
Levo-Corydalmine Alleviates Neuropathic Cancer Pain Induced by Tumor Compression via the CCL2/CCR2 Pathway

Abstract
:1. Introduction
2. Results
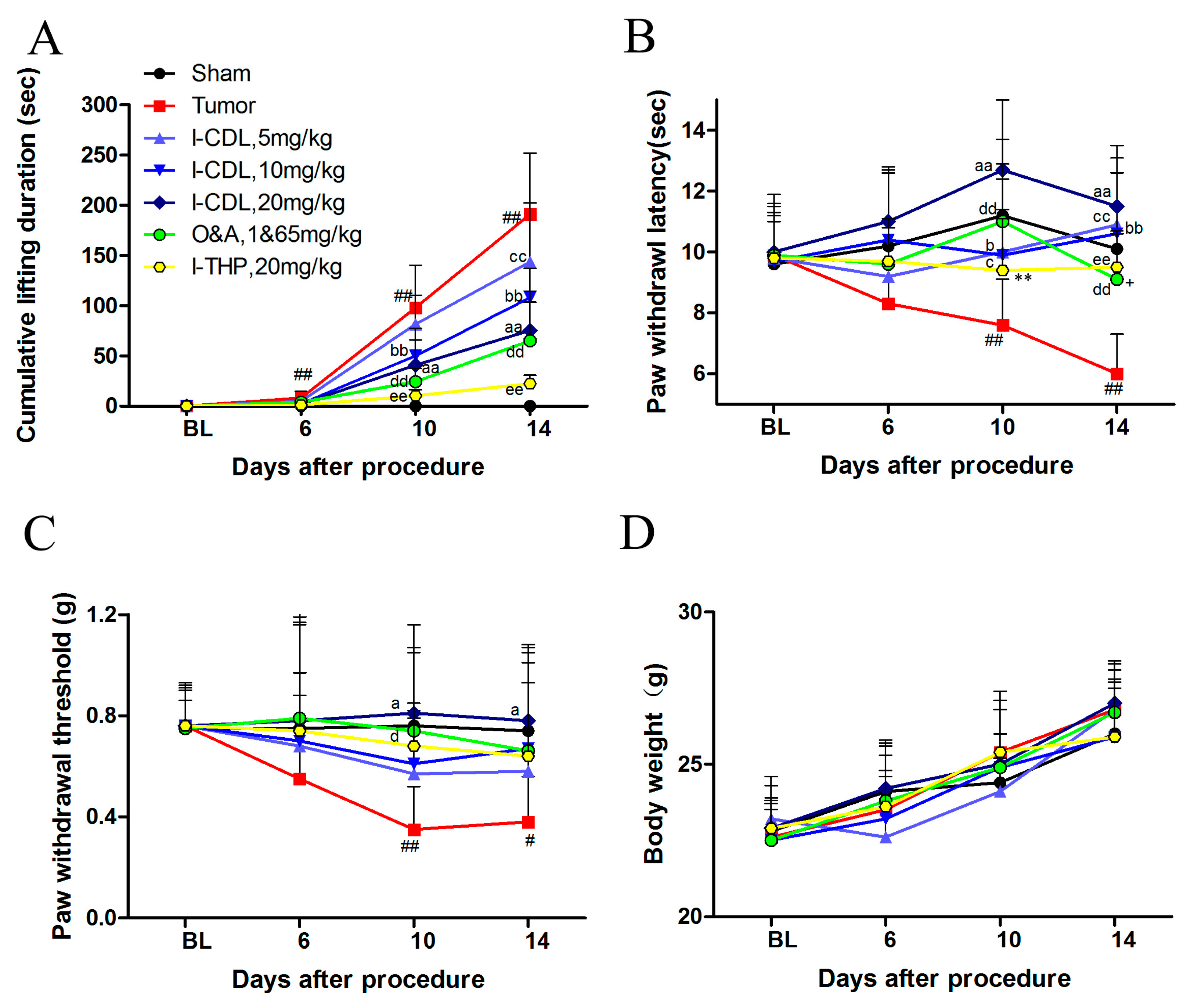
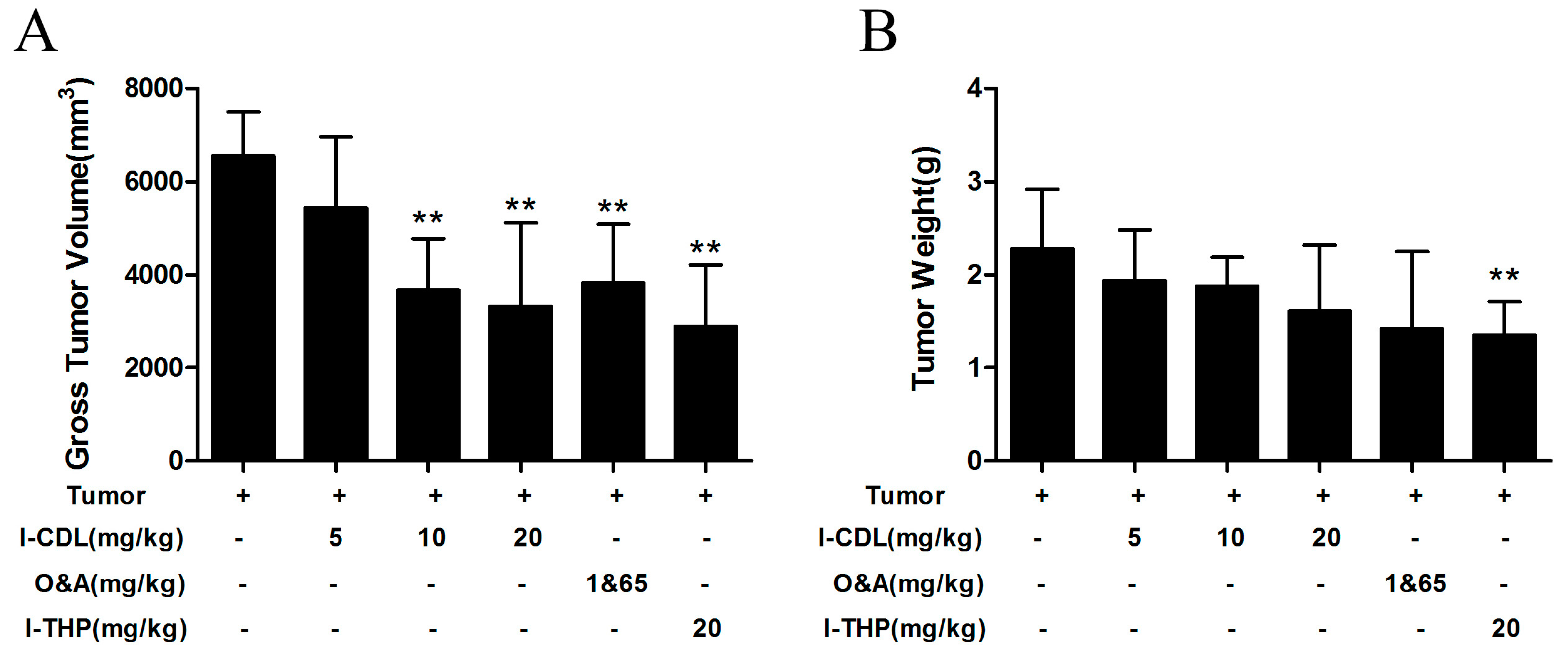
2.1. l-CDL Attenuated Tumor Compression-Induced Pain Hypersensitivity
2.2. l-CDL Decreased the Level of Tumor Compression-Induced TNF-α and IL-1β Expression in Serum and Spinal Cord
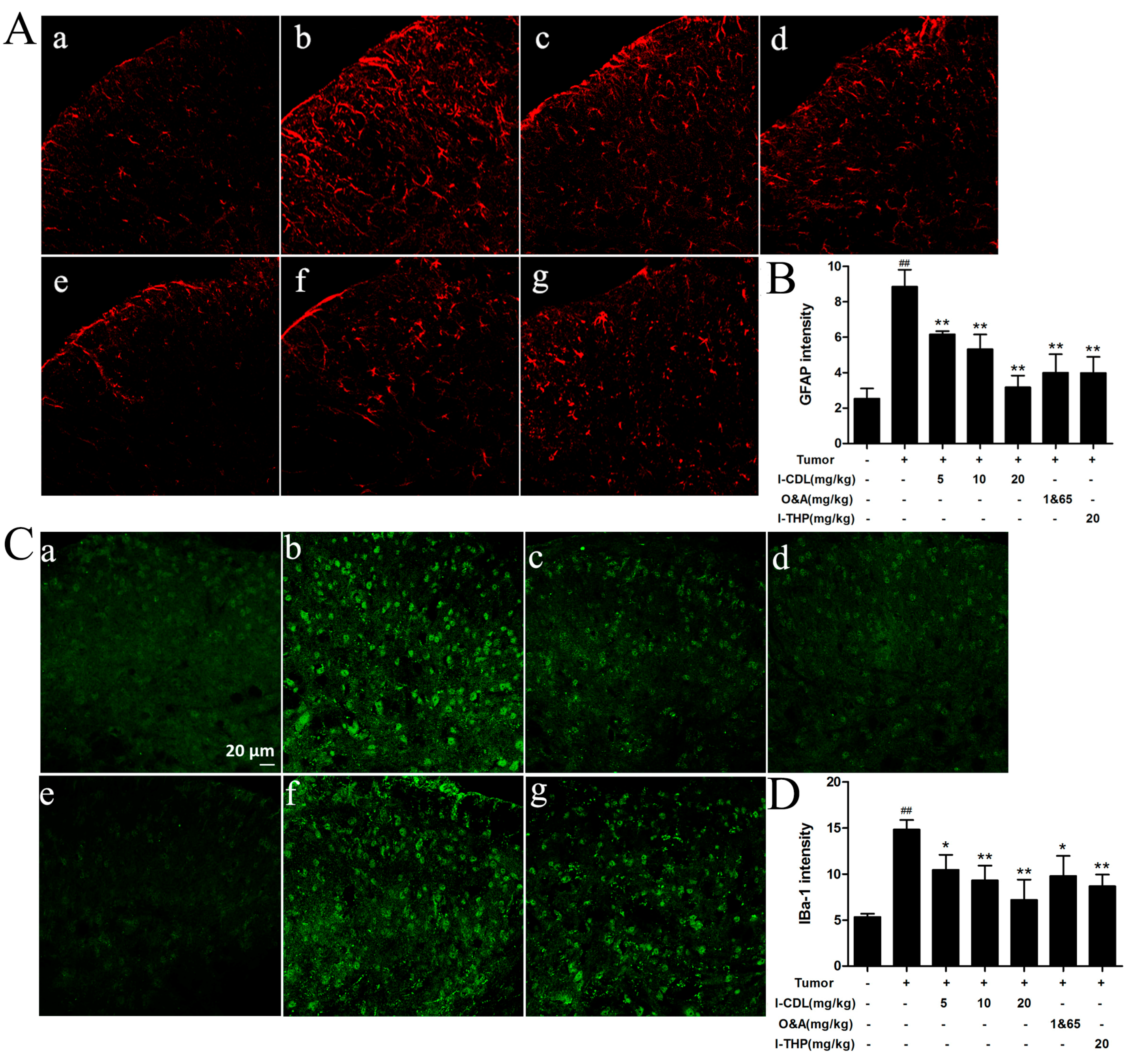
2.3. l-CDL Attenuated Tumor Compression-Induced Glial Cells Activation in the Spinal Cord
2.4. l-CDL Attenuated Tumor Compression-Induced CCL2 and CCR2 Expression in the Spinal Segments
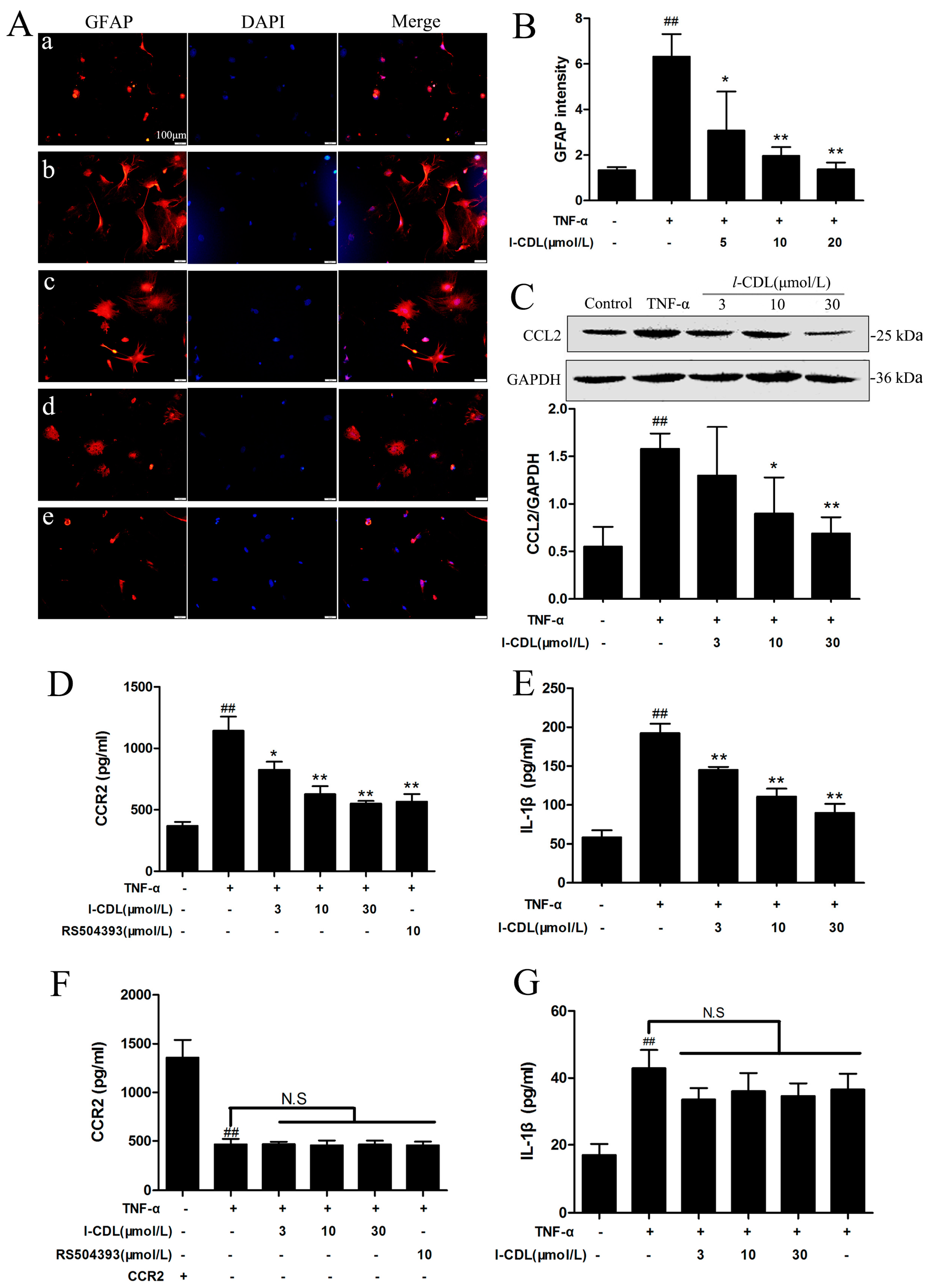
2.5. l-CDL Suppressed the TNF-α-Induced Astrocyte Activation and Downregulated CCL2/CCR2 in the Astrocyte
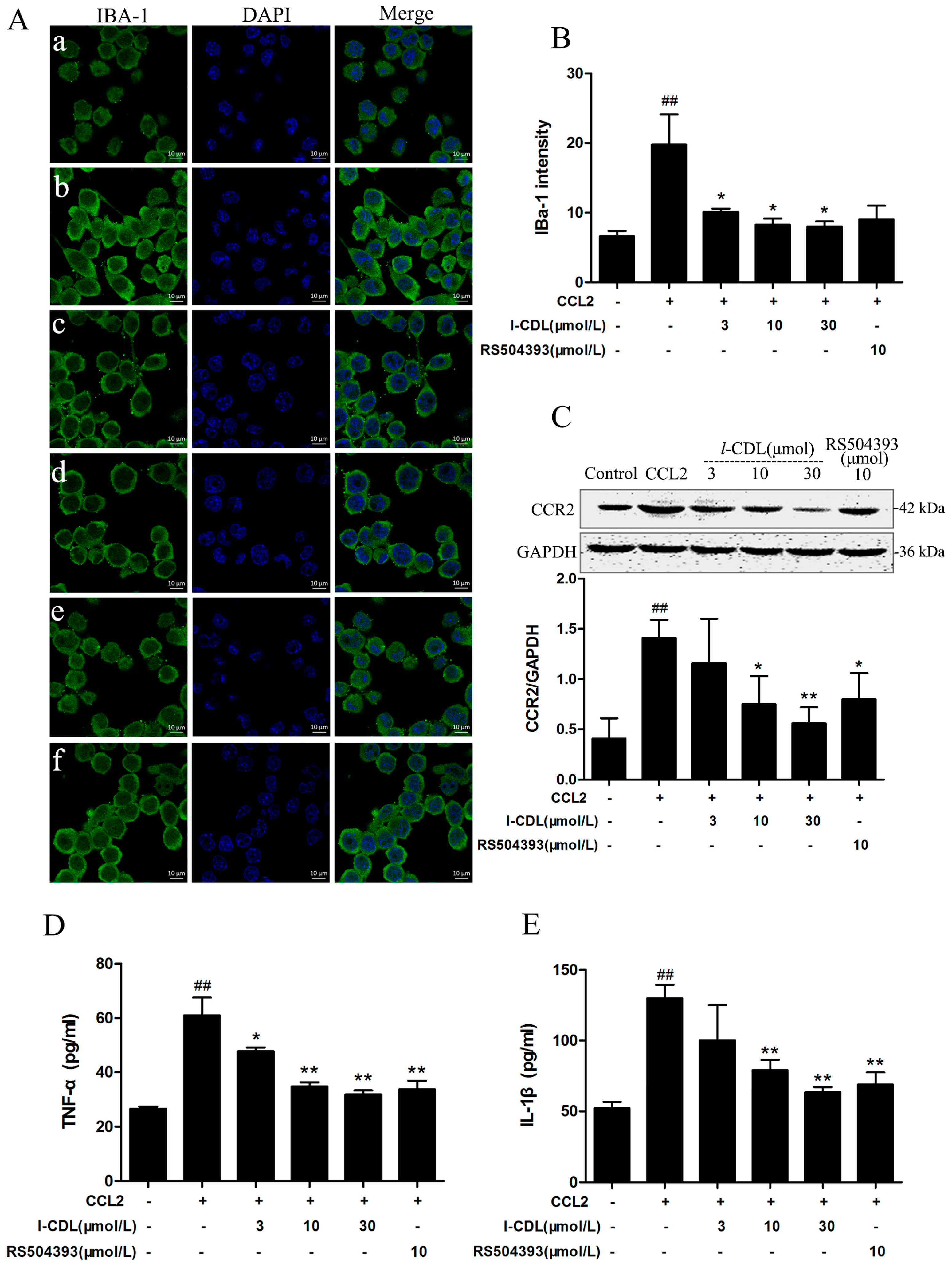
2.6. l-CDL Suppressed the CCL2-Induced BV-Microglia activation, Down-Regulated CCR2 Expression, Reduced TNF-α and IL-1β Secretion
2.7. l-CDL Alleviated the Tumor Compression in the Sciatic Nerve
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Tumor Inoculation
4.3. Drugs and Administration
4.4. Gross Behavior
4.5. Assessment of Thermal Hyperalgesia
4.6. Assessment of Mechanical Allodynia
4.7. Cell Cultures
4.8. Immunofluorescence
4.9. RT-PCR
4.10. Western Blot
4.11. ELISA
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deandrea, S.; Montanari, M.; Moja, L.; Apolone, G. Prevalence of undertreatment in cancer pain: A review of published literature. Ann. Oncol. 2008, 19, 1985–1991. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.L. The neurobiology of cancer pain. Neuroscientist 2014, 20, 546–562. