HPLC-UV/Vis-APCI-MS/MS Determination of Major Carotenoids and Their Bioaccessibility from “Delica” (Cucurbita maxima) and “Violina” (Cucurbita moschata) Pumpkins as Food Traceability Markers

 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Carotenoid Determination by HPLC-UV/Vis-APCI-MS/MS
2.2. Carotenoid Levels in Raw and Cooked Pumpkins
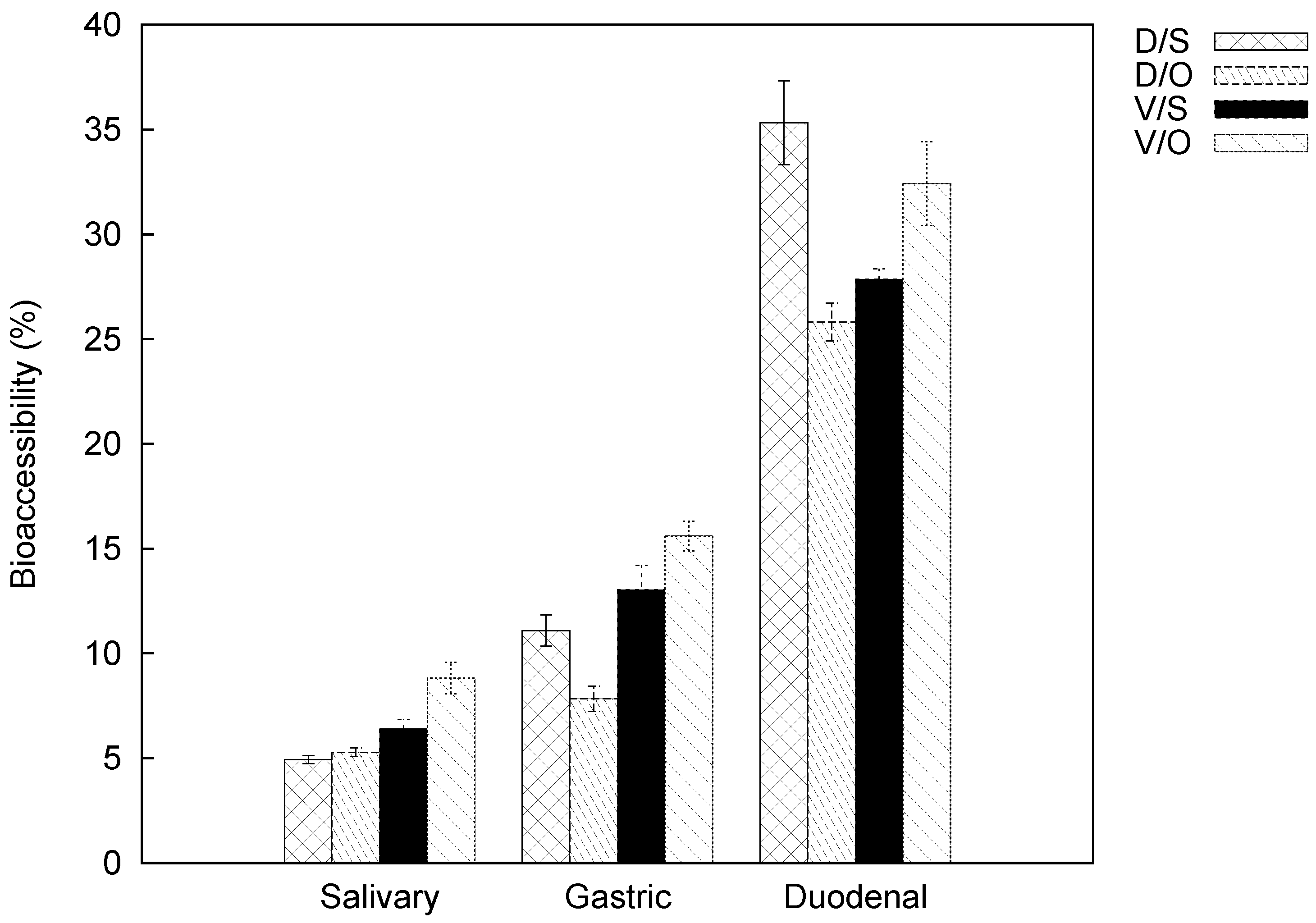
2.3. Carotenoid Bioaccessibility
3. Materials and Methods
3.1. Chemicals
3.2. Instrumentation
3.3. Reagent Preparation
3.4. Sample Collection and Preparation
3.5. Cooking Methods
3.6. Carotenoids Extraction
3.7. In Vitro Static Digestion Model
Carotenoid Extraction from Digesta
3.8. HPLC-UV/Vis-APCI-MS/MS Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Britton, G. Structure and properties of carotenoids in relation to function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Britton, G.; Vicario, I.M.; Heredia, F.J. Relationship between the colour and the chemical structure of carotenoid pigments. Food Chem. 2007, 101, 1145–1150. [Google Scholar] [CrossRef]
- Lako, J.; Trenerry, V.C.; Wahlqvist, M.; Wattanapenpaiboon, N.; Sotheeswaran, S.; Premier, R. Phytochemical flavonols, carotenoids and the antioxidant properties of a wide selection of Fijian fruit, vegetables and other readily available foods. Food Chem. 2007, 101, 1727–1741. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. Antioxidant effects of carotenoids in Vivo and in Vitro: An overview. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 213, pp. 403–420. [Google Scholar]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, A.; Skibsted, L.H.; Sampson, J.; Rice-Evans, C.; Everett, S.A. Comparative mechanisms and rates of free radical scavenging by carotenoid antioxidants. FEBS Lett. 1997, 418, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. BBA-Mol. Cell Biol. 2012, 1821, 70–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.; Gomes, A.; Karakaya, S.; Martínez-Cuesta, M.; McDougall, G.; Requena, T.; et al. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef]
- Balant, L. Is there a need for more precise definitions of bioavailability? Eur. J. Clin. Pharmacol. 1991, 40, 123–126. [Google Scholar]
- Parada, J.; Aguilera, J. Food microstructure affects the bioavailability of several nutrients. J. Food Sci. 2007, 72, R21–R32. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.; Cotter, P.; López-Expósito, I.; Kleiveland, C.; Lea, T.; Mackie, A.; Requena, T.; Swiatecka, D.; Wichers, H. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Berlin, Germany, 2015. [Google Scholar]
