
Carbon (δ13C) and Nitrogen (δ15N) Stable Isotope Composition Provide New Insights into Phenotypic Plasticity in Broad Leaf Weed Rumex acetosa under Allelochemical Stress




Abstract
:
1. Introduction
2. Material and Methods
2.1. Plant Growth Conditions and Treatments
2.2. Carbon Isotope Discrimination and Mass Spectrometry Analysis
2.3. Relative Water Content and Leaf Osmotic Potential
2.4. Harvesting and Plant Growth Bioassays
2.5. Statistical Analysis
3. Results
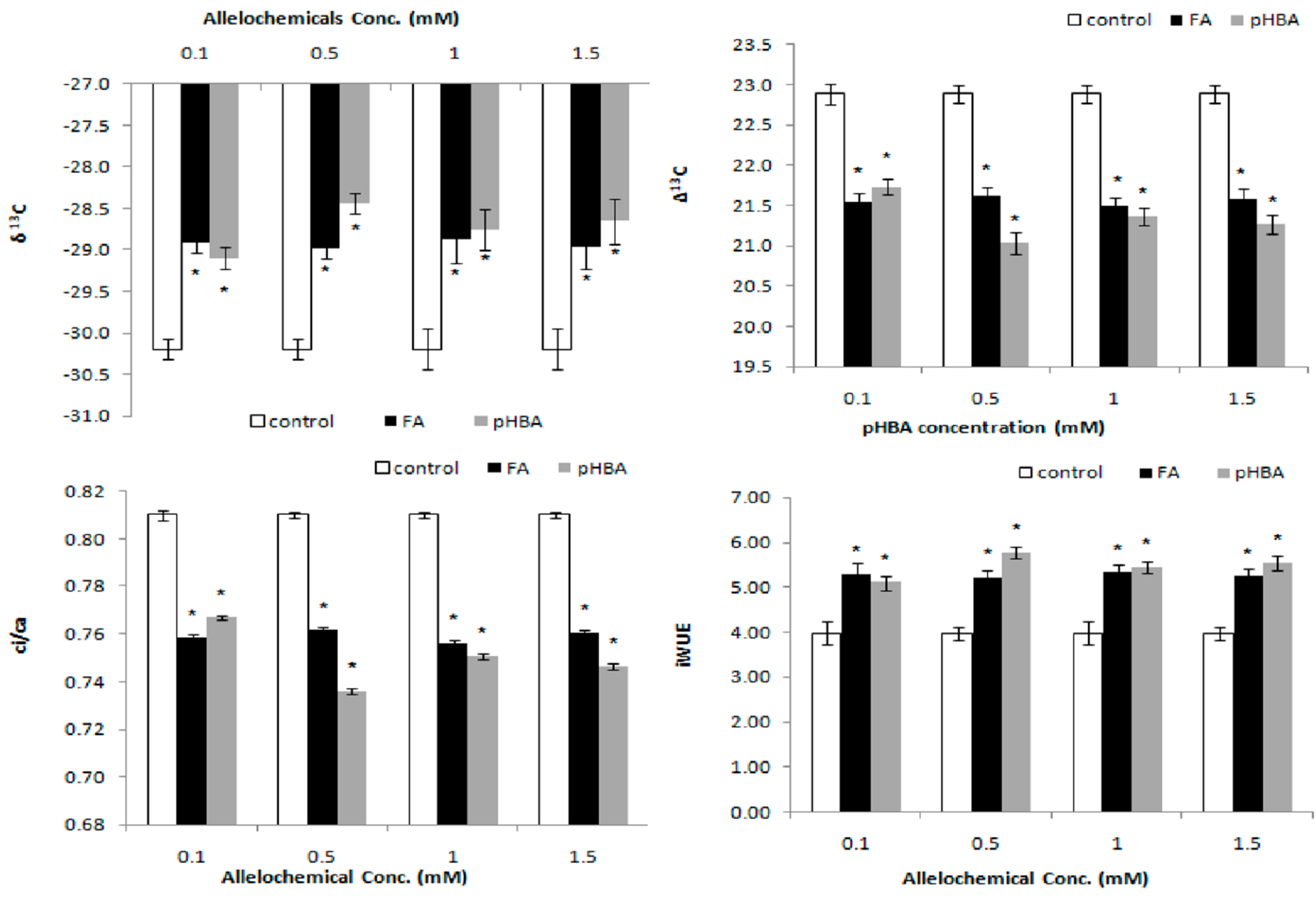
3.1. Effects of Phenolic Acids on Photosynthetic Carbon Isotope Discrimination
3.2. Carbon and Nitrogen Concentrations and Δ15n Isotope Composition
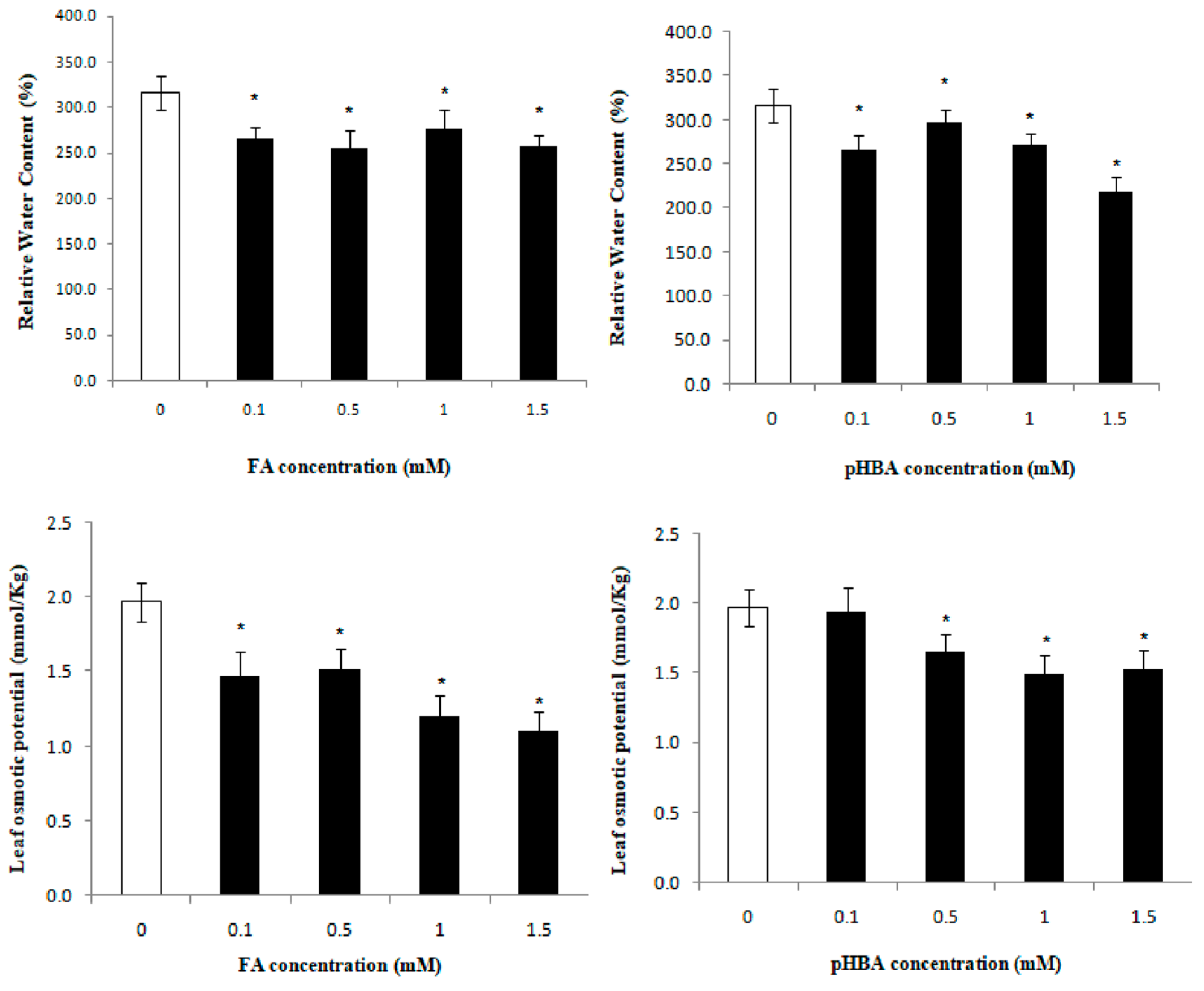
3.3. Elucidation of Inhibitory Effects of Phenolic Acids on Leaf Water Relations
3.4. Unrevealing the Inhibitory Effects of Phenolic Acids in Planta
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| iWUE | intrinsic water use efficiency |
| LOP | leaf osmotic potential |
| RWC | relative water content |
| δ13C | composition of carbon isotope ratios |
| Δ13C | carbon isotope discrimination |
| Ci/Ca | ratio of intercellular CO2 concentration from leaf to air |
References
- Busi, R.; Vila-Aiub, M.M.; Beckie, H.J.; Gaines, T.A.; Goggin, D.E.; Kaundun, S.S.; Lacoste, M.; Neve, P.; Nissen, S.J.; Norsworthy, J.K.; et al. Herbicide-resistant weeds: From research and knowledge to future needs. Evol. Appl. 2013, 6, 1218–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delye, C. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to herbicides: A major challenge for weed science in the forthcoming decade. Pest. Manag. Sci. 2013, 69, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources of herbicides: Current status and future trends. Weed Res. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Jasieniuk, M.; Brûlé-Babel, A.L.; Morrison, I.N. The evolution and genetics of herbicide resistance in weeds. Weed Sci. 1996, 44, 176–193. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.I.; Reigosa, M.J. Allelochemical stress inhibits growth, leaf water relations, PSII photochemistry, non-photochemical fluorescence quenching and heat energy dissipation in three C3 perennial species. J. Exp. Bot. 2011, 62, 4533–4545. