1. Introduction
Catheter-associated urinary tract infection (CA-UTI) is commonly acquired in hospitals around the world [
1]. This infection is caused by uropathogenic
E. coli (UPEC) due to its capacity to adhere to catheters and develop biofilms [
2,
3]. UPEC biofilms on catheters include communities of microorganisms adhered to a silicon surface, embedded in an extracellular polymeric substances matrix, and with altered metabolism compared to the corresponding planktonic cells [
4]. The biofilm-secreted polymeric substances protect the embedded cells against antibiotics, evade the host immune defense, and promote persistence in the environment, causing recurrent infections [
4]. In addition, persistent UTIs can cause pyelonephritis, leading to parenchymal injury or renal scarring, activation of inflammatory mediators, and overproduction of reactive oxygen species [
5]. Consequently, the treatment of UTI is a significant challenge, considering bacterial evolution against conventional treatments, mainly when resistance at the planktonic cellular level and community level occur; besides, the oxidative complications involved with this infection [
6].
UTI is commonly treated with antibiotics, such as ampicillin, trimethoprim, cephalosporin, nalidixic acid, and nitrofurantoin [
7], but alternatives need to be evaluated, considering the rapid emergence of antibiotic resistance, the presence of inflammation, and oxidative damage. It has been reported that planktonic
E. coli isolated from the urine of patients with CA-UTI are resistant to most frequently used antibiotics [
3]. Antibiotics are developed to inhibit bacterial growth or kill bacteria in planktonic cells, but these agents are less active in avoiding cellular adhesion, inactivate formed biofilms, or inactivate free radicals [
8]. The recurrence of UTI, bacterial biofilms resistance, and the oxidative damage of the tissue have promoted the search for alternative antimicrobial-antioxidant therapies [
9,
10].
The consumption of functional plant foods and medicinal plants (for example, cranberry juice rich in proanthocyanidins) has been widely recommended to prevent urinary infections [
9]. However, after the consumption of cranberry, the complex phenolic compounds are metabolized to simple phenols like phenolic acids and flavonoids [
11]. Several clinical studies have reported the presence of these compounds in urine after cranberry ingestion, with protocatechuic acid, vanillic acid, and catechin as the most commonly found [
12,
13]. These results suggest that those specific phenolic compounds could also exert an antibacterial effect, reduce urinary infections, and also act as antioxidants. Thus, the research question of this study is, what type of effect is caused by the combined presence of catechin and vanillic and protocatechuic acid on the growing, adhesion, and biofilm eradication of UPEC, as well as on the antioxidant capacity? The antibacterial potential of catechin, protocatechuic, and vanillic acids against planktonic
E. coli has been tested and proved to be effective [
14,
15,
16]; however, their efficacy to inhibit biofilm formation of uropathogenic
E. coli has not been investigated, nor the effect of their ternary combination. The presence of these phenolic compounds could exert a synergic effect to control uropathogenic
E. coli at different levels, including targeting planktonic cell survival, adhesion, biofilm eradication, and free radical inactivation [
17]. In this context, the purpose of this study was to determine the effect of catechin, vanillic, and protocatechuic acids and their combination to prevent and eradicate uropathogenic
E. coli biofilm on silicone catheters, besides acting as antioxidants.
3. Discussion
The tested phenolic compounds were useful to inhibit the growth of planktonic cells and cause bacterial death. There is evidence that phenolic acids can cause irreversible modifications to the permeability of bacterial membranes and decrease cell surface charges, causing rupture or forming pores, with consequent leakage of intracellular components [
18]. In addition, flavonoids, like catechin, could chelate metals essentials as enzymatic cofactors involved in bacterial growth [
19]. These different targets for the tested phenolic compounds could be the reason for the observed synergy. It has to be noticed that even when catechin showed no bactericidal effect at the highest tested concentrations, it seems to have a role in improving the efficacy of phenolic acids in the combination.
No previous studies have reported the antibacterial effect of these phenolic acids and catechin against UPEC; however, similar MICs have been reported for these individual phenolic acids against other bacteria [
15,
20]. Protocatechuic acid showed an MIC of 19.46 mM against
Listeria monocytogenes, and vanillic acid showed MICs of 17.84 and 14.86 mM against
E. coli O157:H7 and
Salmonella Typhimurium, respectively. On the other hand, flavonoids such as catechins, showed antibacterial potential with MICs ranging from 31 mM to 344 mM against
E. coli,
S. Typhimurium, and
Staphylococcus aureus [
16,
21]. In average, the obtained MIC values are in the range previously reported, and the found variations could be attributed to different bacterial strains.
