
Isolation of Tricin as a Xanthine Oxidase Inhibitor from Sweet White Clover (Melilotus albus) and Its Distribution in Selected Gramineae Species


Abstract
:1. Introduction
2. Results and Discussion
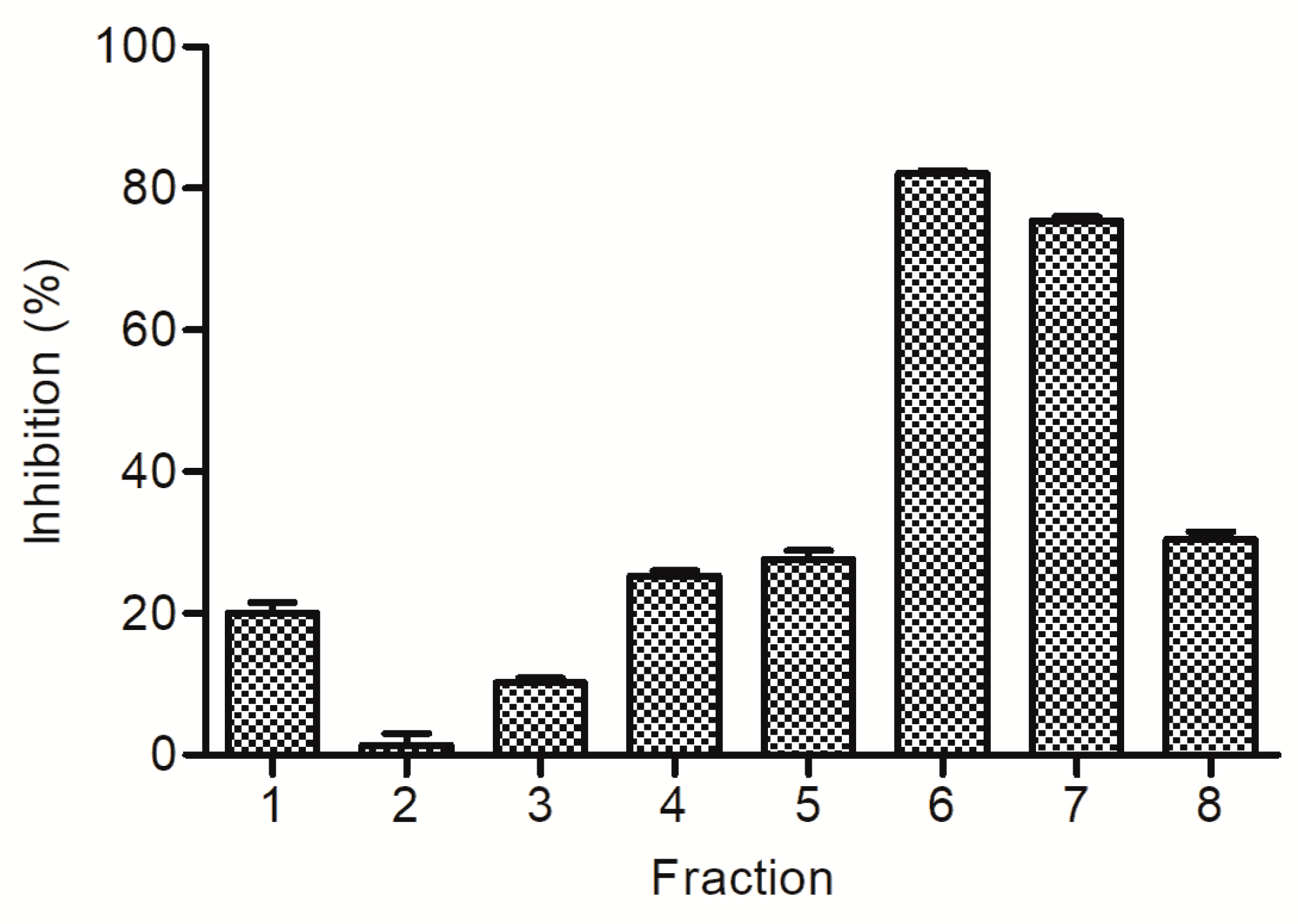
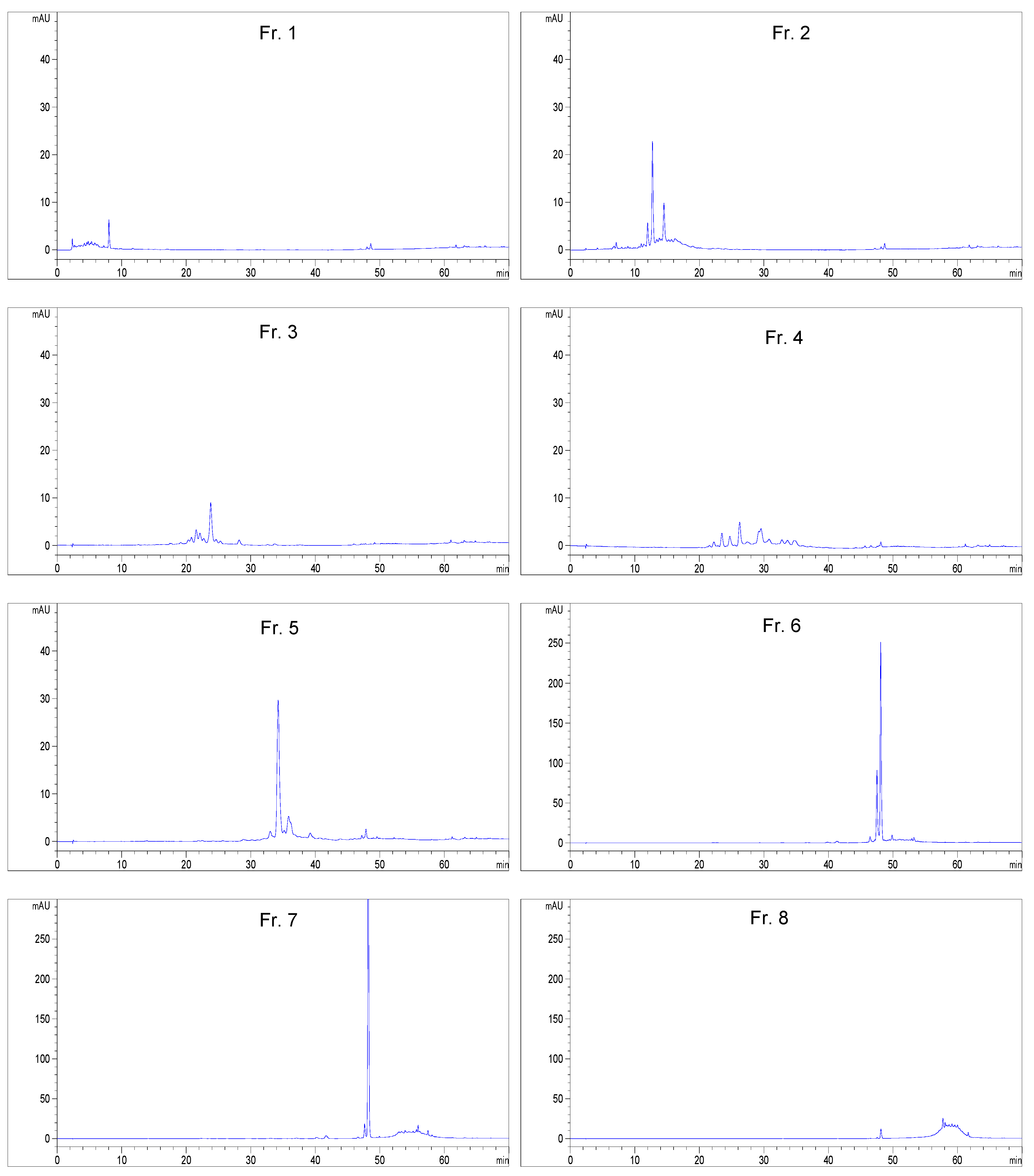
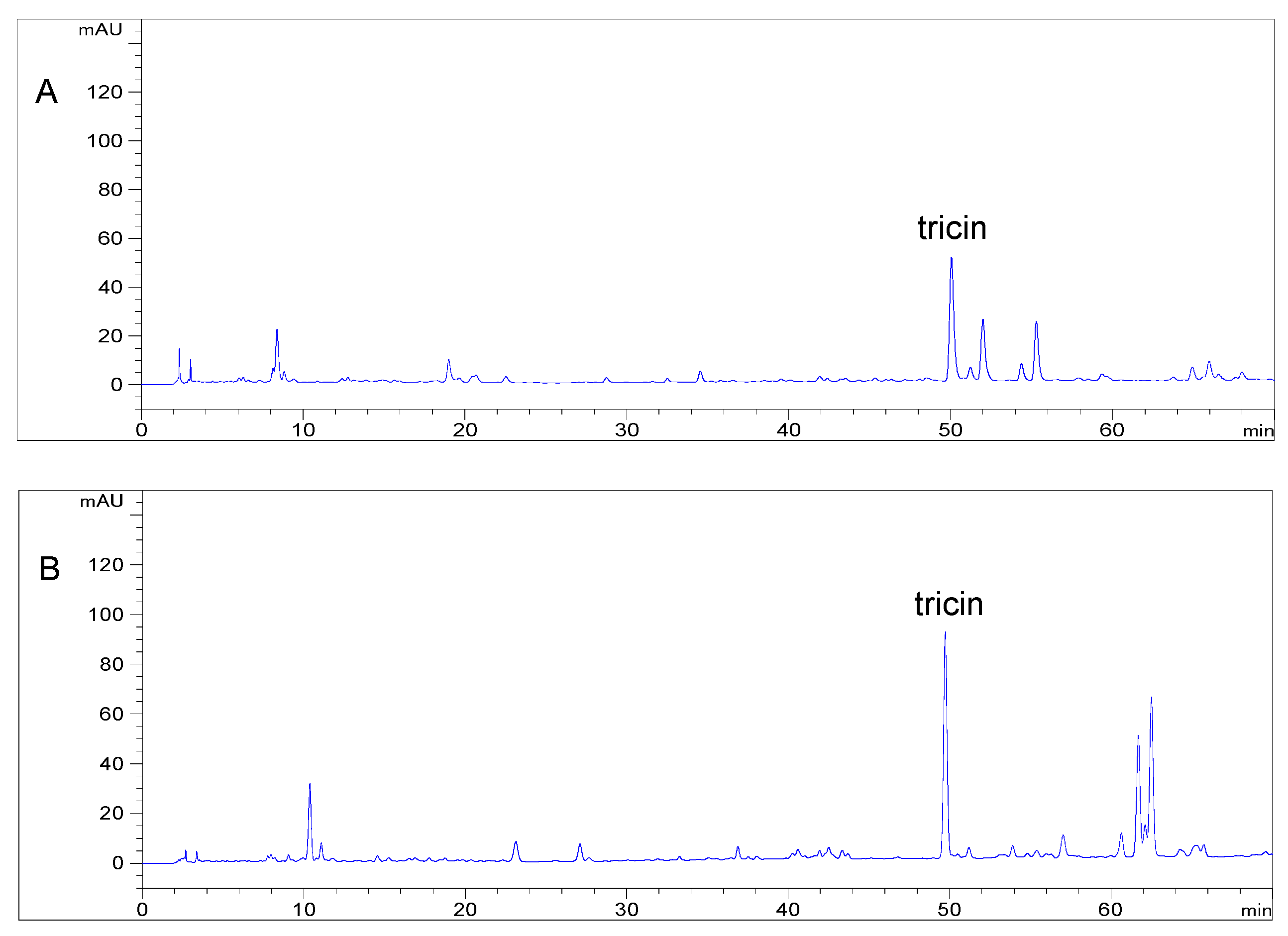
2.1. Bioassay and High Performance Liquid Chromatography Guided Isolation of Tricin
2.2. Distribution of Tricin in Different Tissues of Selected Gramineae Species
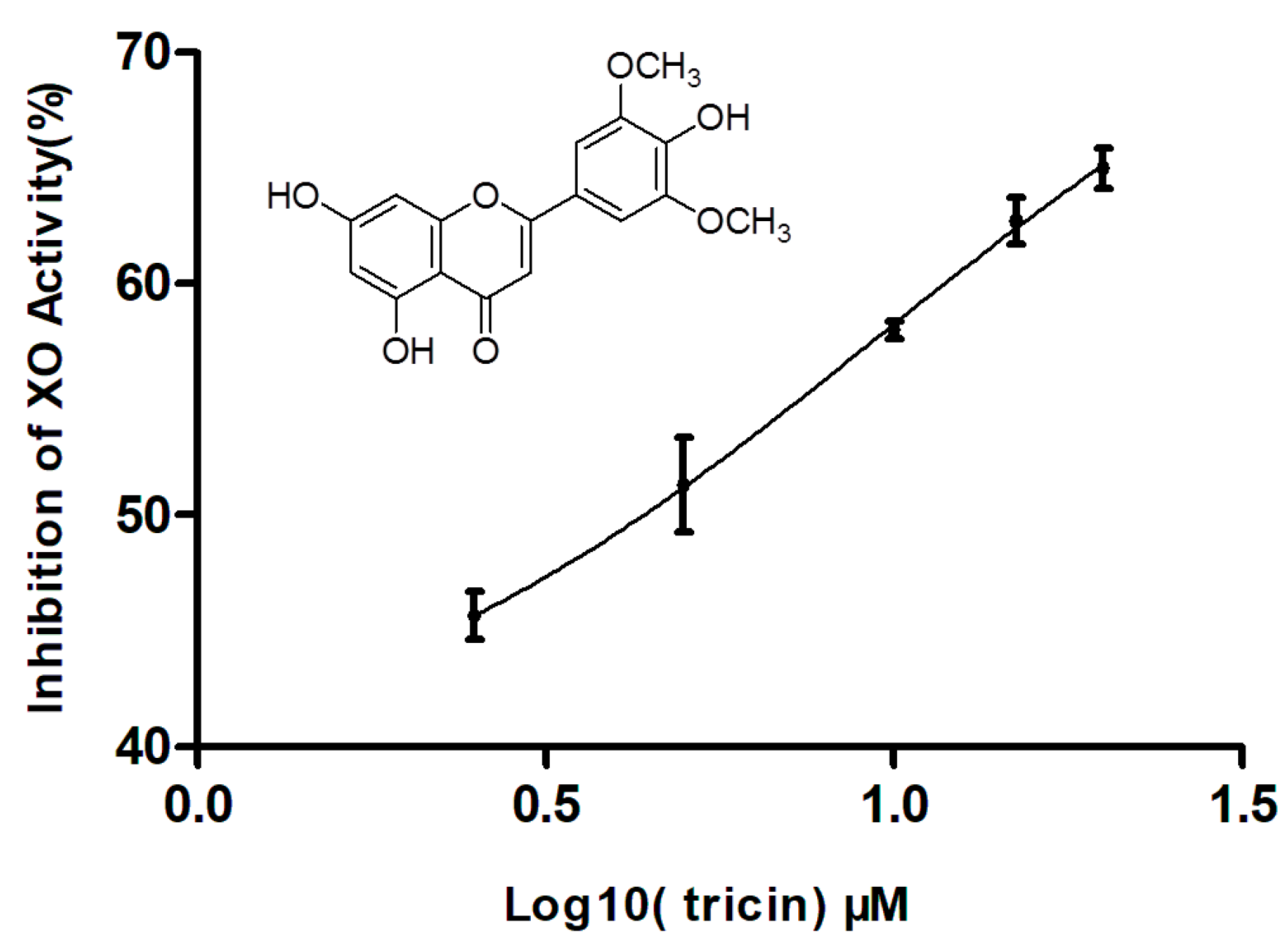
2.3. Effect of Tricin on XO Activity
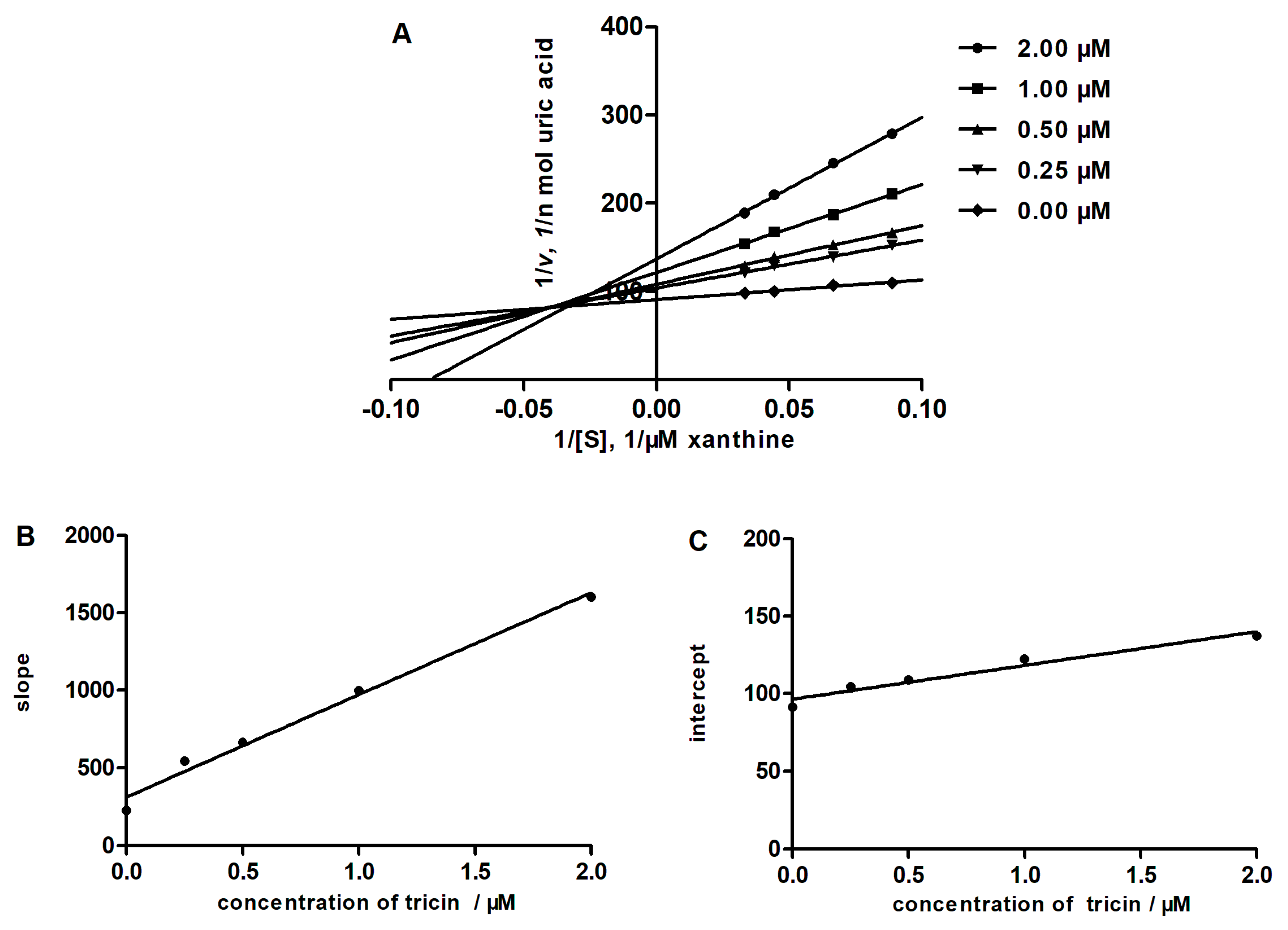
2.4. Determining the Type of Inhibition by Lineweaver-Burk Analysis
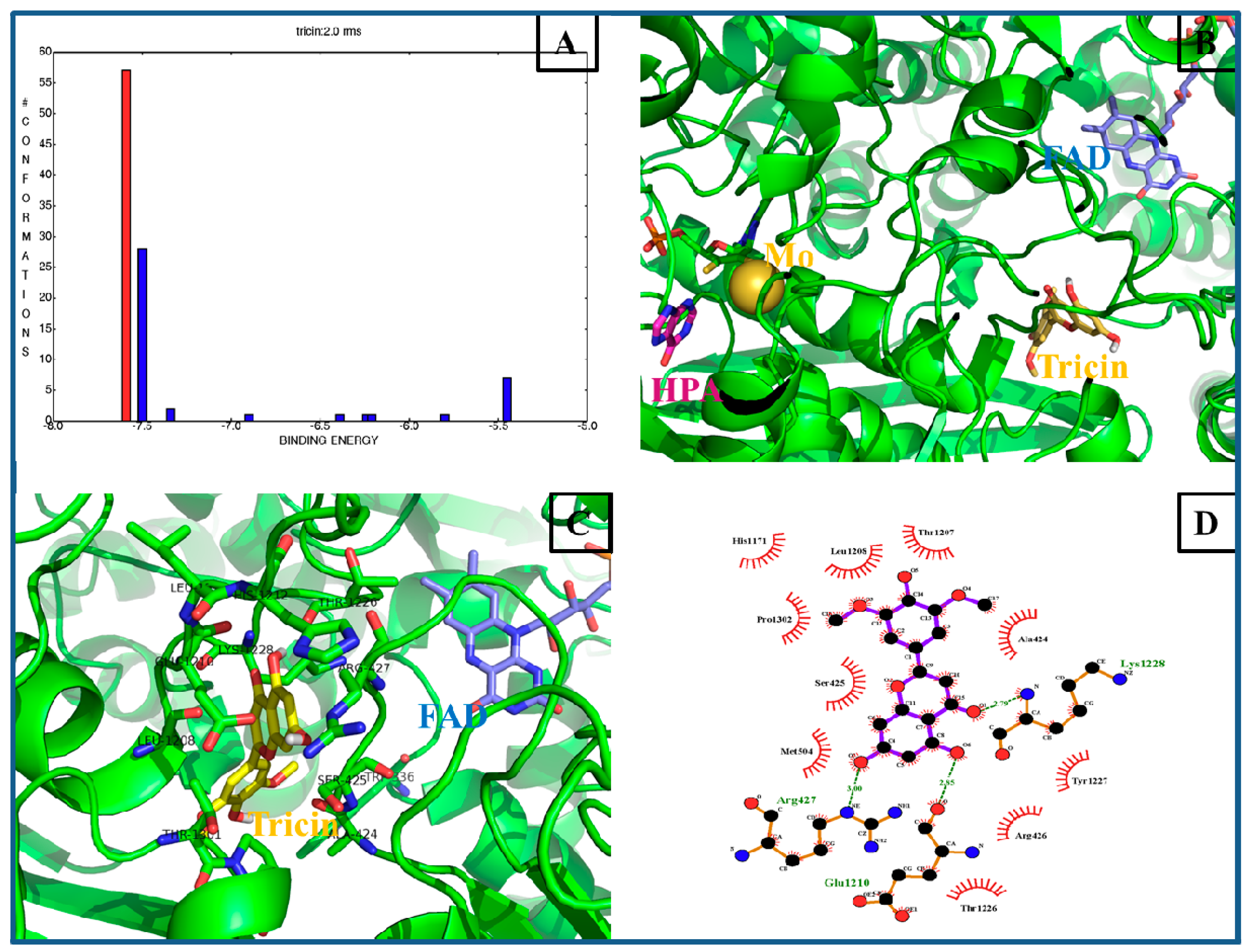
2.5. Computational Docking Analysis
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Chemicals
3.3. Plant Material
3.4. Extraction and Isolation
3.5. HPLC Analysis of the Fractions
3.6. XO Inhibitory Activity Assay and XO Inhibitory Modes of Action Assay
3.7. Quantitative Determination of Tricin
3.8. Docking Studies
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Parvez, M.K. Natural or plant products for the treatment of neurological disorders: Current knowledge. Curr. Drug Metab. 