
Growth Inhibition of Sulfate-Reducing Bacteria in Produced Water from the Petroleum Industry Using Essential Oils

 , ,
, ,
Abstract
:
1. Introduction
2. Results
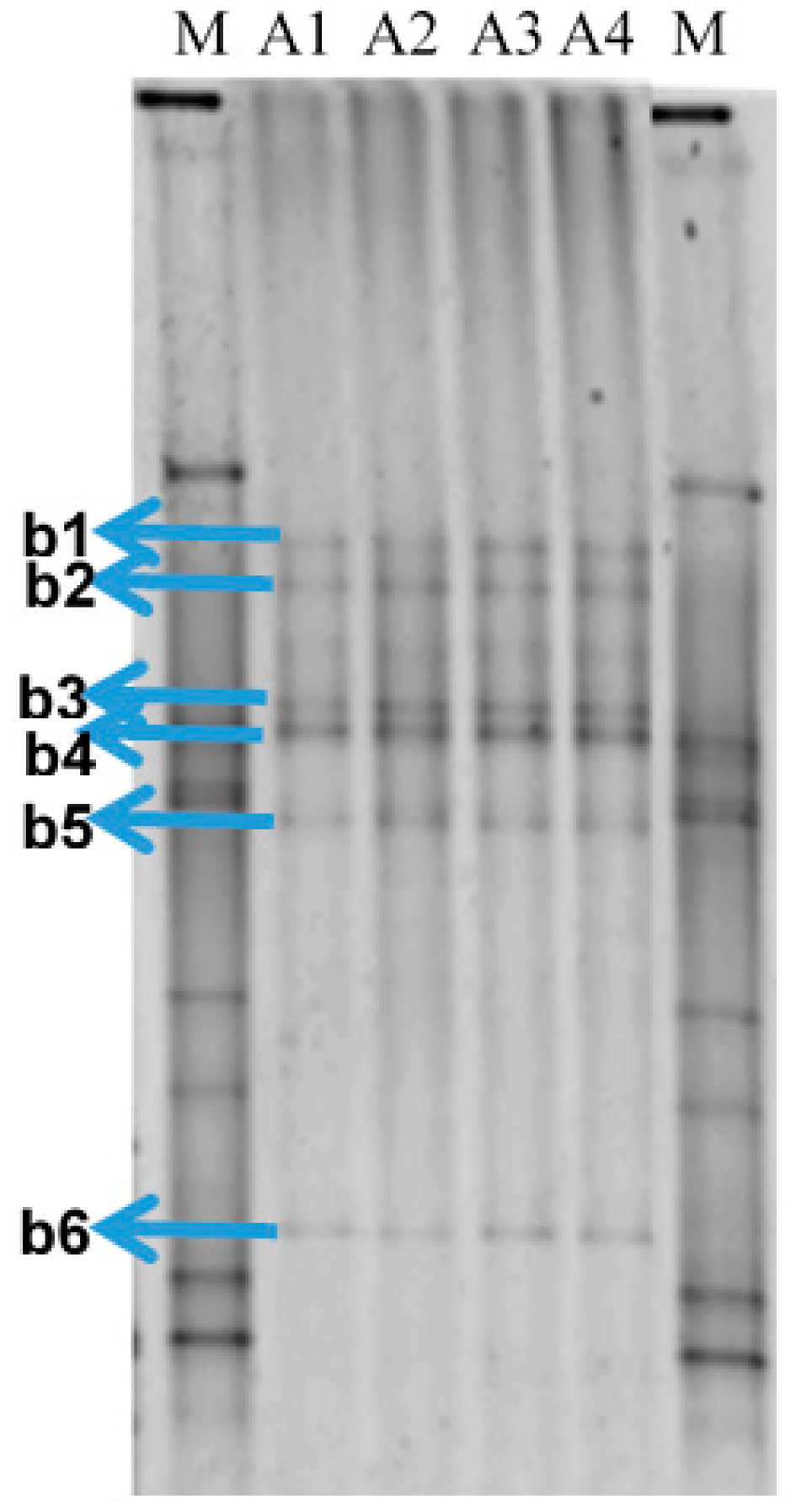
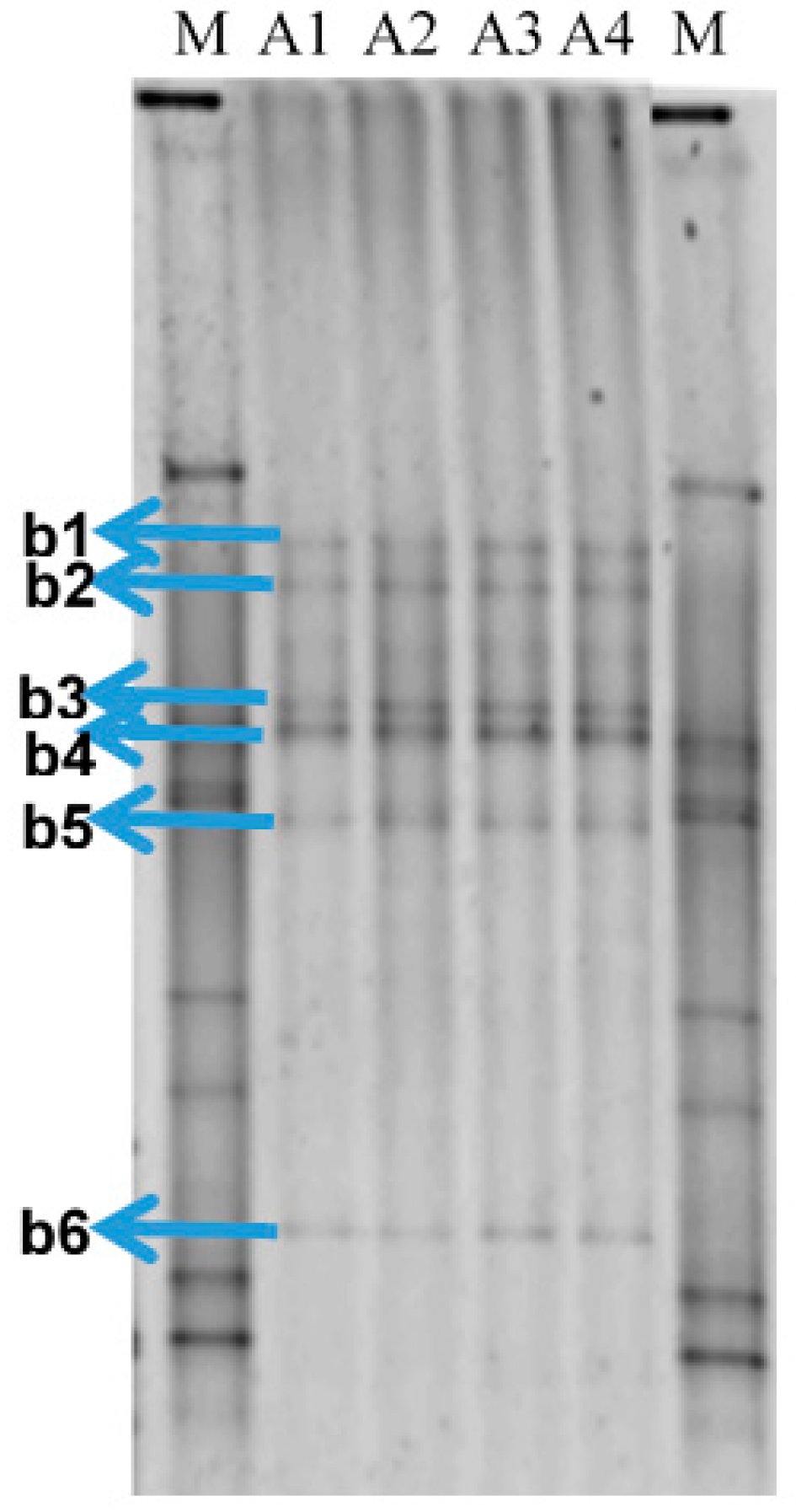
2.1. Bacterial Community in the Produced Water Samples
2.2. Essential Oils and Major Components Against SRB
3. Discussion
4. Materials and Methods
4.1. Produced Water
4.2. Sulfate-Reducing Bacteria
4.3. EO and Major Components
4.4. DNA Extraction
4.5. PCR Amplification of Bacterial rrs and dsrA Genes
4.6. Denaturing Gradient Gel Electrophoresis (DGGE) and Statistical Analyses
4.7. Sequencing of DGGE Bands and dsrA Clone Libraries
4.8. Enumeration of Sulfate-Reducing Bacteria
4.9. Determination of Minimum Inhibitory Concentration (MIC) of EO and Major Components
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fakhru’l-Razi, A.; Pendashteh, A.; Abdullah, L.C.; Biak, D.R.A.; Madaeni, S.S.; Abidin, Z.Z. Review of technologies for oil and gas produced water treatment. J. Hazard. Mater. 2009, 170, 530–551. [Google Scholar] [PubMed]
- Muggeridge, A.; Cockin, A.; Webb, K.; Frampton, H.; Collins, I.; Moulds, T.; Salino, P. Recovery rates, enhanced oil recovery and technological limits. Philos. Trans. R. Soc. A 2014, 372, 20120320. [Google Scholar]
- Fontenot, B.E.; Hunt, L.R.; Hildenbrand, Z.L.; Carlton, D.D.; Oka, H.; Walton, J.L.; Hopkins, D.; Osorio, A.; Bjorndal, B.; et al. An evaluation of water quality in private drinking water wells near natural gas extraction sites in the Barnett shale formation. Environ. Sci. Technol. 2013, 47, 10032–10040. [Google Scholar] [CrossRef] [PubMed]
- Harkness, J.S.; Dwyer, G.S.; Warner, N.R.; Parker, K.M.; Mitch, W.A.; Vengosh, A. Iodide, bromide, and ammonium in hydraulic fracturing and oil and gas wastewaters: Environmental implications. Environ. Sci. Technol. 2015, 49, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Veil, J.; Puder, M.G.; Elcock, D.; Redweik, R.J.J. A White Paper Describing Produced Water from Production of Crude Oil, Natural Gas and Coal Bed Methane. Available online: http://www.netl.doe.gov/publications/oil pubs/prodwaterpaper.pdf (accessed on 26 January 2017).
- Arthur, J.D.; Langhus, B.G.; Patel, C. Technical Summary of Oil and Gas Produced Water Treatment Technologies. Available online: http://www.rrc.state.tx.us/ commissioners/williams/environment/produced water treatment Tech.pdf (accessed on 26 January 2017).