- Carvalho, L.M.J.D.; Smiderle, L.D.A.S.M.; Carvalho, J.L.V.D.; Cardoso, F.D.S.N.; Koblitz, M.G.B. Assessment of carotenoids in pumpkins after different home cooking conditions. Food Sci. Technol. 2014, 34, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Provesi, J.G.; Dias, C.O.; Amante, E.R. Changes in carotenoids during processing and storage of pumpkin puree. Food Chem. 2011, 128, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Azevedo-Meleiro, C.H.; Rodriguez-Amaya, D.B. Qualitative and quantitative differences in carotenoid composition among Cucurbita moschata, Cucurbita maxima, and Cucurbita pepo. J. Agric. Food Chem. 2007, 55, 4027–4033. [Google Scholar] [CrossRef] [PubMed]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of different cooking methods on nutritional and physicochemical characteristics of selected vegetables. J. Agric. Food Chem. 2007, 56, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Yeum, K.J.; Russell, R.M. Carotenoid bioavailability and bioconversion. Annu. Rev. Nutr. 2002, 22, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, L.; Nagao, A. Intestinal absorption of dietary carotenoids. Mol. Nutr. Food Res. 2007, 51, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Turcsi, E.; Nagy, V.; Deli, J. Study on the elution order of carotenoids on endcapped C18 and C30 reverse silica stationary phases. A review of the database. J. Food Compos. Anal. 2016, 47, 101–112. [Google Scholar] [CrossRef]
- Wald, J.P.; Nohr, D.; Biesalski, H.K. Rapid and easy carotenoid quantification in Ghanaian starchy staples using RP-HPLC-PDA. J. Food Compos. Anal. 2018, 67, 119–127. [Google Scholar] [CrossRef]
- Lee, H.; Castle, W.; Coates, G. High-performance liquid chromatography for the characterization of carotenoids in the new sweet orange (Earlygold) grown in Florida, USA. J. Chromatogr. A 2001, 913, 371–377. [Google Scholar] [CrossRef]
- de Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids, by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef] [PubMed]
- Britton, G. Carotenoids. In Natural Food Colorants; Springer: Berlin, Germany, 1996; pp. 197–243. [Google Scholar]
- Rodriguez-Amaya, D.B.; Kimura, M.; Godoy, H.T.; Amaya-Farfan, J. Updated Brazilian database on food carotenoids: Factors affecting carotenoid composition. J. Food Compos. Anal. 2008, 21, 445–463. [Google Scholar] [CrossRef]
- Hornero-Méndez, D.; Mínguez-Mosquera, M.I. Bioaccessibility of carotenes from carrots: Effect of cooking and addition of oil. Innov. Food Sci. Emerg. Technol. 2007, 8, 407–412. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M. The TNO gastro-intestinal model (TIM). In The Impact of Food Bioactives on Health; Springer: Berlin, Germany, 2015; pp. 37–46. [Google Scholar]
- Kurz, C.; Carle, R.; Schieber, A. HPLC-DAD-MS characterisation of carotenoids from apricots and pumpkins for the evaluation of fruit product authenticity. Food Chem. 2008, 110, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Alegría, A.; Garcia-Llatas, G.; Cilla, A. Static digestion models: General introduction. In The Impact of Food Bioactives on Health; Springer: Berlin, Germany, 2015; pp. 3–12. [Google Scholar]