2018, 19, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Hou, Y.; Sun, W.; Yu, J.; Liu, X.; Niu, Y.; Lu, J.J.; Chen, X. Natural products to prevent drug resistance in cancer chemotherapy: A review. Ann. N. Y. Acad. Sci. 2017, 1401, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, B.; Halabalaki, M.; Schwaiger, S.; Adamopoulos, N.; Allouche, N.; Fiebich, B.L.; Hermans, N.; Jansen-Dürr, P.; Kesternich, V.; Pieters, L.; et al. Novel natural products for healthy ageing from the Mediterranean diet and food plants of other global sources-The MediHealth Project. Molecules 2018, 23, 1097. [Google Scholar] [CrossRef] [PubMed]
- Stefanović, O.D.; Tešić, J.D.; Čomić, L.R. Melilotus albus and Dorycnium herbaceum extracts as source of phenolic compounds and their antimicrobial, antibiofilm, and antioxidant potentials. J. Food Drug Anal. 2015, 23, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Zabala, J.M.; Marinoni, L.; Giavedoni, J.A.; Schrauf, G.E. Breeding strategies in Melilotus albus Desr., a salt-tolerant forge legume. Euphytica 2018, 214, 22. [Google Scholar] [CrossRef]
- Kuo, C.F.; Grainge, M.J.; Zhang, W.; Doherty, M. Global epidemiology of gout: Prevalence, incidence and risk factors. Nat. Rev. Rheumatol. 2015, 11, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Bardin, T.; Richette, P. Impact of comorbidities on gout and hyperuricaemia: An update on prevalence and treatment options. BMC Med. 2017, 15, 123. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lü, J.M.; Yao, Q.Z. Hyperuricemia-related diseases and xanthine oxidoreductase (XOR) inhibitors: An overview. Med. Sci. Monit. 