Effects of Agitation, Aeration and Temperature on Production of a Novel Glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and Scale-Up Based on Volumetric Oxygen Transfer Coefficient

Abstract
:1. Introduction
2. Results
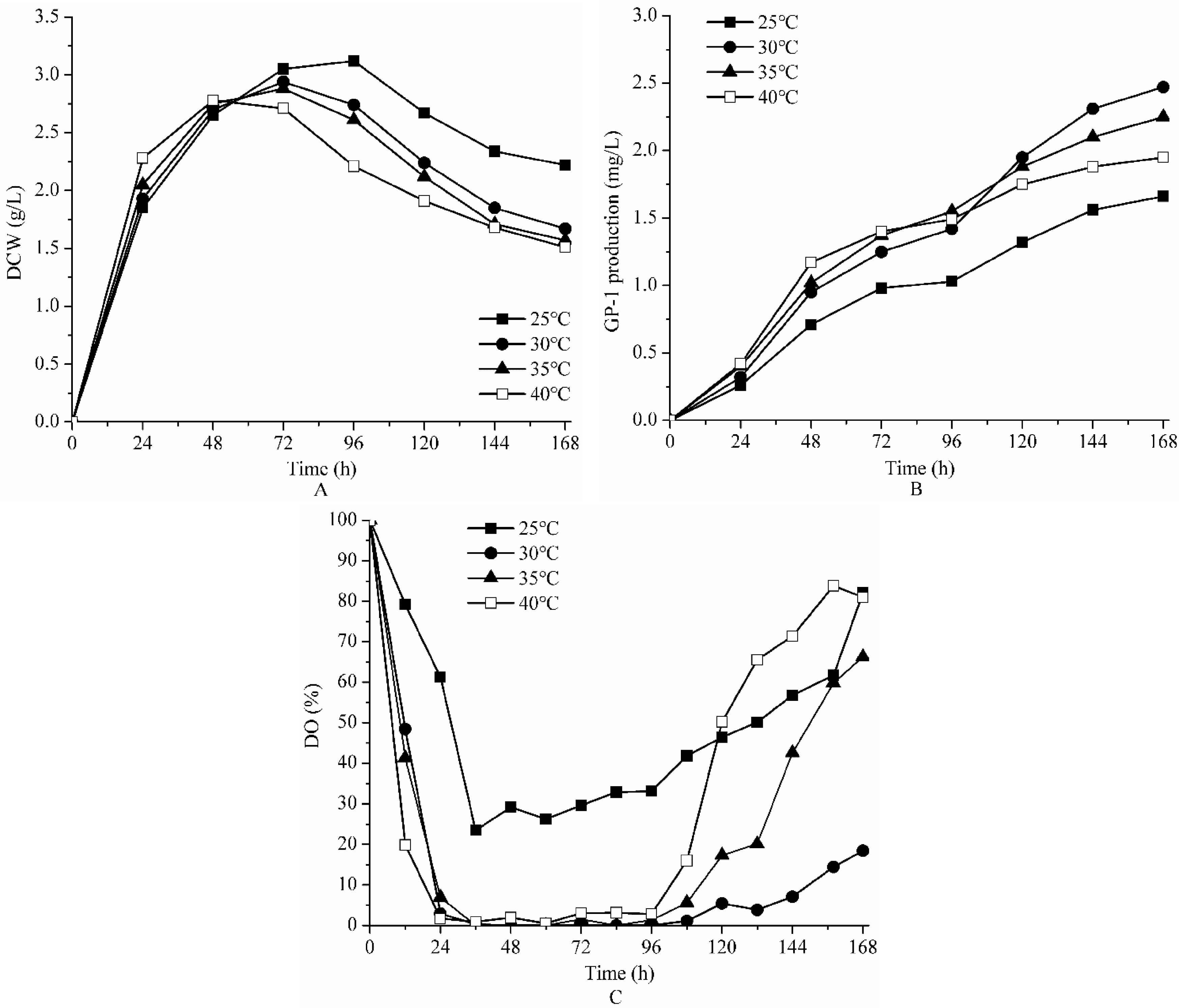
2.1. Effect of Temperature on the Bench Scale Fermentation of Strain ZX01
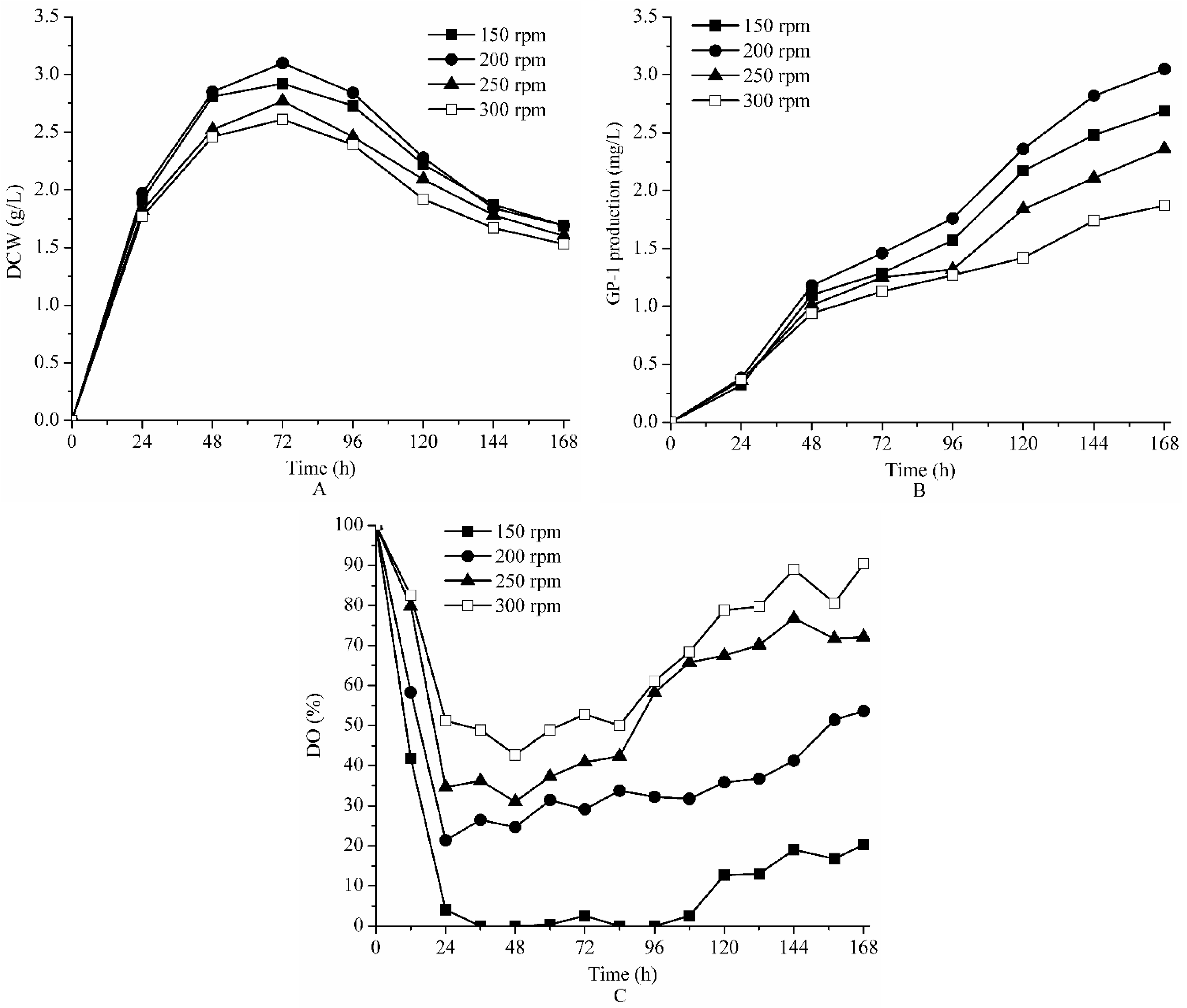
2.2. Effect of Agitation on Fermentation of Strain ZX01 on Bench Scale
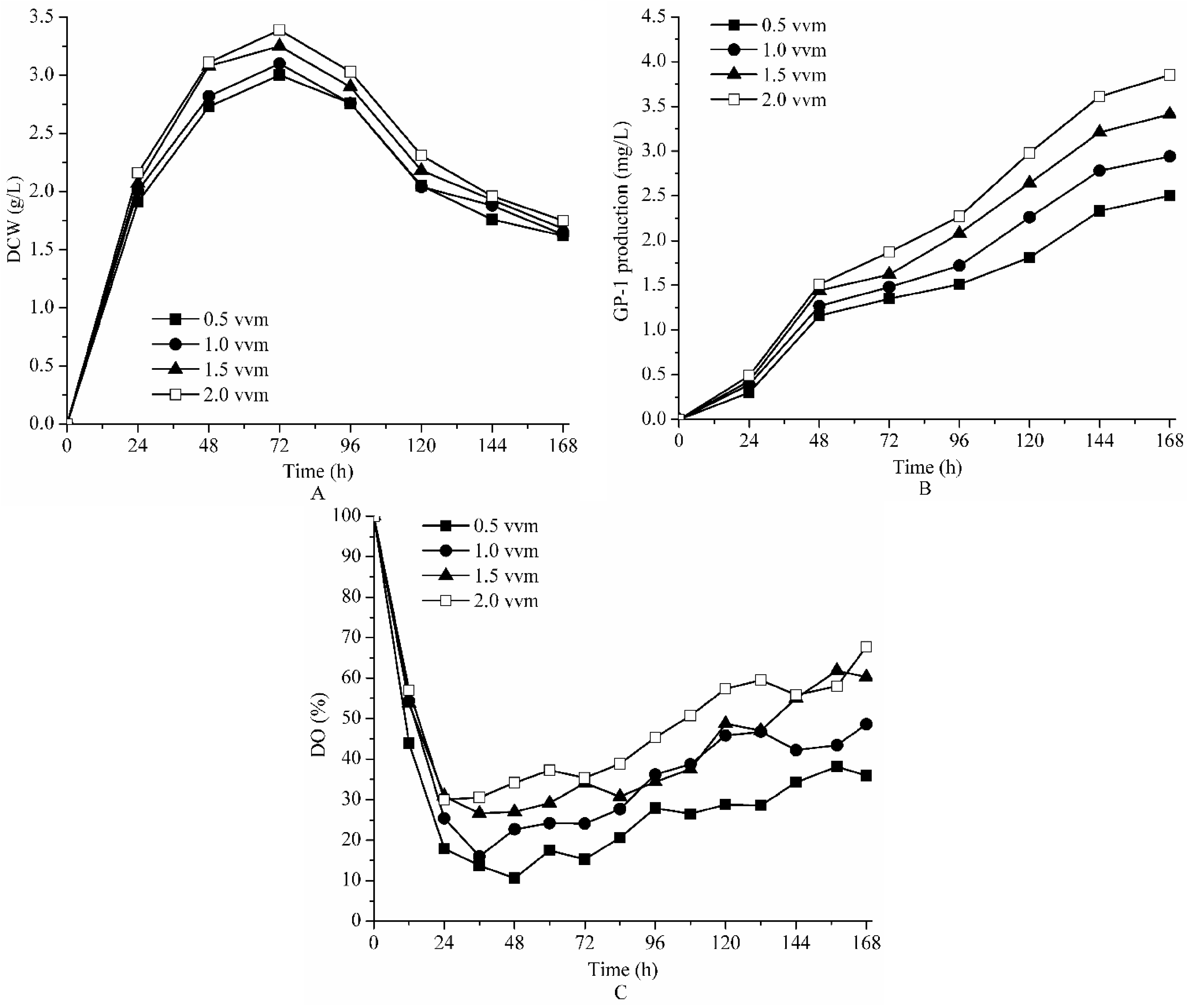
2.3. Effect of Aeration on Fermentation of Strain ZX01 on Bench Scale
2.4. Effects of Agitation and Aeration on kLa on Bench Scale
2.5. Bench Scale Verification Experiments
2.6. Scale-Up Fermentation on Pilot Scale
3. Discussion
4. Materials and Methods
4.1. Microorganism
4.2. Inoculum Preparation and Media
4.3. Fermentors
4.4. Extraction and Determination of Glycoprotein GP-1 Production
4.5. Determination of Dry Cell Weight
4.6. Determination of Volumetric Oxygen Transfer Coefficient
4.7. Effects of Agitation, Aeration and Temperature on GP-1 Production on Bench Scale
4.8. Scale-Up of Fermentation from Bench Scale to Pilot Scale
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Bibb, M.J. Regulation of secondary metabolism in Streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Van Wezel, G.P.; McDowall, K.J. The regulation of the secondary metabolism of Streptomyces: New links and experimental advances. Nat. Prod. Rep. 2011, 28, 1311–1333. [Google Scholar] [CrossRef] [PubMed]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef] [PubMed]
- De Lima Procópio, R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Berdy, J. The discovery of new bioactive microbial metabolites: Screening and identification. Prog. Ind. Microbiol. 1989, 27, 3–27. [Google Scholar]
- Donadio, S.; Monciardini, P.; Alduina, R.; Mazza, P.; Chiocchini, C.; Cavaletti, L.; Sosio, M.; Puglia, A.M. Microbial technologies for the discovery of novel bioactive metabolites. J. Biotechnol. 2002, 99, 187–198. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Agunbiade, M.O.; Green, E.; Mabinya, L.V.; Okoh, A.I. A freshwater Streptomyces, isolated from tyume river, produces a predominantly extracellular glycoprotein bioflocculant. Int. J. Mol. Sci. 2012, 13, 8679–8695. