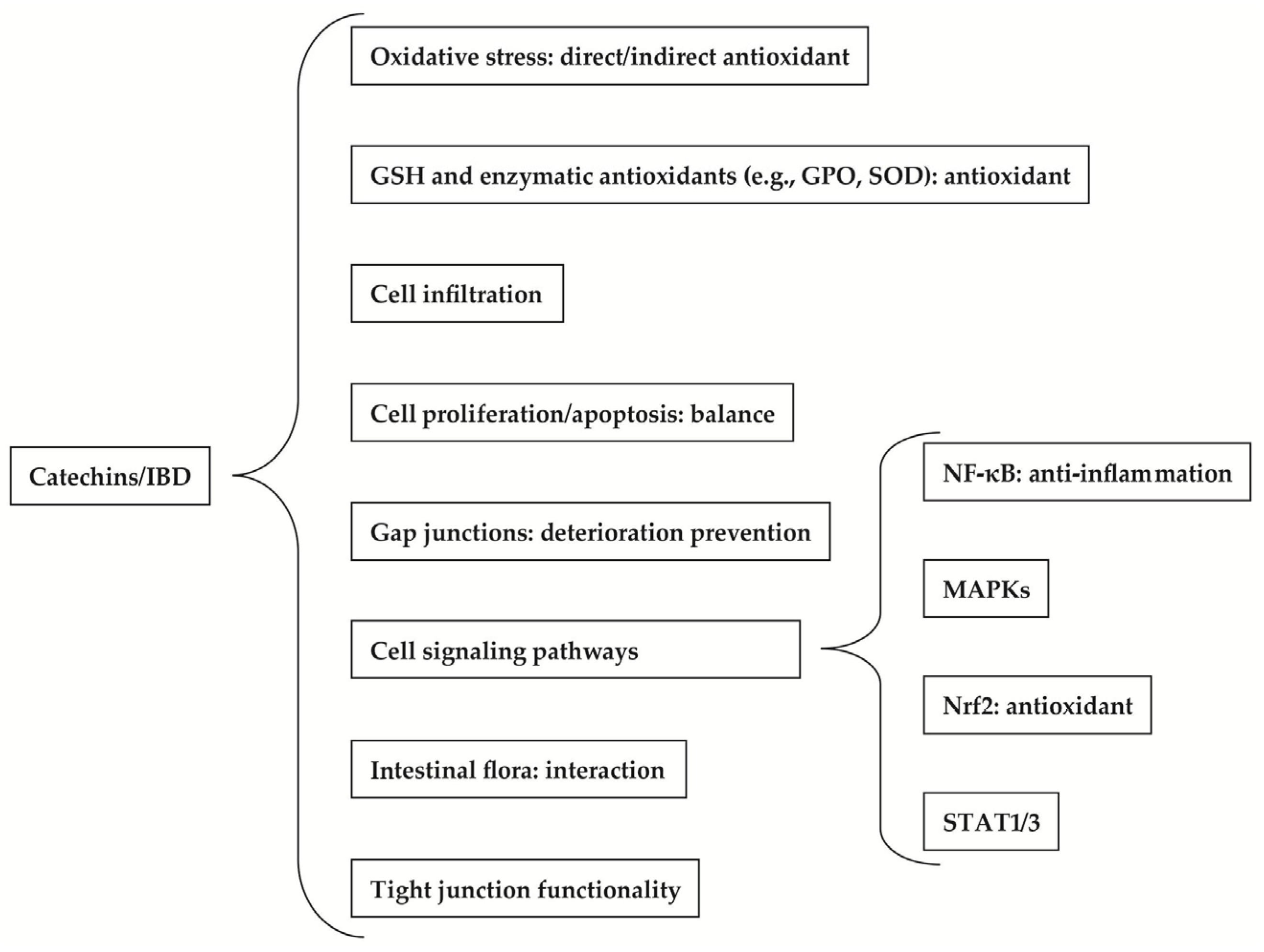
2. Therapeutic Benefits of Catechins to IBD
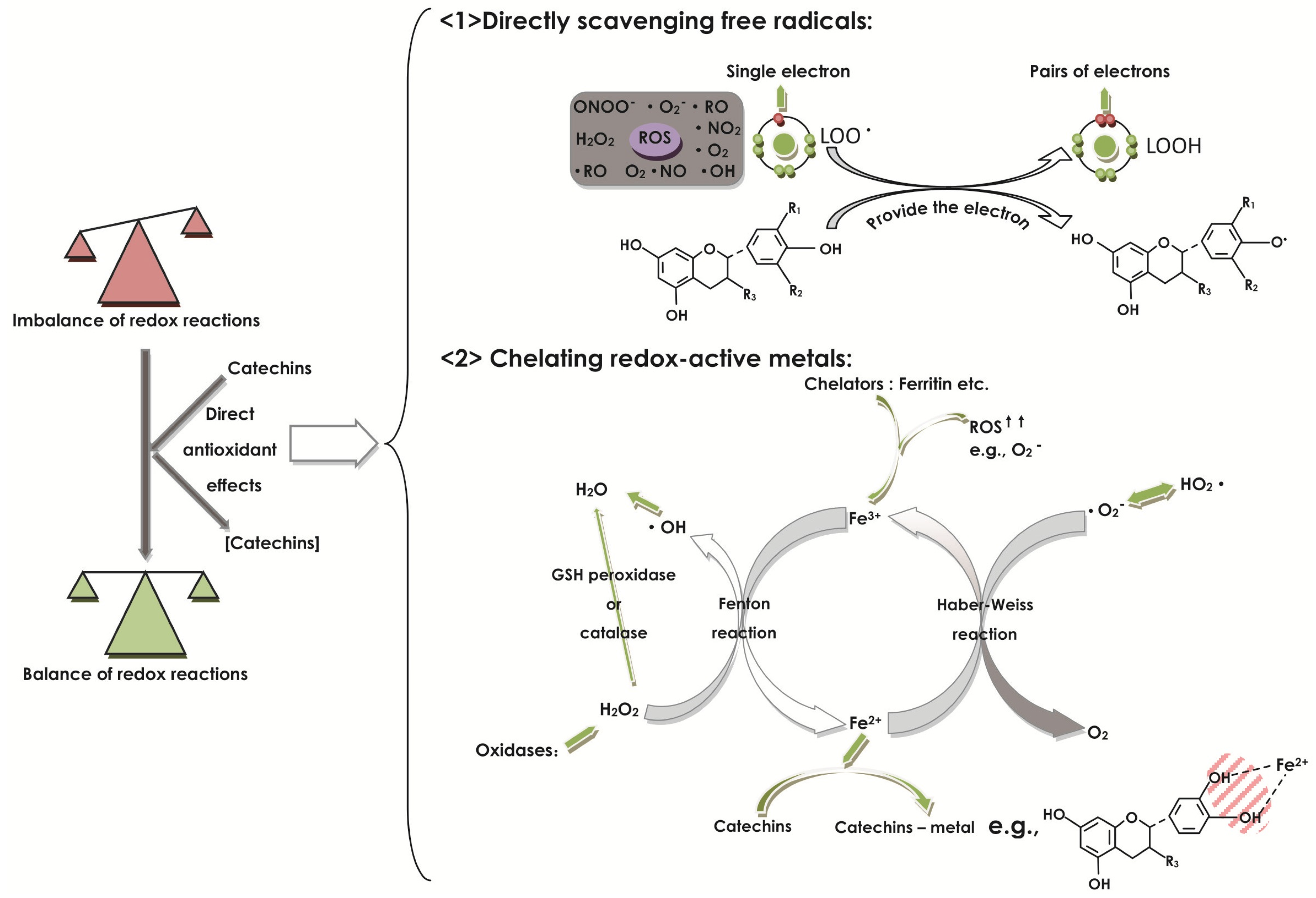
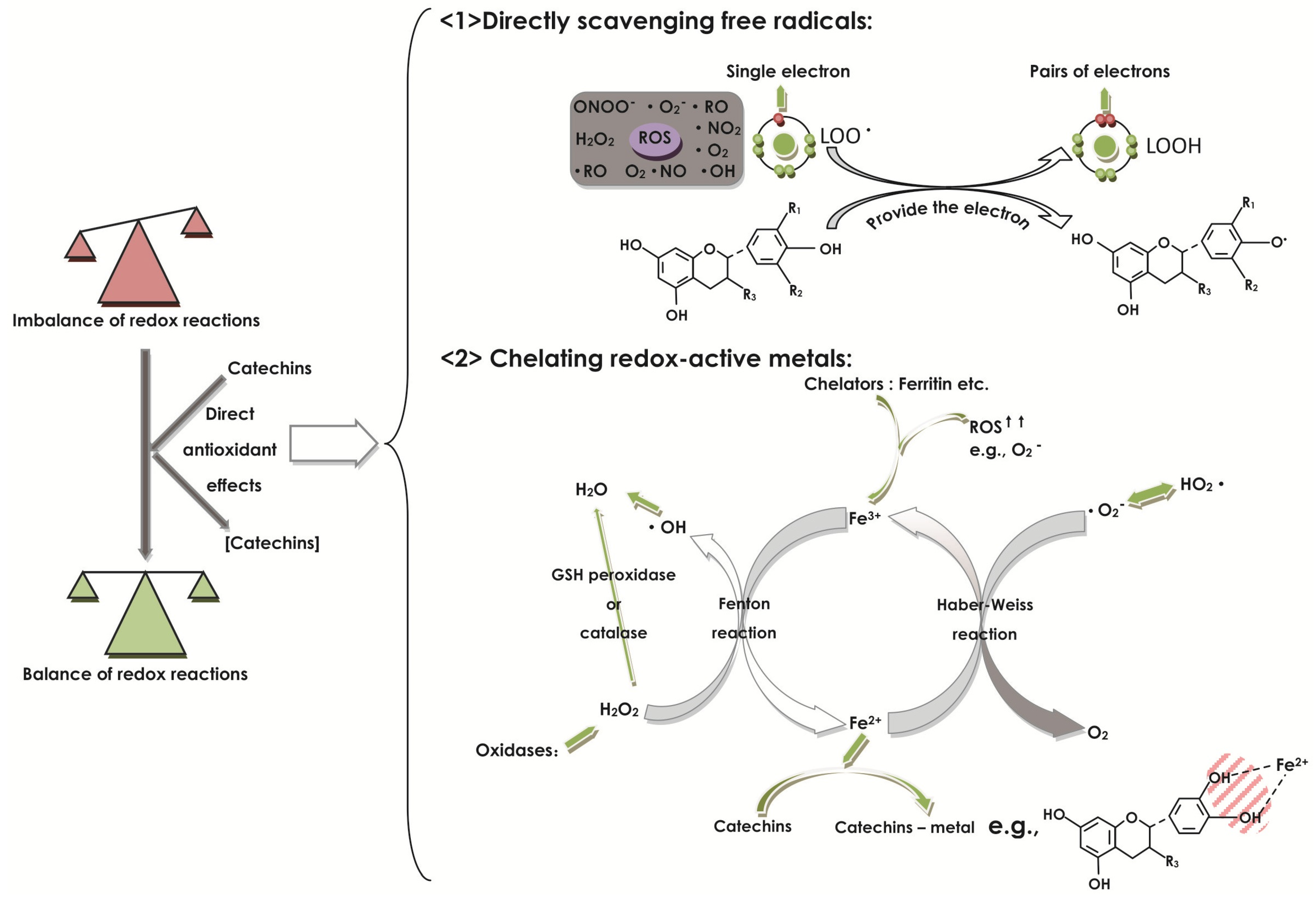
2.1. Effect of Catechins on Oxidative Stress
2.2. Effect of Catechins on GSH and Enzymatic Antioxidants
2.3. Effect of Catechins on Cell Infiltration
2.4. Effect of Catechins on Cell Proliferation and Apoptosis
2.5. Effect of Catechins on Gap Junctions
2.6. Effect of Catechins on Cell Signaling Pathways
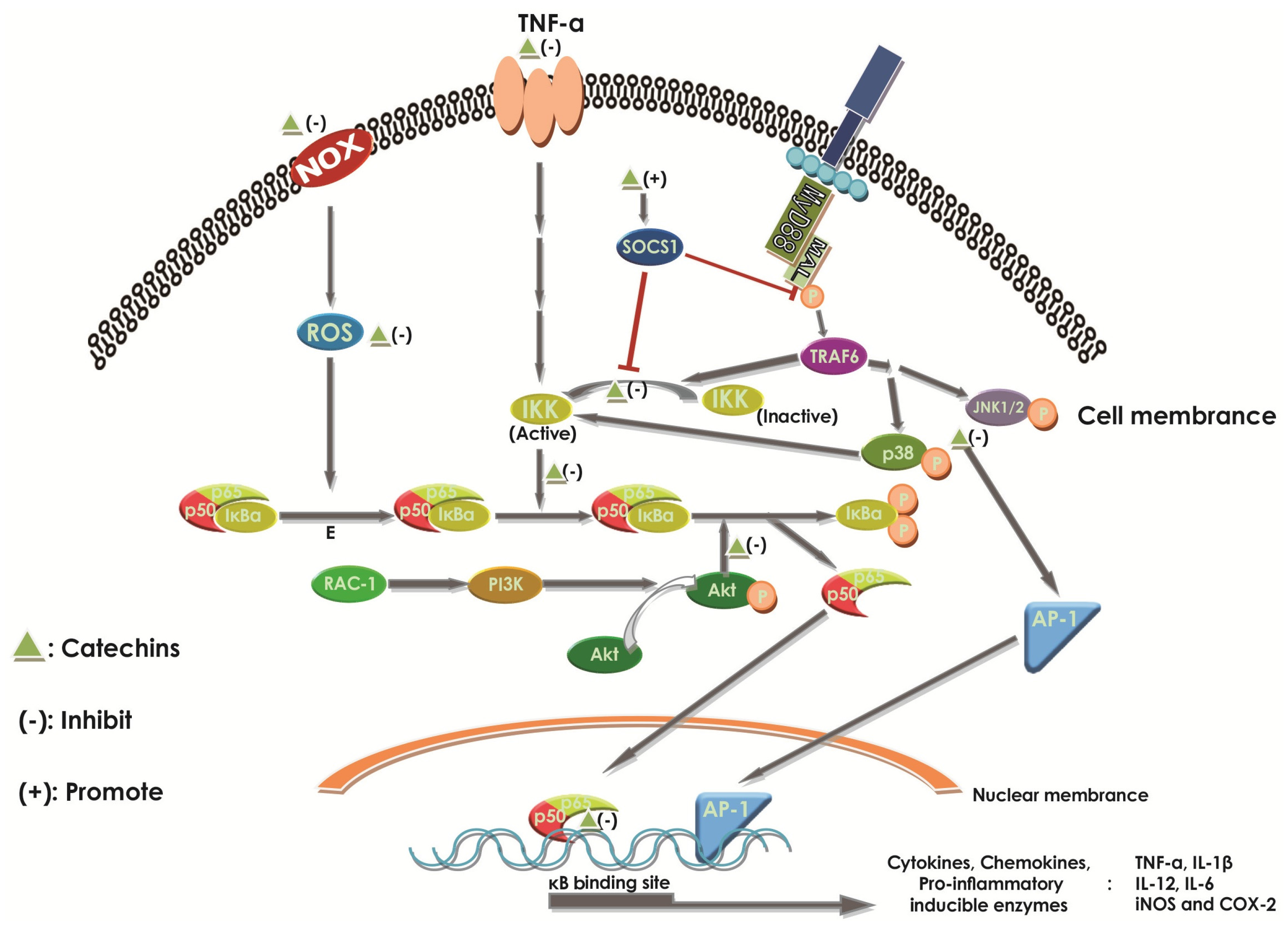
2.6.1. Effect of Catechins on NF-κB
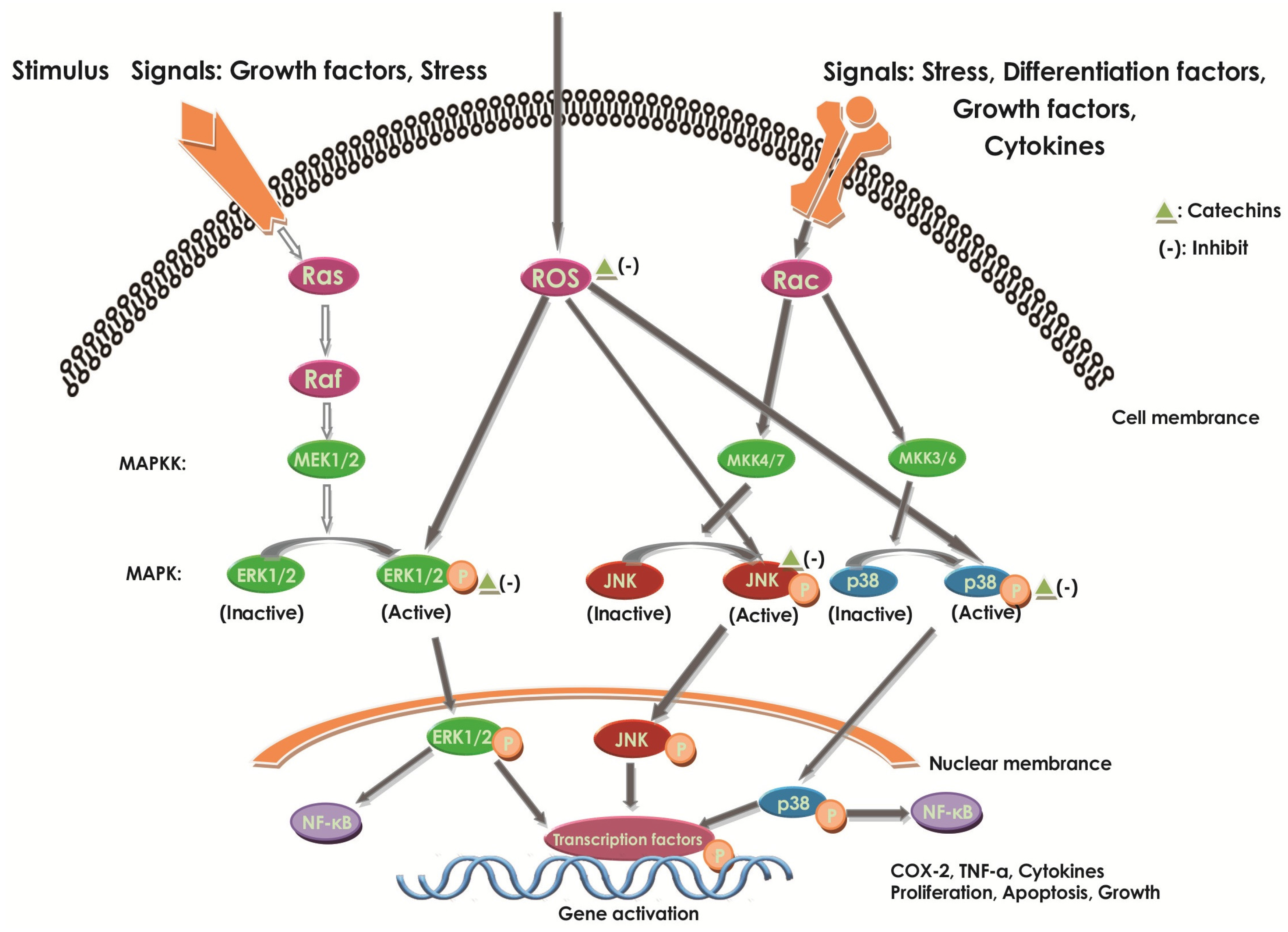
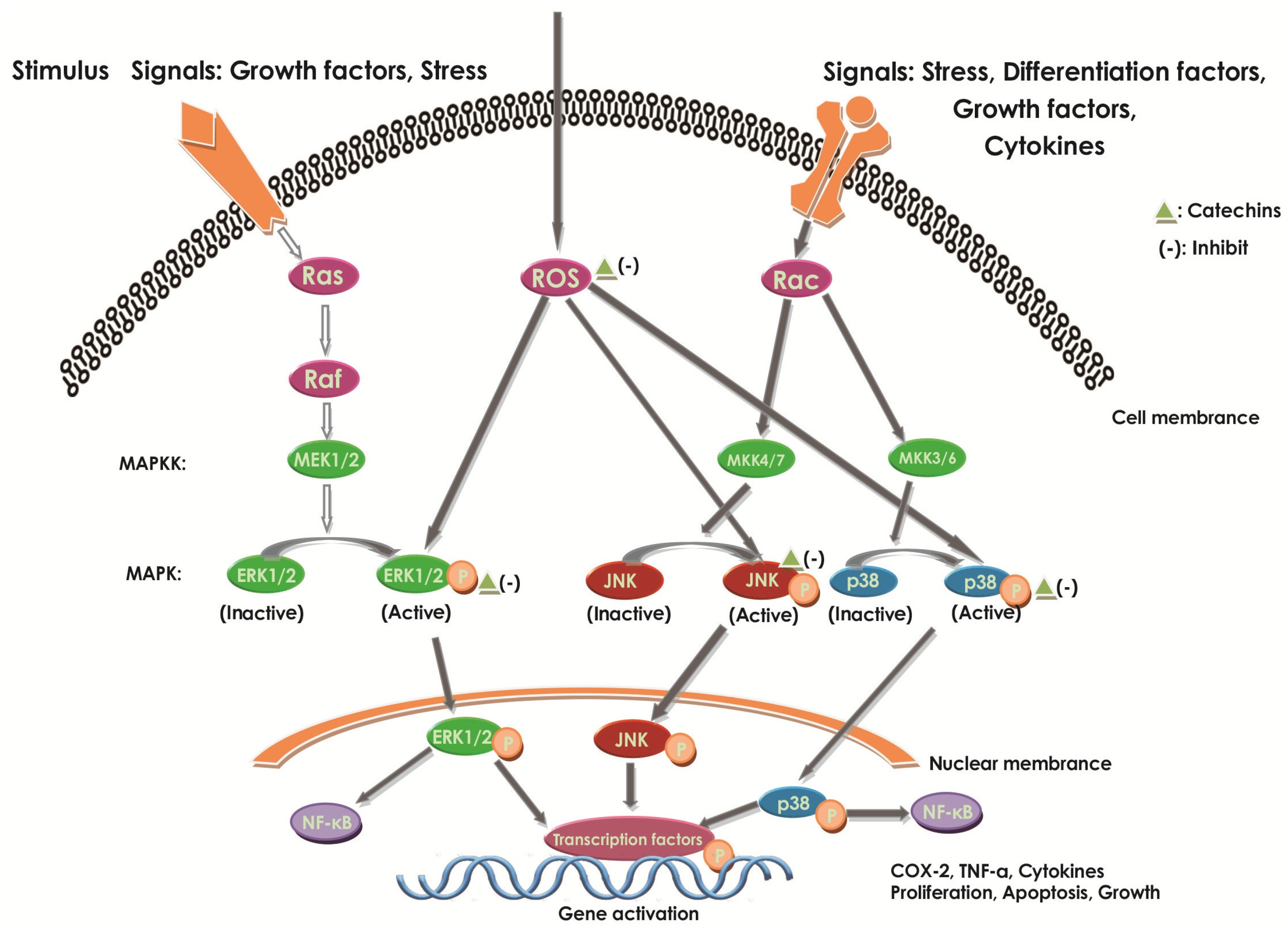
2.6.2. Effect of Catechins on MAPKs
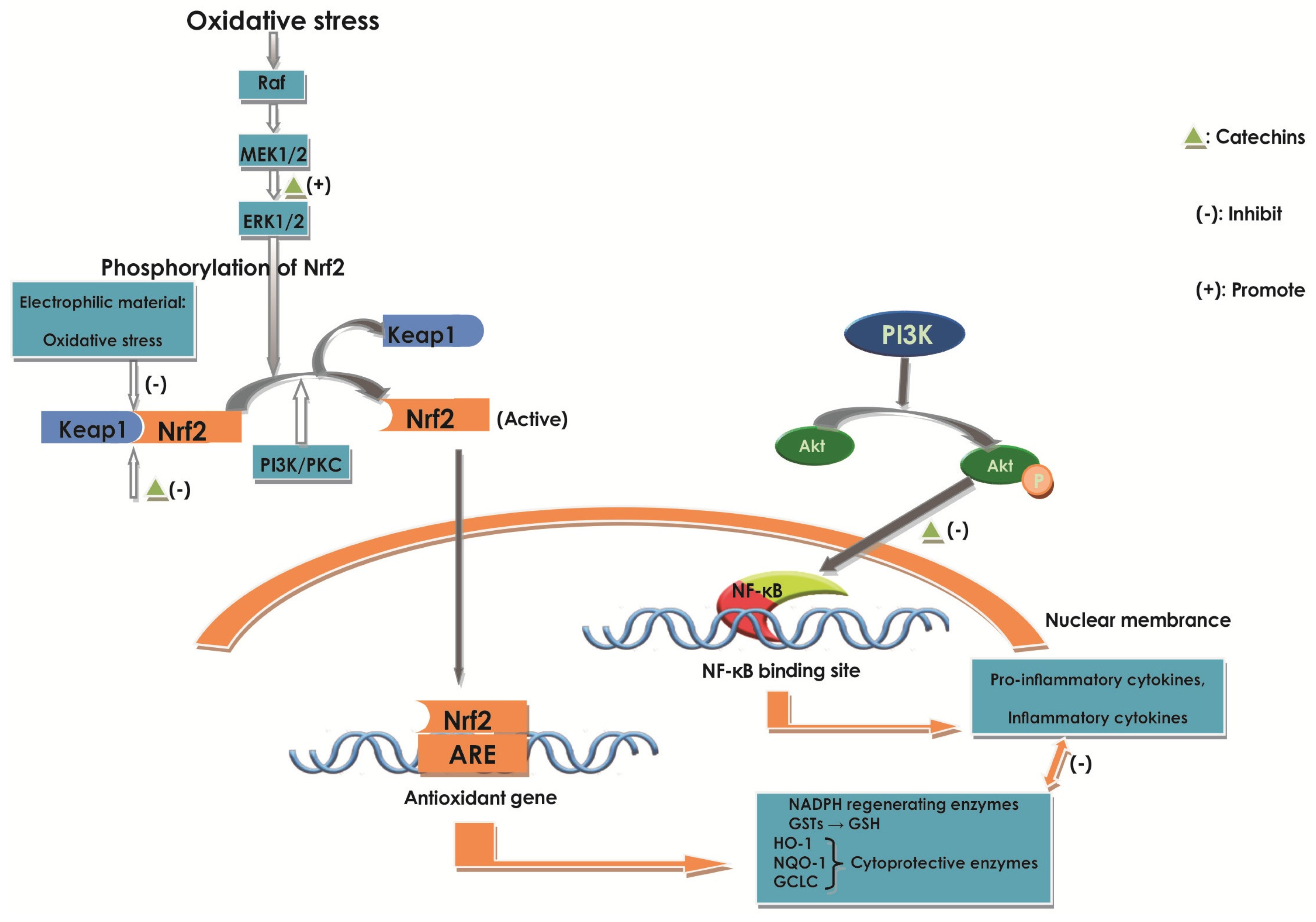
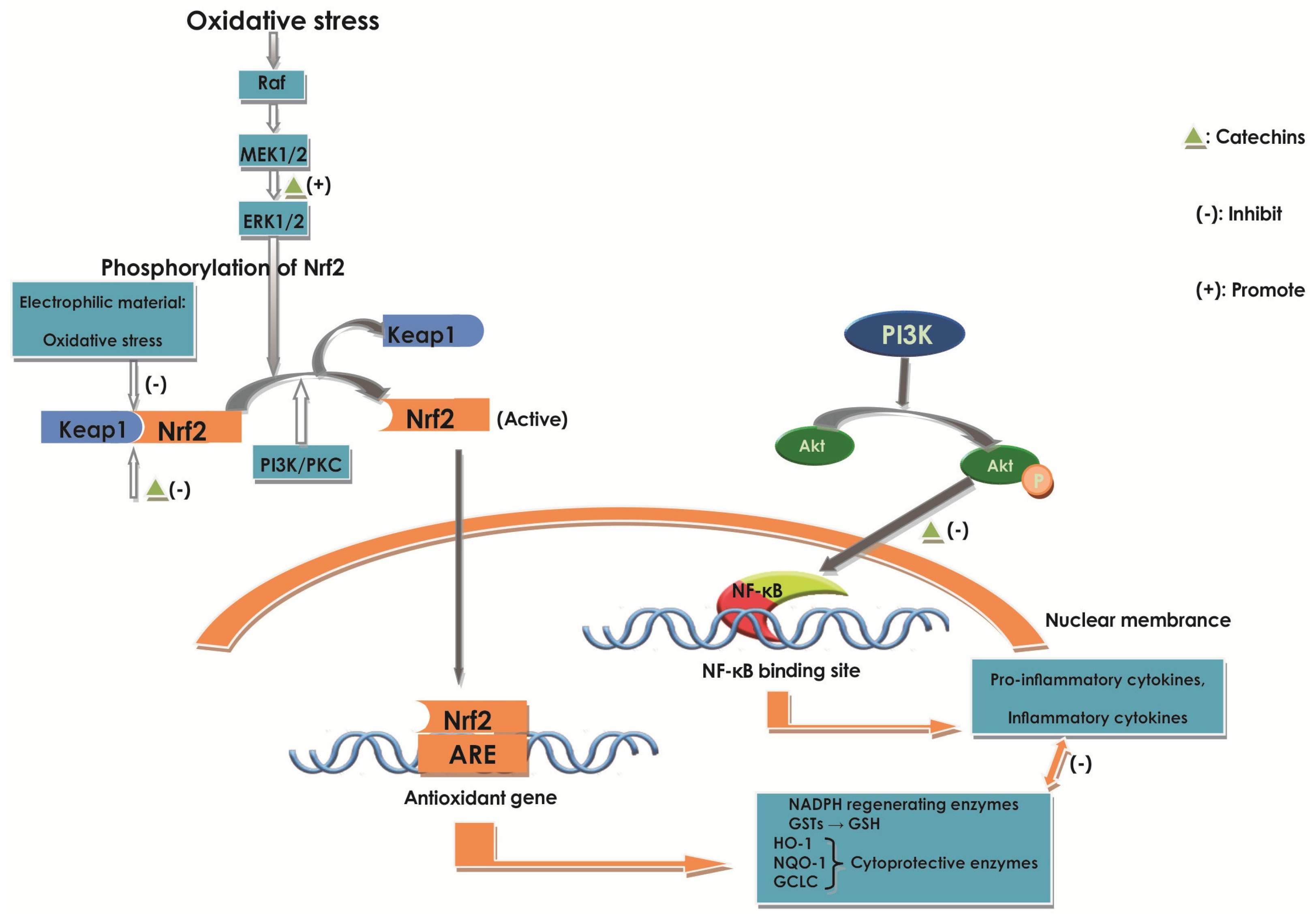
2.6.3. Effect of Catechins on Nrf2
2.6.4. Effect of Catechins on STAT1 and STAT3
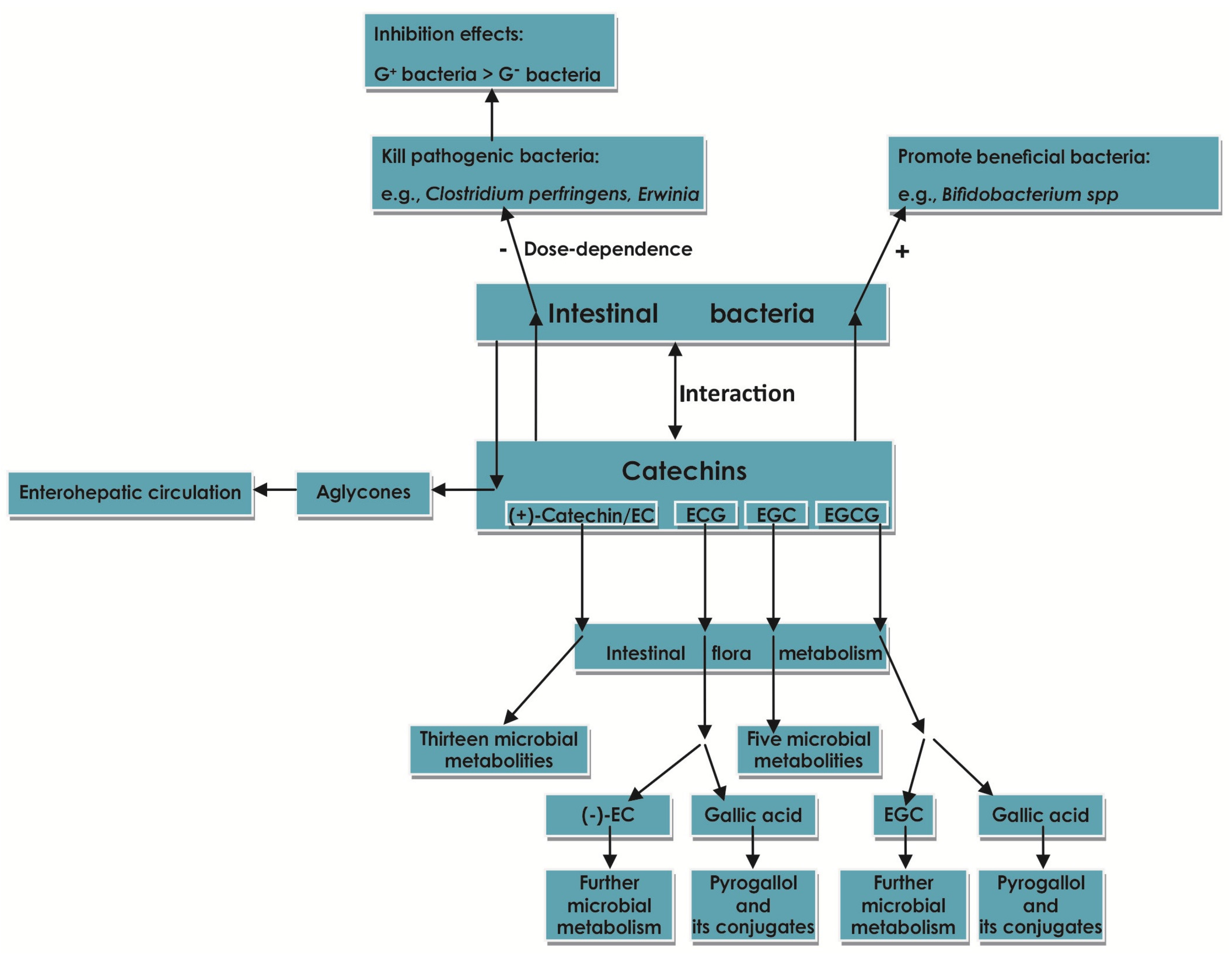
2.7. Effect of Catechins on Intestinal Flora
2.8. Effect of Catechins on Tight Junctions (TJ)
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Sánchez-Fidalgo, S.; Talero, E.; Cárdeno, A.; da Silva, M.A.; Villegas, W.; Souza Brito, A.R.; de La Lastra, C.A. Anti-inflammatory intestinal activity of Abarema cochliacarpos (Gomes) Barneby & Grimes in TNBS colitis model. J. Ethnopharmacol. 2010, 128, 467–475. [Google Scholar] [PubMed]
- Biasi, F.; Astegiano, M.; Maina, M.; Leonarduzzi, G.; Poli, G. Polyphenol Supplementation as a Complementary Medicinal Approach to Treating Inflammatory Bowel Disease. Curr. Med. Chem. 2011, 18, 4851–4865. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.; Bendtzen, K.; Nielsen, O.H. Extraintestinal manifestations of inflammatory bowel disease: Epidemiology, diagnosis, and management. Ann. Med. 2010, 42, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases—Promises, Perspectives, and Pitfalls. Oxidative Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Hold, G.L.; Smith, M.; Grange, C.; Watt, E.R.; EI-Omar, E.M.; Mukhopadhya, I. Role of the gut microbiota in inflammatory bowel disease pathogenesis: What have we learnt in the past 10 years? World J. Gastroenterol. 2014, 20, 1192–1210. [Google Scholar] [CrossRef] [PubMed]
- Yum, H.W.; Zhong, X.; Jin, P.; Na, H.K.; Kim, N.; Lee, H.S.; Surh, Y.J. Oligonol Inhibits Dextran Sulfate Sodium-Induced Colitis and Colonic Adenoma Formation in Mice. Antioxid. Redox Signal. 2013, 19, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, M.; Hasegawa, N. (–)-Epigallocatechin-3-Gallate Reduces Experimental Colon Injury in Rats by Regulating Macrophage and Mast Cell. Phytother. Res. 2010, 24 (Suppl. 1), S120–S122. [Google Scholar] [CrossRef] [PubMed]
- Melgarejo, E.; Medina, M.Á.; Sánchez-Jiménez, F.; Urdiales, J.L. Targeting of histamine producing cells by EGCG: A green dart against inflammation? J. Physiol. Biochem. 2010, 66, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Oteiza, P.I. Dietary flavonoids: Role of (−)-epicatechin and related procyanidins in cell signaling. Free Radic. Biol. Med. 2011, 51, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Buell, M.G.; Berin, M.C. Neutrophil-Independence of the Initiation of Colonic Injury. Comparison of Results from Three Model of Experimental Colitis in the Rat. Dig. Dis. Sci. 1994, 39, 2575–2588. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Zimmerman, B.J.; Specian, R.D.; Grisham, M.B. Role of neutrophils in acetic acid-induced colitis in rats. Inflammation 1991, 15, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Pull, S.L.; Doherty, J.M.; Mills, J.C.; Gordon, J.I.; Stappenbeck, T.S. Activated macrophages are an adaptive element of the colonic epithelial progenitor niche necessary for regenerative responses to injury. Proc. Natl. Acad. Sci. USA 2005, 102, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Dryden, G.W.; Lam, A.; Beatty, K.; Qazzaz, H.H.; Mcclain, C.J. A Pilot Study to Evaluate the Safety and Efficacy of an Oral Dose of (–)-Epigallocatechin-3-Gallate-Rich Polyphenon E in Patients with Mild to Moderate Ulcerative Colitis. Inflamm. Bowel Dis. 2013, 19, 1904–1912. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Saas, P.; Wang, H.; Xu, Y.; Chen, K.; Zhong, J.; Yuan, Y.; Wang, Y.; Sun, Y. Oridonin exerts therapeutic effect by suppressing Th1/Th17 simultaneously in a mouse model of Crohn’s disease. J. Gastroenterol. Hepatol. 