Genuine and Sequestered Natural Products from the Genus Orobanche (Orobanchaceae, Lamiales)



Pharmazeutisches Institut, Abteilung Pharmazeutische Biologie, Christian-Albrechts-Universität zu Kiel, Gutenbergstraße 76, 24118 Kiel, Germany
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(11), 2821; https://doi.org/10.3390/molecules23112821
Submission received: 10 October 2018
/
Revised: 26 October 2018
/
Accepted: 28 October 2018
/
Published: 30 October 2018
(This article belongs to the Special Issue Natural Products and Drug Discovery)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The present review gives an overview about natural products from the holoparasitic genus Orobanche (Orobanchaceae). We cover both genuine natural products as well as compounds sequestered by Orobanche taxa from their host plants. However, the distinction between these two categories is not always easy. In cases where the respective authors had not indicated the opposite, all compounds detected in Orobanche taxa were regarded as genuine Orobanche natural products. From the about 200 species of Orobanche s.l. (i.e., including Phelipanche) known worldwide, only 26 species have so far been investigated phytochemically (22 Orobanche and four Phelipanche species), from 17 Orobanche and three Phelipanche species defined natural products (and not only natural product classes) have been reported. For two species of Orobanche and one of Phelipanche dedicated studies have been performed to analyze the phenomenon of natural product sequestration by parasitic plants from their host plants. In total, 70 presumably genuine natural products and 19 sequestered natural products have been described from Orobanche s.l.; these form the basis of 140 chemosystematic records (natural product reports per taxon). Bioactivities described for Orobanche s.l. extracts and natural products isolated from Orobanche species include in addition to antioxidative and anti-inflammatory effects, e.g., analgesic, antifungal and antibacterial activities, inhibition of amyloid β aggregation, memory enhancing effects as well as anti-hypertensive effects, inhibition of blood platelet aggregation, and diuretic effects. Moreover, muscle relaxant and anti-spasmodic effects as well as anti-photoaging effects have been described.
1. Introduction
Taxa of the genus Orobanche sensu lato (Orobanchaceae) are non-photosynthetic root holoparasites. The genus Orobanche s.l. is distributed worldwide. Most of the about 200 species are native to the Northern Hemisphere [1,2]. The phylogeny of Orobanche s.l. is still discussed controversially. Park et al., (2008) [3], distinguished five lineages within Orobanche s.l. based on morphological, cytological, and (macro-)molecular phylogenetic data. Schneeweiss (2013) [4] suggested treating these lineages as separate genera. From these five lineages, four have traditionally been regarded as sections of Orobanche [the respective generic names suggested by Schneeweiss (2013) [4] are indicated in brackets]: Gymnocaulis Nutt. (=genus Aphyllon Mitch.), Myzorrhiza (Phil.) Beck (=sect. Nothaphyllon (A.Gray) Heckard = genus Myzorrhiza Phil.) [5], Trionychon Wallr. (=genus Phelipanche Pomel), and Orobanche L. (=sect. Osproleon Wallr. = genus Orobanche L. s.str.). The only representative of the fifth clade, Orobanche latisquama Reut. ex Boiss. (=genus Boulardia F.W.Schultz), was formerly assigned to sect. Orobanche. Additionally, the genus Phelypaea is part of the clade comprising Orobanche s.l. [4]. On the basis of recent taxonomic and phylogenetic data, the genus Orobanche