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Xia, F.; Feng, K.; Sun, G.; Gao, X.; Sun, L.; Jiang, R.; Tian, D.; Sun, X. Structural characterization and in vitro antitumor activity of a novel polysaccharide isolated from the fruiting bodies of Pleurotus ostreatus. Bioresour. Technol. 2009, 100, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.C.; Sêmedo, L.T.A.S.; Soares, R.M.A.; Linhares, L.F.; Ulhoa, C.J.; Alviano, C.S.; Coelho, R.R.R. Purification of a thermostable endochitinase from Streptomyces RC1071 isolated from a cerrado soil and its antagonism against phytopathogenic fungi. J. Appl. Microbiol. 2001, 90, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Potumarthi, R.; Ch, S.; Jetty, A. Alkaline protease production by submerged fermentation in stirred tank reactor using Bacillus licheniformis NCIM-2042: Effect of aeration and agitation regimes. Biochem. Eng. J. 2007, 34, 185–192. [Google Scholar] [CrossRef]
- Borges, C.D.; Moreira, A.D.S.; Vendruscolo, C.; Ayub, M.A.Z. Influence of agitation and aeration in xanthan production by Xanthomonas campestris pv pruni strain 101. Rev. Argent. Microbiol. 2008, 40, 81–85. [Google Scholar] [PubMed]
- Giavasis, I.; Harvey, L.M.; McNeil, B. The effect of agitation and aeration on the synthesis and molecular weight of gellan in batch cultures of Sphingomonas paucimobilis. Enzym. Microb. Technol. 2006, 38, 101–108. [Google Scholar] [CrossRef]
- Wecker, A.; Onken, U. Influence of dissolved oxygen concentration and shear rate on the production of pullulan by Aureobasidium pullulans. Biotechnol. Lett. 1991, 13, 155–160. [Google Scholar] [CrossRef]
- Lee, I.; Kim, M.; Lee, J.; Seo, W.; Jung, J.; Lee, H.; Park, Y. Influence of agitation speed on production of curdlan by Agrobacterium species. Bioprocess Biosyst. Eng. 1999, 20, 283–287. [Google Scholar] [CrossRef]
- Bajaj, I.B.; Singhal, R.S. Effect of aeration and agitation on synthesis of poly (γ-glutamic acid) in batch cultures of Bacillus licheniformis NCIM 2324. Biotechnol. Bioprocess Eng. 2010, 15, 635–640. [Google Scholar] [CrossRef]
- Radchenkova, N.; Vassilev, S.; Martinov, M.; Kuncheva, M.; Panchev, I.; Vlaev, S.; Kambourova, M. Optimization of the aeration and agitation speed of Aeribacillus palidus 418 exopolysaccharide production and the emulsifying properties of the product. Process Biochem. 2014, 49, 576–582. [Google Scholar] [CrossRef]