2015, 30, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Martin, B.; Veldhoen, M. Development, regulation and functional capacities of Th17 cells. Semin. Immunopathol. 2010, 32, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Seiderer, J.; Elben, I.; Diegelmann, J.; Glas, J.; Stallhofer, J.; Tillack, C.; Pfenning, S.; Jürgens, M.; Schmechel, S.; Konrad, A.; et al. Role of the Novel Th17 Cytokine IL-17F in Inflammatory Bowel Disease (IBD): Upregulated Colonic IL-17F Expression in Active Crohn’s Disease and Analysis of the IL-17F, p.His161arg Polymorphism in IBD. Inflamm. Bowel Dis. 2008, 14, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Rosillo, M.A.; Sanchez-Hidalgo, M.; Cárdeno, A.; Lastra, C.A.D.L. Protective effect of ellagic acid, a natural polyphenolic compound, in a murine model of Crohn’s disease. Biochem. Pharmacol. 2011, 82, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, N.M. Antigen-Specific CD4+ Regulatory T cells in the Intestine. Inflamm. Allergy Drug Targets 2006, 5, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Tyagi, A.K.; Deshmukh-Taskar, P.; Hinojosa, M.; Prasad, S.; Aggarwal, B.B. Downregulation of tumor necrosis factor and other proinflammatory biomarkers by polyphenols. Arch. Biochem. Biophys. 2014, 559, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ben-horin, S.; Chowers, Y. Tailoring anti-TNF therapy in IBD: Drug levels and disease activity. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Lichtenstein, L.; Assa, A.; Mazor, Y.; Weiss, B.; Levine, A.; Ron, Y.; Kopylov, U.; Bujanover, Y.; Rosenbach, Y.; et al. Levels of Drug and Antidrug Antibodies Are Associated With Outcome of Interventions After Loss of Response to Infliximab or Adalimumab. Clin. Gastroenterol. Hepatol. 2015, 13, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Howell, H.R. Ulcerative colitis: Achieving and maintaining remission. US Pharm. 2008, 33, 30–38. [Google Scholar]
- Faubion, W.A., Jr.; Loftus, E.V., Jr.; Harmsen, W.S.; Zinsmeister, A.R.; Sandborn, W.J. The Natural History of Corticosteroid Therapy for Inflammatory Bowel Disease: A Population-Based Study. Gastroenterology 2001, 121, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Rahimi, R.; Abdollahi, M. The Role of Dietary Polyphenols in the Management of Inflammatory Bowel Disease. Curr. Pharm. Biotechnol. 2015, 16, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Koláček, M.; Muchová, J.; Dvořáková, M.; Paduchová, Z.; Žitňanová, I.; Čierna, I.; Országhová, Z.; Székyová, D.; Jajcaiová-Zedníčková, N.; Kovács, L.; et al. Effect of natural polyphenols (Pycnogenol) on oxidative stress markers in children suffering from Crohn’s disease - a pilot study. Free Radic. Res. 2013, 47, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Iida, T.; Takeuchi, K.; Watanabe, F.; Maruyama, Y.; Andoh, A.; Tsujikawa, T.; Fujiyama, Y.; Mitsuyama, K.; Sata, M.; et al. Curcumin Maintenance Therapy for Ulcerative Colitis: Randomized, Multicenter, Double-Blind, Placebo-Controlled Trial. Clin. Gastroenterol. Hepatol. 2006, 4, 1502–1506. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.H.; Choi, K.C.; Cho, S.W.; Cho, H.K.; Lee, K.D.; Lee, J.C. Catechin-7-O-β-d-glucopyranoside isolated from the seed of Phaseolus calcaratus Roxburgh ameliorates experimental colitis in rats. Int. Immunopharmacol. 2015, 29, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fidalgo, S.; Da Silva, S.M.; Cárdeno, A.; Aparicio-Soto, M.; Salvador, M.J.; Frankland Sawaya, A.C.; Souza-Brito, A.R.M.; Alarcón de la Lastra, C. Abarema cochliacarpos reduces LPS-induced inflammatory response in murine peritoneal macrophages regulating ROS-MAPK signal pathway. J. Ethnopharmacol. 2013, 149, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.C.; Seito, L.N.; Di, S.L.; Akiko, H.C.; Ch, P. Epicatechin Used in the Treatment of Intestinal Inflammatory Disease: An Analysis by Experimental Models. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Ye, M.; Wang, Y.; Ni, Y.; Su, M.; Huang, H.; Qiu, M.; Zhao, A.; Zheng, X.; Chen, T.; et al. Characterization of Pu-erh Tea Using Chemical and Metabolic Profiling Approaches. J. Agric. Food Chem. 2009, 57, 3046–3054. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xiao, R.; Hu, X.; Xu, K.; Hou, Y.; Zhong, Y.; Meng, J.; Fan, B.; Liu, L. Comparative Safety Evaluation of Chinese Pu-erh Green Tea Extract and Pu-erh Black Tea Extract in Wistar Rats. J. Agric. Food Chem. 2010, 58, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Danila, A.M.; Kotani, A.; Hakamata, H.; Kusu, F. Determination of Rutin, Catechin, Epicatechin, and Epicatechin Gallate in Buckwheat Fagopyrum Esculentum Moench by Micro-High-Performance Liquid Chromatography with Electrochemical Detection. J. Agric. Food Chem. 2007, 55, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Satoko, A.; Atsushi, N.; Mari, M.; Mariko, U.; Akira, M. Effects of anthocyanin-rich tea “Sunrouge” on dextran sodium sulfate-induced colitis in mice. BioFactors 2012, 38, 226–233. [Google Scholar]
- Stevens, J.F.; Maier, C.S. The chemistry of gut microbial metabolism of polyphenols. Phytochem. Rev. 2016, 15, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Fathima, A.; Rao, J.R. Selective toxicity of Catechin—A natural flavonoid towards bacteria. Appl. Microbiol. Biotechnol. 2016, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.F.; Fisher, L.J.; Hara, Y.; Harris, T.; Mak, W.B.; Melton, L.D.; Packer, J.E. Green tea catechins partially protect DNA from ·OH radical-induced strand breaks and base damage through fast chemical repair of DNA radicals. Carcinogenesis 2001, 22, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Pegg, R.B.; Bautista, D.A. Antibacterial activity of green tea polyphenols against Escherichia coli K12. Mol. Nutr. Food Res. 2000, 44, 60–62. [Google Scholar] [PubMed]
- Muzolf-Panek, M.; Gliszczyńska-Swigło, A.; de, H.L.; Aarts, J.M.; Szymusiak, H.; Vervoort, J.M.; Tyrakowska, B.; Rietjens, I.M. Role of Catechin Quinones in the Induction of EpRE-Mediated Gene Expression. Chem. Res. Toxicol. 2008, 21, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Brückner, M.; Westphal, S.; Domschke, W.; Kucharzik, T.; Lügering, A. Green tea polyphenol epigallocatechin-3-gallate shows therapeutic antioxidative effects in a murine model of colitis. J. Crohns. Colitis 2012, 6, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Chiou, Y.S.; Wang, Y.J.; Ho, C.T.; Lin, J.K. Multistage carcinogenesis process as molecular targets in cancer chemoprevention by epicatechin-3-gallate. Food Funct. 2011, 2, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Lee, H.J. The roles of polyphenols in cancer chemoprevention. BioFactors 2016, 26, 105–121. [Google Scholar] [CrossRef]
- Silva, M.S.D.; Almeida, A.C.A.D.; Faria, F.M.D.; Luiz-Ferreira, A.; Silva, M.A.D.; Vilegas, W.; Pellizzon, C.H.; Brito, C.H.P. Abarema cochliacarpos: Gastroprotective and Ulcer-Healing Activities. J. Ethnopharmacol. 2010, 132, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Scalia, S.; Marchetti, N.; Bianchi, A. Comparative Evaluation of Different Co-Antioxidants on the Photochemical- and Functional-Stability of Epigallocatechin-3-gallate in Topical Creams Exposed to Simulated Sunlight. Molecules 2013, 18, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Hong, J.; Yang, G.Y.; Liao, J.; Yang, C.S. Inhibition of carcinogenesis by polyphenols: Evidence from laboratory investigations. Am. J. Clin. Nutr. 2005, 81, 284S–291S. [Google Scholar] [PubMed]
- Yang, C.S.; Maliakal, P.; Meng, X. Inhibition of Carcinogenesis by Tea. Annu. Rev. Pharmacol. Toxicol. 2002, 389, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, X.; Lu, G.; Picinich, S.C. Cancer prevention by tea: Animal studies, molecular mechanisms and human relevance. Nat. Rev. Cancer 2009, 9, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Hao, X.; Simi, B.; Ju, J.; Jiang, H.; Reddy, B.S.; Yang, C.S. Green tea polyphenols inhibit colorectal aberrant crypt foci (ACF) formation and prevent oncogenic changes in dysplastic ACF in azoxymethane-treated F344 rats. Carcinogenesis 2008, 29, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Liu, A.B.; Li, G.; Yang, Z.; Sun, Y.; Yang, C.S.; Ju, J. Deleterious Effects of High Concentrations of (–)-Epigallocatechin-3-Gallate and Atorvastatin in Mice With Colon Inflammation. Nutr. Cancer 2012, 64, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.D.; Ellis, L.M. Inhibition of tumour invasion and angiogenesis by epigallocatechin gallate (EGCG), a major component of green tea. Int. J. Exp. Pathol. 