s.l. was split into two separate genera, Orobanche and Phelipanche [6]. Following this decision the names Phelipanche ramosa (L.) Pomel and P. aegyptiaca (Pers.) Pomel, instead of Orobanche ramosa L. and O. aegyptiaca Pers. (of sect. Trionychon) are applicable [7]. In the following paragraphs, the new species names are used and synonyms in the original publications are indicated if applicable. Orobanche species play an important (negative) role in agriculture as they can pose serious threats to major crop plants [8,9]. Many research papers are focused on host-parasite interactions via strigolactones (seed germination promoters) in terms of weed management. Less is known about phytochemical compounds in the parasitic species. This review is intended to give an overview over secondary metabolites synthesized by Orobanche s.l. known so far. For Orobanche sections Gymnocaulis and Myzorrhiza as well as for O. latisquama, no studies on secondary metabolites have been published yet. Members of Orobanche section Orobanche and of Phelipanche that have been phytochemically analyzed are reviewed in alphabetical order. An additional paragraph describes sequestration of secondary metabolites from host plants. Characterizations of bioactivities of Orobanche s.l. extracts and pure compounds isolated from Orobanche species are described separately. Literature data were retrieved from SciFinder and ISI Web of Knowledge databases. Reports until the end of 2017 were taken into account (exact search terms can be found in Supplementary Text S1). All taxon names were reviewed and accepted names according to The Plant List are used throughout the manuscript [10]. Usage of synonyms deviating from The Plant List in the cited literature was mentioned when applicable.
2. Results and Discussion
2.1. Literature Data on Natural Products from Orobanche Species
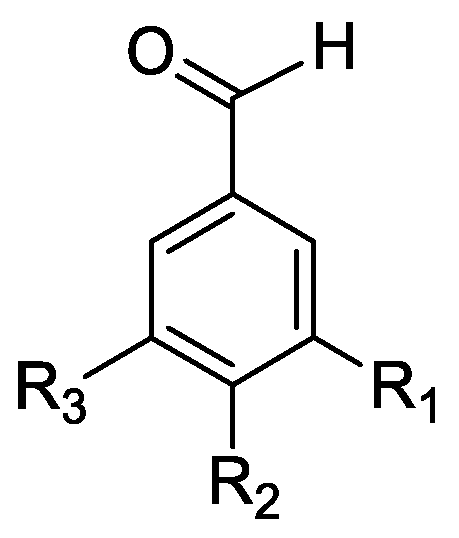
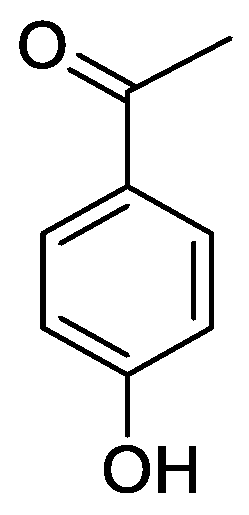
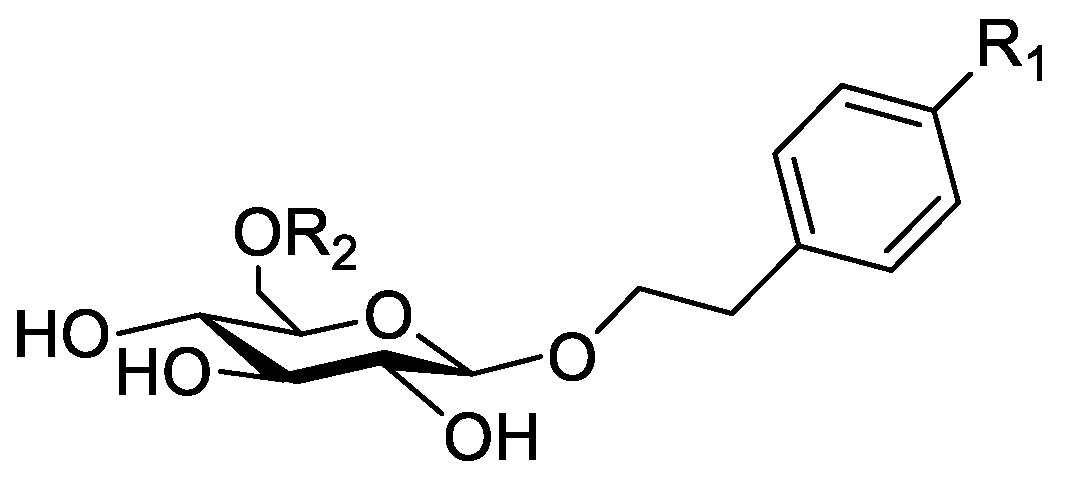
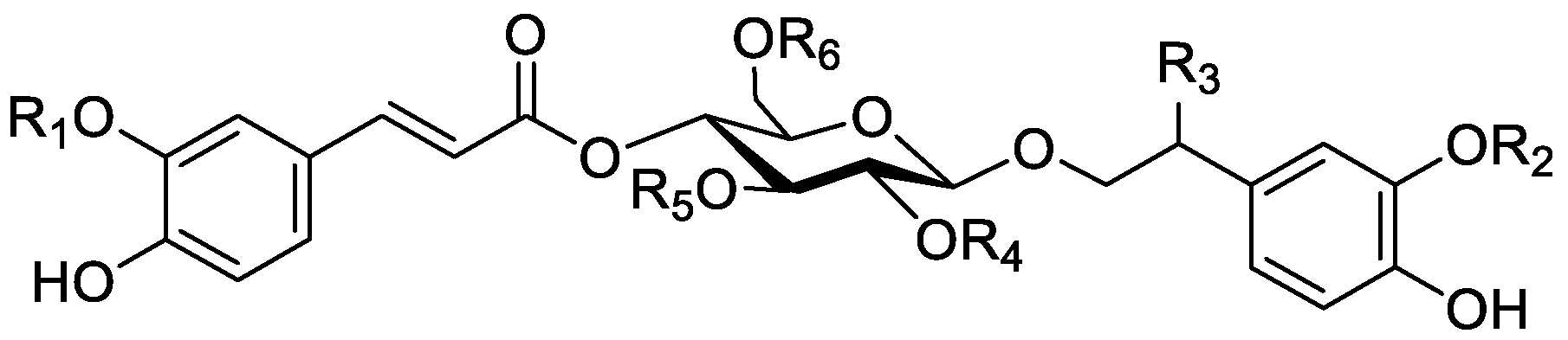
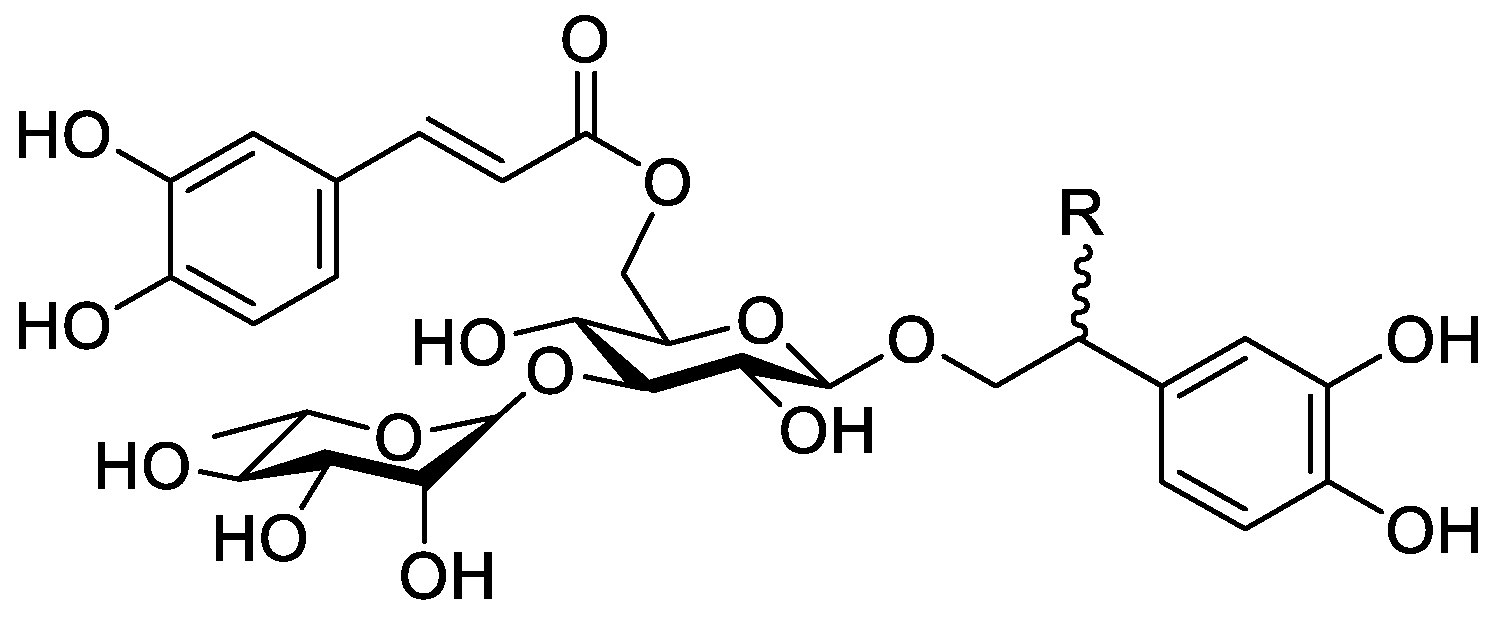
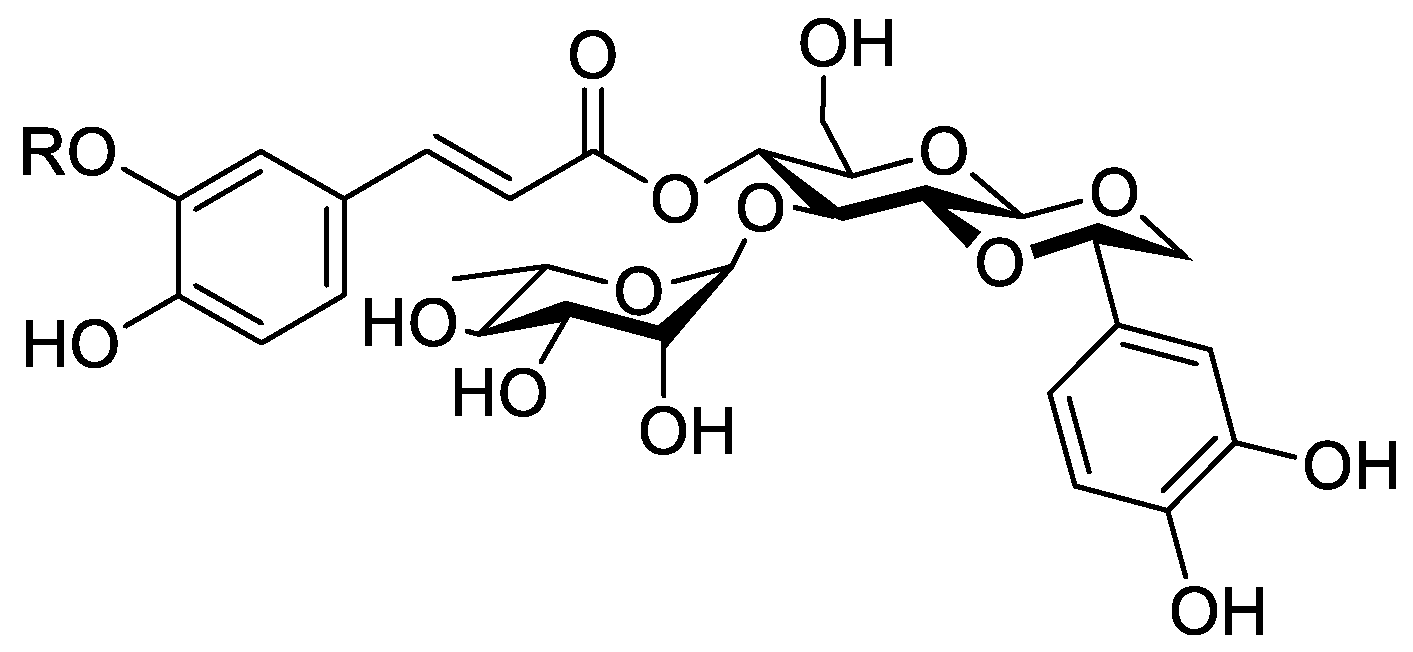
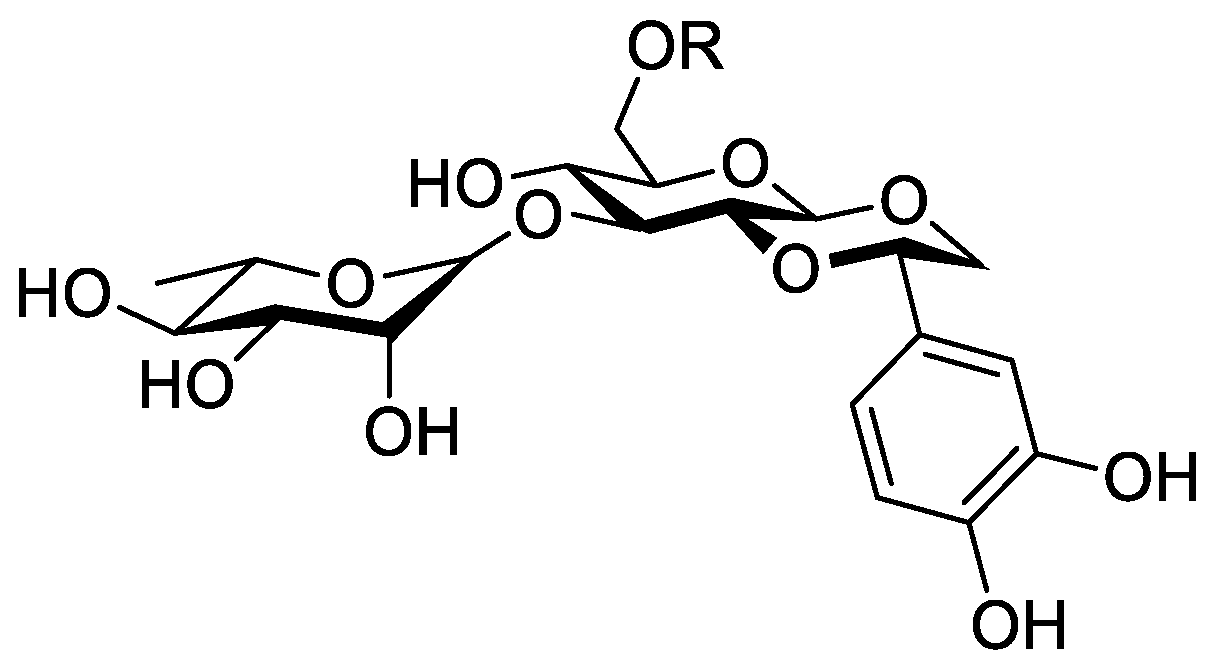
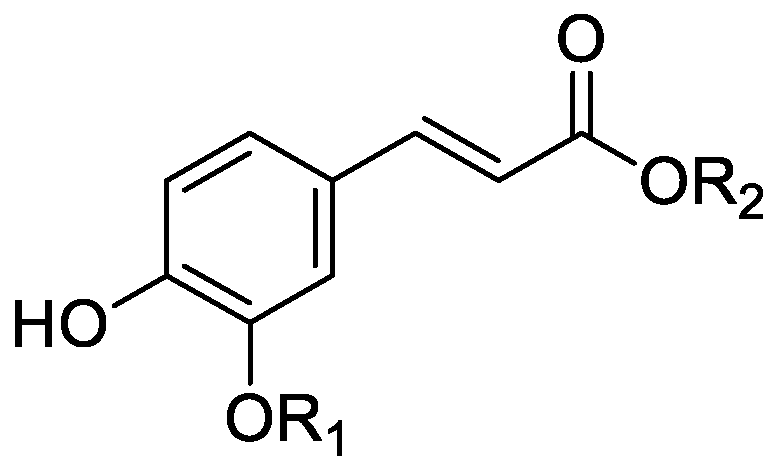
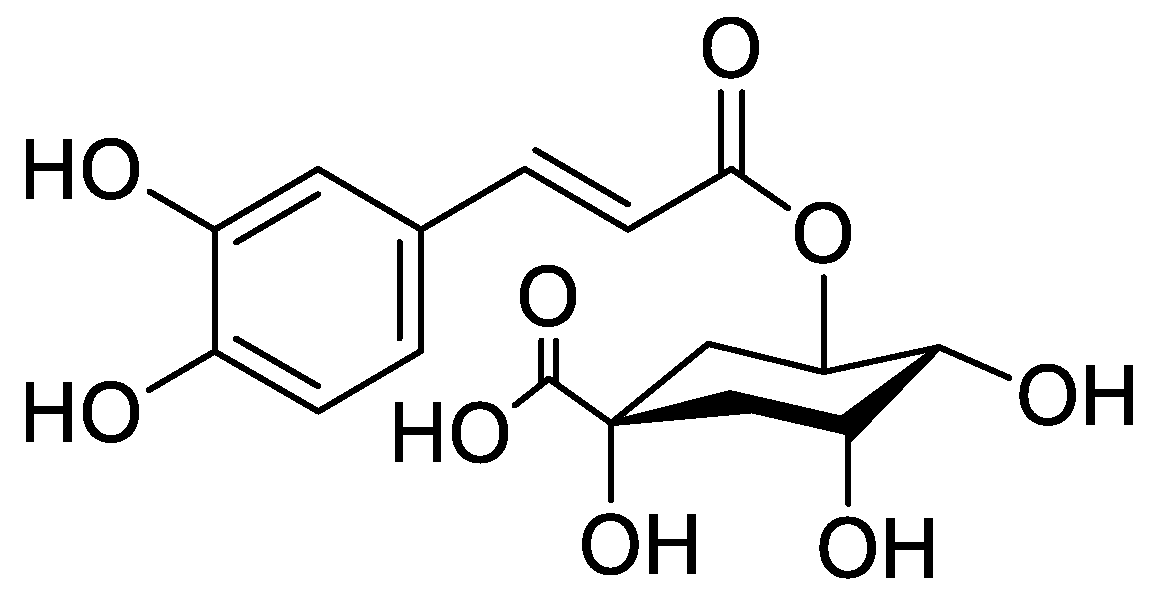
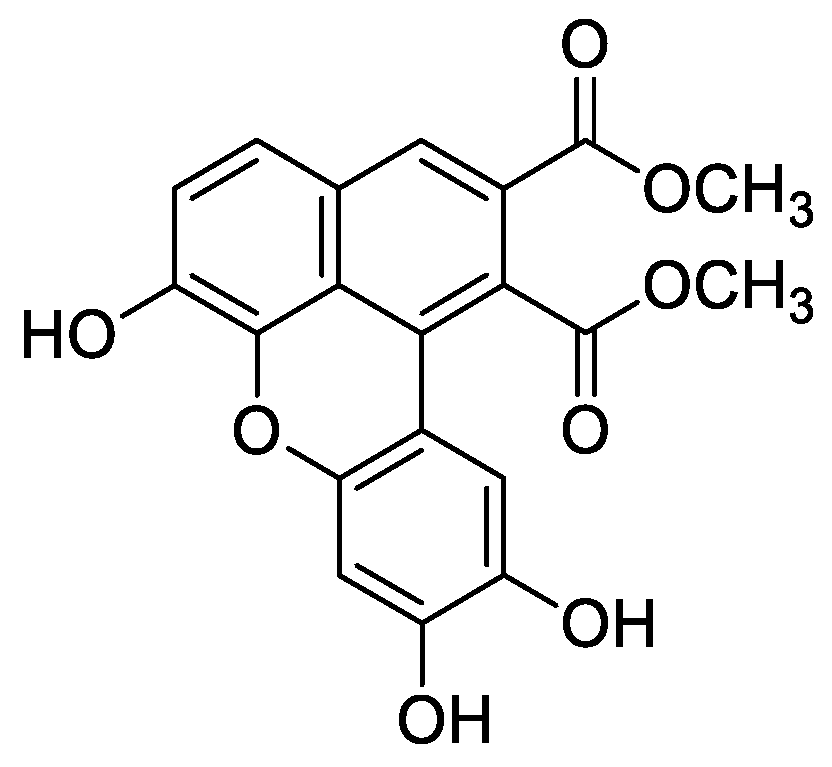
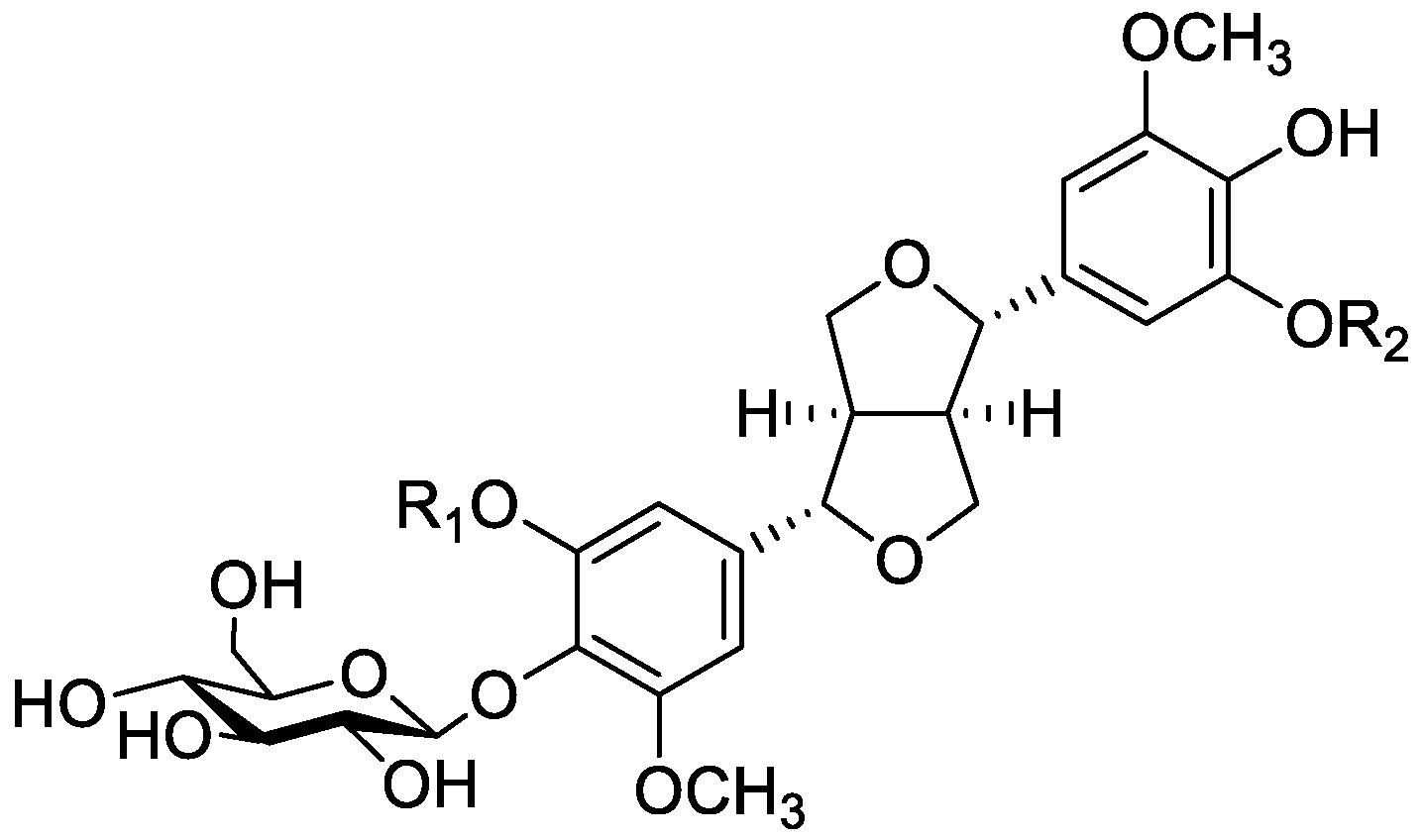
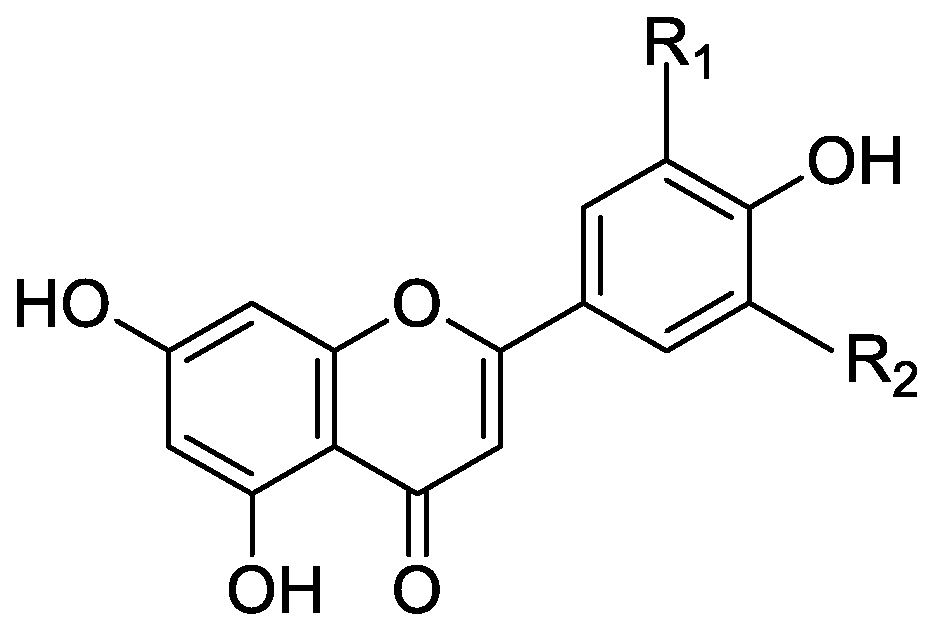
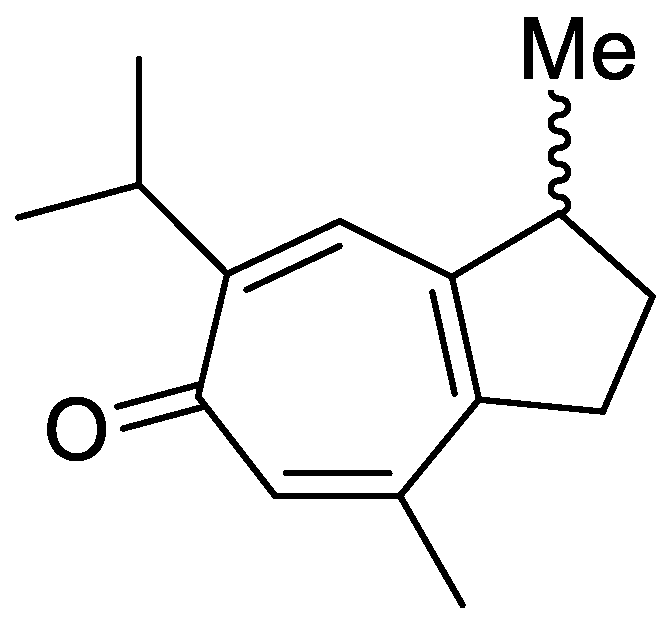
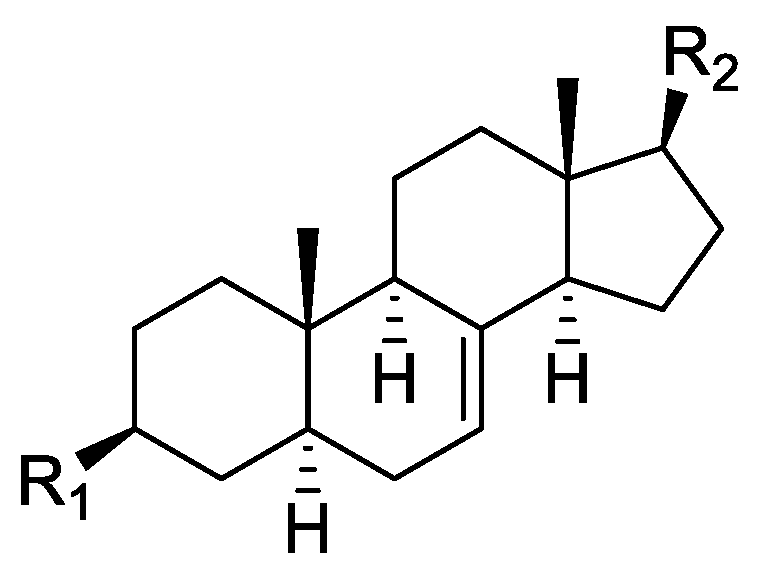
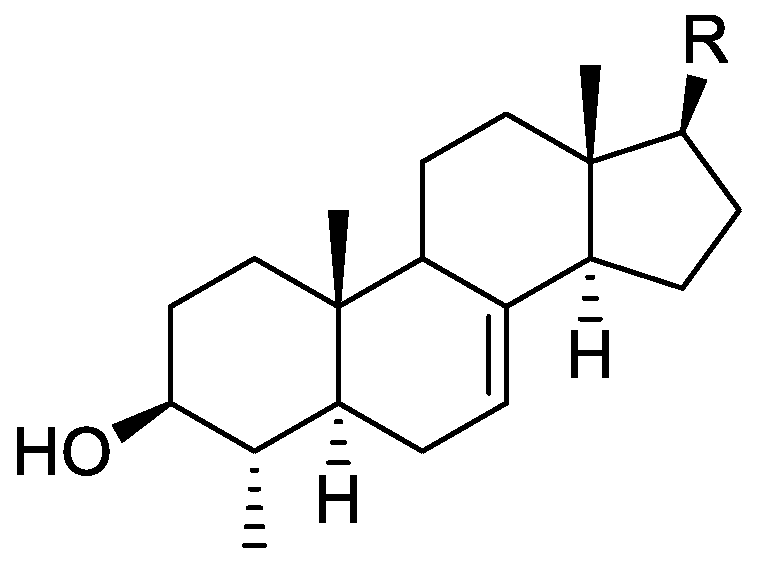
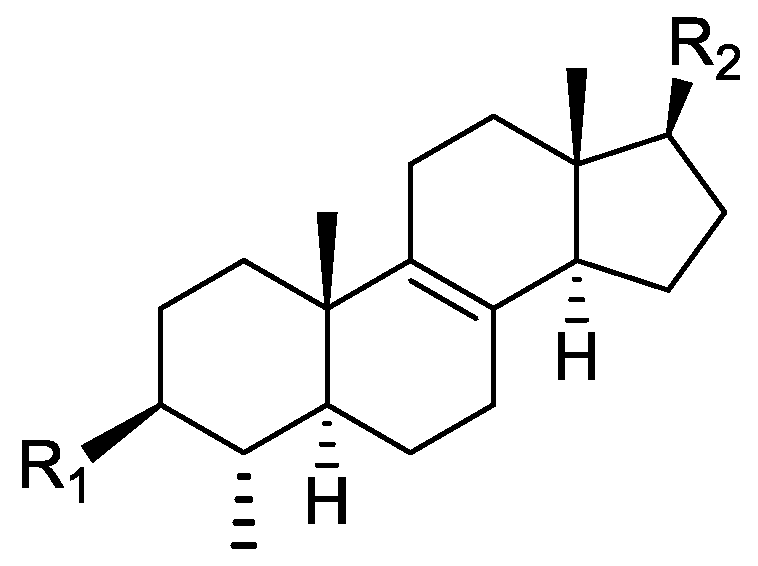
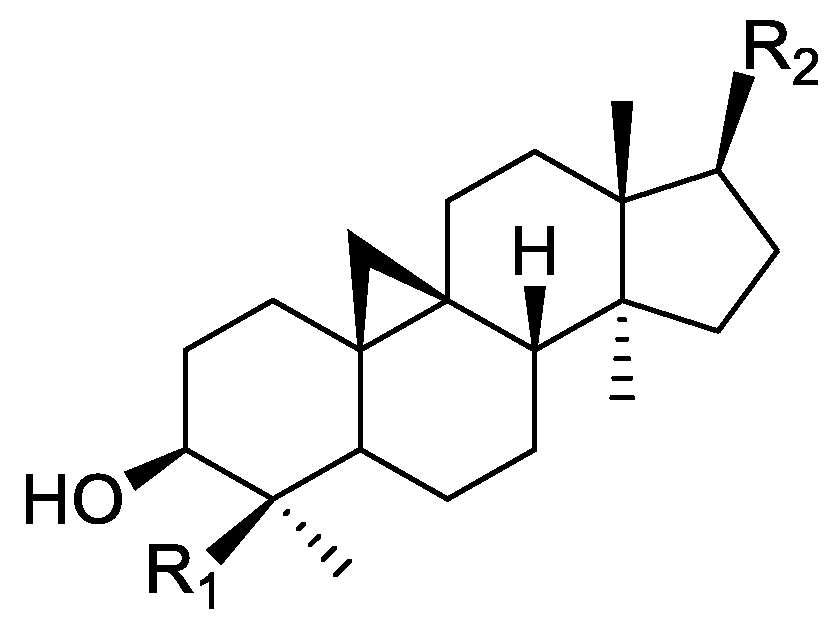
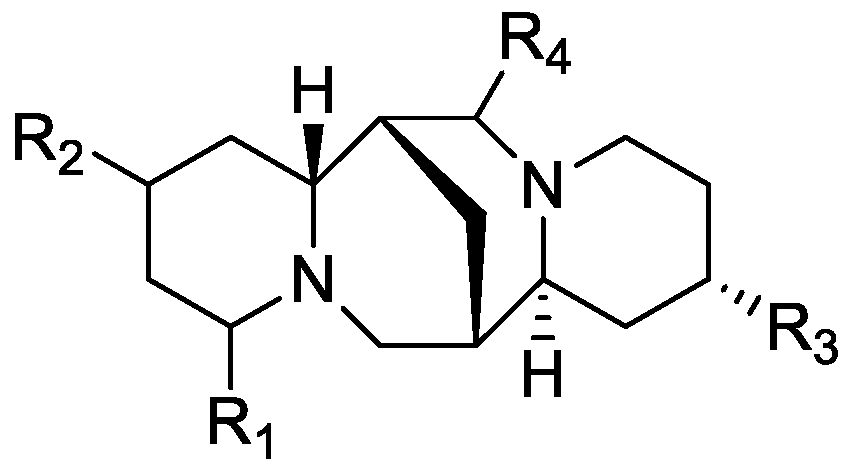
In the genus Orobanche s.l. the following classes of secondary metabolites have been found: aromatic aldehydes, ketones, and phenylmethanoids (Figure 1), phenylethanoids (Figure 2), phenylethanoid glycosides (Figure 3), phenylpropanoid glycosides (Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8), phenolic acids (Figure 9 and Figure 10), lignans (Figure 11 and Figure 12), flavonoids (Figure 13), a tropone derivative (Figure 14), and sterols (Figure 15, Figure 16, Figure 17, Figure 18, Figure 19 and Figure 20). Phenylpropanoid glycosides represent the largest group of secondary metabolites isolated from Orobanche species. Moreover, bi- and tricyclic sesquiterpenes, iridoid glycosides, acyclic, monocyclic, and bicyclic monoterpenes, and carotenoids have been detected in Orobanche. Along with secondary metabolites the isolation of some primary metabolites such as fatty acids, alkanes, alkenes, ketones, fatty alcohols, and sugar alcohols has been described. Species that have been