Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents


Abstract
:1. Introduction
2. Results
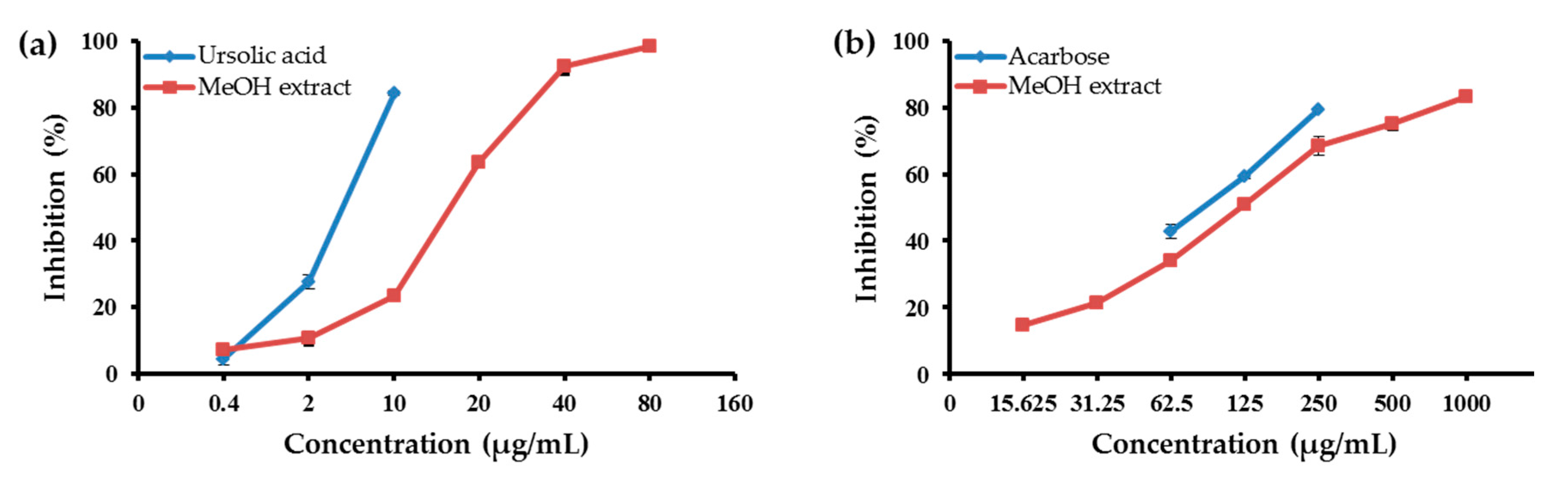
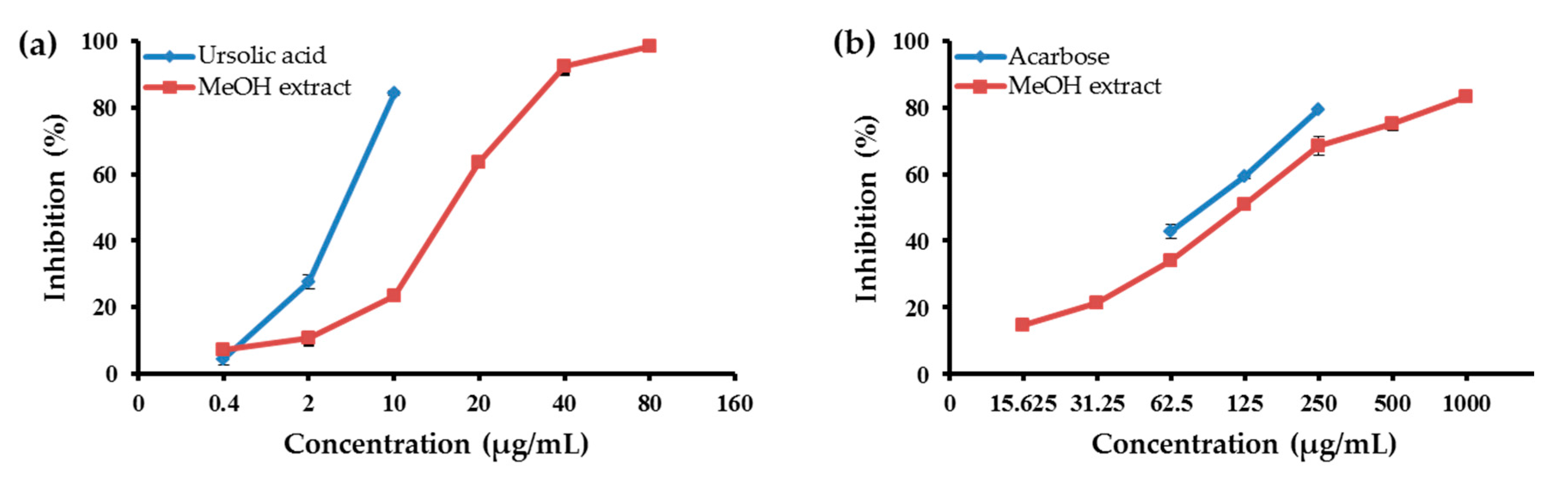
2.1. Abilities of the MeOH Extract of the Aerial Parts of E. koreanum and Its Solvent-Soluble Fractions to Inhibit PTP1B and α-Glucosidase
2.2. Abilities of the Individual Compounds to Inhibit PTP1B and α-Glucosidase Activity
2.3. Enzyme Kinetics Analysis of PTP1B Inhibition by Selected Compounds
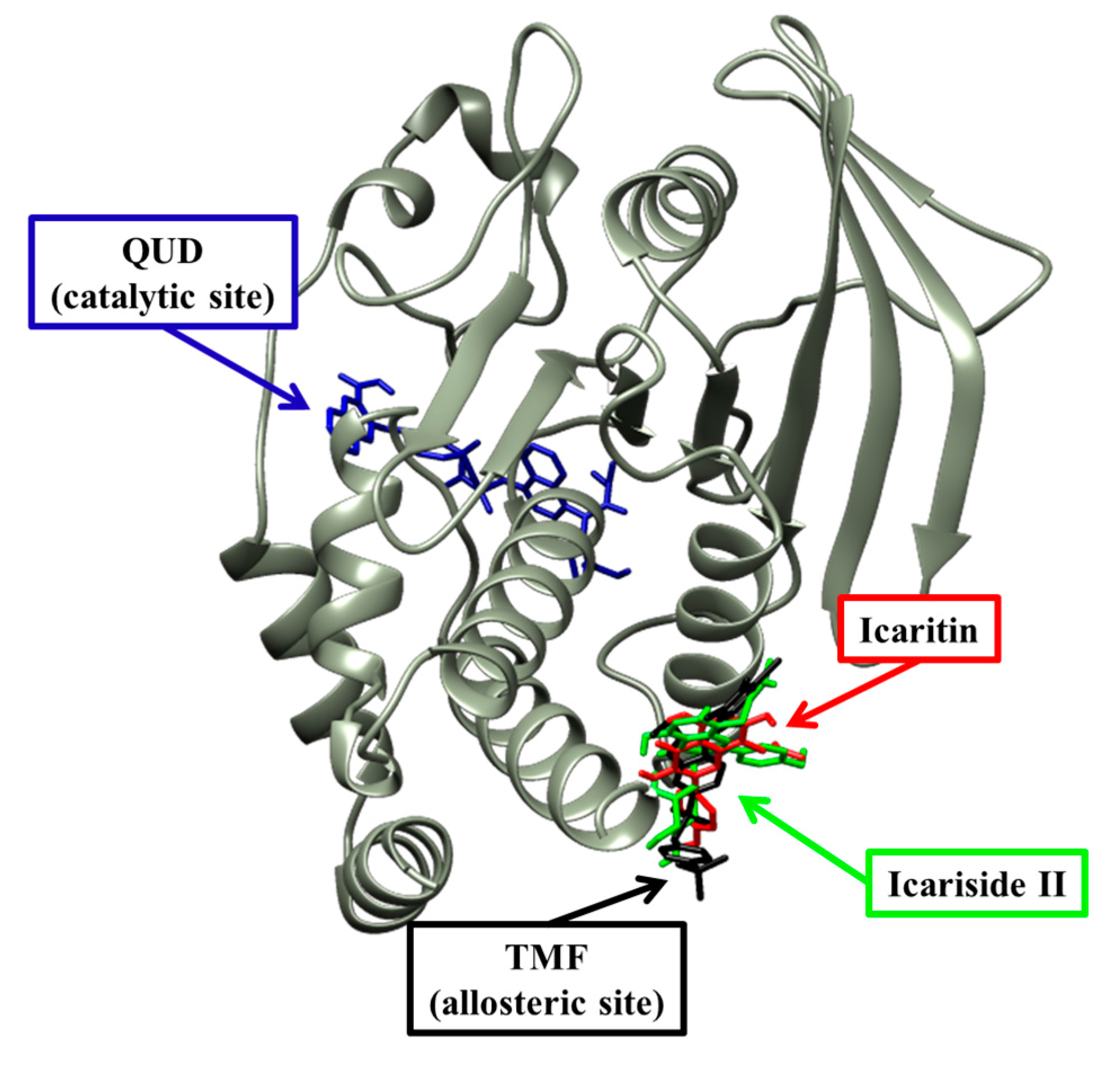
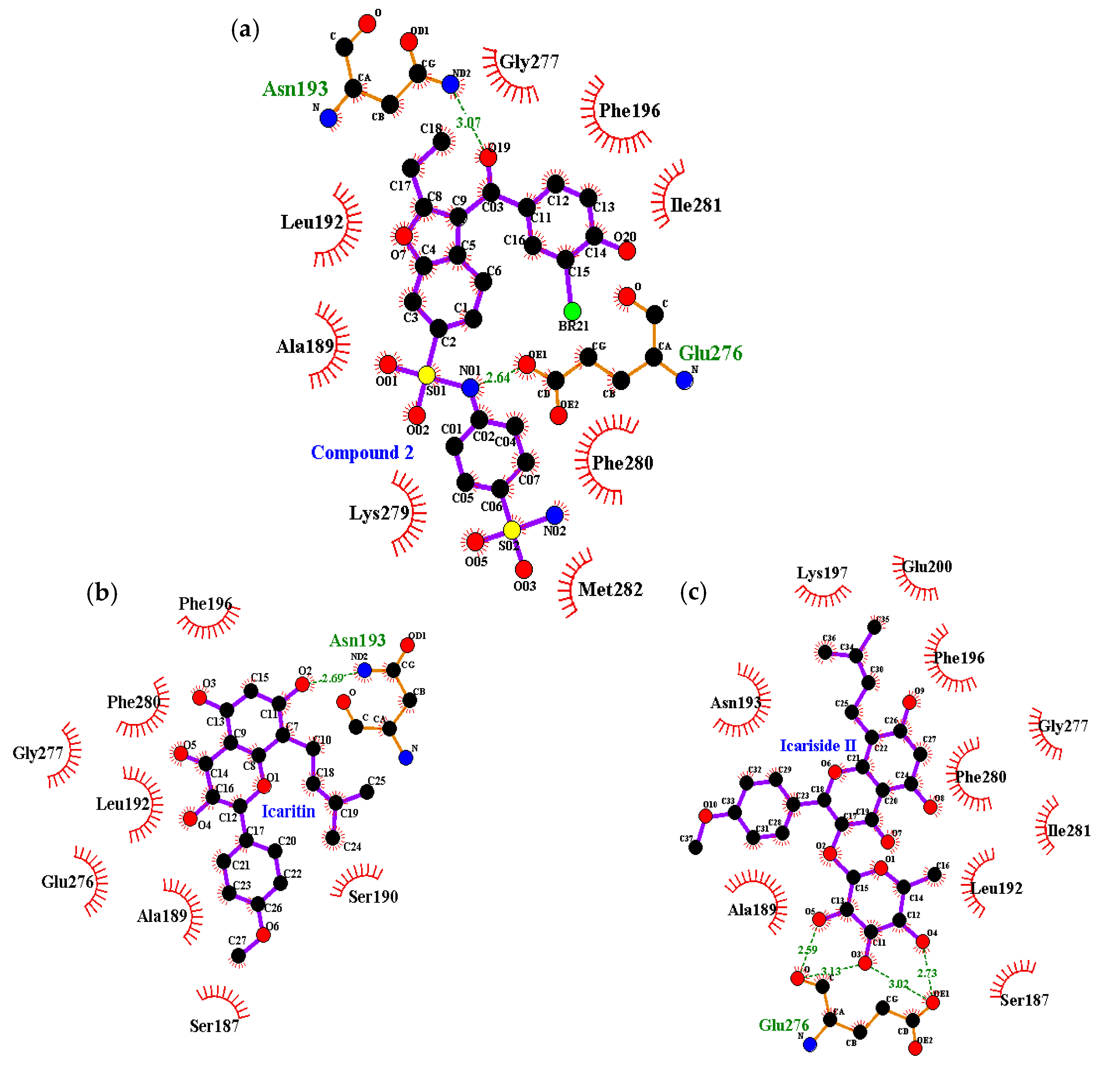
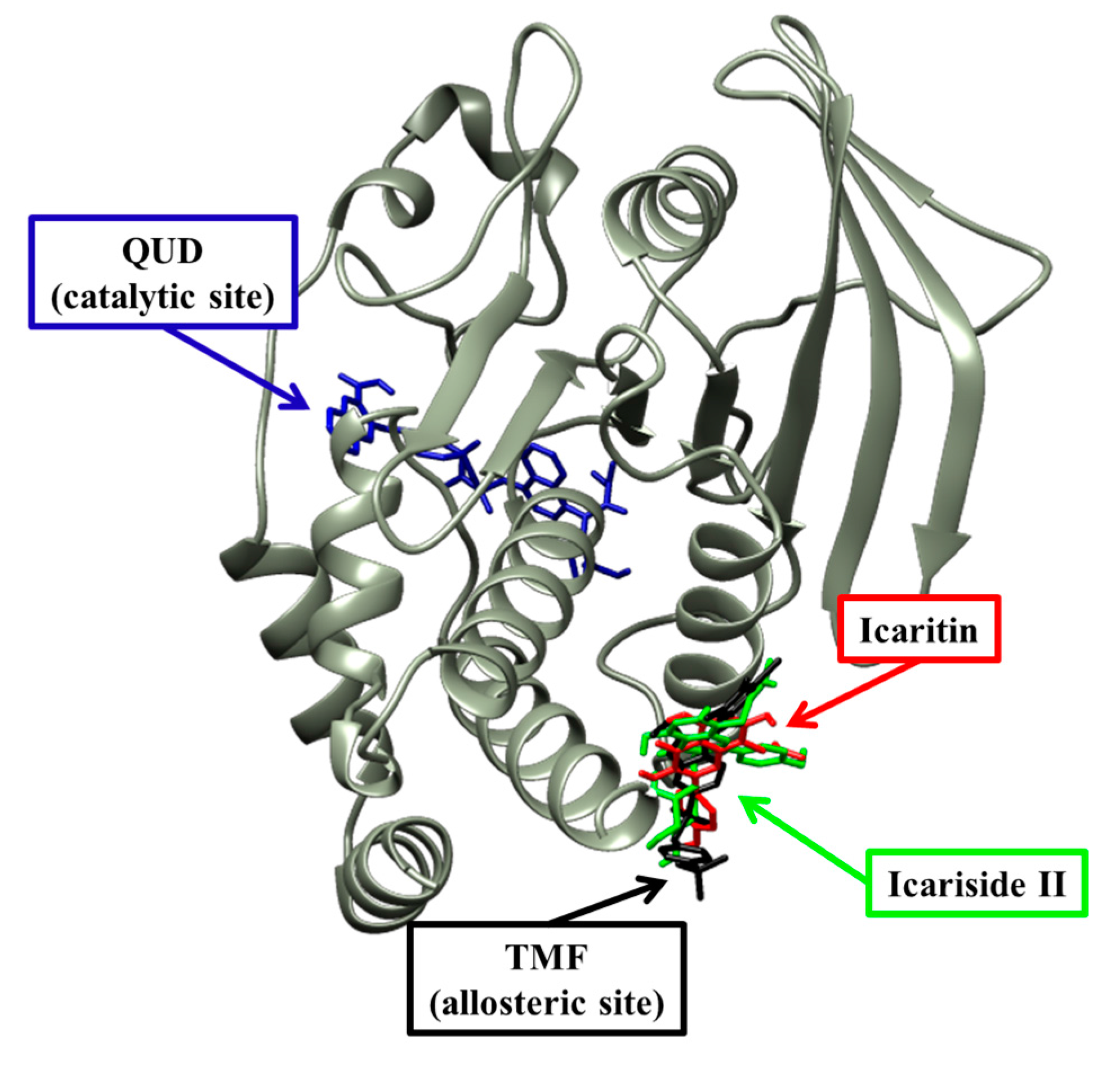
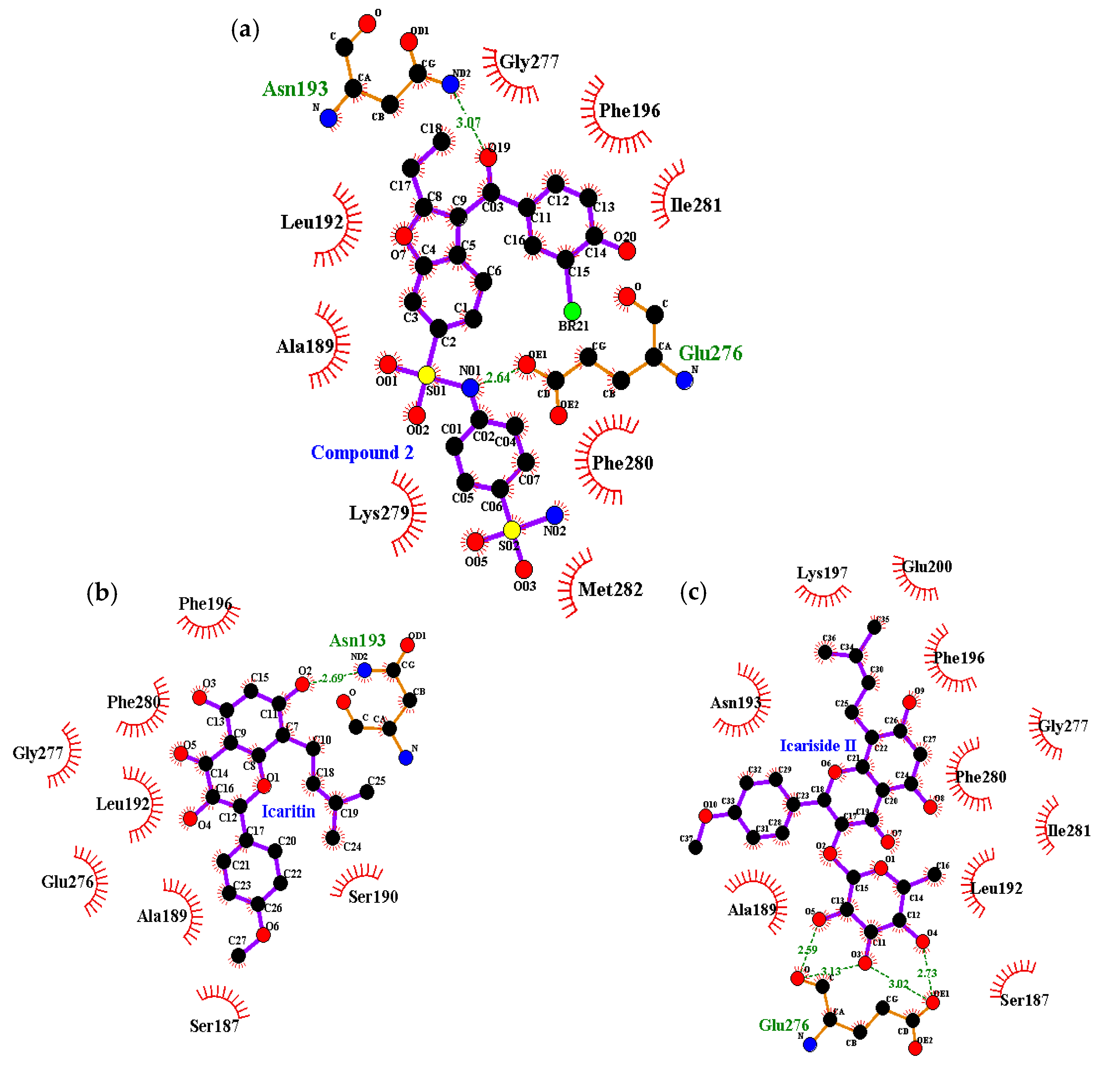
2.4. Molecular Docking Simulation of PTP1B Inhibition
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. General Experimental Procedures
4.3. Chemicals and Reagents
4.4. Extraction and Fractionation
4.5. Isolation of Prenylated Flavonoids from E. koreanum
4.6. PTP1B Activity Assay
4.7. α-Glucosidase Activity Assay
4.8. Enzyme Kinetics Analysis of PTP1B
4.9. Molecular Docking Simulation of PTP1B Inhibition
4.10. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- The Expert Committee on the Diagnosis and Classification of Diabetes Mellitus. Report of the expert committee on the diagnosis and classification of diabetes mellitus. Diabetes Care 1997, 20, 1183–1197. [Google Scholar]
- Parthasarathy, R.; Ilavarasan, R.; Karrunakaran, C.M. Antidiabetic activity of Thespesia Populnea bark and leaf extract against streptozotocin induced diabetic rats. Int. J. PharmTech. Res. 2009, 1, 1069–1072. [Google Scholar]
- Kwon, J.H.; Chang, M.J.; Seo, H.W.; Lee, J.H.; Min, B.S.; Na, M.; Kim, J.C.; Woo, M.H.; Choi, J.S.; Lee, H.K.; et al. Triterpenoids and a sterol from the stem-bark of Styrax japonica and their protein tyrosine phosphatase 1B inhibitory activities. Phytother. Res. 2008, 22, 1303–1306. [Google Scholar] [CrossRef] [PubMed]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Liu, T.; Chen, C.; Li, M.Y.; Wang, Z.Y.; Chen, R.S.; Wei, G.X.; Wang, X.Y.; Luo, D.Q. Fumosorinone, a novel PTP1B inhibitor, activates insulin signaling in insulin-resistance HepG2 cells and shows anti-diabetic effect in diabetic KKAy mice. Toxicol. Appl. Pharmacol. 2015, 285, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Shobana, S.; Sreerama, Y.N.; Malleshi, N.G. Composition and enzyme inhibitory properties of finger millet (Eleusine coracana L.) seed coat phenolics: Mode of inhibition of α-glucosidase and pancreatic amylase. Food Chem. 2009, 115, 1268–1273. [Google Scholar] [CrossRef]