2001, 82, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Jankun, J.; Selman, S.H.; Swiercz, R.; Skrzypczak-Jankun, E. Why drinking green tea could prevent cancer. Nature 1997, 387, 561. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Lu, T.H.; Su, C.C.; Lay, I.S.; Lin, H.Y.; Fang, K.M.; Ho, T.J.; Chen, K.L.; Su, Y.C.; Chiang, W.C.; et al. Lotus Leaf (Nelumbo nucifera) and its Active Constituents Prevent Inflammatory Responses in Macrophages via JNK/NF-κB Signaling Pathway. Am. J. Chin. Med. 2014, 42, 869–889. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Adachi, S.; Hata, K.; Hirose, Y.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. (–)-Epigallocatechin Gallate Suppresses Azoxymethane-Induced Colonic Premalignant Lesions in Male C57BL/KsJ-db/db Mice. Cancer Prev. Res. 2008, 1, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Rodríguezramiro, I.; Martín, M.Á.; Ramos, S.; Bravo, L.; Goya, L. Comparative effects of dietary flavanols on antioxidant defences and their response to oxidant-induced stress on Caco2 cells. Eur. J. Nutr. 2011, 50, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y. Influence of Tea Catechins on the Digestive Tract. J. Cell. Biochem. Suppl. 1997, 52, 8. [Google Scholar]
- Kim, M.; Murakami, A.; Miyamoto, S.; Tanaka, T.; Ohigashi, H. The modifying effects of green tea polyphenols on acute colitis and inflammation-associated colon carcinogenesis in male ICR mice. BioFactors 2010, 36, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.S.; Huang, Q.; Ho, C.T.; Wang, Y.J.; Pan, M.H. Directly interact with Keap1 and LPS is involved in the anti-inflammatory mechanisms of (–)-epicatechin-3-gallate in LPS-induced macrophages and endotoxemia. Free Radic. Biol. Med. 2016, 94, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the Golden Nutraceutical: Multitargeting for Multiple Chronic Diseases. Br. J. Pharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.S.; Lim, I.H.; Yuk, D.Y.; Chung, K.H.; Park, J.B.; Yoo, H.S.; Yun, Y.P. Antithrombotic Activities of Green Tea Catechins and (–)-Epigallocatechin Gallate. Thromb. Res. 1999, 96, 229–237. [Google Scholar] [CrossRef]
- Marinovic, M.P.; Morandi, A.C.; Otton, R. Green tea catechins alone or in combination alter functional parameters of human neutrophils via suppressing the activation of TLR-4/NFκB p65 signal pathway. Toxicol. In Vitro 2015, 29, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Dryden, G.W.; Song, M.; Mcclain, C. Polyphenols and gastrointestinal diseases. Curr. Opin. Gastroenterol. 2006, 22, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Tsuno, N.H.; Kitayama, J.; Okaji, Y.; Yazawa, K.; Asakage, M.; Sasaki, S.; Watanabe, T.; Takahashi, K.; Nagawa, H. Epigallocatechin gallate induces apoptosis of monocytes. J. Allergy Clin. Immunol. 2005, 115, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T. Condensed catechins and their potential health-benefits. Eur. J. Pharmacol. 2015, 765, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.M.; Mattila, I.; Seppänen-Laakso, T.; Miettinen, J.; Oksman-Caldentey, K.M.; Orešič, M. Microbial metabolism of catechin stereoisomers by human faecal microbiota: Comparison of targeted analysis and a non-targeted metabolomics method. Phytochem. Lett. 2008, 1, 18–22. [Google Scholar] [CrossRef]
- Clifford, M.N.; Van der Hooft, J.J.; Crozier, A. Human studies on the absorption, distribution, metabolism, and excretion of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1619S–1630S. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.Y. Metabolism of Green Tea Catechins: An Overview. Curr. Drug Metab. 2006, 7, 755–809. [Google Scholar] [CrossRef] [PubMed]
- Stalmach, A.; Mullen, W.; Steiling, H.; Williamson, G.; Lean, M.E.; Crozier, A. Absorption, metabolism, and excretion of green tea flavan-3-ols in humans with an ileostomy. Mol. Nutr. Food Res. 2010, 54, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zheng, S.; Li, L.; Jiang, H. Metabolism of Flavonoids in Human: A Comprehensive Review. Curr. Drug Metab. 2014, 15, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Dai, Y.; Xun, L.; Hu, M. Enteric Disposition and Recycling of Flavonoids and Ginkgo Flavonoids. J. Altern. Complement. Med. 2003, 9, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Mcdougall, G.J.; Alegría, A.; Alminger, M.; Arrigoni, E.; Aura, A.M.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the gap-deficits in our knowledge of aspects impacting the bioavailability of phytochemicals and their metabolites—A position paper focusing on carotenoids and polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef] [PubMed]