investigated for their phytochemical compounds are subsequently listed in alphabetical order within their corresponding genus. The secondary metabolites synthesized by Orobanche species are shown in Figures 1–20 and compounds are numbered consecutively from 1 to 70. Components of essential oils are listed in the corresponding paragraphs but individual chemical structures are neither displayed nor numbered. Primary metabolites that have been isolated along with secondary metabolites and that are mentioned in the corresponding papers are only mentioned in the text (no figure, no numbers). Secondary metabolites sequestered from host species by Orobanche species, comprising alkaloids (Figure 21, Figure 22, Figure 23 and Figure 24), polyacetylenes (Figure 25), and cannabinoids (Figure 26), are shown in Figure 21, Figure 22, Figure 23, Figure 24, Figure 25 and Figure 26, and these sequestered compounds are numbered consecutively from S1 to S19. Information about collection site, country of origin, analyzed plant parts, and extraction solvents as well as analytical methods used for compound identification and structure elucidation are listed in this order in brackets after the corresponding taxon. Analytical methods include infrared spectroscopy (IR), gas chromatography (GC), gas liquid chromatography (GLC), mass spectrometry (MS), nuclear magnetic resonance spectroscopy (NMR), thin layer chromatography (TLC), high performance thin layer chromatography (HPTLC), and high performance liquid chromatography (HPLC) with ultraviolet (UV) and diode array detectors (DAD). Most authors do not specify the host species of the investigated Orobanche samples. Whenever available, information about the host species is mentioned. An additional section is dedicated to reports dealing with more than one Orobanche species. Natural products described in these reports are also briefly mentioned in the respective paragraphs for each individual species. An additional section is dedicated to the sequestration of secondary metabolites by Orobanche species from their host plants. Moreover, one section deals with bioactivities reported for Orobanche extracts and natural products isolated from such extracts. Synonyms of taxon and compound names are only indicated when being mentioned for the first time. Tables giving an overview of all phytochemically investigated species, natural products detected within these species and sequestered by these species, respectively, are available as Supplementary Material (Tables S1 and S2).
2.2. Secondary Metabolites Synthesized by Orobanche s.l.
2.2.1. Orobanche Sectio Orobanche (=sect. Osproleon Wallr. = Genus Orobanche L. s.str.)
Orobanche alba Stephan ex Willd.—Roudbaraki and Nori-Shargh identified forty different compounds in the essential oil from O. alba [province of Guilan, Iran; aerial parts; hydrodistillation; GC, GC-MS, comparison with authentic samples] [host species not mentioned] [11]. Detected compounds included three monoterpene hydrocarbons, twelve oxygenated monoterpenes, five sesquiterpene hydrocarbons, three oxygenated sesquiterpenes, one oxygenated diterpene, and sixteen non-terpenic compounds (aliphatic hydrocarbons, alcohols, ethers, aldehydes, ketones, carboxylic acids, and esters). The monoterpene fraction encompassed bornylangelate, p-cymene, limonene, γ-terpinene, p-menthone, 1,8-cineol, pinocamphone, linalool, (Z)-iso-citral, nerol, neral (syn. citral B, cis-isomer), geraniol, geranial (syn. citral B, trans-isomer), and geranylacetate; the sesquiterpene fraction included trans-caryophyllene (syn. β-caryophyllene), 