- Kim, S.; Hwang, H.; Xu, C.; Choi, J.; Yun, J. Effect of aeration and agitation on the production of mycelial biomass and exopolysaccharides in an enthomopathogenic fungus Paecilomyces sinclairii. Lett. Appl. Microbiol. 2003, 36, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Mantzouridou, F.; Roukas, T.; Kotzekidou, P. Effect of the aeration rate and agitation speed on β-carotene production and morphology of Blakeslea trispora in a stirred tank reactor: Mathematical modeling. Biochem. Eng. J. 2002, 10, 123–135. [Google Scholar] [CrossRef]
- Qu, L.; Ren, L.-J.; Huang, H. Scale-up of docosahexaenoic acid production in fed-batch fermentation by Schizochytrium sp. based on volumetric oxygen-transfer coefficient. Biochem. Eng. J. 2013, 77, 82–87. [Google Scholar] [CrossRef]
- Schmidt, F. Optimization and scale up of industrial fermentation processes. Appl. Microbiol. Biotechnol. 2005, 68, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Valadez, J.A.; Estrada, M.; Galindo, E.; Serrano-Carreón, L. From shake flasks to stirred fermentors: Scale-up of an extractive fermentation process for 6-pentyl-α-pyrone production by Trichoderma harzianum using volumetric power input. Process Biochem. 2006, 41, 1347–1352. [Google Scholar] [CrossRef]
- Seletzky, J.M.; Noak, U.; Fricke, J.; Welk, E.; Eberhard, W.; Knocke, C.; Büchs, J. Scale-up from shake flasks to fermenters in batch and continuous mode with Corynebacterium glutamicum on lactic acid based on oxygen transfer and pH. Biotechnol. Bioeng. 2007, 98, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Hang, H.-F.; Chu, J.; Zhuang, Y.-P.; Zhang, S.-L. Oxygen uptake rate optimization with nitrogen regulation for erythromycin production and scale-up from 50 L to 372 m3 scale. Bioresour. Technol. 2009, 100, 1406–1412. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor scale-up and oxygen transfer rate in microbial processes: An overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Huong, K.-H.; Azuraini, M.J.; Aziz, N.A.; Amirul, A.-A.A. Pilot scale production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) biopolymers with high molecular weight and elastomeric properties. J. Biosci. Bioeng. 2017, 124, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Galaction, A.-I.; Cascaval, D.; Oniscu, C.; Turnea, M. Prediction of oxygen mass transfer coefficients in stirred bioreactors for bacteria, yeasts and fungus broths. Biochem. Eng. J. 2004, 20, 85–94. [Google Scholar] [CrossRef]