- Choi, C.W.; Choi, Y.H.; Cha, M.R.; Yoo, D.S.; Kim, Y.S.; Yon, G.H.; Hong, K.S.; Kim, Y.H.; Ryu, S.Y. Yeast α-glucosidase inhibition by isoflavones from plants of Leguminosae as an in vitro alternative to acarbose. J. Agric. Food Chem. 2010, 58, 9988–9993. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S. Molecular mechanism in α-glucosidase and glucoamylase. Biosci. Biotechnol. Biochem. 1997, 61, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Van De Laar, F.A.; Lucassen, P.L.; Akkermans, R.P.; Van De Lisdonk, E.H.; Rutten, G.E.; Van Weel, C. α-Glucosidase inhibitors for patients with type 2 diabetes results from a cochrane systematic review and meta-analysis. Diabetes Care 2005, 28, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; de la Garza, A.L.; Campión, J.; Martínez, J.A.; Milagro, F.I. Antidiabetic effects of natural plant extracts via inhibition of carbohydrate hydrolysis enzymes with emphasis on pancreatic alpha amylase. Expert Opin. Ther. Targets 2012, 16, 269–297. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Miyashita, H.; Yoshimitsu, H.; Fujiwara, Y.; Nagai, R.; Ikeda, T. Two new prenylflavonoids from Epimedii Herba and their inhibitory effects on advanced glycation end-products. J. Nat. Med. 2016, 70, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Tang, Z.H.; Li, X.W.; Xie, C.X.; Lu, J.J.; Wang, Y.T. Chemical constituents, quality control, and bioactivity of Epimedii Folium (Yinyanghuo). Am. J. Chin. Med. 2015, 43, 783–834. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lien, E.J.; Lien, L.L. Chemical and pharmacological investigations of Epimedium species: A survey. Prog. Drug Res. 2003, 60, 1–57. [Google Scholar] [PubMed]
- Oh, M.H.; Houghton, P.J.; Whang, W.K.; Cho, J.H. Screening of Korean herbal medicines used to improve cognitive function for anti-cholinesterase activity. Phytomedicine 2004, 11, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.W.; Kang, S.Y.; Kim, K.H.; Song, M.Y.; Park, Y.K. Anti-diabetic effect of medicinal plants used for lower wasting-thirst in streptozotocin-induced diabetic rats. Korea J. Herbol. 2013, 28, 53–60. [Google Scholar] [CrossRef]