- Igunnu, E.T.; Chen, G.Z. Produced water treatment technologies. Int. J. Low-Carbon Technol. 2014, 9, 157–177. [Google Scholar]
- Singer, M.E. Microbial biosurfactants. In Microbes and Oil Recovery; Zajic, J.E., Donaldson, E.C., Eds.; Bioresources Publications: El Paso, TX, USA, 1985; pp. 19–38. [Google Scholar]
- Popoola, L.T.; Grema, A.S.; Latinwo, G.K.; Gutti, B.; Balogun, A.S. Corrosion problems during oil and gas production and its mitigation. Int. J. Ind. Chem. 2013, 4, 35. [Google Scholar]
- Barton, L.L.; Fauque, G.D. Biochemistry, physiology and biotechnology of sulfate-reducing bacteria. Adv. Appl. Microbiol. 2009, 68, 41–98. [Google Scholar] [PubMed]
- Gevertz, D.; Telang, A.J.; Voordouw, G.; Jenneman, G.E. Isolation and characterization of strains CVO and FWKO B, two novel nitrate-reducing, sulfide-oxidizing bacteria isolated from oil field brine. Appl. Environ. Microbiol. 2000, 66, 2491–2501. [Google Scholar] [CrossRef] [PubMed]
- Hubert, C.; Nemati, M.; Jenneman, G.; Voordouw, G. Corrosion risk associated with microbial souring control using nitrate or nitrite. Appl. Microbiol. Biotechnol. 2005, 68, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Videla, H.A. Prevention and control of biocorrosion. Int. Biodeterior. Biodegrad. 2002, 49, 259–270. [Google Scholar]
- Pichtel, J. Oil and gas production wastewater: Soil contamination and pollution prevention. Appl. Environ. Soil Sci. 2016, 2016, 24. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Nemati, M.; Jenneman, G.E.; Voordouw, G. Mechanistic study of microbial control of hydrogen sulfide production in oil reservoirs. Biotechnol. Bioeng. 2001, 74, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Koudou, J.; Abena, A.A.; Ngaissona, P.; Bessière, J.M. Chemical composition and pharmacological activity of essential oil of Canarium schweinfurthii. Fitoterapia 2005, 76, 700–703. [Google Scholar] [CrossRef] [PubMed]
- M’Barek, L.A.; Mouse, H.A.; Jaâfari, A.; Aboufatima, R.; Benharref, A.; Kamal, M.; Bénard, J.; El Abbadi, N.; Bensalah, M.; et al. Cytotoxic effect of essential oil of thyme (Thymus broussonettii) on the IGR-OV1 tumor cells resistant to chemotherapy. Braz. J. Med. Biol. Res. 2007, 40, 1537–1544. [Google Scholar]