- Tobajas, M.; Garcia-Calvo, E. Comparison of experimental methods for determination of the volumetric mass transfer coefficient in fermentation processes. Heat Mass Transf. 2000, 36, 201–207. [Google Scholar] [CrossRef]
- Poughon, L.; Duchez, D.; Cornet, J.; Dussap, C. kLa determination: Comparative study for a gas mass balance method. Bioprocess Biosyst. Eng. 2003, 25, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Moutafchieva, D.; Popova, D.; Dimitrova, M.; Tchaoushev, S. Experimental determination of the volumetric mass transfer coefficient. J. Chem. Technol. Metall. 2013, 48, 351–356. [Google Scholar]
- Tribe, L.; Briens, C.; Margaritis, A. Determination of the volumetric mass transfer coefficient (kLa) using the dynamic “gas out-gas in” method: Analysis of errors caused by dissolved oxygen probes. Biotechnol. Bioeng. 1995, 46, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, G.; Miao, G.; Zhang, X.; Feng, J. Streptomyces kanasensis sp. nov., an antiviral glycoprotein producing actinomycete isolated from forest soil around kanas lake of China. Curr. Microbiol. 2015, 71, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Han, L.; Zhang, G.; Zhang, X.; Feng, J. Purification and characterization of a novel glycoprotein from Streptomyces sp. ZX01. Int. J. Biol. Macromol. 2015, 78, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Feng, J.; Han, L.; Zhang, X. Antiviral activity of glycoprotein GP-1 isolated from Streptomyces kanasensis ZX01. Int. J. Biol. Macromol. 2016, 88, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Sun, Y.-B.; He, H.-W.; Feng, J.-T.; Zhang, X.; Han, L.-R. Optimization of medium compositions to improve a novel glycoprotein production by Streptomyces kanasenisi ZX01. AMB Express 2017, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Bandaiphet, C.; Prasertsan, P. Effect of aeration and agitation rates and scale-up on oxygen transfer coefficient, kLa in exopolysaccharide production from Enterobacter cloacae WD7. Carbohydr. Polym. 2006, 66, 216–228. [Google Scholar] [CrossRef]