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Owen, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest. Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Massalha, H.; Korenblum, E.; Tholl, D.; Aharoni, A. Small molecules below-ground: The role of specialized metabolites in the rhizosphere. Plant J. 2017, 90, 788–807. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K.; Hutzler, J.; Tresch, S.; Christiansen, N.; Looser, R.; Ehrhardt, T. On the mode of action of the herbicides cinmethylin and 5-benzyloxymethyl-1,2-isoxazolines: Putative inhibitors of plant tyrosine aminotransferase. Pest. Manag. Sci. 2012, 68, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Holzwarth, A.R.; Muller, M.G.; Reus, M.; Nowaczyk, M.; Sander, J.; Rogner, M. Kinetics and mechanism of electron transfer in intact photostystem II and the isolated reaction center: Pheophytin is the primary electron acceptor. Proc. Natl. Acad. Sci. USA 2006, 103, 6895–6900. [Google Scholar] [CrossRef] [PubMed]
- Doblinski, P.M.F.; Ferrarese, M.L.L.; Huber, D.A.; Scapim, C.A.; Braccini, A.L.; Ferrarese-Filho, O. Peroxidase and lipid peroxidation of soybean roots in response to p-coumaric and p-hydroxybenzoic acids. Braz. Arch. Biol. Technol. 2003, 46, 193–198. [Google Scholar] [CrossRef]
- Ai Hamdi, B.; Inderjit; Olofsdotter, M.; Streibig, J.C. Laboratory bioassay for phytotoxicity: An example from wheat straw. Agron. J. 2001, 93, 43–48. [Google Scholar]
- Pérez, F.J.; Ormeño-Nuñez, J. Root exudates of wild oats: Allelopathic effect on spring wheat. Phytochemistry 1991, 30, 2199–2202. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. Seedling growth, leaf water status and signature of stable carbon isotopes in C3 perennials exposed to natural phytochemicals. Aust. J. Bot. 2012, 60, 676–684. [Google Scholar] [CrossRef]
- Hussain, M.I.; González, L.; Reigosa, M.J. Allelopathic potential of Acacia melanoxylon R. Br. on the germination and root growth of native species. Weed Biol. Manag. 2011, 11, 18–28. [Google Scholar] [CrossRef]