6,9-guaiadiene, isobornyl-2-methyl-butyrate, δ-cadinene α-copaene, β-bourbonene, and caryophyllene oxide. The only diterpene detected was manool and the only phenylpropanoid methyl chavicol (syn. estragol). Detected non-terpene compounds comprised n-nonanal, neryl acetate, myristic acid, palmitic acid, linoleic acid, linolenic acid, 6-methyl-5-hepten-2-one, 6,10,14-trimethyl-2-pentadecanone, octadecane, heneicosane, docosane, tricosane, tetracosane, pentacosane, nonadecene, isobutyl phthalate, and bis(2-ethylhexyl)phthalate. Fruchier et al. reported the isolation of the tropone derivative orobanone (45) from O. alba (as O. epithymum DC.) [whole plants; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] [12].
Orobanche amethystea Thuill.—In O. amethystea Serafini et al. [Sardinia, Italy; flowering plants; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] detected the phenylpropanoid glycosides verbascoside (syn. acteoside, orobanchin, [13]) (10) and oraposide (syn. crenatoside and orobanchoside) (29) [14].
Orobanche anatolica Boiss. & Reut.—The occurrence of saponins in O. anatolica was mentioned by Aynehchi et al. [Iran; whole plant] [host species not mentioned] but no further specification of structures nor of any analytical methods were indicated in the report [15].
Orobanche artemisiae-campestris Gaudin subsp. picridis (F.W.Schultz) O. Bolòs, Vigo, Masalles & Ninot—Fruchier et al. isolated the tropone derivative orobanone (45) [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [no information about the host] from O. artemisiae-campestris subsp. picridis (using the synonym O. picridis F.W.Schultz) [12].
Orobanche caryophyllacea Sm.—The tropone derivative orobanone (45) was isolated from O. caryophyllacea (using the synonym O. major L.) by Fruchier et al. [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [no information about the host] [12].
Orobanche cernua Loefl.—Qu et al. characterized 17 compounds from O. cernua (syn. O. cumana Wallr., O. cernua var. cumana Wallr.) [Jilin province, China; fresh whole plants; methanol; MS, 1D and 2D NMR, comparison with literature data] [host species not mentioned] [17]. Eleven compounds were identified as phenylpropanoid glycosides [salidroside (8), acteoside (10), 2′-O-acetylacteoside (14), campneoside (18), leucosceptoside A (19), isoacteoside (23), isocampneoside (24), oraposide (29), 3‴-O-methylcrenatoside (30), descaffeoyl crenatoside (31), and isocrenatoside (32)]. Furthermore, three phenolic acids [caffeic acid (34), trans-ferulic acid (36), and chlorogenic acid (37)], one lignan [dimethyl-6,9,10-trihydroxybenzol[kl]xanthene-1,2-dicarboxylate (38)], and two flavonoids [apigenin (42) and luteolin (43)], were found. In a second report Qu et al. described the isolation of a novel phenylethanoid glycoside, 3′-O-methylisocrenatoside (33) as well as of the known compounds protocatechuic aldehyde (2) and methyl caffeate (35) from O. cernua [Jilin Province, China; fresh whole plant; methanol; IR, MS, NMR] [host species not mentioned] [18]. Yang et al. examined O. cernua [Neimenggu province, China; whole plant; ethanol 70%, reflux; HPLC-MS, NMR] [host species not mentioned] isolating twelve compounds: eight phenylpropanoid glycosides [acteoside (10), campneoside II (17), campneoside I (18), leucosceptoside A (19), isoacteoside (23), cistanoside F (27), oraposide (29), and isocrenatoside (32)]; three lignans [(+)-pinoresinol 4′-O-β-D-glucopyranoside (39), isoeucommin A (40), and (+)-syringaresinol 4′-O-β-D-glucopyranoside (41)], and one steroid [stigmasterol 3-O-β-D-glucoside (54)] [19].