Adhesion of uropathogenic
E. coli on the silicone surface of catheters is responsible for its persistence in hospital environments. The inhibition of adhesion is the first step in interrupting biofilm formation and its complications, including UTI and pyelonephritis. Regrettably, few antibiotics have this ability, including nitrofurantoin, and the observed anti-adhesive effect of protocatechuic and vanillic acids must be recognized, as well as the eradication of pre-formed biofilms and contribution of catechin to antioxidant capacity. Trying to relate the observed effect of the tested phenolic compounds with their molecular properties, it has to be mentioned that vanillic acid is more lipophilic (log
p = 1.43) than protocatechuic acid (Log
p = 0.86), and catechin (Log
p = 0.4), and, because of this, it could easily interfere with the cell membrane and possibly affect bacterial adhesion. Previous reports indicated that gallic, ferulic, caffeic, and chlorogenic acids prevented bacterial adhesion, increased superficial hydrophobicity, and decreased the cell adhesion potential [
22,
23,
24]. Furthermore, bacterial adhesion can be affected by inhibiting the secretion of polymeric substances [
25]. Epigallocatechin gallate can eliminate biofilm matrix of
E. coli by interfering with the assembly of curli subunits into amyloid fibers and cellulose biosynthesis, and by reducing the expression of CsgD—a crucial activator of curli and cellulose [
25].
Phenolic compounds can interfere with cell-cell communication by various mechanisms, such as inhibiting the synthesis of signal molecules or receptors, which in turn affects the synthesis of polymeric extracellular substances and biofilm development [
26]. It is known that compounds like quercetin, kaempferol, naringenin, and apigenin reduced
E. coli biofilm formation because they suppressed the autoinducer-2 activity, which is responsible for cell-to-cell communication involved in biofilm production [
26]. In addition, Lee et al. [
27] confirmed that phloretin belonging to flavonoids suppressed autoinducer-2 importer genes of
E. coli O157:H7 biofilm cells, confirming the inhibition of cell communication. Thus, it would be interesting to test whether some of these mechanisms are involved in the observed UPEC responses caused by phenolic compounds.
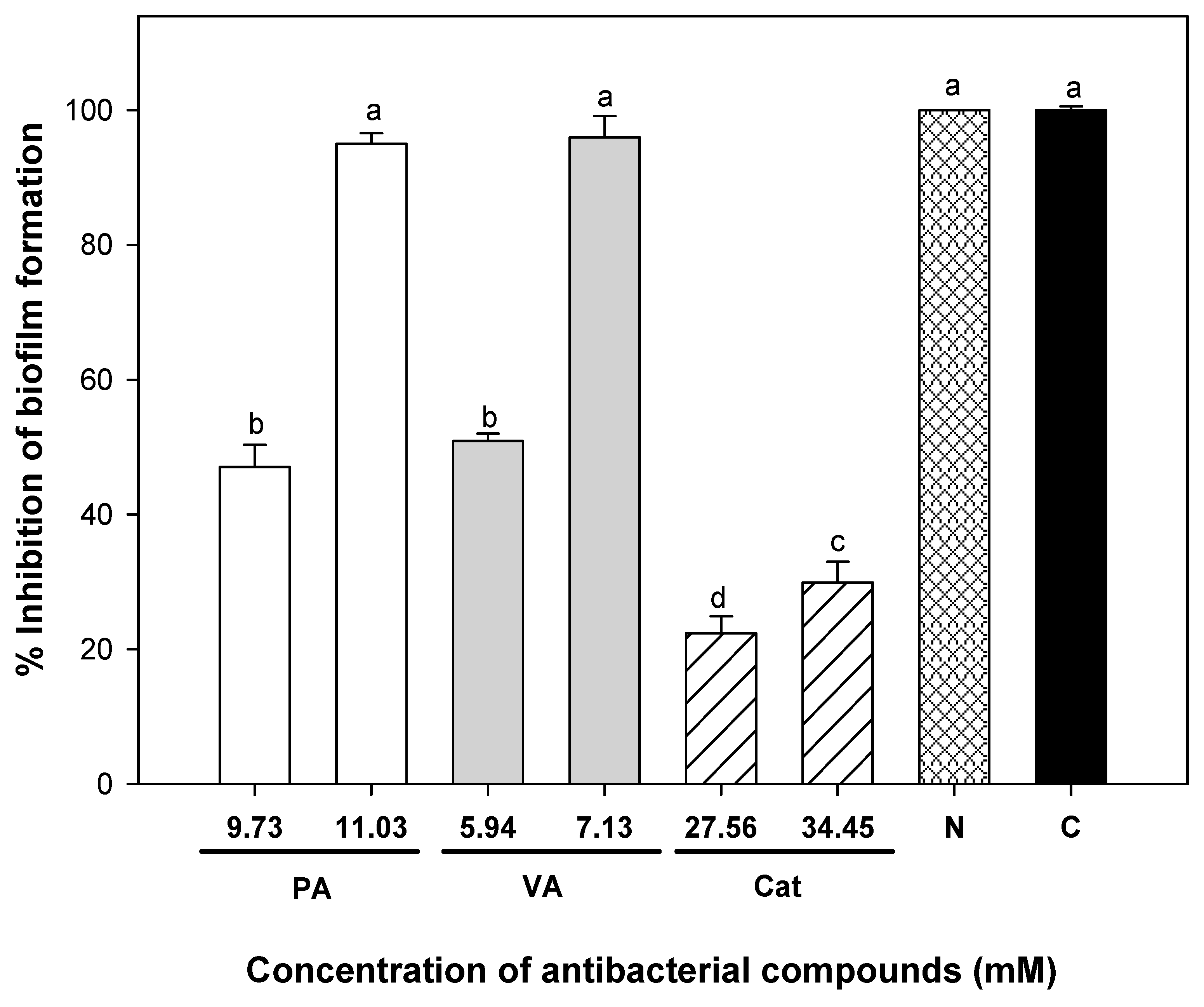
The achieved dose reductions when combining phenolic compounds were 6.89, 9.63, and 1398 times for protocatechuic acid, vanillic acid, and catechin against UPEC adhesion, respectively. At this point, the effective doses to avoid cell adhesion were lower than those needed to have a bactericidal effect. Another interesting fact is that nitrofurantoin did not seem to affect any mechanism of adhesion but instead attacked the viability of planktonic cells; this could be attributed to its mode of action. It does not interfere with motility, adhesion, or cellular communication, which are factors influencing biofilm development [