- Loizzoa, M.R.; Saabb, A.; Tundisa, R.; Stattia, G.A.; Lamprontic, I.; Menichinia, F.; Gambarid, R.; Cinatle, J.; Doer, H.W. Phytochemical analysis and in vitro evaluation of the biological activity against herpes simplex virus type 1 (HSV-1) of Cedrus libani A. Rich. Phytomedicine 2008, 15, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Alviano, D.S.; Alviano, C.S. Plant extracts: Search for new alternatives to treat microbial diseases. Curr. Pharm. Biotechnol. 2009, 10, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Goulart, F.R.V.; Rodrigues, I.A.; Abreu, F.; Lins, U.; Alves, P.B.; Blank, A.F.; Valoni, E.; Sebastián, G.V.; et al. Antimicrobial action and anti-corrosion effect against sulfate reducing bacteria by lemongrass (Cymbopogon citratus) essential oil and its major component, the citral. AMB Express 2013, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Simas, D.L.R.; de Amorim, S.H.B.M.; Goulart, F.R.V.; Alviano, C.S.; Alviano, D.S.; da Silva, A.J.R. Citrus species essential oils and their components can inhibit or stimulate fungal growth in fruit. Ind. Crops Prod. 2017, 98, 108–115. [Google Scholar] [CrossRef]
- Chaves, F.M.C.; Bizzo, H.R.; Angelo, P.C.S.; Xavier, J.J.B.N.; Sá Sobrinho, A.F. Yield and chemical composition of essential oil from leaves of two varieties of sacaca (Croton cajucara Benth). Rev. Bras. Plant Med. Botucatu 2006, 8, 117–119. [Google Scholar]
- Blank, A.F.; Camêlo, L.C.A.; Arrigoni-Blank, M.F.; Pinheiro, J.B.; Andrade, T.M.; Niculau, E.S.; Alves, P.B. Chemical diversity in Lippia alba (Mill.) N.E. brown germplasm. Sci. World J. 2015, 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Farias-Junior, P.A.; Rios, M.C.; Moura, T.A.; Almeida, R.P.; Alves, P.B.; Blank, A.F.; Fernandes, R.P.M.; Scher, R. Leishmanicidal activity of carvacrol-rich essential oil from Lippia sidoides Cham. Biol. Res. 2012, 45, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Aleksic, V.; Knezevic, P. Antimicrobial and antioxidative activity of extracts and essential oils of Myrtus communis L. Microbiol. Res. 2014, 169, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Kerekes, E.-B.; Deák, É.; Takó, M.; Tserennadmid, R.; Petkovits, T.; Vágvölgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Borrego, B.; Valdés, O.; Vivar, I.; Lavin, P.; Guiamet, P.; Battistoni, P.; De Saravia, S.G.; Borges, P. Essential oils of plants as biocides against microorganisms isolated from Cuban and Argentine documentary heritage. ISRN Microbiol. 2012, 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Feio, M.J.; Zinkevich, V.; Beech, I.B.; Llobet-Brossa, E.; Eaton, P.; Schmitt, J.; Guezennec, J. Desulfovibrio alaskensis sp. nov., a sulfate-reducing bacterium from a soured oil reservoir. Int. J. Syst. Evol. Microbiol. 