- Del, R.D.; Calani, L.; Cordero, C.; Salvatore, S.; Pellegrini, N.; Brighenti, F. Bioavailability and catabolism of green tea flavan-3-ols in humans. Nutrition 2010, 26, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Momma, T.Y.; Heiss, C.; Kwik-Uribe, C.; Schroeter, H.; Keen, C.L. The sterochemical configuration of flavanols influences the level and metabolism of flavanols in humans and their biological activity in vivo. Free Radic. Biol. Med. 2011, 50, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.D.; Rodriguezmateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [PubMed]
- Neilson, A.P.; Sapper, T.N.; Janle, E.M.; Rudolph, R.; Matusheski, N.V.; Ferruzzi, M.G. Chocolate Matrix Factors Modulate the Pharmacokinetic Behavior of Cocoa Flavan-3-OL Phase-II Metabolites Following Oral Consumption by Sprague-Dawley Rats. J. Agric. Food Chem. 2010, 58, 6685–6691. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.S.; Ma, J.L.; Sang, S.; Ho, C.T.; Wang, Y.J.; Pan, M.H. Peracetylated (–)-Epigallocatechin-3-gallate (AcEGCG) Potently Suppresses Dextran Sulfate Sodium-Induced Colitis and Colon Tumorigenesis in Mice. J. Agric. Food Chem. 2012, 60, 3441–3451. [Google Scholar] [CrossRef] [PubMed]
- Alzoghaibi, M.A. Concepts of oxidative stress and antioxidant defense in Crohn’s disease. World J. Gastroenterol. 2013, 19, 6540–6547. [Google Scholar] [CrossRef] [PubMed]
- Kruidenier, L.; Kuiper, I.; Lamers, C.B.; Verspaget, H.W. Intestinal oxidative damage in inflammatory bowel disease: semi-quantification, localization, and association with mucosal antioxidants. J. Pathol. 2003, 201, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Maiorino, M.; Ursini, F. Signaling Functions of Reactive Oxygen Species. Biochemistry 2010, 49, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Soobrattee, M.A.; Bahorun, T.; Aruoma, O.I. Chemopreventive actions of polyphenolic compounds in cancer. BioFactors 2006, 27, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Najafzadeh, M.; Reynolds, P.D.; Baumgartner, A.; Anderson, D. Flavonoids inhibit the genotoxicity of hydrogen peroxide (H2O2) and of the food mutagen 2-amino-3-methylimadazo[4,5-f]-quinoline (IQ) in lymphocytes from patients with inflammatory bowel disease (IBD). Mutagenesis 2009, 24, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.B.; Sousa, C.; Costa, A.; Andrade, P.B.; Valentão, P. Glutathione and the Antioxidant Potential of Binary Mixtures with Flavonoids: Synergisms and Antagonisms. Molecules 2013, 18, 8858–8872. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Flavonoids as Antioxidants: Determination of Radical-Scavenging Efficiencies. Methods Enzymol. 1990, 186, 343–355. [Google Scholar] [PubMed]
- Kim, C.Y.; Lee, C.; Park, G.H.; Jang, J.H. Neuroprotective Effect of Epigallocatechin-3-gallate against β-Amyloid-induced Oxidative and Nitrosative Cell Death via Augmentation of Antioxidant Defense Capacity. Arch. Pharm. Res. 2009, 32, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.H.; Xu, J.; Hu, F.L.; Wan, X.C.; Deng, S.X.; Barasch, J. EGCG inhibit chemical reactivity of iron through forming an Ngal-EGCG-iron complex. BioMetals 2013, 26, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Raza, H.; John, A. In vitro protection of reactive oxygen species-induced degradation of lipids, proteins and 2-deoxyribose by tea catechins. Food Chem. Toxicol. 2007, 45, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhao, B.; Li, M.; Shen, S.; Xin, W. Studies on protective mechanisms of four components of green tea polyphenols against lipid peroxidation in synaptosomes. Biochim. Biophys. Acta 1996, 1304, 210–222. [Google Scholar] [CrossRef]
- Meng, Q.; Velalar, C.N.; Ruan, R. Effects of epigallocatechin-3-gallate on mitochondrial integrity and antioxidative enzyme activity in the aging process of human fibroblast. Free Radic. Biol. Med. 2008, 44, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Esworthy, R.S.; Aranda, R.; Martín, M.G.; Doroshow, J.H.; Binder, S.W.; Chu, F.F. Mice with combined disruption of Gpx1 and Gpx2 genes have colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, 848–855. [Google Scholar]