- Hussain, M.I.; González, L.; Souto, C.; Reigosa, M.J. Ecophysiological responses of native plants to phytotoxic effect of Acacia melanoxylon R. Br. Agrofor. Syst. 2011, 83, 149–166. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. Evaluation of photosynthetic performance and carbon isotope discrimination in perennial ryegrass (Lolium perenne L.) under allelochemicals stress. Ecotoxicology 2017, 26, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Castellano, D.; Simonet, A.M.; Molinillo, J.M. Structure−activity relationship (SAR) studies of Benzoxazinones, their degradation products, and analogues. Phytotoxicity on problematic weeds Avena fatua L. and Lolium rigidum Gaud. J. Agric. Food Chem. 2006, 54, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Ann. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Funct. Plant Biol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- CDIAC. Monthly Atmospheric 13C/12C Isotopic Ratios for 11 SIO Stations; Carbon Dioxide Information Analysis Center (CDIAC): Oak Ridge, TN, USA, 2015. [Google Scholar]
- Robertson, A.; Overpeck, J.; Rind, D.; Mosley-Thompson, E.; Zielinski, G.; Lean, J.; Koch, D.; Penner, J.; Tegen, I.; Healy, R. Hypothesized climate forcing time series for the last 500 years. J. Geophys. Res. Atmos. 2001, 106, 14783–14803. [Google Scholar] [CrossRef] [Green Version]
- Chandramohan, S.; Charudattan, R.; Sonoda, R.M.; Singh, M. Field evaluation of a fungal pathogen mixture for the control of seven weed grasses. Weed Sci. 2002, 50, 204–213. [Google Scholar] [CrossRef]
- Macías, F.A.; Oliveros-Bastidas, A.; Marín, D.; Chinchilla, N.; Castellano, D.; Molinillo, J.M.G. Evidence for an allelopathic interaction between rye and wild oats. J. Agric. Food Chem. 2014, 62, 9450–9457. [Google Scholar] [CrossRef] [PubMed]
- DellaGreca, M.; Fiorentino, A.; Izzo, A.; Napoli, F.; Purcaro, R. Phytotoxicity of secondary metabolites from Aptenia cordifolia. Chem. Biodivers. 2007, 4, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Cheema, Z.A. Weed Control in Wheat through Sorghum Allelochemicals. Ph.D. Thesis, University of Agriculture, Faisalabad, Pakistan, 1988. [Google Scholar]