2004, 54, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Adukwu, E.C.; Allen, S.C.; Phillips, C.A. The anti-biofilm activity of lemongrass (Cymbopogon flexuosus) and grapefruit (Citrus paradisi) essential oils against five strains of Staphylococcus aureus. J. Appl. Microbiol. 2012, 113, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; Wood, W.A.; Key, R.; Nakamura, K.; Stahl, D.A. Isolation of three species of Geotoga and Petrotoga: Two new genera, representing a new lineage in the bacterial line of descent distantly related to the “Thermotogales”. Syst. Appl. Microbiol. 1993, 16, 191–200. [Google Scholar] [CrossRef]
- Bowman, J.P.; Rea, S.M.; McCammon, S.A.; McMeekin, T.A. Diversity and community structure within anoxic sediment from marine salinity meromictic lakes and a coastal meromictic marine basin, Vestfold Hills, Eastern Antarctica. Environ. Microbiol. 2000, 2, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Gieg, L.M.; Parisi, V.A.; Tanner, R.S.; Tringe, S.G.; Bristow, J.; Suflita, J.M. Biocorrosive thermophilic microbial communities in Alaskan North slope oil facilities. Environ. Sci. Technol. 2009, 43, 7977–7984. [Google Scholar] [CrossRef] [PubMed]
- Hubert, C.R.J.; Oldenburg, T.B.P.; Fustic, M.; Gray, N.D.; Larter, S.R.; Penn, K.; Rowan, A.K.; Seshadri, R.; Sherry, A.; et al. Massive dominance of Epsilonproteobacteria in formation waters from a Canadian oil sands reservoir containing severely biodegraded oil. Environ. Microbiol. 2012, 14, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, A.; Alsop, E.B.; Chambers, B.; Lomans, B.P.; Head, I.M.; Tsesmetzis, N. Complementary microorganisms in highly corrosive biofilms from an offshore oil production facility. Appl. Environ. Microbiol. 2016, 82, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Tian, H.; Li, G.; Sun, H.; Ma, T. Microbial diversity and abundance in the Xinjiang Luliang long-term water-flooding petroleum reservoir. Microbiologyopen 2015, 4, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kojima, H.; Fukui, M. Desulfoplanes formicivorans gen. nov., sp. nov., a novel sulfate-reducing bacterium isolated from a blackish meromictic lake, and emended description of the family Desulfomicrobiaceae. Int. J. Syst. Evol. Microbiol. 