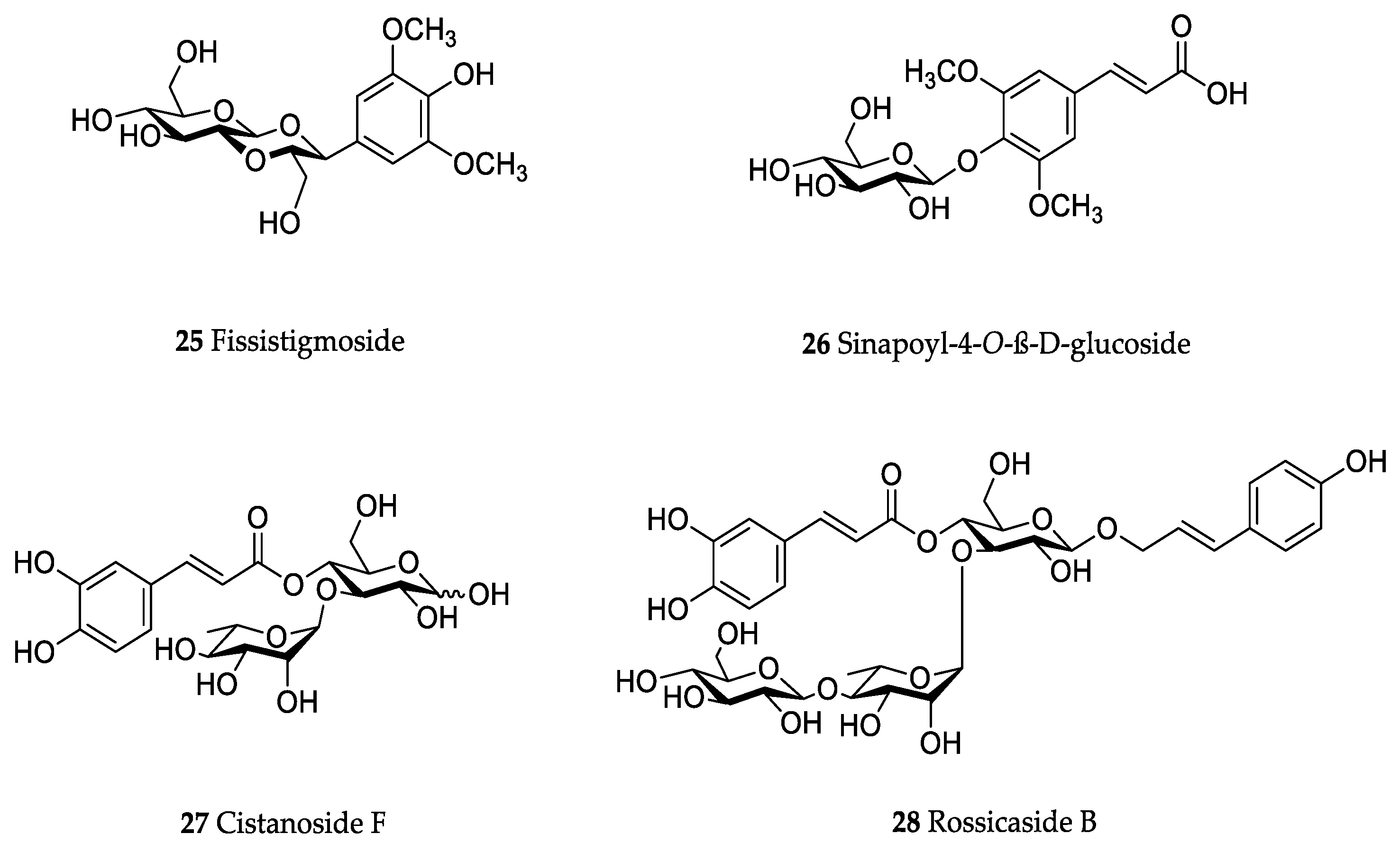
Orobanche coerulescens Stephan ex Willd.—Zhao et al. isolated the phenylpropanoid glycoside acteoside (10) from O. coerulescens [Xinjiang province, China; ethanol] [host species not mentioned] [20]. The isolation of isoacteoside (23), sinapoyl 4-O-β-D-glucoside (26), cistanoside F (27), oraposide (29), and adenosine was described in a second report [21] [Xinjiang province, China; rhizome; ethanol 95%, reflux; TLC, NMR] [host species not mentioned]. Acteoside (10), cistanoside F (27), and oraposide (29) were also isolated by Murayama et al. along with 2-phenylethyl β-primeveroside (7), rhodioloside (syn. salidroside) (8), descaffeoyl crenatoside (31), and isocrenatoside (32) from O. coerulescens [Niigata prefecture, Japan; whole plant; methanol; IR, UV, HR-FAB-MS, 1H,1H-DQF, COSY, 1H,1H-relayed COSY, HMQC, and HMBC NMR] [host species not mentioned] [22]. Lin et al. isolated two new phenylpropanoid glycosides, caerulescenoside (13) and 3′-methyl crenatoside (30), along with five known phenylpropanoid glycosides, desrhamnosyl acteoside (9), acteoside (10), campneoside (18), isoacteoside (23), and oraposide (29) from O. coerulescens [Taipei, Taiwan; whole plant; ethanol 95%; IR, UV, 1H NMR, 13C NMR, ESI-MS, FAB-MS, HR-FAB-MS] [host species not mentioned] [23]. In a second report the authors mentioned the occurrence of another phenylpropanoid glycoside, rossicaside B (28), in O. coerulescens; ethanol 95%, preparation in accordance with former report, no information about analytical methods) [24]. Wang et al. also isolated a new phenylpropanoid glycoside from the whole plant of O. coerulescens [Neimenggu province, China; whole plant; ethanol 50%; TLC, HPLC, MS, NMR] [host species not mentioned] [25]. The structure was identified as 2-(3-methoxy-4-hydroxy)-phenyl-ethanol-1-O-α-L-[(1→3)-4-O-acetyl-rhamnopyranosyl-4-O-feruloyl]-O-β-D-glucopyranoside and named orobancheoside A (22). Additionally, Zhao et al. described the isolation protocatechuic aldehyde (2) and caffeic acid (34) as well as of β-sitosterol (50) and daucosterol (53) from O. coerulescens [Xinjiang province, China; rhizome; ethanol 95%, reflux; NMR] [host species not mentioned] [26]. Shao et al. identified the phenylpropanoid glycosides acteoside (10) and oraposide (29), as well as sterols β-sitosterol (50), stigmasterol (51), and β-daucosterol (53) [Neimenggu province, China; ethanol 95%, reflux; TLC, MS, NMR] [host species not mentioned] [27]. Moreover, primary metabolites D-mannitol, glyceryl arachidate, succinic acid, and D-pinitol were reported. Recently, Zhang characterized a new phenethyl alcohol glycoside named orobancheoside B (21) from O. coerulescens [Neimenggu province, China; whole plant; ethanol 50%; IR, UV, MS, NMR] [host species not mentioned] [28].
Orobanche crenata Forssk.—El-Shabrawy et al. found two phenylpropanoid glycosides in O. crenata growing on Vicia faba L. [Egypt; chloroform for removal of non-polar compounds, aqueous ethanol 70%; TLC, NMR] [29]. However, their structures were not fully characterized. Afifi et al. extracted secondary metabolites from O. crenata parasitizing on V. faba [Mansoura, Egypt; aerial parts; ethanol 90%; melting point, TLC, IR, UV, 1H NMR, 13C NMR, 13C NMR-DEPT, FAB-MS] and isolated the known phenylpropanoid glycoside acteoside (10) as well as the new phenylpropanoid glycoside oraposide (29) [30]. Acteoside (10) was also extracted by Gatto et al. [parasitizing on V. faba; Apulia, Italy; stems; methanol 80% under reflux; comparison of UV spectra and retention times with standard substances, HPLC-DAD] who furthermore found isoverbascoside (syn. isoacteoside; an isomer of verbascoside) (23) and an unidentified caffeic acid derivative [31,32,33]. Serafini et al. isolated verbascoside (10), poliumoside (11), and orobanchoside (29) from O. crenata [Sardinia, Italy; flowering plants; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14]. Orobanchoside (29) was proven to be structurally identical with oraposide and oraposide by Nishibe et al. in the same year [34]. Nada and El-Chaghaby analyzed ethanolic (80%) extracts of O. crenata grown on V. faba by GC-MS and postulated the occurrence of 6-monohydroxyflavone, glycitein, actinobolin, hexestrol, and 2,4-di-tert-butylphenyl benzoate [Egypt; ethanol 80%; GC-MS] [35]. However, none of these compounds are considered any further here, because the data on the used GC system are incomplete and data on peak identification procedures are completely missing. Until contrary evidence will have been procured, we do not consider 6-monohydroxyflavone, glycitein, actinobolin, hexestrol, and 2,4-di-tert-butylphenyl benzoate natural products which have been detected in the genus Orobanche. Dini et al. investigated the phytochemistry of O. crenata using the synonym O. speciosa DC. [Molise, Italy; aerial parts; petroleum ether, chloroform, methanol; comparison of UV, IR, 1H and 13C NMR spectral data with literature data] [36]. The three phenylpropanoid glycosides verbascoside (10), poliumoside (11), and oraposide (29) were detected. Along with these compounds the authors also isolated p-hydroxy benzaldehyde (1), isovanillin (3), vanillin (4), syringaldehyde (5), and p-hydroxy acetophenone (6). Fruchier et al. reported the isolation of the tropone derivative orobanone (45) from O. crenata in their investigation of the occurrence of this secondary metabolite in various Orobanche species [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] (see extra paragraph and supplementary material) [12]. Abbes et al. described the extraction of polyphenols and tannins from O. crenata growing on V. faba without any further specification of the substances [Beja and Ariana Governorates, Tunisia; aerial parts; methanol; water] [37].
Orobanche denudata Moris—Serafini et al. isolated the phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) from O. denudata [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14].
Orobanche foetida Poir.—Abbes et al. described the extraction of polyphenols and tannins from O. foetida growing on V. faba without any further specification of the substances [Beja and Ariana Governorates, Tunisia; aerial parts; methanol; water] [37].
Orobanche gracilis Sm.—Fruchier et al. isolated the tropone derivative orobanone (45) from O. gracilis (using the synonym O. cruenta Bertol.) [whole plants; water, chloroform; IR, UV, MS, CI-MS, and 1H and 13C NMR] [no information about the host] [12].
Orobanche grisebachii Reut.—Aynilian et al. screened several Orobanche species for their contents of alkaloids, tannins, and saponins, without reporting any particular structures of the metabolites. O. grisebachii [plant material obtained from The Post Herbarium of the American University of Beirut, Lebanon; ethanol 95%, ethanol 80%] contained alkaloids and tannins [40].
Orobanche hederae Duby—Pieretti et al. isolated the phenylpropanoid verbascoside (10) and orobanchoside (29) from O. hederae [Lazio, Italy; whole plants; ethanol; 1H NMR, HPLC-UV] [host species not mentioned] [41]. Capasso et al. also found these two compounds [whole plants; ethanol] [host species not mentioned] [42]. As well as in eleven other species of the genus Orobanche (see extra paragraph) Fruchier et al. found the tropone derivative orobanone (45) in O. hederae [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] [12]. Baccarini and Melandri isolated and analyzed seven pigments from O. hederae growing on Hedera helix L. (Araliaceae) [whole plant; acetone 80%; TLC, comparison with standard substances]. These pigments were β-carotene, α-carotene-5,6-epoxide, flavochrome, lutein-5,6-epoxide, flavoxanthin, and taraxanthin [43]. The seventh compound was tentatively identified as neoxanthin. There is also a report of the sequestration of substances from its host species by O. hederae. This is described in a separate paragraph below.
Orobanche loricata Rchb.—The isolation of the phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) from O. loricata was described by Serafini et al. [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14]. Fruchier et al. isolated the tropone derivative orobanone (45) [whole plants; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [no information about the host] [12].
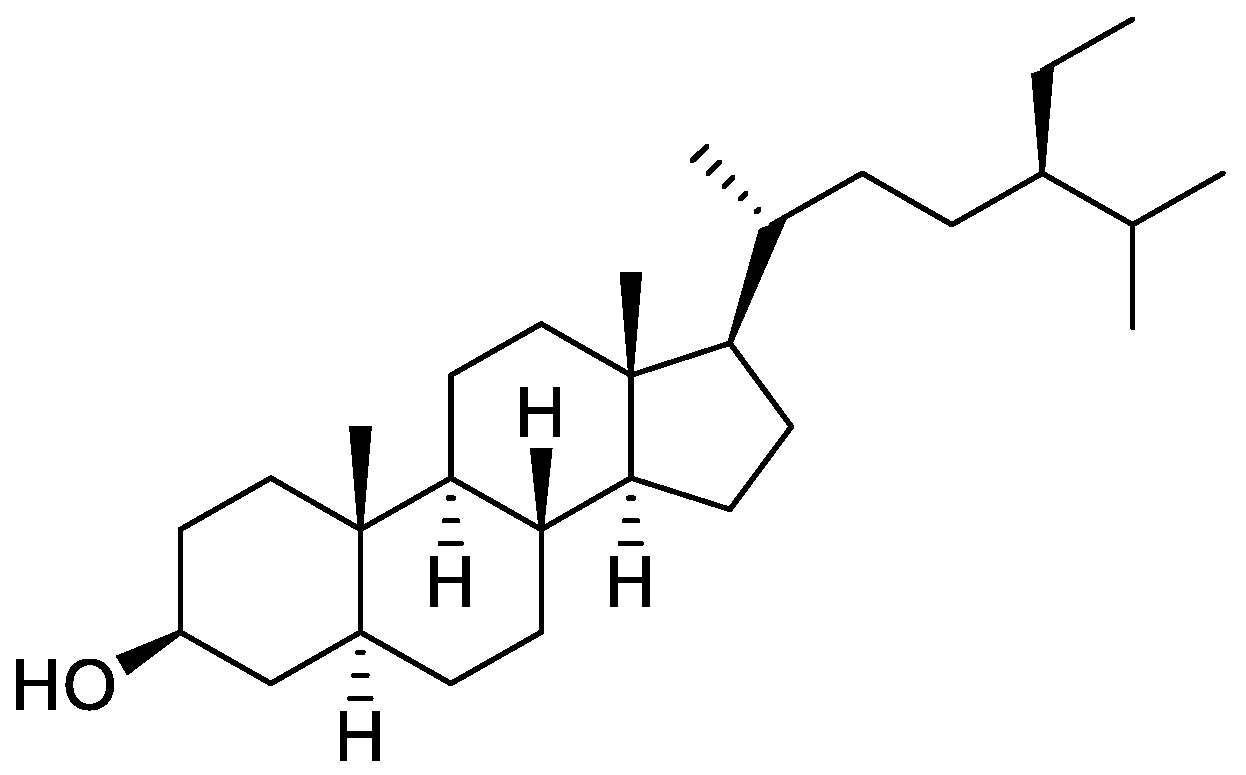
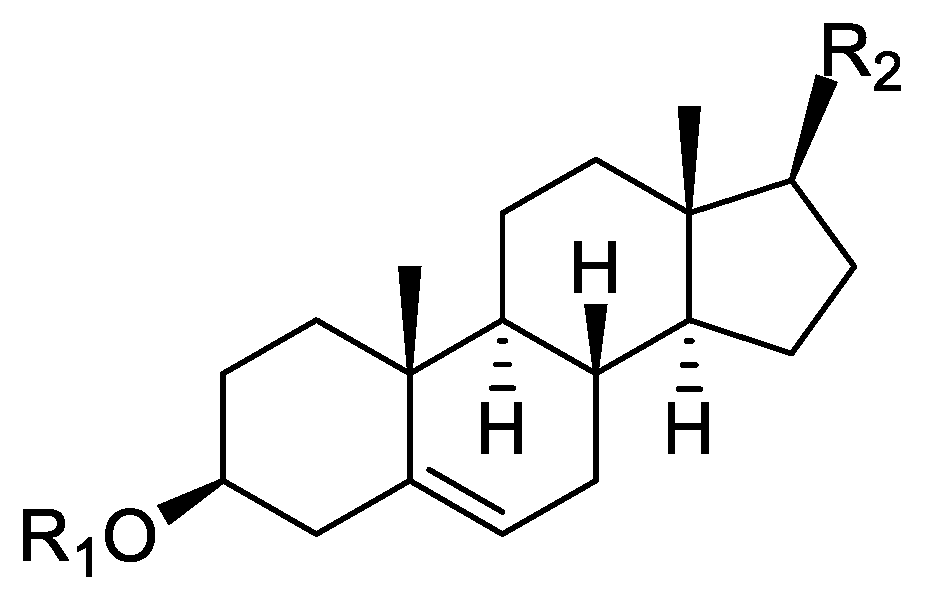
Orobanche lutea Baumg.—Rohmer et al. analyzed several parasitic species for their sterol contents and sterol biosynthesis [44]. The following compounds were isolated and identified from O. lutea [Alsace, France; stems; acetone, chloroform:methanol (2:1); GLC, GLC-MS] [host species not mentioned]: sterols [stigmastanol (46), cholesterol (47), campesterol (48), 24-methylene cholesterol (49), sitosterol (50), stigmasterol (51), isofucosterol (52), ∆7-campestenol (55), episterol (56), ∆7-stigmastenol (57), and ∆7-avenasterol (58)], 4α-methylsterols [24- 24-methyl lophenol (59), methylene lophenol (60), 24-ethyl lophenol (61), 24-ethylidene lophenol (62), 4α-methyl ∆8-campestenol (63), 4α-methyl ∆8-stigmastenol (64), 24,28-dihydro obtusifoliol (65), obtusifoliol (66), 24,28-dihydrocycloeucalenol (67), cycloeucalenol (68), and 4,4-dimethylsterols cycloartenol (69)], and 24-methylene cycloartenol (70).