- Vyvyan, W.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1632–1646. [Google Scholar] [CrossRef]
- Hubick, K.; Farquhar, G. Carbon isotope discrimination and the ratio of carbon gained to water lost in barley cultivars. Plant. Cell. Environ. 1989, 12, 795–804. [Google Scholar] [CrossRef]
- Frank, A.B.; Berdahl, J.D. Gas exchange and water relations in diploid and tetraploid Russian wildrye. Crop. Sci. 2001, 41, 87–92. [Google Scholar] [CrossRef]
- Zhao, B.; Kondo, M.; Maeda, M.; Ozaki, Y.; Zhang, J. Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes. Plant Soil 2004, 261, 61–75. [Google Scholar] [CrossRef]
- Erice, G.; Louahlia, S.; Irigoyen, J.J.; Sánchez-Díaz, M.; Alami, I.T.; Avice, J.C. Water use efficiency, transpiration and net CO2 exchange of four alfalfa genotypes submitted to progressive drought and subsequent recovery. Environ. Exp. Bot. 2011, 72, 123–130. [Google Scholar] [CrossRef]
- Li, C. Carbon isotope composition, water-use efficiency and biomass productivity of Eucalyptus microtheca populations under different water supplies. Plant Soil 1999, 214, 165–171. [Google Scholar] [CrossRef]
- Virgona, J.M.; Farquhar, D.G. Genotypic variation in relative growth rate and carbon isotope discrimination in sunflower is related to photosynthetic capacity. Funct. Plant. Biol. 1996, 23, 227–236. [Google Scholar] [CrossRef]
- Barkosky, R.R.; Einhellig, F.A. Allelopathic interference of plant-water relationships by para-hydroxybenzoic acid. Bot. Bull. Acad. Sin. 2003, 44, 53–58. [Google Scholar]
- Vaughan, D.; Ord, B.G. Influence of phenolic acids on morphological changes in roots of Pisum sativum. J. Sci. Food Agric. 2006, 52, 289–299. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Sorgonà, A.; Muscolo, A. Morphophysiological changes in tissue culture of Petunia hybridain response to the allelochemical coumarin. Allelop. J. 2001, 8, 171–178. [Google Scholar]
- Anjum, T.; Bajwa, R. Field appraisal of herbicide potential of sunflower leaf extract against Rumex dentatus. Field Crops Res. 2007, 100, 139–142. [Google Scholar] [CrossRef]