2016, 22, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Wang, S.W.; Zhu, M.Q.; Chen, J.Z.; Zhu, X.X. Effect of genistein, apigenin, quercetin, rutin and astilbin on serum uric acid levels and xanthine oxidase activities in normal and hperuricemic mice. Food Chem. Toxicol. 2011, 49, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Fujishima, M.A.T.; da Silva, N.D.R.; Ramos, R.D.; Ferreira, E.F.B.; dos Santos, K.L.B.; da Silva, C.H.T.D.; da Silva, J.O.; Rosa, J.M.C.; dos Santos, C.B.R. An antioxidant potential, quantum-chemical and molecular docking study of the major chemical constituents present in the leaves of Curatella americana Linn. Pharmaceuticals 2018, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Moheb, A.; Grondin, M.; Ibrahim, R.K.; Roy, R.; Sarhan, F. Winter wheat hull (husk) is a valuable source for tricin, a potential selective cytotoxic agent. Food Chem. 2013, 138, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.J.; Zhang, Y.; Liu, C.M.; Liu, J.E.; Wu, X.Q.; Zhang, Y. Spearation and purification of tricin from an antioxidant product derived from bamboo leaves. J. Agric. Food Chem. 2007, 55, 10008–10092. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pu, Y.Q.; Yoo, C.G.; Ragauskas, A.J. The occurrence of tricin and its derivatives in plants. Green Chem. 2016, 18, 1439–1454. [Google Scholar] [CrossRef]