- Redón, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of growth temperature on yeast lipid composition and alcoholic fermentation at low temperature. Eur. Food Res. Technol. 2011, 232, 517–527. [Google Scholar] [CrossRef]
- Abdel-Banat, B.M.; Hoshida, H.; Ano, A.; Nonklang, S.; Akada, R. High-temperature fermentation: How can processes for ethanol production at high temperatures become superior to the traditional process using mesophilic yeast? Appl. Microbiol. Biotechnol. 2010, 85, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Andorrà, I.; Landi, S.; Mas, A.; Esteve-Zarzoso, B.; Guillamón, J.M. Effect of fermentation temperature on microbial population evolution using culture-independent and dependent techniques. Food Res. Int. 2010, 43, 773–779. [Google Scholar] [CrossRef]
- Kshirsagar, P.R.; Suttar, R.; Nilegaonkar, S.S.; Pradhan, S.; Kanekar, P.P. Scale up production of polyhydroxyalkanoate (PHA) at different aeration, agitation and controlled dissolved oxygen levels in fermenter using Halomonas campisalis mcm b-1027. J. Biochem. Technol. 2013, 4, 512–517. [Google Scholar]
- Kamble, A.; Meena, V.; Banerjee, U. Effect of agitation and aeration on the production of nitrile hydratase by Rhodococcus erythropolis MTCC 1526 in a stirred tank reactor. Lett. Appl. Microbiol. 2010, 51, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Xia, J.-Y.; Chu, J.; Zhuang, Y.-P.; Zhang, S.-L. Real-time fluid dynamics investigation and physiological response for erythromycin fermentation scale-up from 50 L to 132 m3 fermenter. Bioprocess Biosyst. Eng. 2012, 35, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Kanda, M.; Yamamoto, E.; Hayashi, A.; Yabutani, T.; Yamashita, M.; Honda, H. Scale-up fermentation of echinocandin type antibiotic FR901379. J. Biosci. Bioeng. 2010, 109, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Albaek, M.O.; Gernaey, K.V.; Hansen, M.S.; Stocks, S.M. Modeling enzyme production with Aspergillus oryzae in pilot scale vessels with different agitation, aeration, and agitator types. Biotechnol. Bioeng. 2011, 108, 1828–1840. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Yuan, G.; Zhao, Y.; Ni, W.; Luo, H.; Shi, Z.; Liu, F. Simulation of computational fluid dynamics and comparison of cephalosporin C fermentation performance with different impeller combinations. Korean J. Chem. Eng. 2013, 30, 1097–1104. [Google Scholar] [CrossRef]