- Biehler, E.; Kaulmann, A.; Hoffmann, L.; Krause, E.; Bohn, T. Dietary and host-related factors influencing carotenoid bioaccessibility from spinach (Spinacia oleracea). Food Chem. 2011, 125, 1328–1334. [Google Scholar] [CrossRef]
- Böhm, V. Use of column temperature to optimize carotenoid isomer separation by C30 high performance liquid chromatography. J. Sep. Sci. 2001, 24, 955–959. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Retention Time (min) | Compound | % III/II | [M + H] (m/z) | Fragment Ions (m/z) | |
|---|---|---|---|---|---|---|
| 1 | violaxanthin | 87 | 601 | 534, 583, 429 | ||
| 2 | astaxanthin | 82 | 597 | 579, 285, 379 | ||
| 3 | antheraxanthin | 27 | ||||
| 4 | zeaxanthin | 53 | 569 | 551, 489, 477, 416 | ||
| 5 | lutein | 53 | 569 | 495, 459, 430, 477 | ||
| 6 | lycopene | 72 | 537 | 457, 413, 177 | ||
| 7 | 58 | |||||
| 8 | 54 | |||||
| 9 | 53 | |||||
| 10 | -carotene | 51 | 537 | 457, 413, 177 | ||
| 11 | -carotene | 18 | 537 | 457, 445, 413, 400, 269, 177 |
| Peak | Absolute Quantification | Relative Quantification | ||
|---|---|---|---|---|
| C. maxima (Delica) | C. moschata (Violina) | C. maxima (Delica) | C. moschata (Violina) | |
| violaxanthin | ||||
| astaxanthin | ||||
| antheraxanthin | ||||
| zeaxanthin | nd | nd | ||
| lutein | ||||
| lycopene | ||||
| 7 | ||||
| 8 | ||||
| 9 | ||||
| -carotene | nd | nd | ||
| β-carotene | ||||
| C. maxima (Delica) | C. moschata (Violina) | |||
|---|---|---|---|---|
| Lutein | -carotene | Lutein | -carotene | |
| Oven-cooked | ||||
| Steam-cooked | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergantin, C.; Maietti, A.; Tedeschi, P.; Font, G.; Manyes, L.; Marchetti, N. HPLC-UV/Vis-APCI-MS/MS Determination of Major Carotenoids and Their Bioaccessibility from “Delica” (Cucurbita maxima) and “Violina” (Cucurbita moschata) Pumpkins as Food Traceability Markers. Molecules 2018, 23, 2791. https://doi.org/10.3390/molecules23112791
Bergantin C, Maietti A, Tedeschi P, Font G, Manyes L, Marchetti N. HPLC-UV/Vis-APCI-MS/MS Determination of Major Carotenoids and Their Bioaccessibility from “Delica” (Cucurbita maxima) and “Violina” (Cucurbita moschata) Pumpkins as Food Traceability Markers. Molecules. 2018; 23(11):2791. https://doi.org/10.3390/molecules23112791
Chicago/Turabian StyleBergantin, Caterina, Annalisa Maietti, Paola Tedeschi, Guillermina Font, Lara Manyes, and Nicola Marchetti. 2018. "HPLC-UV/Vis-APCI-MS/MS Determination of Major Carotenoids and Their Bioaccessibility from “Delica” (Cucurbita maxima) and “Violina” (Cucurbita moschata) Pumpkins as Food Traceability Markers" Molecules 23, no. 11: 2791. https://doi.org/10.3390/molecules23112791