- Makarova, M.N.; Pozharitskaya, O.N.; Shikov, A.N.; Tesakova, S.V.; Makarov, V.G.; Tikhonov, V.P. Effect of lipid-based suspension of Epimedium koreanum Nakai extract on sexual behavior in rats. J. Ethnopharmacol. 2007, 114, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Choi, Y.H.; Kwon, H.; Lee, S.B.; Kim, D.H.; Sung, C.K.; Park, Y.I.; Dong, M.S. Estrogenic/antiestrogenic activities of a Epimedium koreanum extract and its major components: In vitro and in vivo studies. Food Chem. Toxicol. 2012, 50, 2751–2759. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Kim, H.; Choi, Y.J.; Yim, N.H.; Yang, H.J.; Ma, J.Y. Epimedium koreanum Nakai water extract exhibits antiviral activity against porcine epidermic diarrhea virus in vitro and in vivo. Evid. Based Complement. Alternat. Med. 2012, 2012, 985151. [Google Scholar] [CrossRef] [PubMed]
- Keum, J.H.; Han, H.Y.; Roh, H.S.; Seok, J.H.; Lee, J.K.; Jeong, J.; Kim, J.A.; Woo, M.H.; Choi, J.S.; Min, B.S. Analysis and stability test of the extracts from Epimedii Herba, Atractylodis Rhizoma Alba and Polygalae Radix for toxicity study. Korea J. Pharmacogn. 2014, 45, 135–140. [Google Scholar]
- Li, W.K.; Xiao, P.G.; Pan, J.Q. Complete assignment of 1H- and 13C-NMR spectra of ikarisoside A and epimedoside C. Magn. Reson. Chem. 1998, 36, 303–304. [Google Scholar] [CrossRef]
- Li, W.K.; Pan, J.Q.; Lü, M.J.; Zhang, R.Y.; Xiao, P.G. A 9,10-dihydrophenanthrene derivate from Epimedium koreanum. Phytochemistry 1995, 39, 231–233. [Google Scholar] [CrossRef]
- Lee, M.K.; Choi, Y.J.; Sung, S.H.; Shin, D.I.; Kim, J.W.; Kim, Y.C. Antihepatotoxic activity of icariin, a major constituent of Epimedium koreanum. Planta Med. 1995, 61, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wu, J.; Chen, X.; Fortenbery, N.; Eksioglu, E.; Kodumudi, K.N.; Pk, E.B.; Dong, J.; Djeu, J.Y.; Wei, S. Icariin and its derivative, ICT, exert anti-inflammatory, anti-tumor effects, and modulate myeloid derived suppressive cells (MDSCs) functions. Int. Immunopharmacol. 2011, 11, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.B.; Huang, Z.Q. Vasorelaxant effects of icariin on isolated canine coronary artery. J. Cardiovasc. Pharmacol. 2007, 49, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Tohda, C.; Nagata, A. Epimedium koreanum extract and its constituent icariin improve motor dysfunction in spinal cord injury. Evid. Based Complement. Alternat. Med. 2012, 2012, 731208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shen, C.; Chu, J.; Zhang, R.; Li, Y.; Li, L. Icariin decreases the expression of APP and BACE-1 and reduces the β-amyloid burden in an APP transgenic mouse model of Alzheimer’s disease. Int. J. Biol. Sci. 