- Macías, F.A.; Galindo, J.C.G.; Molinillo, J.M.G.; Castellano, D. Dehydrozaluzanin C: A potent plant growth regulator with potential use as a natural herbicide template. Phytochemistry 2000, 54, 165–171. [Google Scholar] [CrossRef]
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Expt. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, S.M.; Su, J.H.; Zho, L.L.; Ba, X.G.; Zhan, H.G.; Zhang, F.S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. USA 2007, 104, 11192–11196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, L.L.; Robinson, D.; Forster, B.P.; Ellis, R.P.; Scrimgeour, C.M.; Gordon, D.C.; Nero, E.; Raven, J.A. Shoot δ15N correlates with genotype and salt stress in barley. Planta 1997, 201, 100–102. [Google Scholar] [CrossRef]
- Robinson, D.; Handley, L.L.; Scrimgeour, C.M.; Gordon, D.C.; Forster, B.P.; Ellis, R.P. Using stable isotope natural abundances (δ15N and δ13C) to integrate the stress responses of wild barley (Hordeum spontaneum C. Koch.) genotypes. J. Exp. Bot. 2000, 51, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.; Araus, J.L. Nitrogen source and water regime effects on durum wheat photosynthesis, and stable carbon and nitrogen isotope composition. Physiol. Plant. 2006, 126, 435–445. [Google Scholar] [CrossRef]
- Evans, R.D. Physiological mechanism influencing plant nitrogen isotope composition. Trends Plant. Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Gronle, A.; Heb, J.; Böhm, H. Weed suppressive ability in sole and intercrops of pea and oat and its interaction with ploughing and crop interference in organic farming. Org. Agric. 2015, 5, 39–51. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Treatments | Concentration (mM) | C% | N% | C/N ratio | δ15N |
|---|---|---|---|---|---|
| Control | 48.14 ± 1.9 a | 1.97 ± 0.31 a | 24.43 ± 0.6 b | 15.02 ± 0.4 a | |
| FA | 0.1 | 46.02 ± 5.5 b | 1.90 ± 0.39 a | 24.22 ± 0.4 b | 11.09 ± 3.5 b |
| 0.5 | 46.37 ± 1.9 b | 1.52 ±0.4 c | 30.5 ± 0.4 a | 10.46 ± 1.1 c | |
| 1 | 43.66 ± 2.5 c | 1.82 ±0.63 b | 23.98 ± 0.5 c | 11.59 ± 0.9 b | |
| 1.5 | 44.28 ± 1.2 c | 1.47 ± 0.32 c | 30.12 ± 0.6 a | 10.77 ± 0.9 c | |
| Control | 48.14 ± 1.9 a | 1.97 ± 0.31 a | 24.43 ± 0.6 c | 15.02 ± 0.4 a | |
| pHBA | 0.1 | 44.88 ± 3.5 b | 1.95 ± 0.68 a | 23.01 ± 0.43 c | 11.56 ± 1.3 b |
