
HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
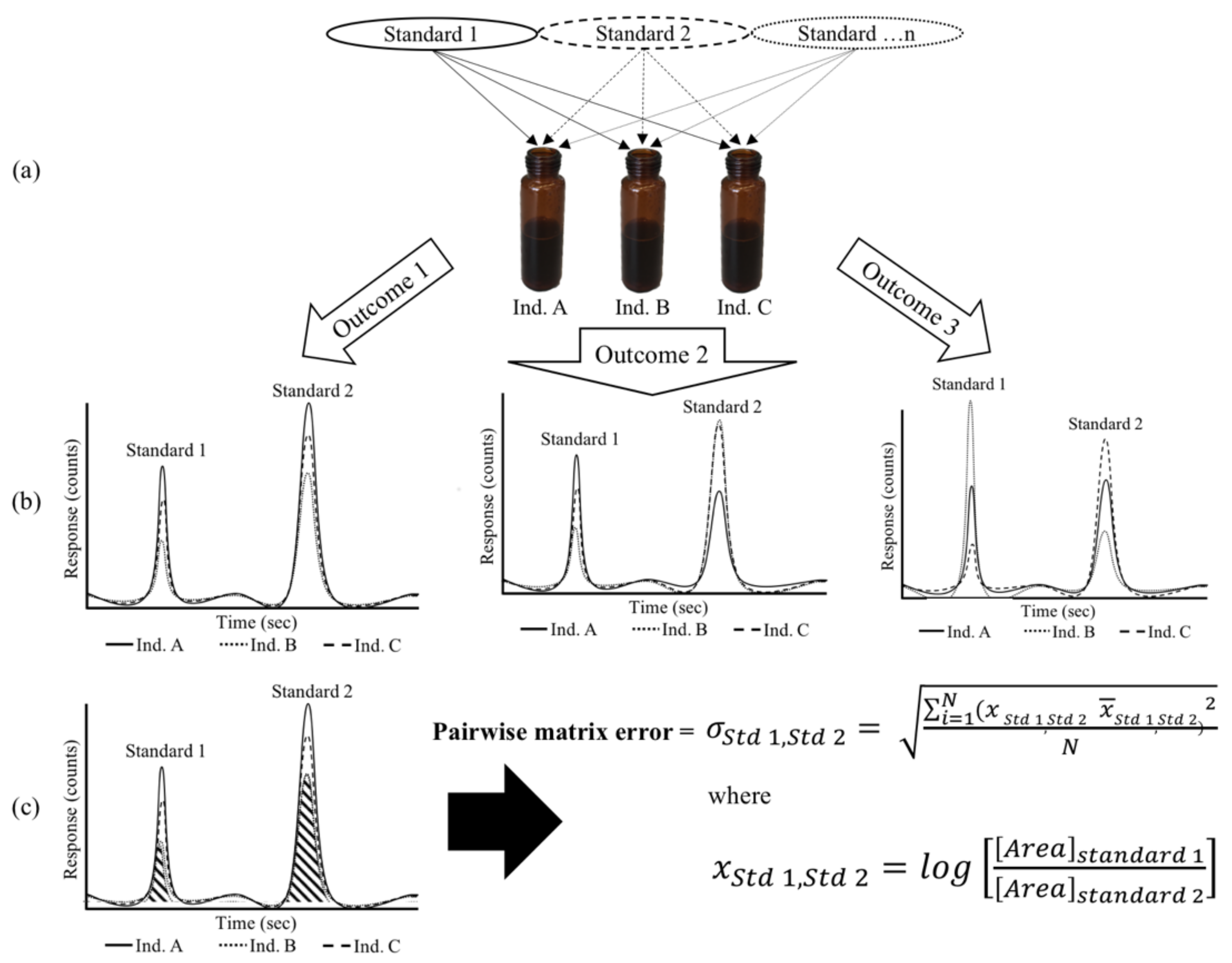
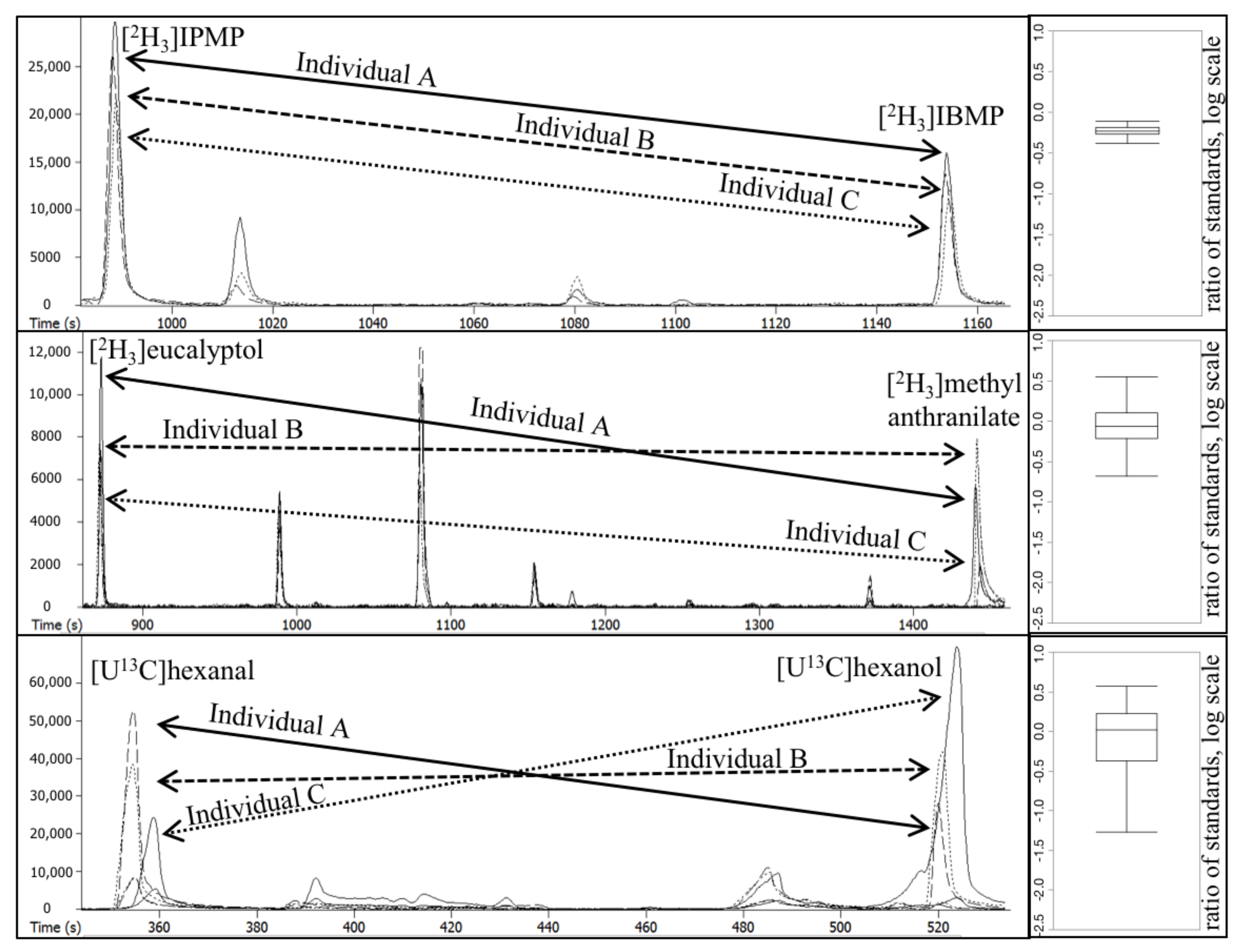
2.1. General Approach to Estimating Compound × Sample Interactions () in Plant Populations
- Samples are spiked with a cocktail of internal standards prior to SPME-GC-MS analysis (Figure 1). In the present work, as we were studying plant populations, these standards were either isotopic analogues of plant-derived odorants, or non-labeled surrogate standards previously reported for use in plant volatile profiling.
- Pairwise matrix error () is calculated as described in the Methods section and Figure 1, where the Compound × Individual interaction is assessed across a population for each internal standard, and quantified by the value.