2014, 10, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Zhou, F.; Liu, T.; Li, G.Y.; Liu, J.; Gao, Z.Z.; Bai, G.Y.; Lu, H.; Xin, Z.C. Icariin ameliorates streptozotocin-induced diabetic retinopathy in vitro and in vivo. Int. J. Mol. Sci. 2012, 13, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Zheng, H.Y.; Huang, X.X.; Han, S.X.; Zhang, D.S.; Ni, J.Z.; He, X.Y. Neuroprotective effects of icariin on brain metabolism, mitochondrial functions, and cognition in triple-transgenic Alzheimer’s disease mice. CNS Neurosci. Ther. 2016, 22, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, W.K.; Xiao, P.G. Effects of icariside II from Epimedium koreanum on tumour cell lines in vitro. Pharm. Pharmacol. Commun. 1999, 5, 701–703. [Google Scholar] [CrossRef]
- Cho, N.J.; Sung, S.H.; Lee, H.S.; Jeon, M.H.; Kim, Y.C. Anti-hepatotoxic activity of icariside II, a constituent of Epimedium koreanum. Arch. Pharm. Res. 1995, 18, 289–292. [Google Scholar] [CrossRef]
- Yin, C.; Deng, Y.; Gao, J.; Li, X.; Liu, Y.; Gong, Q. Icariside II, a novel phosphodiesterase-5 inhibitor, attenuates streptozotocin-induced cognitive deficits in rats. Neuroscience 2016, 328, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu, D.; Lou, Y. A novel anticancer agent, icaritin, induced cell growth inhibition, G 1 arrest and mitochondrial transmembrane potential drop in human prostate carcinoma PC-3 cells. Eur. J. Pharmacol. 2007, 564, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, X.; Meng, J.; Wang, Z.Y. An anticancer agent icaritin induces sustained activation of the extracellular signal-regulated kinase (ERK) pathway and inhibits growth of breast cancer cells. Eur. J. Pharmacol. 2011, 658, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Wang, H.; Qi, L.; Lou, Y. Neuroprotective effects of icaritin against beta amyloid-induced neurotoxicity in primary cultured rat neuronal cells via estrogen-dependent pathway. Neuroscience 2007, 145, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Oh, M.; Kim, S.; Kim, J.; Kim, H.; Kim, S.; Houghton, P.J.; Whang, W. Epimediphine, a novel alkaloid from Epimedium koreanum inhibits acetylcholinesterase. Nat. Prod. Res. 2013, 27, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock 4 and AutoDockTools 4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation (IDF). IDF Diabetes Atlas, 7th ed.; International Diabetes Federation: Brussels, Belgium, 2015. [Google Scholar]