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zhang, Y.Q. Spinal glial activation contributes to pathological pain states. Neurosci. Biobehav. Rev. 2008, 32, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, M.; Tanaka, K.; Hasue, F.; Shimoyama, N. A mouse model of neuropathic cancer pain. Pain 2002, 99, 167–174. [Google Scholar] [CrossRef]
- Khasabova, I.A.; Stucky, C.L.; Harding-Rose, C.; Eikmeier, L.; Beitz, A.J.; Coicou, L.G.; Hanson, A.E.; Simone, D.A.; Seybold, V.S. Chemical Interactions between Fibrosarcoma Cancer Cells and Sensory Neurons Contribute to Cancer Pain. J. Neurosci. 2007, 27, 10289–10298. [Google Scholar] [CrossRef] [PubMed]
- Vissers, K.C.P.; Besse, K.; Wagemans, M.; Zuurmond, W.; Giezeman, M.J.M.M.; Lataster, A.; Mekhail, N.; Burton, A.W.; van Kleef, M.; Huygen, F. 23 Pain in Patients with Cancer. Pain Pract. 2011, 11, 453–475. [Google Scholar] [CrossRef] [PubMed]
- Day, C.-P.; Merlino, G.; van Dyke, T. Preclinical mouse cancer models: A Maze of opportunities and challenges. Cell 2015, 163, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Colleoni, M.; Sacerdote, P. Murine models of human neuropathic pain. Biochimi. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Pacharinsak, C.; Beitz, A. Animal Models of Cancer Pain. Comp. Med. 2008, 58, 220–233. [Google Scholar] [PubMed]
- McCaffrey, G.; Thompson, M.L.; Majuta, L.; Fealk, M.N.; Chartier, S.; Longo, G.; Mantyh, P.W. NGF blockade at early times during bone cancer development attenuates bone destruction and increases limb use. Cancer Res. 2014, 74, 7014–7023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Boyette-Davis, J.A.; Kosturakis, A.K.; Li, Y.; Yoon, S.Y.; Walters, E.T.; Dougherty, P.M. Induction