2015, 65, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.G.; Hulem, C.; Colgate, J.; Anselmo, C. Antibacterial susceptibility of plaque bacteria. J. Dent. Res. 1979, 58, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Postgate, J.R. The Sulphate-Reducing Bacteria, 2nd ed.; Cambridge University Press: Cambridge, UK, 1984; pp. 9–55. [Google Scholar]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M.H. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Wagner, M.; Roger, A.J.; Flax, J.L.; Brusseau, G.A.; Stahl, D.A. Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration. J. Bacteriol. 1998, 180, 2975–2982. [Google Scholar] [PubMed]
- Kondo, R.; Nedwell, D.B.; Purdy, K.J.; Silva, S.D. Detection and enumeration of sulphate reducing bacteria in estuarine sediments by competitive PCR. Geomicrobiol. J. 2004, 21, 145–157. [Google Scholar] [CrossRef]
- Taylor, J. The estimation of members of bacteria by tenfold dilution series. J. Appl. Microbiol. 1962, 25, 54–61. [Google Scholar]
Sample Availability: Not Available. |

{kind=link}
{kind=link}
| Bands | Closest Database Match to A Cultivable Bacterium (Accession Number) | Identity (%) |
|---|---|---|
| B1 | Pelobacter sp. (CP000142.2) | 98 |
| B2 | Geotoga petraea (L10658.1) | 100 |
| B3 | Desulfoplanes formicivorans (LC017841.1) | 99 |
| B4 | Pelobacter carbinilicus (U23141.1) | 98 |
| B5 | Marinobacterium sp. (KU052621.1) | 99 |
| B6 | Marinobacterium sp. (HG315015.1) | 99 |
| Plant | Popular Name | EO Distilled from (Plant Part) | EO Major Components | Reference | MIC of EO Against D. alaskensis (µg/mL) | MIC of EO in Produced Water (µg/mL) |
|---|---|---|---|---|---|---|
| Citrus aurantifolia | “Mirim” lime | fruit peels | 31% R-Limonene, 8.5% β-pinene, 16% citral | [23] | 78 | 156 |
| C. latifolia | “Tahiti” lime | fruit peels | 35% R-Limonene, 5% β-pinene, 5% citral | [23] | 625 | - |
| C. limon | “Siciliano” lemon | fruit peels | 53% R-Limonene, 13% β-pinene, 4% citral | [23] | 156 | - |
| C. limonia | “Cravo” lime | fruit peels | 65% R-Limonene, 9% β-pinene | [23] | 625 | - |
| Croton cajucara | White “sacaca” | leaves | 28% Linalool | [24] | 1250 | - |
| C. cajucara | Red “sacaca” | leaves | 18% Linalool, 25% 7-hydroxy-calamenene | [24] | 2500 | - |
| Cymbopogon citratus | Lemongrass | leaves | 75% Citral | [22] | 78 | 156 |
| Lippia alba (LA10) * | Lemon balm | leaves | 77% Citral | [25] | 156 | 312 |
| L. alba (LA13) | Lemon balm | leaves | 45% Limonene, 40% Carvone | [25] | 1250 | - |