- Wang, L.J.; Jiang, B.; Wu, N.; Wang, S.Y.; Shi, D.Y. Natural and semisynthetic protein tyrosine phosphatase 1B (PTP1B) inhibitors as anti-diabetic agents. RSC Adv. 2015, 5, 48822–48834. [Google Scholar] [CrossRef]
- Bongard, R.D.; Lepley, M.; Thakur, K.; Talipov, M.R.; Nayak, J.; Lipinski, R.A.J.; Bohl, C.; Sweeney, N.; Ramchandran, R.; Rathore, R.; et al. Serendipitous discovery of lightinduced (In Situ) formation of an Azo-bridged dimeric sulfonated naphthol as a potent PTP1B inhibitor. BMC Biochem. 2017, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Wijeratne, E.M.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A. An epigenetic modifier induces production of (10'S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Ali, M.Y.; Choi, J.S. Promising inhibitory effects of anthraquinones, naphthopyrone, and naphthalene glycosides, from Cassia obtusifolia on α-glucosidase and human protein tyrosine phosphatases 1B. Molecules 2016, 22, 28. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Xiong, Z.; Jiang, Z.; Li, F. Osteoblastic proliferation stimulating activity of Epimedium koreanum. Nakai extracts and its flavonol glycosides. Pharm. Biol. 2005, 43, 92–95. [Google Scholar] [CrossRef]
- Huang, D.; Yang, J.; Lu, X.; Deng, Y.; Xiong, Z.; Li, F. An integrated plasma and urinary metabonomic study using UHPLC–MS: Intervention effects of Epimedium koreanum on ‘Kidney-Yang Deficiency syndrome’ rats. J. Pharm. Biomed. Anal. 2013, 76, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Kim, U.; Kim, D.H.; Dong, M.S.; Yoo, H.H. High-performance liquid chromatography-based multivariate analysis to predict the estrogenic activity of an Epimedium koreanum extract. Biosci. Biotechnol. Biochem. 2012, 76, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.A.T.; Wang, J.; Tang, J.; Lee, Y.Z.; Ng, K. Evaluation of α-glucosidase inhibition potential of some flavonoids from Epimedium brevicornum. LWT-Food Sci. Technol. 2013, 53, 492–498. [Google Scholar] [CrossRef]
- Kumar, K.M.; Anbarasu, A.; Ramaiah, S. Molecular docking and molecular dynamics studies on β-lactamases and penicillin binding proteins. Mol. BioSyst. 2014, 10, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Roy, A.; Jung, H.A.; Jung, H.J.; Choi, J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory activities of Pueraria lobata root and its constituents. J. Ethnopharmacol. 