28]. However, nitrofurantoin’s mechanism of action is complex and not completely understood, but it appears to be caused by the production of nitrofurans (reactive intermediates) that can damage ribosomal proteins, DNA, and other macromolecules [
29]. For this reason, it is important to consider alternative antibacterial agents to this antibiotic. Besides, as observed previously, cell adhesion and biofilm formation of UPEC took a short time (4 h) in catheters (
Figure 1), and nitrofurantoin only affected planktonic cells. Its administration must occur before cellular attachment does.
Higher doses of phenolic compounds were needed to eradicate biofilms compared to planktonic cell adhesion, and this reflects the attributed resistance granted to this association against the tested antimicrobial agents. However, vanillic and protocatechuic acids were effective at eliminating viable cells from 24 h pre-formed biofilms of uropathogenic
E. coli on silicone surfaces; while catechin seemed to disrupt the biofilm biomass. The achieved reductions combining phenolic compounds were 8, 8, and 40 times for protocatechuic acid, vanillic acid, and catechin, respectively. No reduction of viable cells from pre-formed biofilms was achieved using nitrofurantoin, and these results reinforced the need for using alternatives to prevent biofilm formation and substitutes for this antibiotic. It has been reported that biofilm removal and biofilm cell inactivation are done by distinct processes. Biofilm eradication refers to inactivating bacteria, but those can remain attached to the surface; on the other hand, biofilm removal has to be done by the elimination of the attached extracellular polymeric substances. Any agent with biofilm eradication activity must pass the matrix created by extracellular polymeric substances and attack the embedded viable cells. Biofilm bacteria can be inactivated by the antimicrobial activity of phenolic acids, which include disruption of cytoplasmic membranes, enzyme inhibition through reaction with the sulfhydryl group, or non-specific interactions with proteins and synthesis of nucleic acids [
18]. Nevertheless, the mechanism in embedded biofilm bacteria could be different, considering the interference of the biofilm constituents. These results highlight the importance of combining antibacterial agents with a different mode of action, with at least one able to disrupt the polymer matrix and another able to cause bacterial cell death.