2.2. Quantitating Compound × Individual Matrix Effects in a Grape Population
2.3. Quantifying Compound × Individual Matrix Effects in a Tomato RIL Population
2.4. Consequences of Compound × Individual Matrix Effects within Plant Populations
2.5. Conclusions
3. Materials and Methods
3.1. Chemical Reagents and Standards
3.2. Sample Collection of Grapes and Tomatoes
3.3. Sample Preparation of Grapes and Tomatoes
3.4. Analysis of Grape Volatiles by HS-SPME-GC-MS
3.5. Analysis of Tomato Volatiles by HS-SPME-GC-MS
3.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rambla, J.L.; Trapero-Mozos, A.; Diretto, G.; Rubio-Moraga, A.; Granell, A.; Gómez-Gómez, L.; Ahrazem, O. Gene-Metabolite Networks of Volatile Metabolism in Airen and Tempranillo Grape Cultivars Revealed a Distinct Mechanism of Aroma Bouquet Production. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, J.; Xu, Y.; Liang, J.; Chang, P.; Yan, F.; Li, M.; Liang, Y.; Zou, Z. Genome-Wide Association Mapping for Tomato Volatiles Positively Contributing to Tomato Flavor. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Slegers, A.; Angers, P.; Ouellet, É.; Truchon, T.; Pedneault, K. Volatile compounds from grape skin, juice and wine from five interspecific hybrid grape cultivars grown in Québec (Canada) for wine production. Molecules 2015, 20, 10980–11016. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.-W.; Su, M.-Q.; Wu, H.-X.; Zhou, Y.-G.; Wang, S.-B. Analysis of the Volatile Profile of Core Chinese Mango Germplasm by Headspace Solid-Phase Microextraction Coupled with Gas Chromatography-Mass Spectrometry. Molecules 2018, 23, 1480. [Google Scholar] [CrossRef] [PubMed]
- Marsili, R. Flavor, Fragrance, and Odor Analysis, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Van Nocker, S.; Gardiner, S.E. Breeding better cultivars, faster: applications of new technologies for the rapid deployment of superior horticultural tree crops. Hortic. Res. 2014, 1, 14022. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fresnedo-Ramírez, J.; Wang, M.; Cote, L.; Schweitzer, P.; Barba, P.; Takacs, E.M.; Clark, M.; Luby, J.; Manns, D.C.; et al. A next-generation marker genotyping platform (AmpSeq) in heterozygous crops: A case study for marker-assisted selection in grapevine. Hortic. Res. 2016, 3, 16002. [Google Scholar] [CrossRef] [PubMed]
- Chaparro-Torres, L.A.; Bueso, M.C.; Fernández-Trujillo, J.P. Aroma volatiles obtained at harvest by HS-SPME/GC-MS and INDEX/MS-E-nose fingerprint discriminate climacteric behaviour in melon fruit. J. Sci. Food Agric. 2016, 96, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Obando-Ulloa, J.M.; Ruiz, J.; Monforte, A.J.; Fernández-Trujillo, J.P. Aroma profile of a collection of near-isogenic lines of melon (Cucumis melo L.). Food Chem. 2010, 118, 815–822. [Google Scholar] [CrossRef]
- Dunemann, F.; Ulrich, D.; Boudichevskaia, A.; Grafe, C.; Weber, W.E. QTL mapping of aroma compounds analysed by headspace solid-phase microextraction gas chromatography in the apple progeny “Discovery” × “Prima”. Mol. Breed. 2009, 23, 501–521. [Google Scholar] [CrossRef]
- Vogt, J.; Schiller, D.; Ulrich, D.; Schwab, W.; Dunemann, F. Identification of lipoxygenase (LOX) genes putatively involved in fruit flavour formation in apple (Malus × domestica). Tree Genet. Genomes 2013, 9, 1493–1511. [Google Scholar] [CrossRef]
- Battilana, J.; Costantini, L.; Emanuelli, F.; Sevini, F.; Segala, C.; Moser, S.; Velasco, R.; Versini, G.; Grando, M.S. The 1-deoxy-d-xylulose 5-phosphate synthase gene co-localizes with a major QTL affecting monoterpene content in grapevine. Theor. Appl. Genet. 2009, 118, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Doligez, A.; Audiot, E.; Baumes, R.; This, P. QTLs for muscat flavor and monoterpenic odorant content in grapevine (Vitis vinifera L.). Mol. Breed. 2006, 18, 109–125. [Google Scholar] [CrossRef]