2016, 194, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, S.K.; Goswami, N.; Selvaraj, S.; Muthusamy, V.S.; Lakshmi, B.S. Molecular dynamics approach to probe the allosteric inhibition of PTP1B by chlorogenic and cichoric acid. J. Chem. Inf. Model. 2012, 52, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Mou, L.; Shen, Q.; Lu, S.; Li, C.; Liu, X.; Wang, G.; Li, S.; Geng, L.; Liu, Y.; et al. ASD v2. 0: Updated content and novel features focusing on allosteric regulation. Nucleic Acids Res. 2014, 42, D510–D516. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, J.; Lu, S.; Huang, W.; Geng, L.; Shen, Q.; Zhang, J. The mechanism of allosteric inhibition of protein tyrosine phosphatase 1B. PLoS ONE 2014, 9, e97668. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Wang, Q. Recent development of small molecular specific inhibitor of protein tyrosine phosphatase 1B. Med. Res. Rev. 2007, 27, 553–573. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Ali, M.Y.; Jung, H.A.; Oh, S.H.; Choi, R.J.; Kim, E.J. Protein tyrosine phosphatase 1B inhibitory activity of alkaloids from Rhizoma Coptidis and their molecular docking studies. J. Ethnopharmacol. 2015, 171, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Kim, M.; Han, J. Icariin metabolism by human intestinal microflora. Molecules 2016, 21, 1158. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Chen, L. Icariin reduces mitochondrial oxidative stress injury in diabetic rat hearts. Zhongguo Zhong Yao Za Zhi 2011, 36, 1503–1507. [Google Scholar] [PubMed]
- Ma, P.; Zhang, S.; Su, X.; Qiu, G.; Wu, Z. Protective effects of icariin on cisplatin-induced acute renal injury in mice. Am. J. Transl. Res. 2015, 7, 2105. [Google Scholar] [PubMed]
- Tian, W.; Lei, H.; Guan, R.; Xu, Y.; Li, H.; Wang, L.; Yang, B.; Gao, Z.; Xin, Z. Icariside II ameliorates diabetic nephropathy in streptozotocin-induced diabetic rats. Drug Des. Devel. Ther. 2015, 9, 5147–5157. [Google Scholar] [PubMed]
- Zhang, W.; Xing, B.; Yang, L.; Shi, J.; Zhou, X. Icaritin attenuates myocardial ischemia and reperfusion injury via anti-inflammatory and anti-oxidative stress effects in rats. Am. J. Chin. Med. 2015, 43, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Zhang, Z.; Sun, E.; Li, S.; Jia, X. Statistically designed enzymatic hydrolysis of an icariin/β-cyclodextrin inclusion complex optimized for production of icaritin. Acta Pharm. Sin. B 2012, 2, 83–89. [Google Scholar] [CrossRef]