- Xia, J.-Y.; Wang, Y.-H.; Zhang, S.-L.; Chen, N.; Yin, P.; Zhuang, Y.-P.; Chu, J. Fluid dynamics investigation of variant impeller combinations by simulation and fermentation experiment. Biochem. Eng. J. 2009, 43, 252–260. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compound Glycoprotein GP-1 is available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Parameters | The Final GP-1 Production (mg/L) | The Maximum DCW (g/L) | kLa (h−1) | |
|---|---|---|---|---|
| Temperature (°C) | 25 | 1.32 | 3.15 | - |
| 30 | 2.47 | 2.94 | - | |
| 35 | 2.20 | 2.85 | - | |
| 40 | 1.95 | 2.78 | - | |
| Agitation (rpm) | 150 | 2.59 | 3.02 | 14.53 |
| 200 | 3.05 | 3.12 | 18.23 | |
| 250 | 2.16 | 2.67 | 27.21 | |
| 300 | 1.87 | 2.61 | 32.82 | |
| Aeration (vvm) | 0.5 | 2.50 | 3.00 | 13.21 |
| 1.0 | 2.96 | 3.10 | 16.70 | |
| 1.5 | 3.41 | 3.25 | 18.91 | |
| 2.0 | 3.86 | 3.39 | 22.43 |
| Fermentor | Fermentation Condition | The Final GP-1 Production (mg/L) | The Maximum DCW (g/L) | kLa (h−1) |
|---|---|---|---|---|
| 5 L | 200 rpm, 2.0 vvm | 3.89 | 3.55 | 22.83 |
| 4.05 | 3.42 | 21.31 | ||
| 3.90 | 3.41 | 21.57 | ||
| 3.83 | 3.45 | 20.78 | ||
| Average | 3.92 | 3.46 | 21.62 |
| Fementor | Agitation (rpm) | The Final GP-1 Production (mg/L) | The Maximum DCW (g/L) | kLa (h−1) |
|---|---|---|---|---|
| 15 L | 200 | 3.75 | 3.55 | 19.15 |
| 225 | 4.03 | 3.54 | 22.52 | |
| 250 | 3.92 | 3.40 | 25.74 | |
| 275 | 3.57 | 3.20 | 29.51 | |
| 300 | 3.20 | 3.14 | 31.43 | |
| 70 L | 200 | 3.55 | 3.60 | 17.53 |
| 225 | 3.75 | 3.60 | 20.32 | |
| 250 | 3.89 | 3.76 | 23.11 | |
| 275 | 3.50 | 3.58 | 26.18 | |
| 300 | 3.16 | 3.32 | 29.78 | |
| 500 L | 200 | 3.81 | 3.77 | 15.04 |
| 225 | 3.96 | 3.81 | 18.38 | |
| 250 | 4.13 | 4.01 | 22.31 | |
| 275 | 4.00 | 3.85 | 27.04 | |
| 300 | 3.80 | 3.66 | 32.47 |
| Fermentors | Fermentation Condition | The Final GP-1 Production (mg/L) | The Maximum DCW (g/L) | kLa (h−1) |
|---|---|---|---|---|
| 15 L | 225 rpm, 2.0 vvm | 3.98 | 3.52 | 23.63 |
| 4.10 | 3.55 | 23.71 | ||
| 4.10 | 3.61 | 22.72 | ||
| 3.94 | 3.45 | 21.14 | ||
| Average | 4.03 | 3.53 | 22.80 | |