- Martín, M.Á.; Serrano, A.B.; Ramos, S.; Pulido, M.I.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzyme activity via the ERK1/2 pathway to protect against oxidative stress-induced apoptosis in HepG2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Abboud, P.A.; Hake, P.W.; Burroughs, T.J.; Odoms, K.; O’Connor, M.; Mangeshkar, P.; Wong, H.R.; Zingarelli, B. Therapeutic effect of epigallocatechin-3-gallate in a mouse model of colitis. Eur. J. Pharmacol. 2008, 579, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Seegert, D.; Rosenstiel, P.; Pfahler, H.; Pfefferkorn, P.; Nikolaus, S.; Schreiber, S. Increased expression of IL-16 in inflammatory bowel disease. Gut 2001, 48, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shao, F.; Yang, Y.; Gu, L.; Zheng, W.; Wu, X.; Gu, Y.; Shu, Y.; Sun, Y.; Xu, Q. Epigallocatechin-3-gallate sensitizes IFN-γ-stimulated CD4+ T cells to apoptosis via alternative activation of STAT1. Int. Immunopharmacol. 2014, 23, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Kang, B.S.; Lee, B.J.; Che, J.H.; Li, G.X.; Trosko, J.E.; Lee, Y.S. Preventive effect of epicatechin and ginsenoside Rb2 on the inhibition of gap junctional intercellular communication by TPA and H2O2. Cancer Lett. 2002, 152, 97–106. [Google Scholar] [CrossRef]
- Trosko, J.E.; Chang, C.C.; Upham, B.; Wilson, M. Epigenetic toxicology as toxicant-induced changes in intracellular signalling leading to altered gap junctional intercellular communication. Toxicol. Lett. 1998, 95, 4–5. [Google Scholar] [CrossRef]
- Kang, N.J.; Lee, K.M.; Kim, J.H.; Bo, K.L.; Kwon, J.Y.; Lee, K.W.; Lee, H.J. Inhibition of Gap Junctional Intercellular Communication by the Green Tea Polyphenol (–)-Epigallocatechin Gallate in Normal Rat Liver Epithelial Cells. J. Agric. Food Chem. 2008, 56, 10422–10427. [Google Scholar] [CrossRef] [PubMed]
- Sai, K.; Kanno, J.; Hasegawa, R.; Hasegawa, R.; Trosko, J.E.; Inoue, T. Prevention of the down-regulation of gap junctional intercellular communication by green tea in the liver of mice fed pentachlorophenol. Carcinogenesis 2000, 21, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Ale-Agha, N.; Stahl, W.; Sies, H. (–)-Epicatechin effects in rat liver epithelial cells: Stimulation of gap junctional communication and counteraction of its loss due to the tumor promoter 12-O-tetradecanoylphorbol-13-acetate. Biochem Pharmacol. 2002, 63, 2145. [Google Scholar] [CrossRef]
- Sigler, K.; Ruch, R.J. Enhancement of gap junctional intercellular communication in tumor promoter-treated cells by components of green tea. Cancer Lett. 1993, 69, 15–19. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, K.W.; Lee, H.J. Abies nephrolepis leaf phenolics prevent the inhibition of gap junction intercellular communication by hydrogen peroxide in rat liver epithelial cells. BioFactors 2004, 21, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, L.; Xu, S.; Qiu, F.; Fan, Y.; Fu, G. Down-regulation of connexin43 gap junction by serum deprivation in human endothelial cells was improved by (–)-epigallocatechin gallate via ERK MAP kinase pathway. Biochem. Biophys. Res. Commun. 2011, 404, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhao, Y.; Fan, Y.; Wang, M.; Xu, S.; Fu, G. Epigallocatechin-3 Gallate, a Green Tea Catechin, Attenuated the Downregulation of the Cardiac Gap Junction Induced by High Glucose in Neonatal Rat Cardiomyocytes. Cell. Physiol. Biochem. 2010, 26, 403–412. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Ghosh, S. Signal transduction through NF-κB. Immunol. Today 1998, 19, 80–88. [Google Scholar] [CrossRef]
- Ukil, A.; Maity, S.; Das, P.K. Protection from experimental colitis by theaflavin-3,3′-digallate correlates with inhibition of IKK and NF-κB activation. Br. J. Pharmacol. 2006, 149, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Andújar, I.; Recio, M.C.; Giner, R.M.; Cienfuegos-Jovellanos, E.; Laghi, S.; Muguerza, B.; Ríos, J.L. Inhibition of Ulcerative Colitis in Mice after Oral Administration of a Polyphenol-Enriched Cocoa Extract Is Mediated by the Inhibition of STAT1 and STAT3 Phosphorylation in Colon Cells. J. Agric. Food Chem. 2011, 59, 6474–6483. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Triebel, S.; Anke, T.; Richling, E.; Erkel, G. Influence of apple polyphenols on inflammatory gene expression. Mol. Nutr. Food Res. 2009, 53, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Khajah, M.A.; Fateel, M.M.; Ananthalakshmi, K.V.; Luqmani, Y.A. Anti-Inflammatory Action of Angiotensin 1-7 in Experimental Colitis. PLoS ONE 2016, 11, e0150861. [Google Scholar] [CrossRef] [PubMed]