- Liu, R.; Li, A.; Sun, A.; Cui, J.; Kong, L. Preparative isolation and purification of three flavonoids from the Chinese medicinal plant Epimedium koreamum Nakai by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1064, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Ma, X.; Wu, Y.; Chen, Y.; Zeng, L.; Liu, J.; Sun, W.; Wang, D.; Hu, Y. Determine the structure of phosphorylated modification of icariin and its antiviral activity against duck hepatitis virus A. BMC Vet. Res. 2015, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Hirayama, F.; Suto, K.; Sagara, K.; Yoshida, T. Three flavonol glycosides from Epimedium koreanum. Phytochemistry 1988, 27, 911–913. [Google Scholar] [CrossRef]
- Cui, L.; Na, M.; Oh, H.; Bae, E.Y.; Jeong, D.G.; Ryu, S.E.; Kim, S.; Kim, B.Y.; Oh, W.K.; Ahn, J.S. Protein tyrosine phosphatase 1B inhibitors from Morus root bark. Bioorg. Med. Chem. Lett. 2006, 16, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, X.D.; Song, Y.W.; Liu, J.W. A microplate-based screening method for alpha-glucosidase inhibitors. Chin. J. Clin. Pharmacol. Ther. 2005, 10, 1128–1134. [Google Scholar]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Dixon, Á. The determination of enzyme inhibitor constants. Biochem. J. 1953, 55, 170. [Google Scholar] [CrossRef] [PubMed]
- Cornish-Bowden, A. A simple graphical method for determining the inhibition constants of mixed, uncompetitive and non-competitive inhibitors. Biochem. J. 1974, 137, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, F.C.; Koetzle, T.F.; Williams, G.J.; Meyer, E.F.; Brice, M.D.; Rodgers, J.R.; Kennard, O.; Shimanouchi, T.; Tasumi, M. The protein data bank: A computer-based archival file for macromolecular structures. Eur. J. Biochem. 1977, 80, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The protein data bank. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, C.; Barr, K.J.; Kung, J.; Zhu, J.; Erlanson, D.A.; Shen, W.; Fahr, B.J.; Zhong, M.; Taylor, L.; Randal, M.; et al. Allosteric inhibition of protein tyrosine phosphatase 1B. Nat. Struct. Mol. Biol. 2004, 11, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds (icaritin, icariside II, icariin, epimedin A, epimedin B and epimedin C) are available from the authors and commercial sources. |
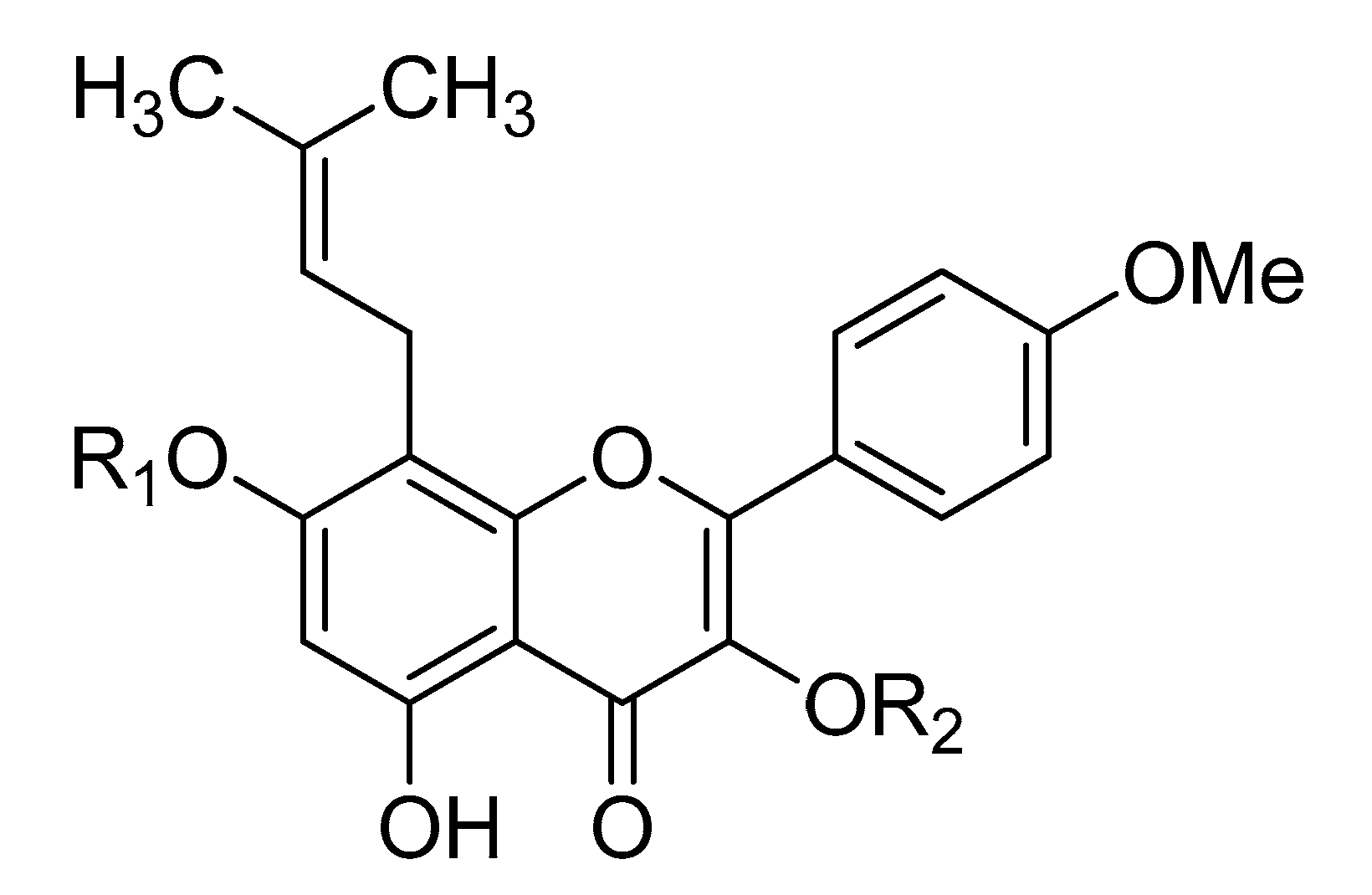
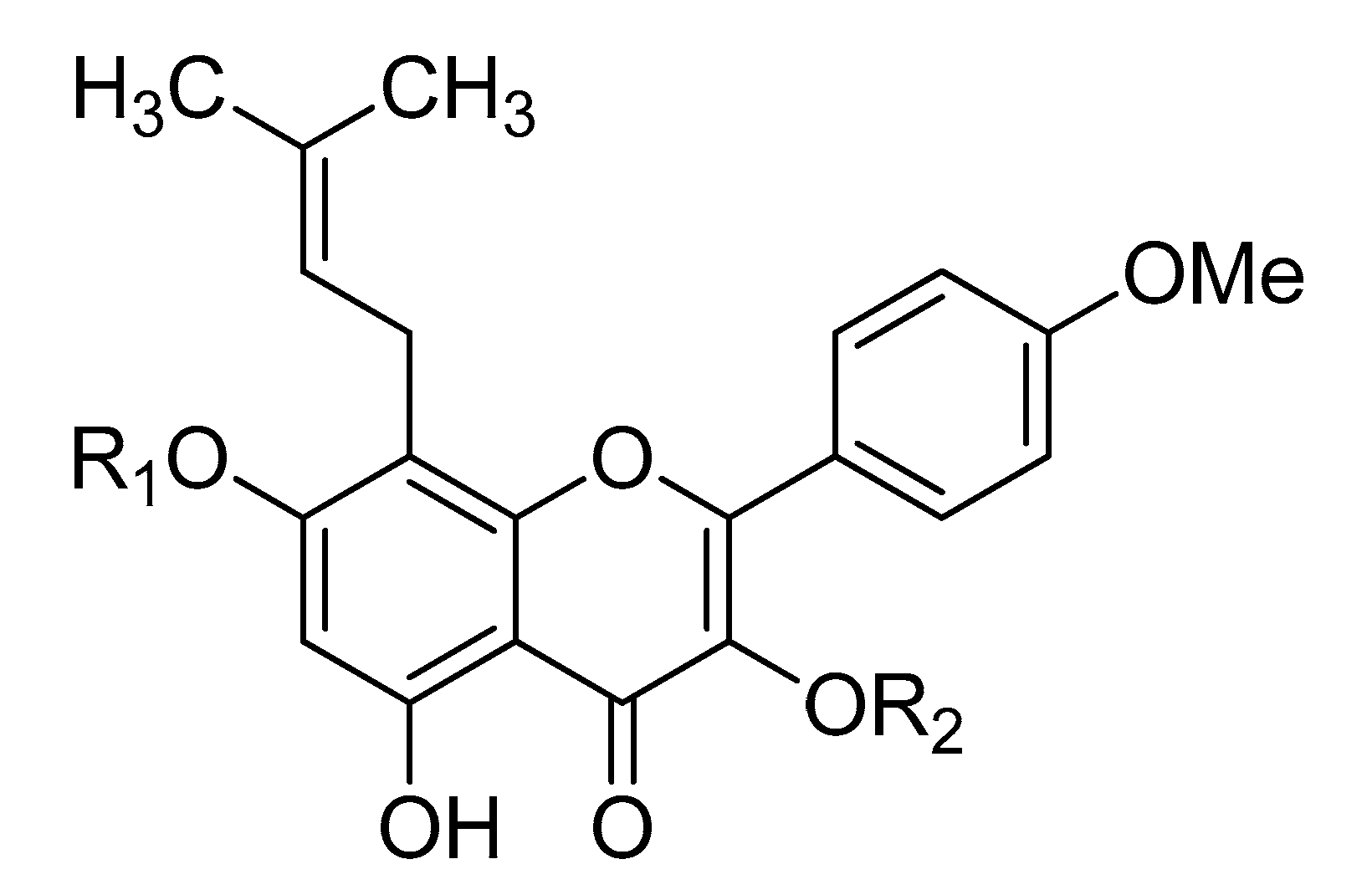
| Icariside II | R1 = H | R2 = rha |
| Icariin | R1 = glc | R2 = rha |
| Epimedin A | R1 = glc | R2 = rha-glc |
| Epimedin B | R1 = glc | R2 = rha-xyl |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Yield (g) | IC50 (μg/mL) a | |