| 70 L | 250 rpm, 2.0 vvm | 3.81 | 3.71 | 23.42 |
| 3.93 | 3.76 | 24.27 | ||
| 3.78 | 3.65 | 23.51 | ||
| 3.77 | 3.63 | 21.59 | ||
| Average | 3.82 | 3.69 | 23.20 | |
| 500 L | 250 rpm, 2.0 vvm | 4.23 | 3.93 | 22.20 |
| 4.28 | 3.90 | 23.28 | ||
| 4.09 | 4.06 | 21.81 | ||
| 4.20 | 3.91 | 21.68 | ||
| Average | 4.20 | 3.95 | 22.24 |
| Fermentor | Bench-Scale | Pilot-Scale | ||
|---|---|---|---|---|
| Total volume (L) | 5 | 15 | 70 | 500 |
| Working volume (L) | 3.5 | 10 | 50 | 350 |
| Diameter of fermentor (m) | 0.15 | 0.25 | 0.38 | 0.75 |
| Diameter of impeller (m) | 0.07 | 0.10 | 0.15 | 0.30 |
| Height of fermentor (m) | 0.30 | 0.50 | 0.75 | 1.50 |
| Baffle | 3 | 4 | 4 | 6 |
| Impeller | Two impellers with four-flat-blade | Two impellers with six-flat-blade | ||
| Type of drive | Magnetic stirred | Mechanical stirred | ||
| Sterilization | Ex-situ | In-situ | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Han, L.-R.; He, H.-W.; Sang, B.; Yu, D.-L.; Feng, J.-T.; Zhang, X. Effects of Agitation, Aeration and Temperature on Production of a Novel Glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and Scale-Up Based on Volumetric Oxygen Transfer Coefficient. Molecules 2018, 23, 125. https://doi.org/10.3390/molecules23010125
Zhou Y, Han L-R, He H-W, Sang B, Yu D-L, Feng J-T, Zhang X. Effects of Agitation, Aeration and Temperature on Production of a Novel Glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and Scale-Up Based on Volumetric Oxygen Transfer Coefficient. Molecules. 2018; 23(1):125. https://doi.org/10.3390/molecules23010125
Chicago/Turabian StyleZhou, Yong, Li-Rong Han, Hong-Wei He, Bu Sang, Dai-Lin Yu, Jun-Tao Feng, and Xing Zhang. 2018. "Effects of Agitation, Aeration and Temperature on Production of a Novel Glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and Scale-Up Based on Volumetric Oxygen Transfer Coefficient" Molecules 23, no. 1: 125. https://doi.org/10.3390/molecules23010125
APA StyleZhou, Y., Han, L. -R., He, H. -W., Sang, B., Yu, D. -L., Feng, J. -T., & Zhang, X. (2018). Effects of Agitation, Aeration and Temperature on Production of a Novel Glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and Scale-Up Based on Volumetric Oxygen Transfer Coefficient. Molecules, 23(1), 125. https://doi.org/10.3390/molecules23010125