- Nørbæk, R.; Aaboer, D.B.F.; Bleeg, I.S.; Christensen, B.T.; Kondo, T.; Brandt, K. Flavone C-glycoside, phenolic acid, and nitrogen contents in leaves of barley subject to organic fertilization treatments. J. Agric. Food Chem. 2003, 51, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.K.; Xu, J.K.; Zhang, L.; Du, G.H. Flavonoids from the bran of Avena sativa. Chin. J. Nat. Med. 2012, 10, 110–114. [Google Scholar] [CrossRef]
- Mohanlal, S.; Parvathy, R.; Shalini, V.; Helen, A.; Jayalekshmy, A. Isolation, characterization and quantification of tricin and flavonolignans in the medicinal rica Njavara (Oryza sativa L.), as compared to staple varieties. Plant Food Hum. Nutr. 2011, 66, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Imm, J.Y. Antiobesity effect of tricin, a methylated cereal flavone, in high-fat-diet-induced obese mice. J. Agric. Food Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Park, H.Y.; Kim, S.; Park, Y.; Bang, M.H.; Imm, J.Y. Anti-adipogenic effect of oat hull extract containing tricin on 3T3-L1 adipocytes. Process. Biochem. 2015, 50, 2314–2321. [Google Scholar] [CrossRef]
- Sadanari, H.; Fujimoto, K.J.; Sugihara, Y.; Ishida, T.; Takemoto, M.; Daikoku, T.; Murayama, T. The anti-human cytomegalovirus drug tricin inhibits cyclin-dependent kinase 9. FEBS Open Bio 2018, 8, 646–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Seki, M.; Kobayashi, K. Inhibition of xanthine oxidase by flavonoids. Biosci. Biotechnol. Biochem. 1990, 63, 1787–1790. [Google Scholar] [CrossRef]