Orobanche minor Sm.—Kurisu et al. isolated the phenylpropanoid glycosides acteoside (10) and oraposide (29) from O. minor [whole plant; methanol; NMR (1H, 13C, COSY, HMQC, HMBC, NOESY), HR-ESI-MS] [host species not mentioned] [45]. Serafini et al. also extracted verbascoside (10) and orobanchoside (29) [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14]. Kidachi et al. isolated acteoside (10), cistanoside D (20), isoacteoside (23), oraposide (29), 3‴-O-methyl crenatoside (30), and isocrenatoside (32) [whole plant; methanol] [host species not mentioned] [46]. The tropone derivative orobanone (45) was found in O. minor by Fruchier et al. [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] [12].
Orobanche owerinii Beck—Dzhumyrko and Sergeeva detected several carotenoids in O. owerinii [epigeal parts; n-hexane, petrol ether; co-chromatography with reference compounds and UV spectroscopy] growing on the hypogeal organs of Fraxinus [47]. Violaxanthin, auroxanthin, the ester of violaxanthin and palmitic acid, as well as α- and β- carotenes were detected.
Orobanche pubescens d´Urv.—Aynilian et al. screened several Orobanche species for their contents of alkaloids, tannins, and saponins, without investigating any particular structures of the metabolites. Tannins were found in O. pubescens (using the synonym O. versicolor F.W.Schultz) [plant material obtained from The Post Herbarium of the American University of Beirut, Lebanon; petroleum benzine (defatting), ethanol 95%, ethanol 80%] [40].
Orobanche pycnostachya Hance—Han et al. isolated eight compounds from O. pycnostachya [Neimenggu province, China; ethanol 95%; MS, NMR] [host species not mentioned] [48]. The primary metabolites n-nonacosane acid, n-hexacosyl alcohol, D-allitol, 2,3,4,6-α-D-galactopyranose tetramethyl ether, and secondary metabolites acteoside (10), fissistigmoside (25), β-sitosterol (50), and daucosterol (53) were identified. Li et al. also isolated the phenylpropanoid glycosides acteoside (10) as well as 2′-O-acetylacteoside (14), and oraposide (29) [Mengu, Anhui and Hebei, China; methanol 70%; HPLC] [host species not mentioned] [49].
Orobanche rapum-genistae Thuill.—Several secondary metabolites such as phenylproanoid glycosides, alkaloids, and a tropone derivative have been isolated from O. rapum-genistae. The isolated alkaloids are presumably not synthesized by the parasite itself but sequestered from the host species. Sequestration of secondary metabolites by O. rapum-genistae from its host is also reported by other authors and described in a separate paragraph below. Fruchier et al. isolated the tropone derivative orobanone (45) from O. rapum-genistae parasitizing on Cytisus scoparius (L.) Link and Cytisus purgans (L.) Spach (both Fabaceae) [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [12]. Eleven other Orobanche species were also screened for orobanone (45) (see separate paragraph and supplementary material). Several sources report the isolation of the two phenylpropanoid glycosides verbascoside (10) and oraposide (29) from O. rapum-genistae (Andary et al., [50,51] (host: C. scoparius, C. purgans), [39] [phenolic extract; hydrolysis, high resolution 1H and 13C NMR], [38] [1H and 13C NMR, X-ray crystal analysis]; Bridel and Charaux, [52,53] [tubers/bulbs; alcohol]; Serafini et al., [14] [Sardinia, Italy; flowering plants; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned]; Viron et al., [54] [whole plant; aqueous ethanol 70%; HPTLC, HPLC] [host species not mentioned], [55]). Andary et al. described the differentiation of two ecotypes of O. rapum-genistae: O. rapum-cytisi scoparii (parasitizing C. scoparius) and O. rapum-cytisi purgantis (parasitizing C. purgans) based on morphological and phytochemical characteristics [51]. Phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) as well as quinolizidine alkaloids sparteine (S2) and lupanine (S4) were found in both ecotypes in different concentrations.
Orobanche sanguinea C.Presl.—Serafini et al. isolated the phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) from O. sanguinea [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14].
Orobanche variegata Wallr.—Fruchier et al. isolated the tropone derivative orobanone (45) from O. variegata [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [no information about the host] [12].
2.2.2. Phelipanche (=Orobanche Sectio Trionychon)
Phelipanche aegyptiaca (Pers.) Pomel (formerly O. aegyptiaca Pers.)—Afifi et al. isolated acteoside (10), poliumoside (11), 2′-O-acetylacteoside (14), and 2′-O-acetylpoliumoside (15) from P. aegyptiaca (as O. aegyptiaca) [butanol extraction] [host species not mentioned [56]. Sharaf and Youssef described the extraction of alkaloids and tannins (organic acids, reducing sugars, glucosides resins, and unsaturated substances) from P. aegyptiaca (as O. aegyptiaca) [whole plant; 30% aqueous extract, hexane, chloroform, ethyl alcohol] [host species not mentioned] [57]. The extracted substances were not characterized any further.
Phelipanche arenaria Pomel (Syn.: O. arenaria Borkh.)—Andary et al. isolated the caffeic acid glycosides arenarioside (12) and pheliposide (16) from P. arenaria (as O. arenaria) growing on Artemisia campestris Ledeb. var. glutinosa (J.Gay ex Besser) Y.R.Ling [syn. of A. campestris subsp. glutinosa (Besser) Batt.] [Hérault, France; ethanol 80%; high resolution 1H and 13C NMR]. Orobanone (45) was found in P. arenaria (as O. arenaria) by Fruchier et al., (1981) [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] [12].
Phelipanche oxyloba (Reut.) Soják (Syn.: O. oxyloba (Reut.) Beck)—Aynilian et al. screened several Orobanche species for their contents of alkaloids, tannins, and saponins, without reporting fully characterized metabolites. Tannins were found in P. oxyloba (as O. nana Noe ex G.Beck) [plant material obtained from The Post Herbarium of the American University of Beirut, Lebanon; petroleum benzine (defatting), ethanol 95%, ethanol 80%] [40].
Phelipanche ramosa (L.) Pomel (Syn.: O. ramosa L.)—Lahloub et al. extracted known compounds acteoside (10) and 2′-O-acetylacteoside (14) as well as the formerly undescribed phenylpropanoid glycoside 2′-O-acetylpoliumoside (syn. brandioside, [14]) (15) from Phelipanche ramosa (using the synonym Orobanche ramosa) parasitizing on Lycopersicon esculentum Mill. [El-Behera Governorate, Egypt; whole plant; percolation in ethanol; comparison of melting point, IR, UV, 1H and 13C NMR with literature data] [58]. Serafini et al. isolated the phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) from P. ramosa subsp. ramosa (as O. ramosa subsp. ramosa) and P. nana (Reut.) Soják (as O.ramosa subsp. nana (Reut.) Cout., syn. O. nana (Reut.) Beck, according to [10]) as well as poliumoside (11) from P. nana [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14]. The tropone derivative orobanone (45) was isolated from P. ramosa by Fruchier et al. (as O. ramosa) [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [host species not mentioned] [12]. Afifi et al. also found poliumoside (11) in P. ramosa (as O. ramosa) [butanol extraction] [host species not mentioned] [56]. Two papers report tricin (44) (5,7,4′-trihydroxy-3,5′-dimethoxyflavone) from the seeds and aerial parts of P. ramosa (as Orobanche ramosa), respectively [59,60]. Compound identification [59]: spectra of the pigments and their acetates, Rf values, color properties on paper chromatograms and mixed melting points; [60]: UV and MS analyses, demethylation to tricetin and column chromatography]; host species not mentioned. Sequestration of secondary metabolites from its host by P. ramosa is described in a separate section below.
2.2.3. Reports Describing the Investigation of more than One Species
Besides investigating the phytochemical composition of one species there are also several reports dealing with more than one species. Serafini et al. investigated the secondary metabolite contents of several Orobanche species [Sardinia, Italy; flowering samples; alcoholic extract; 1H and 13C NMR, HPLC, co-elution of extracts with isolated and identified phenylpropanoid glycosides] [host species not mentioned] [14].
The authors reported the occurrence of the phenylpropanoid glycosides verbascoside (10) and orobanchoside (29) in O. amethystea, O. crenata, O. denudata, O. hederae, O. loricata, O. minor, O. rapum-genistae subsp. rigens (Loisel.) Arcang. (as O. rigens Loisel. [10]), O. sanguinea, P. nana (as O. ramosa subsp. nana), and P. ramosa subsp. ramosa (as O. ramosa subsp. ramosa). In O. crenata and P. nana poliumoside (11) was additionally reported. Moreover, the isolation of verbascoside (10), 2′-O-acetylacteoside (14), and 2′-O-acetylpoliumoside (15) from P. ramosa (as O. ramosa) [58], and pheliposide (16) and orobanchoside (29) from O. arenaria [61] were mentioned in the report from Serafini et al. [14]. The reference about orobanchoside (29) occurring in O. arenaria by Andary et al. [61] is questionable because in the named report arenarioside (12) is described instead of orobanchoside (29). Fruchier et al. describe the isolation of the guaian type sesquiterpene tropone derivative orobanone (3,8-dimethyl-S-isopropyl-2,3-dihydro(lH)azulen-6-one) from O. rapum-genistae parasitizing on C. scoparius and C. purgans [whole plant; water, chloroform; IR, UV, MS, CI-MS, 1H and 13C NMR] [12]. No traces of orobanone were found in the hosts. This compound was also found in eleven other Orobanche species: O. alba (as O. epithymum), O. arenaria, O. artemisiae-campestris subsp. picridis (as O. picridis), O. caryophyllacea (as O. major), O. crenata, O. gracilis (as O. cruenta), O. hederae, O. loricata, O. minor, O. ramosa, and O. variegata. Except for O. rapum-genistae, no information about the host species were indicated.