| 0.5 | 43.96 ± 0.4 c | 1.65 ± 0.36 b | 26.64 ± 0.6 b | 10.21 ± 0.7 c | |
| 1 | 43.34 ± 4.2 c | 1.53 ± 0.38 c | 28.32 ± 0.4 a | 10.58 ± 1.3 c | |
| 1.5 | 43.61 ± 3.2 c | 1.40 ± 0.40 d | 31.15 ± 0.5 a | 9.94 ± 1.0 d |
| Treatments | Concentration (mM) | LFW | LDW | RFW | RDW | SL | RL |
|---|---|---|---|---|---|---|---|
| Control | Control | 2.39 a | 0.33 a | 3.05 a | 0.98 a | 15.32 a | 19.18 a |
| FA | 0.1 | 1.11 c | 0.27 b | 1.99 c | 0.633 b | 11.83 c | 16 b |
| 0.5 | 1.01 c | 0.21 b | 1.13 c | 0.71 b | 12.16 b | 14.33 c | |
| 1 | 0.99 c | 0.12b c | 2.62 b | 0.816 b | 10.56 c | 17 b | |
| 1.5 | 1.23 b | 0.11 b | 3.11 a | 0.396 c | 12.73 b | 14.5 c | |
| pHBA | 0.1 | 0.55 c | 0.09 c | 2.12 a | 0.793 a | 12 b | 14.83 b |
| 0.5 | 0.77 b | 0.16 b | 2.01 a | 0.646 b | 12.5 b | 14.33 b | |
| 1 | 0.93 a | 0.18 a | 1.78 b | 0.606 b | 12.26 b | 19 a | |
| 1.5 | 0.98 a | 0.14 c | 1.6 b | 0.506 c | 13a | 12.76 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.I.; Reigosa, M.J.; Muscolo, A. Carbon (δ13C) and Nitrogen (δ15N) Stable Isotope Composition Provide New Insights into Phenotypic Plasticity in Broad Leaf Weed Rumex acetosa under Allelochemical Stress. Molecules 2018, 23, 2449. https://doi.org/10.3390/molecules23102449
Hussain MI, Reigosa MJ, Muscolo A. Carbon (δ13C) and Nitrogen (δ15N) Stable Isotope Composition Provide New Insights into Phenotypic Plasticity in Broad Leaf Weed Rumex acetosa under Allelochemical Stress. Molecules. 2018; 23(10):2449. https://doi.org/10.3390/molecules23102449
Chicago/Turabian StyleHussain, M. Iftikhar, Manuel J. Reigosa, and Adele Muscolo. 2018. "Carbon (δ13C) and Nitrogen (δ15N) Stable Isotope Composition Provide New Insights into Phenotypic Plasticity in Broad Leaf Weed Rumex acetosa under Allelochemical Stress" Molecules 23, no. 10: 2449. https://doi.org/10.3390/molecules23102449
APA StyleHussain, M. I., Reigosa, M. J., & Muscolo, A. (2018). Carbon (δ13C) and Nitrogen (δ15N) Stable Isotope Composition Provide New Insights into Phenotypic Plasticity in Broad Leaf Weed Rumex acetosa under Allelochemical Stress. Molecules, 23(10), 2449. https://doi.org/10.3390/molecules23102449