- Cos, P.; Ying, L.; Calomme, M.; Hu, J.P.; Cimanga, K.; Poel, B.V.; Pieters, L.; Vlietinck, A.J.; Berghe, D.V. Structure-activity relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Spanou, C.; Veskoukis, A.S.; Kerasioti, T.; Kontou, M.; Angelis, A.; Aligiannis, N.; Skaltsounis, A.L.; Kouretas, D. Flavonoid glycosides isolated from unique legume plant extracts as novel inhibitors of xanthine oxidase. PLoS ONE 2012, 7, e32214. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Awale, S.; Tezuka, Y.; Ueda, J.Y.; Tran, Q.L.; Kadota, S. Xanthine oxidase inhibitors from the flowers of Chrysanthemum sinense. Planta Med. 2006, 72, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Walle, T. Methylated flavonoids have greatly improved intestinal absorption and metabolic stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, R.; Zhang, G.W.; Gong, D.M. Mechanistic insights into the inhibition of quercetin on xanthine oxidase. Int. J. Biol. Macromol. 2018, 112, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Zhang, G.W.; Pan, J.H.; Gong, D.M. Novel insights into the inhibitory mechanism of kaempferol on xanthine oxidase. J. Agric. Food Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.Q.; Chen, S.D.; Xie, Y.Z.; Shuai, O.; Li, X.M.; Chen, D.L.; Su, J.Y.; Jiao, C.W.; Liang, Y.L. Hypouricemic effects of extracts from Agrocybe aegerita on hyperuricemia mice and virtual prediction of bioactives by molecular docking. Front. Pharmacol. 