- Krens, S.F.G.; Spaink, H.P.; Snaar-Jagalska, B.E. Functions of the MAPK family in vertebrate development. FEBS Lett. 2006, 580, 4984–4990. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Wu, C.H.; Yen, G.C. Effects of flavonoids on the expression of the pro-inflammatory response in human monocytes induced by ligation of the receptor for AGEs. Mol. Nutr. Food Res. 2006, 50, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jin, H.; Shim, H.E.; Kim, H.N.; Ha, H.; Lee, Z.H. Epigallocatechin-3-gallate Inhibits Osteoclastogenesis by Down-Regulating c-Fos Expression and Suppressing the Nuclear Factor-κB Signal. Mol. Pharmacol. 2010, 77, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Danesi, F.; Philpott, M.; Huebner, C.; Bordoni, A.; Ferguson, L.R. Food-derived bioactives as potential regulators of the IL-12/IL-23 pathway implicated in inflammatory bowel diseases. Mutat. Res. 2010, 690, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Choi, S.R.; Lee, C.M.; Kim, G.Y.; Lee, H.J.; Park, Y.M. Epigallocatechin-3-gallate Suppresses Galactose-α1,4-galactose-β1,4-glucose Ceramide Expression in TNF-α Stimulated Human Intestinal Epithelial Cells Through Inhibition of MAPKs and NF-κB. J. Korean Med. Sci. 2005, 20, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N. Nrf2: Friend or foe for chemoprevention? Carcinogenesis 2010, 31, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Bahia, P.K.; Rattray, M.; Williams, R.J. Dietary flavonoid (–)-epicatechin stimulates phosphatidylinositol 3-kinase-dependent antioxidant response element activity and up-regulates glutathione in cortical astrocytes. J. Neurochem. 2008, 106, 2194–2204. [Google Scholar] [PubMed]
- Wakabayashi, N.; Slocum, S.L.; Skoko, J.J.; Shin, S.; Kensler, T.W. When NRF2 Talks, who’s Listening? Antioxid. Redox Signal. 2010, 13, 1649–1663. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Qu, J.; Shen, X. NF-κB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim. Biophys. Acta 2008, 1783, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.; Rosenstiel, P.; Hampe, J.; Nikolaus, S.; Groessner, B.; Schottelius, A.; Kühbacher, T.; Hämling, J.J.; Fölsch, U.R.; Seegert, D. Activation of signal transducer and activator of transcription (STAT) 1 in human chronic inflammatory bowel disease. Gut 2002, 51, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K. Role of STAT3 in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, S110–S114. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Mohebali, N.; Esa, N.M.; Looi, C.Y.; Ismail, S.; Saadatdoust, Z. Gallic acid suppresses inflammation in dextran sodium sulfate-induced colitis in mice: Possible mechanisms. Int. Immunopharmacol. 2015, 28, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Xie, J.; Huang, J.; Chen, L.; Gao, L.; Ou, S.; Wang, Y.; Peng, X. Plant Polyphenols Alter a Pathway of Energy Metabolism by Inhibiting Fecal Bacteroidetes and Firmicutes in vitro. Food Funct. 2016, 7, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.; Khazanehei, H.; Jones, P.J.; Khafipour, E. Interactions between Obesity Status and Dietary Intake of Monounsaturated and Polyunsaturated Oils on Human Gut Microbiome Profiles in the Canola Oil Multicenter Intervention Trial (COMIT). Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Sanderson, J.D.; Churcher, C.; Parkes, G.C.; Hudspith, B.N.; Rayment, N.; Brostoff, J.; Parkhill, J.; Dougan, G.; Petrovska, L. High-throughput clone library analysis of the mucosa-associated microbiota reveals dysbiosis and differences between inflamed and non-inflamed regions of the intestine in inflammatory bowel disease. BMC Microbiol. 2011, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.N.; St. Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals—Just antioxidants or much more? Cell. Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Archana, S.; Abraham, J. Comparative analysis of antimicrobial activity of leaf extracts from fresh green tea and black tea on pathogens. J. Appl. Pharm. Sci. 2011, 1, 149–152. [Google Scholar]