of monocyte chemoattractant protein-1 (MCP-1) and its receptor CCR2 in primary sensory neurons contributes to paclitaxel-induced peripheral neuropathy. J. Pain 2013, 14, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Melli, G.; Taiana, M.; Camozzi, F.; Triolo, D.; Podini, P.; Quattrini, A.; Taroni, F.; Lauria, G. Alpha-lipoic acid prevents mitochondrial damage and neurotoxicity in experimental chemotherapy neuropathy. Exp. Neurol. 2008, 214, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Polomano, R.C.; Mannes, A.J.; Clark, U.S.; Bennett, G.J. A painful peripheral neuropathy in the rat produced by the chemotherapeutic drug, paclitaxel. Pain 2001, 94, 293–304. [Google Scholar] [CrossRef]
- Quasthoff, S.; Hartung, H.P. Chemotherapy-induced peripheral neuropathy. J. Neurol. 2002, 249, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Brigatte, P.; Sampaio, S.C.; Gutierrez, V.P.; Guerra, J.L.; Sinhorini, I.L.; Curi, R.; Cury, Y. Walker 256 tumor-bearing rats as a model to study cancer pain. J. Pain 2007, 8, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.-R.; Tan, P.-H.; Cheng, J.-K.; Liu, Y.-C.; Ji, R.-R. Microglia: A Promising Target for Treating Neuropathic and Postoperative Pain, and Morphine Tolerance. J. Formos. Med. Assoc. 2011, 110, 487–494. [Google Scholar] [CrossRef]
- Tsuda, M.; Beggs, S.; Salter, M.W.; Inoue, K. Microglia and intractable chronic pain. Glia 2013, 61, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Liu, Y.P.; Zhang, L.Y.; Yue, D.M.; Qi, D.Y.; Liu, G.J.; Liu, S. Levo-tetrahydropalmatine Attenuates Bone Cancer Pain by Inhibiting Microglial Cells Activation. Mediat. Inflamm. 2015, 2015, 752512. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, H.; Li, T.T.; Luo, H.; Gu, X.Y.; Lu, N.; Ji, R.R.; Zhang, Y.Q. Delayed activation of spinal microglia contributes to the maintenance of bone cancer pain in female Wistar rats via P2X7 receptor and IL-18. J. Neurosci. 2015, 35, 7950–7963. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Inoue, K.; Salter, M.W. Neuropathic pain and spinal microglia: A big problem from molecules in “small” glia. Trends Neurosci. 2005, 28, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Norden, D.M.; Trojanowski, P.J.; Villanueva, E.; Navarro, E.; Godbout, J.P. Sequential activation of microglia and astrocyte cytokine expression precedes increased Iba-1 or GFAP immunoreactivity following systemic immune challenge. Glia 2016, 64, 300–316. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154, S10–S28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Cao, S.; Zhu, M.D.; Liu, J.Q.; Chen, J.J.; Gao, Y.J. Contribution of chemokine CCL2/CCR2 signaling in the dorsal root ganglion and spinal cord to the maintenance of neuropathic pain in a rat model of lumbar disc herniation. J. Pain 2014, 15, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Conductier, G.; Blondeau, N.; Guyon, A.; Nahon, J.L.; Rovere, C. The role of monocyte chemoattractant protein MCP1/CCL2 in neuroinflammatory diseases. J. Neuroimmunol. 2010, 224, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Pevida, M.; González-Rodríguez, S.; Lastra, A.; García-Suárez, O.; Hidalgo, A.; Menéndez, L.; Baamonde, A. Involvement of spinal chemokine CCL2 in the hyperalgesia evoked by bone cancer in mice: A role for astroglia and microglia. Cell. Mol. Neurobiol. 