| L. alba (LA22) | Lemon balm | leaves | 84% Linalool | [25] | 156 | 156 |
| L. alba (LA29) | Lemon balm | leaves | 68% Citral | [25] | 312 | - |
| L. alba (LA44) | Lemon balm | leaves | 82% Citral | [25] | 78 | 78 |
| L. alba (LA57) | Lemon balm | leaves | 19% Limonene, 77% Carvone | [25] | 625 | - |
| Lippia sidoides (LSID001) | Pepper rosmarin | leaves | 83% Thymol | [26] | 625 | - |
| L. sidoides (LSID003) | Pepper rosmarin | leaves | 80% Thymol | [26] | 625 | - |
| L. sidoides (LSID004) | Pepper rosmarin | leaves | 80% Thymol | [26] | 1250 | - |
| L. sidoides (LSID005) | Pepper rosmarin | leaves | 76% Thymol | [26] | 2500 | - |
| L. sidoides (LSID006) | Pepper rosmarin | leaves | 81% Thymol | [26] | 625 | - |
| L. sidoides (LSID104) | Pepper rosmarin | leaves | 8% Thymol, 55% Carvacrol | [26] | 2500 | - |
| L. sidoides (LSID301) | Pepper rosmarin | leaves | 76% Thymol | [26] | 2500 | - |
| Major Components | Purity, Sigma Aldrich Reference Number | MIC Against D. alaskensis (µg/mL) | MIC in Produced Water (µg/mL) |
|---|---|---|---|
| Citral | 95%, C83007 | 78 | 78 |
| Linalool | 97%, L2602 | 78 | 625 |
| Geraniol | 98%, 163333 | 78 | 312 |
| Nerol | ~98%, N7761 | 625 | - |
| Eugenol | 99%, E51791 | 78 | 312 |
| R-Limonene | 97%, 183164 | 2500 | - |
| S-Limonene | 96%, 218367 | 2500 | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, P.M.d.; Goulart, F.R.d.V.; Marques, J.M.; Bizzo, H.R.; Blank, A.F.; Groposo, C.; Sousa, M.P.d.; Vólaro, V.; Alviano, C.S.; Moreno, D.S.A.; et al. Growth Inhibition of Sulfate-Reducing Bacteria in Produced Water from the Petroleum Industry Using Essential Oils. Molecules 2017, 22, 648. https://doi.org/10.3390/molecules22040648
Souza PMd, Goulart FRdV, Marques JM, Bizzo HR, Blank AF, Groposo C, Sousa MPd, Vólaro V, Alviano CS, Moreno DSA, et al. Growth Inhibition of Sulfate-Reducing Bacteria in Produced Water from the Petroleum Industry Using Essential Oils. Molecules. 2017; 22(4):648. https://doi.org/10.3390/molecules22040648
Chicago/Turabian StyleSouza, Pamella Macedo de, Fátima Regina de Vasconcelos Goulart, Joana Montezano Marques, Humberto Ribeiro Bizzo, Arie Fitzgerald Blank, Claudia Groposo, Maíra Paula de Sousa, Vanessa Vólaro, Celuta Sales Alviano, Daniela Sales Alviano Moreno, and et al. 2017. "Growth Inhibition of Sulfate-Reducing Bacteria in Produced Water from the Petroleum Industry Using Essential Oils" Molecules 22, no. 4: 648. https://doi.org/10.3390/molecules22040648