- Guillaumie, S.; Ilg, A.; Réty, S.; Brette, M.; Trossat-Magnin, C.; Decroocq, S.; Léon, C.; Keime, C.; Ye, T.; Baltenweck-Guyot, R.; et al. Genetic Analysis of the Biosynthesis of 2-Methoxy-3-Isobutylpyrazine, a Major Grape-Derived Aroma Compound Impacting Wine Quality. Source Plant Physiol. 2013, 162, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Bezman, Y.; Mayer, F.; Takeoka, G.R.; Buttery, R.G.; Ben-oliel, G.; Rabinowitch, H.D.; Naim, M. Differential effects of tomato (Lycopersicon esculentum Mill) matrix on the volatility of important aroma compounds. J. Agric. Food Chem. 2003, 51, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Vandendriessche, T.; Nicolai, B.M.; Hertog, M.L.A.T.M. Optimization of HS SPME Fast GC-MS for High-Throughput Analysis of Strawberry Aroma. Food Anal. Methods 2013, 6, 512–520. [Google Scholar] [CrossRef]
- García-Vico, L.; Belaj, A.; Sánchez-Ortiz, A.; Martínez-Rivas, J.M.; Pérez, A.G.; Sanz, C. Volatile Compound Profiling by HS-SPME/GC-MS-FID of a Core Olive Cultivar Collection as a Tool for Aroma Improvement of Virgin Olive Oil. Molecules 2017, 22, 141. [Google Scholar] [CrossRef] [PubMed]
- Souza-Silva, É.A.; Gionfriddo, E.; Pawliszyn, J. A critical review of the state of the art of solid-phase microextraction of complex matrices II. Food analysis. TrAC–Trends Anal. Chem. 2015, 71, 236–248. [Google Scholar] [CrossRef]
- Lloyd, N.; Johnson, D.L.; Herderich, M.J. Metabolomics approaches for resolving and harnessing chemical diversity in grapes, yeast and wine. Aust. J. Grape Wine Res. 2015, 21, 723–740. [Google Scholar] [CrossRef]
- Schieberle, P.; Molyneux, R.J. Quantitation of Sensory-Active and Bioactive Constituents of Food: A Journal of Agricultural and Food Chemistry Perspective. J. Agric. Food Chem. 2012, 60, 2404–2408. [Google Scholar] [CrossRef] [PubMed]
- Hopfer, H.; Jodari, F.; Negre-Zakharov, F.; Wylie, P.L.; Ebeler, S.E. HS-SPME-GC-MS/MS Method for the Rapid and Sensitive Quantitation of 2-Acetyl-1-pyrroline in Single Rice Kernels. J. Agric. Food Chem. 2016, 64, 4114–4120. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Cortés, P.; Brenna, J.T.; Sacks, G.L. Production of isotopically labeled standards from a uniformly labeled precursor for quantitative volatile metabolomic studies. Anal. Chem. 2012, 84, 5400–5406. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Natera, R.; Benitez, P.; Barroso, C.G. Comparative analysis of volatile compounds of “fino” sherry wine by rotatory and continuous liquid-liquid extraction and solid-phase microextraction in conjunction with gas chromatography-mass spectrometry. Anal. Chim. Acta 2004, 513, 141–150. [Google Scholar] [CrossRef]
- Klee, H.J.; Tieman, D.M. Genetic challenges of flavor improvement in tomato. Trends Genet. 2013, 29, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Herrero, P.; Zapata, J.; Escudero, A. Coping with matrix effects in headspace solid phase microextraction gas chromatography using multivariate calibration strategies. J. Chromatogr. A 2015, 1407, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Badenes, M.L.; Byrne, D.H. Fruit breeding; Springer Science & Business Media: Berlin, Germany, 2012; ISBN 9781441907639. [Google Scholar]
- Hyma, K.E.; Barba, P.; Wang, M.; Londo, J.P.; Acharya, C.B.; Mitchell, S.E.; Sun, Q.; Reisch, B.; Cadle-Davidson, L. Heterozygous mapping strategy (HetMappS) for high resolution genotyping-by-sequencing markers: A case study in grapevine. PLoS ONE 2015, 10, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Gonda, I.; Ashrafi, H.; Lyon, D.A.; Strickler, S.R.; Hulse-Kemp, A.M.; Ma, Q.; Sun, H.; Stoffel, K.; Powell, A.F.; Futrell, S.; et al. A GBS-based high-density genetic map of a tomato RIL population, facilitating high-resolution QTL mapping and candidate gene identification. Plant Genome 2018, in press. [Google Scholar]