2014, 34, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Reuben, S.S.; Ekman, E.F.; Raghunathan, K.; Steinberg, R.B.; Blinder, J.L.; Adesioye, J. The effect of cyclooxygenase-2 inhibition on acute and chronic donor-site pain after spinal-fusion surgery. Reg. Anesth. Pain Med. 2006, 31, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Man, Y.; Wang, X.; Jin, H.; Sun, X.; Su, X.; Hao, J.; Mi, W. Levo-tetrahydropalmatine attenuates oxaliplatin-induced mechanical hyperalgesia in mice. Sci. Rep. 2014, 4, 3905. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Ding, Y.; Wang, Z.; Ma, J.; Hong, T.; Zhu, Y.; Li, H.; Pan, S. Impacts of anti-nerve growth factor antibody on pain-related behaviors and expressions of opioid receptor in spinal dorsal horn and dorsal root ganglia of rats with cancer-induced bone pain. Mol. Pain 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.A.; Jha, R.N.; Pandey, V.B.; Singh, U.P. Effect of 1-corydalmine, an Alkaloid Isolated from Corydalis chaerophylla Roots on Spore Germination of Some Fungi. Mycobiology 2007, 35, 69–71. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Mao-Ying, Q.L.; Wang, J.; Wang, Z.F.; Mi, W.L.; Wang, X.W.; Jiang, J.W.; Huang, Y.L.; Wu, G.C.; Wang, Y.Q. Lipoxins and aspirin-triggered lipoxin alleviate bone cancer pain in association with suppressing expression of spinal proinflammatory cytokines. J. Neuroinflamm. 2012, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pineau, I.; Lacroix, S. Proinflammatory cytokine synthesis in the injured mouse spinal cord: Multiphasic expression pattern and identification of the cell types involved. J. Comp. Neurol. 