- Rastmanesh, R. High polyphenol, low probiotic diet for weight loss because of intestinal microbiota interaction. Chem. Biol. Interact. 2011, 189, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Trevanich, S.; Tsujimoto, Y.; Miki, T.; Miyabe, S.; Sugiyama, K.; Izawa, S.; Kimura, A. Evaluation of catechin and its derivatives as antioxidant: Recovery of growth arrest of Escherichia coli, under oxidative conditions. J. Sci. Food Agric. 1996, 71, 297–300. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Kutschera, M.; Engst, W.; Blaut, M.; Braune, A. Isolation of catechin-converting human intestinal bacteria. J. Appl. Microbiol. 2011, 111, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Beaud, D.; Tailliez, P.; Anbamondoloni, J. Genetic characterization of the β-glucuronidase enzyme from a human intestinal bacterium, Ruminococcus gnavus. Microbiology 2005, 151, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Girón, A.; Ibáñez, C.; Cifuentes, A.; Simó, C.; Muñoz-González, I.; Martín-Álvarez, P.J.; Bartolomé, B.; Moreno-Arribas, M.V. Faecal metabolomic fingerprint after moderate consumption of red wine by healthy subjects. J. Proteome Res. 2015, 14, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.S.; Lee, K.C.; Park, I.S.; Kim, K.K.; Ahn, J.S.; Benno, Y.; Hattori, M.; Lee, J.S. Gordonibacter faecihominis sp. nov., isolated from human faeces. Antonie Leeuwenhoek. 2014, 106, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.S.; Hattori, M. Isolation and Characterization of a Human Intestinal Bacterium Eggerthella sp. CAT-1 Capable of Cleaving the C-Ring of (+)-Catechin and (–)-Epicatechin, Followed by p-Dehydroxylation of the B-Ring. Biol. Pharm. Bull. 2012, 35, 2252–2256. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Meselhy, M.R.; Li, Y.; Nakamura, N.; Min, B.S.; Qin, G.W.; Hattori, M. The Heterocyclic Ring Fission and Dehydroxylation of Catechins and Related Compounds by Eubacterium sp. Strain SDG-2, a Human Intestinal Bacterium. Chem. Pharm. Bull. 2002, 33, 1640–1643. [Google Scholar] [CrossRef]
- Silverman, R.B. The Organic Chemistry of Enzyme Catalyzed Reactions. Richard B. Silverman. The Organic Chemistry of Enzyme-Catalyzed Reactions; Academic Press: New York, NY, USA, 2002; Volume 172. [Google Scholar]
- Du, L.; Wang, Z.; Zhao, Y.; Huang, J.; Pang, H.; Wei, Y.; Lin, L.; Huang, R. A β-glucosidase from Novosphingobium sp. GX9 with high catalytic efficiency toward isoflavonoid glycoside hydrolysis and (+)-catechin transglycosylation. Appl. Microbiol. Biotechnol. 2014, 98, 7069–7079. [Google Scholar] [CrossRef] [PubMed]
- Steed, E.; Balda, M.S.; Matter, K. Dynamics and functions of tight junctions. Trends Cell Biol. 2010, 20, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Contreras, T.C.; Ricciardi, E.; Cremonini, E.; Oteiza, P.I. (–)-Epicatechin in the prevention of tumor necrosis alpha-induced loss of Caco-2 cell barrier integrity. Arch. Biochem. Biophys. 2015, 573, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.Y.; Söderholm, J.D. Importance of Disrupted Intestinal Barrier in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2011, 17, 362–381. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Pozo, C.; Morales, P.; Gotteland, M. Polyphenols Protect the Epithelial Barrier Function of Caco-2 Cells Exposed to Indomethacin through the Modulation of Occludin and Zonula Occludens-1 Expression. J. Agric. Food Chem. 2013, 61, 5291–5297. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, F.-Y.; Sang, L.-X.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. https://doi.org/10.3390/molecules22030484
Fan F-Y, Sang L-X, Jiang M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules. 2017; 22(3):484. https://doi.org/10.3390/molecules22030484
Chicago/Turabian StyleFan, Fei-Yan, Li-Xuan Sang, and Min Jiang. 2017. "Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease" Molecules 22, no. 3: 484. https://doi.org/10.3390/molecules22030484
APA StyleFan, F.-Y., Sang, L.-X., & Jiang, M. (2017). Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules, 22(3), 484. https://doi.org/10.3390/molecules22030484