- Galpaz, N.; Gonda, I.; Shem-Tov, D.; Barad, O.; Tzuri, G.; Lev, S.; Fei, Z.; Xu, Y.; Mao, L.; Jiao, C.; et al. Deciphering genetic factors that determine melon fruit-quality traits using RNA-Seq-based high-resolution QTL and eQTL mapping. Plant J. 2018, 94, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Canuti, V.; Conversano, M.; Calzi, M.L.; Heymann, H.; Matthews, M.A.; Ebeler, S.E. Headspace solid-phase microextraction-gas chromatography-mass spectrometry for profiling free volatile compounds in Cabernet Sauvignon grapes and wines. J. Chromatogr. A 2009, 1216, 3012–3022. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, F.J.; Aguilar, I.; Zamora, R. Model Studies on the Effect of Aldehyde Structure on Their Selective Trapping by Phenolic Compounds. J. Agric. Food Chem. 2017, 65, 4736–4743. [Google Scholar] [CrossRef] [PubMed]
- Ryona, I.; Pan, B.S.; Sacks, G.L. Rapid measurement of 3-Alkyl-2-methoxypyrazine content of winegrapes to predict levels in resultant wines. J. Agric. Food Chem. 2009, 57, 8250–8257. [Google Scholar] [CrossRef] [PubMed]
- Aronson, J.; Ebeler, S.E. Effect of polyphenol compounds on the headspace volatility of flavors. Am. J. Enol. Vitic. 2004, 55, 13–21. [Google Scholar]
- Dunlevy, J.; Kalua, C.; Keyzers, R.; Boss, P. The Production of Flavour & Aroma Compounds in Grape Berries. In Grapevine Molecular Physiology & Biotechnology; Roubelakis-Angelakis, K.A., Ed.; Springer: Dordrecht, The Netherland, 2009; pp. 293–340. ISBN 978-90-481-2304-9. [Google Scholar]
- Souza-Silva, E.A.; Jiang, R.; Rodriguez-Lafuente, A.; Gionfriddo, E.; Pawliszyn, J. A critical review of the state of the art of solid-phase microextraction of complex matrices I. Environmental analysis. TrAC–Trends Anal. Chem. 2015, 71, 224–235. [Google Scholar] [CrossRef]
- Ashrafi, H.; Kinkade, M.; Foolad, M.R. A new genetic linkage map of tomato based on a Solanum lycopersicum × S. pimpinellifolium RIL population displaying locations of candidate pathogen response genes. Genome 2009, 52, 935–956. [Google Scholar] [CrossRef] [PubMed]
- Tikunov, Y. A Novel Approach for Nontargeted Data Analysis for Metabolomics. Large-Scale Profiling of Tomato Fruit Volatiles. Plant Physiol. 2005, 139, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of fruit from the grapevine population are currently available if advance arrangements are made. Samples of fruit from the tomato population are not available. |




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burzynski-Chang, E.A.; Ryona, I.; Reisch, B.I.; Gonda, I.; Foolad, M.R.; Giovannoni, J.J.; Sacks, G.L. HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects. Molecules 2018, 23, 2436. https://doi.org/10.3390/molecules23102436
Burzynski-Chang EA, Ryona I, Reisch BI, Gonda I, Foolad MR, Giovannoni JJ, Sacks GL. HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects. Molecules. 2018; 23(10):2436. https://doi.org/10.3390/molecules23102436
Chicago/Turabian StyleBurzynski-Chang, Elizabeth A., Imelda Ryona, Bruce I. Reisch, Itay Gonda, Majid R. Foolad, James J. Giovannoni, and Gavin L. Sacks. 2018. "HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects" Molecules 23, no. 10: 2436. https://doi.org/10.3390/molecules23102436
APA StyleBurzynski-Chang, E. A., Ryona, I., Reisch, B. I., Gonda, I., Foolad, M. R., Giovannoni, J. J., & Sacks, G. L. (2018). HS-SPME-GC-MS Analyses of Volatiles in Plant Populations—Quantitating Compound × Individual Matrix Effects. Molecules, 23(10), 2436. https://doi.org/10.3390/molecules23102436