|---|---|---|---|
| PTP1B | α-Glucosidase | ||
| MeOH extract | 665.2 | 18.83 ± 0.14 | 120.75 ± 3.06 |
| CH2Cl2 fraction | 56.94 | 3.51 ± 0.05 | 47.69 ± 0.32 |
| EtOAc fraction | 78.20 | 27.09 ± 0.30 | 75.87 ± 0.38 |
| n-BuOH fraction | 237.12 | 14.44 ± 0.68 | 138.90 ± 0.02 |
| H2O fraction | 268.06 | 38.36 ± 1.29 | 310.01 ± 1.29 |
| Ursolic acid b | - | 5.54 ± 0.13 | |
| Acarbose b | - | 87.83 ± 1.08 | |
| Compound | PTP1B | α-Glucosidase | ||
|---|---|---|---|---|
| IC50 (μM) a | Ki (μM) b | Inhibition Type c | IC50 (μM) a | |
| Icaritin | 11.59 ± 1.39 | 11.41 | Non-competitive | 74.42 ± 0.01 |
| Icariside II | 9.94 ± 0.15 | 11.66 | Non-competitive | 106.59 ± 0.44 |
| Icariin | >300 | - | - | >300 |
| Epimedin A | >300 | - | - | >300 |
| Epimedin B | >300 | - | - | >300 |
| Epimedin C | >300 | - | - | >300 |
| Ursolic acid d | 8.24 ± 0.30 | |||
| Acarbose d | 101.16 ± 3.69 | |||
| Compound | Binding Energy a (kcal/mol) | No. of H-Bonds | H-Bond Interacting Residues b | Van der Waals Bond Interacting Residues c |
|---|---|---|---|---|
| Compound 23 d (catalytic inhibitor) | −11.23 | 11 | Tyr46, Asp48, Arg24, Ser216, Ala217, Arg221, Arg254, Gln262 | Ser28, Val49, Lly116, Lly120, Cys215, Ile219, Gly220, Met258, Gly259 |
| Compound 2 d (allosteric inhibitor) | −10.98 | 2 | Asn193, Glu276 | Phe196, Gly277, Phe280, Ile281, Met282, Lys279, Ala189, Leu192 |
| Icaritin | −6.24 | 1 | Asn193 | Ser187, Ala189, Ser190, Leu192, Phe196, Glu276, Gly277, Phe280 |
| Icariside II | −8.77 | 4 | Glu276 | Ser187, Ala189, Leu192, Asn193, Phe196, Lys197, Glu200, Gly277, Phe280, Ile281 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.H.; Jung, H.A.; Sohn, H.S.; Kim, J.W.; Choi, J.S. Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents. Molecules 2017, 22, 986. https://doi.org/10.3390/molecules22060986
Kim DH, Jung HA, Sohn HS, Kim JW, Choi JS. Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents. Molecules. 2017; 22(6):986. https://doi.org/10.3390/molecules22060986
Chicago/Turabian StyleKim, Da Hye, Hyun Ah Jung, Hee Sook Sohn, Jin Woong Kim, and Jae Sue Choi. 2017. "Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents" Molecules 22, no. 6: 986. https://doi.org/10.3390/molecules22060986
APA StyleKim, D. H., Jung, H. A., Sohn, H. S., Kim, J. W., & Choi, J. S. (2017). Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents. Molecules, 22(6), 986. https://doi.org/10.3390/molecules22060986