2018, 9, 498. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.I.; Fujita, T.; Akaho, E.; Nagamatsu, T. A comparative study of AutoDock and PMF scoring performances, and SAR of 2-substituted pyrazolotriazolopyrimidines and 4-substituted pyrazolopyrimidines as potent xanthine oxidase inhibitors. J. Comput. Aided Mol. Des. 2010, 24, 57–75. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.N.; Pauff, J.M.; Hille, R. Substrate orientation and catalytic specificity in the action of xanthine oxidase: The sequential hydroxylation of hypoxanthine to uric acid. J. Biol. Chem. 2010, 285, 28044–28053. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.S.; Ballou, D.P.; Palmer, G.; Massey, V. The mechanism of action of xanthine oxidase. J. Biol. Chem. 1974, 249, 4363–4382. [Google Scholar] [PubMed]
- Sun, S.W.; Dai, X.Y.; Sun, J.; Bu, X.G.; Weng, C.H.; Li, H.; Zhu, H. A diketopiperazine factor from Rheinheimera aquimaris QSI02 exhibits anti-quorum sensing activity. Sci. Rep. 2016, 6, 39637. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.N.; Wang, W.; Zhang, C.Y.; Shi, H.B.; Liu, Y.; Liu, X.H.; Guo, B.H.; Zhao, D.M.; Gao, H. Bioassay-guided isolation and identification of xanthine oxidase inhibitory constituents from the leaves of Perilla frutescens. Molecules 2015, 20, 17848–17859. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, W.; Guo, B.H.; Gao, H.; Liu, Y.; Liu, X.H.; Yao, H.L.; Cheng, K. Chemical evidence for potent xanthine oxidase inhibitory activity of ethyl acetate extract of Citrus aurantium L. dried immature fruits. Molecules 2016, 21, 302. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of tricin is available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XO Inhibition (Mean ± S.D.%) | |||
|---|---|---|---|
| 200 μg/mL | 100 μg/mL | 50 μg/mL | |
| 70% ethanol extract | 38.26 ± 1.07 | 21.19 ± 1.14 | 8.33 ± 0.69 |
| Petroleum ether-soluble part | 27.84 ± 1.00 | 17.12 ± 1.97 | 12.69 ± 2.15 |
| Ethyl acetate-soluble part | 66.93 ± 1.52 | 57.00 ± 1.65 | 42.26 ± 0.79 |
| n-Butanol-soluble part | 18.11 ± 1.41 | 10.23 ± 0.29 | N.I. 1 |
| Water-soluble part | N.I. | N.I. | N.I. |
| Species | Plant Parts | Common Name | mg Tricin/kg Dry Weight (Mean ± S.D.%) | |
|---|---|---|---|---|
| Before Hydrolysis | After Hydrolysis | |||
| Oryza sativa | hull | rice hull | 155.16 ± 1.03 | 188.32 ± 2.27 |
| straw | rice straw | 722.78 ± 22.82 | 1143.86 ± 54.70 | |
| Triticum aestivum | hull | wheat hull | 511.35 ± 15.80 | 869.98 ± 33.76 |
| straw | wheat straw | 940.09 ± 13.50 | 1925.05 ± 17.89 | |
| bran | wheat bran | N.D. 1 | N.D. | |
| Hordeum vulgare | bran | barley bran | 33.14 ± 2.44 | 36.82 ± 0.28 |