The antibacterial synergism observed in the combination of phenolic compounds could be attributed to different antibacterial mechanisms to inhibit biofilm formation. A possible synergistic mechanism proposed in our study is that vanillic acid and protocatechuic acid interact with the lipid layer of the bacterial membrane, affecting its structural integrity or functionality, and as a consequence, affect bacterial adhesion. This could facilitate the entrance of catechin, and this compound can interact with other molecules such as enzymes within the cytoplasm, affecting bacterial metabolism. The simultaneous action of these two mechanisms could be responsible for the observed synergistic antibacterial effect that inhibits cell density in biofilm formation, but more research is needed to test this hypothesis.
It was observed that the phenolic combinations used against planktonic and biofilm inhibition had a synergistic effect, and their combination at the used proportions in the biofilm eradication test had a synergistic effect in the DPPH and FRAP assays. The difference in the results observed between the used reactive specie could be attributed to their different reaction mechanism. Due to the complexity of simulating the characteristics of an oxidative reaction, it has been widely reported that multiple methods are necessary to test antioxidant activity [
30]. Phenolic compounds are considered to be efficient hydrogen donors due to their number, arrangement of hydroxyl groups, and the specific carboxyl group, which can be easily ionized. A positive correlation has been found between the number of hydroxyl groups and antioxidant activity [
31]. This molecular attribute could be the reason that catechin with five hydroxyls was the most effective antioxidant, followed by protocatechuic with two. Vanillic acid was the least antioxidant with only one hydroxyl group.
Furthermore, antioxidants can interact with each other when combined, showing some effects as synergistic, additive, and antagonistic. Some authors consider that a synergistic effect occurs when two or more antioxidants show a total effect higher than the sum of the individual effects. An additive effect refers to the sum of the effects of each compound, while an antagonistic effect reflects a lower effectiveness than the sum of the individual effects [
32]. There could be three different mechanisms in the synergistic effect: 1. one compound makes the antioxidant effect, and the other regenerates it to have that effect again; 2. sacrificial oxidation of an antioxidant to protect the other; and 3. combination of two or more agents with different antioxidant mechanisms.
Our results suggest that the studied phenolic compounds acted synergistically, depending on their concentration and the testing method, and they were able to donate hydrogen or transfer electrons to the reactive species and also regenerate or protect their partner compounds. Similar antioxidant capacities have been reported for some of these phenolic compounds: Palafox-Carlos, Gil-Chavez, Sotelo-Mundo, Namiesnik, Gorinstein and Gonzalez-Aguilar [
31] reported that protocatechuic acid showed higher antioxidant capacity than vanillic acid against DPPH
• radical inhibition. In addition, these authors reported several binary, ternary, and quaternary combinations of phenolic acids, and most of these combinations had a synergistic effect. In addition, Skroza, Mekinic, Svilovic, Simar and Katalinic [
30] investigated the antioxidant activity of phenolic binary mixtures. Their results suggested that some differences in the activity of the mixture were found depending on the method used. They demonstrated that the combination of catechin with resveratrol had a synergistic effect by DPPH
• and FRAP assays and that caffeic acid and resveratrol mixture showed synergistic activity by FRAP method, having an antagonistic effect by DPPH
• method.
As stated before, persistent UTIs can cause pyelonephritis and overproduction of free radicals [
33]; then, the addition of antioxidant compounds in its treatment is an alternative to relieving the symptoms caused by reactive oxygen species [
5,
10,
34]. In this context, treatments with caffeic acid phenethyl ester (0.01 mM) were related to the repair of renal tissue damage after pyelonephritis, in addition to reducing chronic inflammation by enforcing the enzymatic antioxidant defense system [
35]. Caffeic acid phenethyl ester administrated at 10 µM/kg body-weight reduced malondialdehyde and nitric oxide levels as well as xanthine oxidase activity, although it increased superoxide dismutase and glutathione peroxidase activities during pyelonephritis caused by
E. coli in rats. In addition, a single dose of cranberry beverages with 111 g of phenolic compounds improved indices of oxidative stress, such as elevating blood glutathione peroxidase and superoxide dismutase activity, and exerted an ex vivo anti-adhesion activity against P-fimbriated
E. coli compared to placebo in healthy humans, given evidence of the efficacy of the use of the therapeutic effect of extracts rich in phenolic compounds [
36]. However, more information regarding the contribution of the studied compounds is needed. In summary, our results suggested that the combination of protocatechuic acid, vanillic acid, and catechin could be a promising alternative to use in UTI treatment; however, this study could be taken as the initial step to continue searching for more evidence to sustain the real impact of the alleged responses.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}