2007, 500, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wang, H.; Watanabe, M.; Shimizu, K.; Zou, S.; LaGraize, S.C.; Wei, F.; Dubner, R.; Ren, K. Glial-cytokine-neuronal interactions underlying the mechanisms of persistent pain. J. Neurosci. 2007, 27, 6006–6018. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, M.; Tatsuoka, H.; Ohtori, S.; Tanaka, K.; Shimoyama, N. Change of dorsal horn neurochemistry in a mouse model of neuropathic cancer pain. Pain 2005, 114, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Hansson, E. Could chronic pain and spread of pain sensation be induced and maintained by glial activation? Acta Physiol. 2006, 187, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Hald, A.; Nedergaard, S.; Hansen, R.R.; Ding, M.; Heegaarda, A.M. Differential activation of spinal cord glial cells in murine models of neuropathic and cancer pain. Eur. J. Pain 2009, 13, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Demir, I.E.; Tieftrunk, E.; Schorn, S.; Saricaoglu, O.C.; Pfitzinger, P.L.; Teller, S.; Wang, K.; Waldbaur, C.; Kurkowski, M.U.; Wormann, S.M.; et al. Activated Schwann cells in pancreatic cancer are linked to analgesia via suppression of spinal astroglia and microglia. Gut 2016, 65, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare Mannelli, L.; Pacini, A.; Micheli, L.; Femia, A.P.; Maresca, M.; Zanardelli, M.; Vannacci, A.; Gallo, E.; Bilia, A.R.; Caderni, G.; et al. Astragali radix: Could it be an adjuvant for oxaliplatin-induced neuropathy? Sci. Rep. 2017, 7, 42021. [Google Scholar] [CrossRef] [PubMed]