| Sorghum bicolor | bran | sorghum bran | N.D. | N.D. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.-X.; Sun, S.-W.; Yuan, W.-J.; Gao, H.; Si, Y.-Y.; Liu, K.; Zhang, S.; Liu, Y.; Wang, W. Isolation of Tricin as a Xanthine Oxidase Inhibitor from Sweet White Clover (Melilotus albus) and Its Distribution in Selected Gramineae Species. Molecules 2018, 23, 2719. https://doi.org/10.3390/molecules23102719
Liu X-X, Sun S-W, Yuan W-J, Gao H, Si Y-Y, Liu K, Zhang S, Liu Y, Wang W. Isolation of Tricin as a Xanthine Oxidase Inhibitor from Sweet White Clover (Melilotus albus) and Its Distribution in Selected Gramineae Species. Molecules. 2018; 23(10):2719. https://doi.org/10.3390/molecules23102719
Chicago/Turabian StyleLiu, Xiao-Xiao, Shi-Wei Sun, Wen-Jing Yuan, Hua Gao, Yue-Yue Si, Kun Liu, Shuang Zhang, Yang Liu, and Wei Wang. 2018. "Isolation of Tricin as a Xanthine Oxidase Inhibitor from Sweet White Clover (Melilotus albus) and Its Distribution in Selected Gramineae Species" Molecules 23, no. 10: 2719. https://doi.org/10.3390/molecules23102719
APA StyleLiu, X.-X., Sun, S.-W., Yuan, W.-J., Gao, H., Si, Y.-Y., Liu, K., Zhang, S., Liu, Y., & Wang, W. (2018). Isolation of Tricin as a Xanthine Oxidase Inhibitor from Sweet White Clover (Melilotus albus) and Its Distribution in Selected Gramineae Species. Molecules, 23(10), 2719. https://doi.org/10.3390/molecules23102719