Aynilian et al. [plant material obtained from The Post Herbarium of the American University of Beirut, Lebanon; petroleum benzine (defatting), ethanol 95%, ethanol 80%] screened several Orobanche species for their contents of alkaloids, tannins, and saponins without reporting any defined chemical compounds [40]. Tannins were found in all six analyzed species, O. aegyptiaca, O. crenata, O. grisebachii, O. oxyloba (as O. nana), O. pubescens (as O. versicolor), and P. ramosa (as O. ramosa). Saponins were absent in all investigated taxa, and alkaloids were detected in O. crenata, O. grisebachii, O. pubescens, and O. ramosa.
2.3. Sequestration of Secondary Metabolites by Orobanche s.l. from Their Host Species
As obligate holoparasites Orobanche species drain water and essential nutrients from their host plants. Also the sequestration bioactive natural products from the hosts seems likely [62], but up to now little is known about the uptake of other substances by holoparasitic Orobanche species from the plants they parasitize on and only very few reports deal with the sequestration of secondary metabolites from hosts.
2.3.1. Orobanche Section Orobanche (=sect. Osproleon Wallr. = Genus Orobanche L. s.str.)
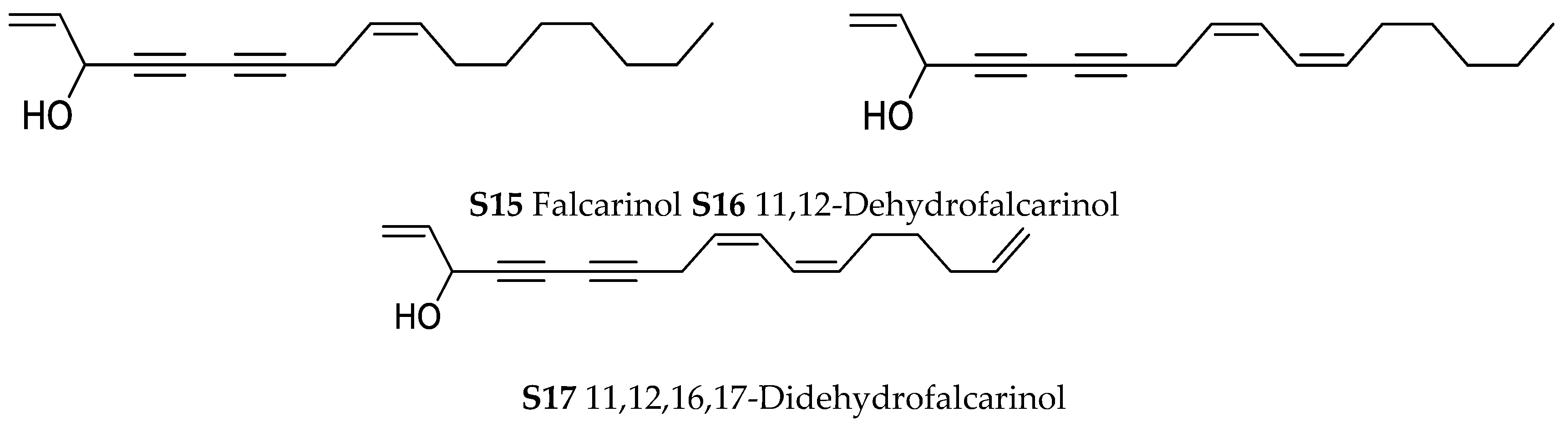
Orobanche hederae Duby—Sequestration of minerals and fatty acids by Orobanche hederae from its host Hedera helix was described by Lotti and Paradossi [Tuscany, Italy; whole plants; petrol ether, soxhlet; GC [63,64]. Sareedenchai and Zidorn reported the uptake of polyacetylenes falcarinol (S15), 11,12-dehydrofalcarinol (S16) and 11,12,16,17-didehydrofalcarinol (S17) from the roots of Hedera helix by Orobanche hederae [Trentino-Alto Adige, Italy; whole plants; dichloromethane; HPLC-DAD, HPLC-MS, comparison with authentic reference compounds and literature data] [65].
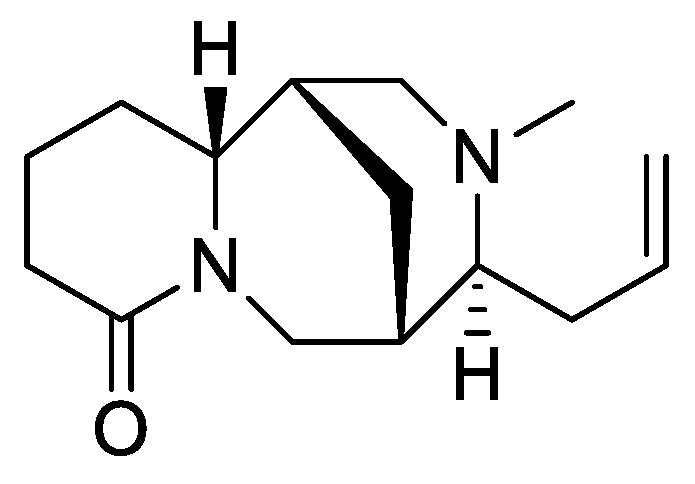
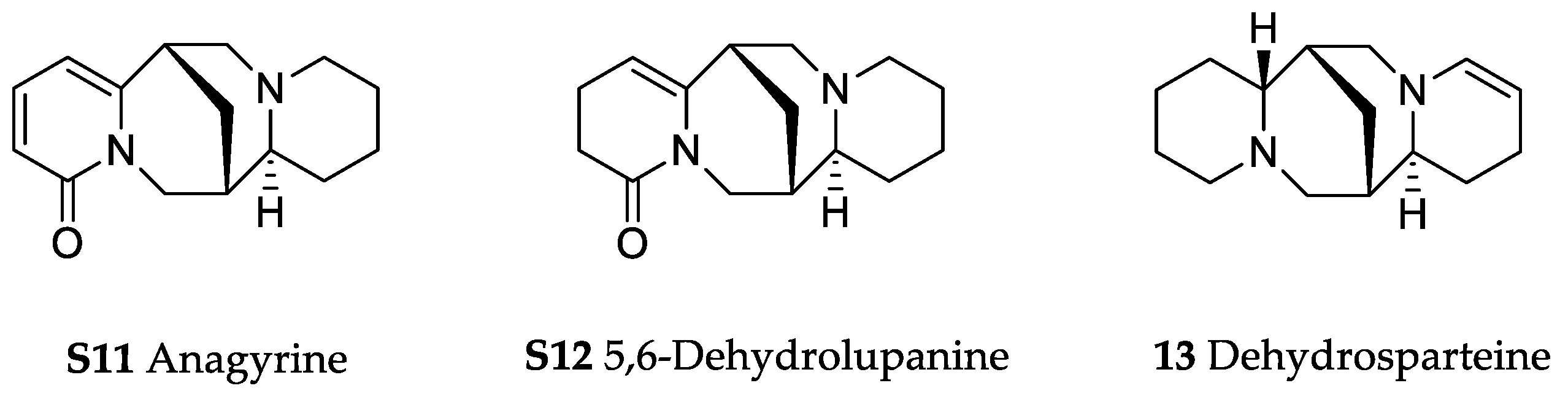
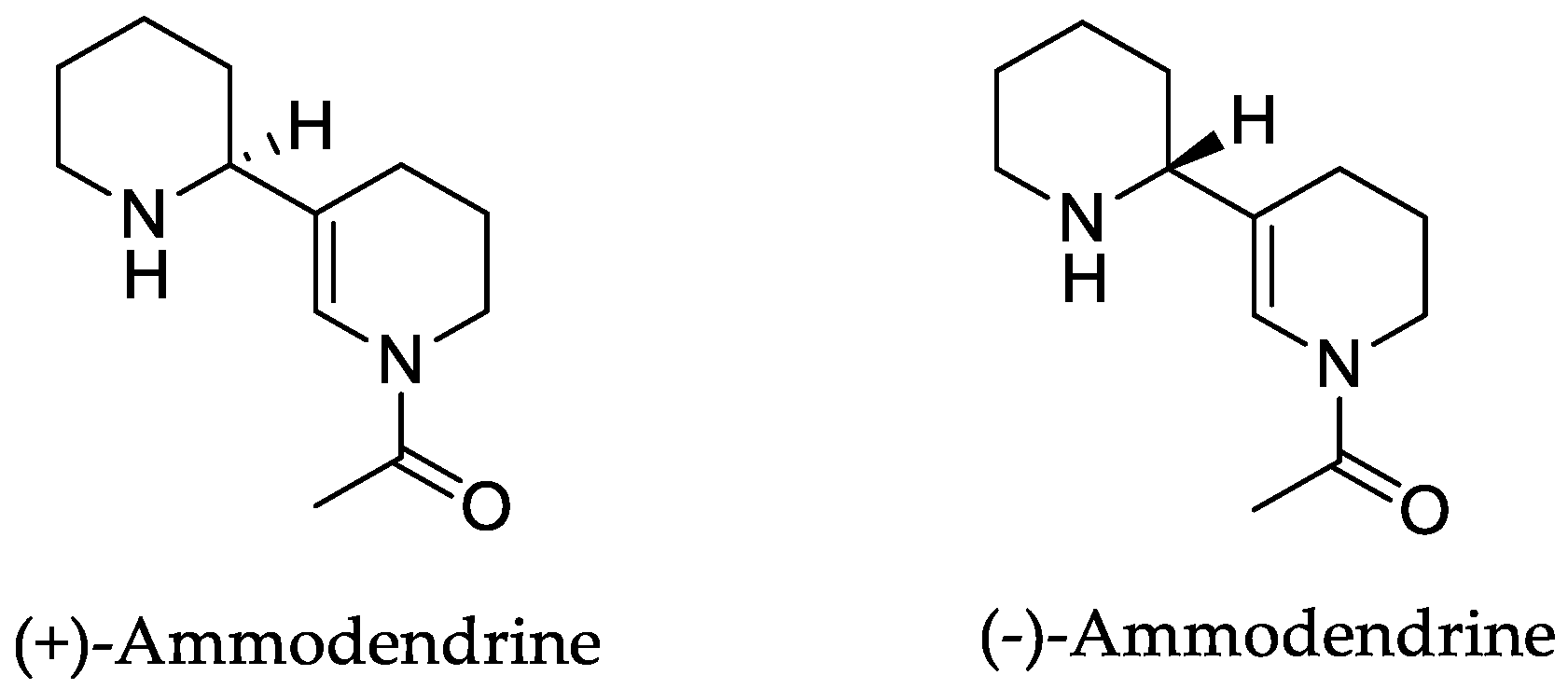
Orobanche rapum-genistae Thuill.—Wink et al. reported O. rapum-genistae sequestering quinolizidin alkaloids N-methylangustifoline (S1), sparteine (S2), 17-oxosparteine (S3), lupanine (S4), 17-oxolupanine (S5), hydroxylupanine (S6), 13-tigloyloxylupanine (S7), 13-angeloyloxylupanine (S8), 13-benzoyloxylupanine (S9), 4-hydroxylupanine (S10), anagyrine (S11), 13-5,6-dehydrolupanine (S12), and dehydrosparteine (S13) as well the piperidine alkaloid ammodendrine (S14) from Cytisus scoparius [as Sarothamnus scoparius (L.) W.D.J. Koch] [Rhineland-Palatinate, Germany; stems, leaves, pods, seeds, bulbs, shoots; HCl, alkalization, Extrelut-column, dichloromethane; GLC/GLC-MS, comparison with retention indices of known reference alkaloids] [66]. From the report it is not clearly visible whether 13-tigloyloxylupanine (S7), 13-angeloyloxylupanine (S8), 13-benzoyloxylupanine (S9), and 4-hydroxylupanine (S10) are present in the α- or the β-form, respectively. Rascol et al. also described the isolation of quinolizidine alkaloids (−)-sparteine (S2), (+)-lupanine (S4), and (+)-13-hydroxylupanine (S6) from O. rapum-genistae growing on Cytisus [67]. In a second report the authors investigated the effects of alkaloid contents of the host (Cytisus scoparius and Cytisus purgans) on the alkaloid contents of the parasite and vice versa [68].