- Mayo, L.; Trauger, S.A.; Blain, M.; Nadeau, M.; Patel, B.; Alvarez, J.I.; Mascanfroni, I.D.; Yeste, A.; Kivisakk, P.; Kallas, K.; et al. Regulation of astrocyte activation by glycolipids drives chronic CNS inflammation. Nat. Med. 2014, 20, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, X.Q.; Echeverry, S.; Mogil, J.S.; de Koninck, Y.; Rivest, S. Expression of CCR2 in both resident and bone marrow-derived microglia plays a critical role in neuropathic pain. J. Neurosci. 2007, 27, 12396–12406. [Google Scholar] [CrossRef] [PubMed]
- Van Steenwinckel, J.; Reaux-Le Goazigo, A.; Pommier, B.; Mauborgne, A.; Dansereau, M.A.; Kitabgi, P.; Sarret, P.; Pohl, M.; Melik Parsadaniantz, S. CCL2 released from neuronal synaptic vesicles in the spinal cord is a major mediator of local inflammation and pain after peripheral nerve injury. J. Neurosci. 2011, 31, 5865–5875. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Dong, H.; Huang, Y.; Lu, S.; Zhang, S.; Qian, Y.; Jin, W. Astrocyte-Derived CCL2 is Associated with M1 Activation and Recruitment of Cultured Microglial Cells. Cell. Physiol. Biochem. 2016, 38, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.L.; Miska, J.; Wainwright, D.A.; Dey, M.; Rivetta, C.V.; Yu, D.; Kanojia, D.; Pituch, K.C.; Qiao, J.; Pytel, P.; et al. CCL2 Produced by the glioma microenvironment is essential for the recruitment of regulatory T cells and myeloid-derived suppressor cells. Cancer Res. 2016, 76, 5671–5682. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.W.; Choi, H.J.; Park, M.J.; Choi, H.J.; Lee, S.O.; Kim, K.J.; Kim, C.H.; Hong, C.; Kim, K.H.; Joo, M.; et al. The function of cancer-shed gangliosides in macrophag. Oncotarget 2017, 8, 4436–4448. [Google Scholar] [PubMed]
- Xu, J.; Zhu, M.D.; Zhang, X.; Tian, H.; Zhang, J.H.; Wu, X.B.; Gao, Y.J. NFκB-mediated CXCL1 production in spinal cord astrocytes contributes to the maintenance of bone cancer pain in mice. J. Neuroinflamm. 2014, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Gao, X.; Xia, J.; Munoz, F.M.; Manners, M.T.; Pan, R.; Meucci, O.; Dai, Y.; Hu, H. STIMs and Orai1 regulate cytokine production in spinal astrocytes. J. Neuroinflamm. 2016, 13, 126. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds Levo-corydalmine (l-CDL) are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Drug Dose | n | Surgery |
|---|---|---|---|
| Sham | - | 7 | - |
| Tumor | - | 7 | Tumor inoculation |
| l-CDL | 5 mg/kg | 7 | Tumor inoculation |
| 10 mg/kg | 7 | Tumor inoculation | |
| 20 mg/kg | 7 | Tumor inoculation | |
| O&A | 1&65 mg/kg | 7 | Tumor inoculation |
| l-THP | 20 mg/kg | 7 | Tumor inoculation |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Kodithuwakku, N.D.; Zhou, L.; Li, C.; Han, D.; Fang, W.; Liu, J.; Li, Y. Levo-Corydalmine Alleviates Neuropathic Cancer Pain Induced by Tumor Compression via the CCL2/CCR2 Pathway. Molecules 2017, 22, 937. https://doi.org/10.3390/molecules22060937
Hu Y, Kodithuwakku ND, Zhou L, Li C, Han D, Fang W, Liu J, Li Y. Levo-Corydalmine Alleviates Neuropathic Cancer Pain Induced by Tumor Compression via the CCL2/CCR2 Pathway. Molecules. 2017; 22(6):937. https://doi.org/10.3390/molecules22060937
Chicago/Turabian StyleHu, Yahui, Nandani Darshika Kodithuwakku, Lin Zhou, Chengyuan Li, Dan Han, Weirong Fang, Jihua Liu, and Yunman Li. 2017. "Levo-Corydalmine Alleviates Neuropathic Cancer Pain Induced by Tumor Compression via the CCL2/CCR2 Pathway" Molecules 22, no. 6: 937. https://doi.org/10.3390/molecules22060937