2.3.2. Phelipanche (=Orobanche Section Trionychon)
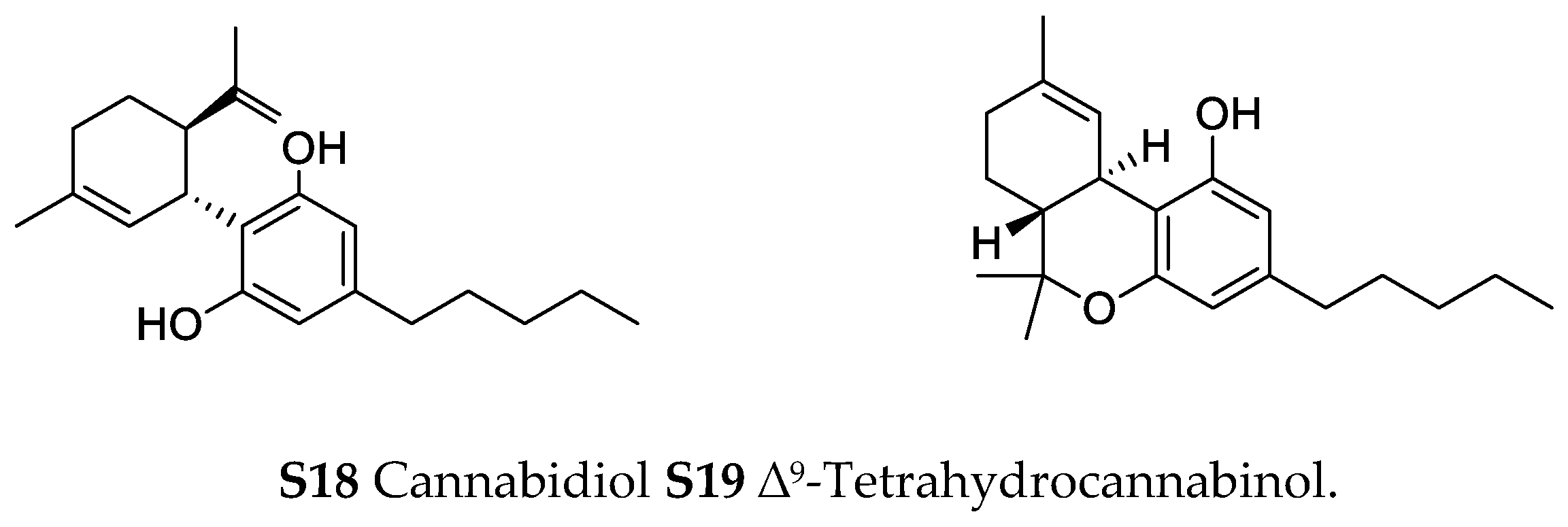
Phelipanche ramosa (L.) Pomel (syn.: Orobanche ramosa L.)—Cannabinoids cannabidiol (S18) and ∆9-tetrahydrocannabinol (S19) were found by Fournier and Paris in traces in Orobanche ramosa growing on Cannabis sativa L. (Cannabaceae) [petrolether; TLC, GC-MS] [69].
2.4. Sequestration of Secondary Metabolites from Host Plant Species by Other Hemiparasitic and Holoparasitic Plant Taxa
Sequestration is defined as the uptake, accumulation, and eventual use of substances, especially toxins, by animals or plant species from other organisms (microbes, plants or other animals) [70]. Most reports deal with interactions between plants and herbivorous insects, but sequestration is also known from some gastropods (feeding on e.g., algae, sponges, and bacteria) as well as from hemiparasitic and holoparasitic plants [71]. In most reported cases toxic secondary plant metabolites such as cardenolides, iridoid glycosides, and pyrrolizidine alkaloids or their precursors are sequestered by herbivorous insects and utilized as a defense against predators. Insects actively sequestering toxic compounds have, during their evolution, developed mechanisms to maintain the chemical stability of the sequestered toxins, while tolerating these toxins, without being negatively affected by their toxicity [72,73]. Sequestered metabolites can play a role in reproduction when utilized as sex pheromones [74]. There are also reports of sequestration of non-toxic substances like carotenoids or flavonoids by herbivorous insects [75]. Besides plant-animal interactions, some authors describe plant-plant interactions. Mainly alkaloids, but also iridoid glycosides, cardenolides, cardiac glycosides, and glucosinolates have been described to be sequestered. Most of the parasitic taxa analyzed so far are hemiparasites. Sequestration of iridoid glycosides by the hemiparasite Euphrasia stricta J.F.Lehm (Orobanchaceae, formerly Scrophulariaceae) from its host Galium verum L. (Rubiaceae) [76], and by the hemiparasite Castilleja integra A.Gray (Orobanchaceae) from Penstemon teucrioides Greene (Plantaginaceae) were described [77]. Pyrrolizidine and quinolizidine alkaloids are sequestered by hemiparasitic Castilleja Mutis ex L.f. species (Orobanchaceae, formerly Scrophulariaceae) from their hosts Senecio atratus Greene and Senecio triangularis Hook. (both Asteraceae), Lupinus L. species (Fabaceae), and Thermopsis montana Torr. & A.Gray (Fabaceae) [78,79,80]. Quinolizidine alkaloids are sequestered by Orthocarpus Nutt. species (Orobanchaceae) from Lupinus species, and by holoparasitic Cuscuta L. species (Convolvulaceae) from Genista acanthoclada DC. (Fabaceae), Lupinus, Cytisus L., and Spartium L. species (all Fabaceae) as well as from several other Fabaceae species [81,82,83]. Hemiparasitic Pedicularis L. species (Orobanchaceae, formerly Scrophulariaceae) were reported to sequester alkaloids from Senecio L. and Thermopsis R.Br. species as well as from Picea engelmannii Parry ex Engelm. (Pinaceae) [84]. The uptake of norditerpenoid alkaloids by Castilleja sulphurea Rydb. from Delphinium L. species (Ranunculaceae) was described. Hemiparasitic Tristerix verticillatus (Ruiz & Pav.) Barlow & Wiens (Loranthaceae) was found to take up isoquinoline alkaloids from Berberis montana Gay (Berberidaceae), and bipiperidyl and quinolizidine alkaloids synthesized by Retama sphaerocarpa (L.) Boiss. and Lygos sphaerocarpa (L.) Heyw. (both Fabaceae) were found to be sequestered by hemiparasitic Viscum cruciatum Sieber ex Boiss. (Santalaceae) [85,86,87,88,89]. An interesting study of community-level interactions described the transfer of alkaloids produced by an endophytic fungus [Neotyphodium uncinatum (W. Gams, Petrini & D. Schmidt) Glenn, C.W. Bacon & Hanlin) (Clavicipitaceae)] via the host grass Festuca pratensis Huds. (using the synonym Lolium pratense (Huds.) Darbysh.) (Poaceae) to the hemiparasite Rhinantus serotinus (Schönh.) Oborny (Orobanchaceae). The hemiparasite utilizes the alkaloids for protection against the herbivorous aphid Aulacorthum solani (Aphididae) [90]. Furthermore the transfer of glucosinolates from Arabidopsis thaliana (L.) Heynh. (Brassicaceae) to holoparasitic Cuscuta gronovii Willd. ex Roem. & Schult. (Convolvulaceae), the uptake of cardiac glycosides by hemiparasitic mistletoes [Muellerina celastroides (Schult. & Schult. f.) Tiegh., as Phrygilanthus celastroides (Schult. & Schult. f.) Eichler], Dendrophthoe falcata (L.f.) Ettingsh., and Amyema congener (Sieber ex Schult. & Schult.f.) Tiegh. (all Loranthaceae) from Nerium oleander L. (Apocynaceae), and the sequestration of cardenolides by holoparasitic Cuscuta species from Digitalis lanata Ehrh. and Digitalis purpurea L. (both Plantaginaceae) (after artificial infection) [91,92,93] were described as examples for plant-plant interactions.
2.5. Bioactivities Reported for Extracts Obtained from Orobanche Taxa and for Natural Products Isolated from Orobanche Taxa
Furthermore, biological activities of extracts or isolated pure compounds have been reported for some Orobanche species. Bioactivities of Orobanche species or of Orobanche natural products are compiled and listed alphabetically by activity in the following paragraphs.
2.5.1. Analgesic Effects
Potent analgesic effects of the phenylpropanoid containing fraction of O. crenata extract (oral application) were observed in mice using the hot plate method for testing. O. crenata phenylpropanoid containing fraction was administered in doses of 50 mg/100 g body weight (b.w.) and 100 mg/100 g b.w. Paracetamol was used for comparison with a dose of 50 mg/100 g b.w. Reaction times were measured right after application and after 10, 20, 30, 45, 60, and 120 min after application. At the beginning of the experiment (0 min) the control group without any analgetic treatment, the O. cernua 50 mg/100 g b.w., O. cernua 100 mg/100 g b.w., and the paracetamol group showed reaction times of 16.1 ± 0.0, 17.8 ± 0.6, 14.3 ± 3.0, and 16.0 ± 1.3 s, respectively. Reaction times of the control group during the experiment varied slightly between 14.1 ± 1.1 s and 16.3 ± 0.1 s. Reaction times after application of 50 mg/100 g b.w. O. cernua extract increased from 17.8 ± 0.6 s at 0 min to reach their maximum of 22.6 ± 0.9 s after 20 min and decreased again afterwards. After application of 100 mg/100 g b.w. O. cernua extract, reaction times increased from 14.3 ± 3.0 s at 0 min to 33.7 ± 1.7 s after 45 min, and then decreased again. Paracetamol application (50 mg/100 g b.w.) led to an increase of the reaction time from 16.0 ± 1.3 s at 0 min to 28.4 ± 0.7 s after 45 min, followed by a decrease. The results implicate a clear analgesic effect of O. cernua phenylpropanoid containing fraction after oral application in mice [29]. Other studies on pharmacological effects of phenylpropanoids also revealed analgesic effects, especially of acteoside (10), one of the main phenylpropanoid glycosides found in O. species. Acteoside showed an analgesic potency almost equal to aminopyrine when tested in mice [94], and was effective against neuropathic pain in rats [95].
2.5.2. Antimicrobial Activities
The phenolic composition of O. crenata 80% methanolic extract and its in vivo efficacy against fungal postharvest diseases were studied in an attempt to find new strategies for reducing postharvest diseases in sweet cherry fruit and replacing or integrating the use of synthetic fungicides. Sweet cherry fruit were sprayed with O. crenata extract (different concentrations: 1×, 2×, 4×; the 1× concentration corresponding to 0.170 mg dry matter/mL of buffer), O. crenata extract added with salts (CaCl2 or NaHCO3, 1% w/v), salt solutions, and the same buffer solution used to prepare the plant extracts (0.1 M K-phosphate, pH 5.5) as a positive control few hours after harvesting. Afterwards they were stored under controlled conditions. Rot incidence, expressed as the percentage of rotten fruit with respect to the total number of fruit in each tray, was assessed daily. At a rot incidence of around 50% the inhibition values of different treatments were evaluated. O. crenata extract inhibited postharvest rot in higher extract concentrations. An increase in extract concentration produced an increase in the percentage of inhibition from 64% to 76% for O. crenata. Addition of salt to the most concentrated extract further increased the inhibition of postharvest rot to 82% and 84% for NaHCO3 and CaCl2, respectively, and hereby proved to have a high antifungal efficacy [33]. Antifungal activity of O. aegyptiaca ethanolic and acetone extracts against Fusarium oxysporum Schlechtend., Cladosporium harbarum (Pers.) Link, Trichothecium roseum (Pers.) Link, and Trichoderma viride Pers. have been described by Nagaraja et al. [96]. Saadoun et al. tested O. aegyptiaca, O. cernua, and O. crenata ethanolic extracts against Agrobacterium tumefaciens (Smith & Townsend) Conn and Erwinia plant pathogens using the hole-plate diffusion method, the dilution method, and the Bauer–Kirby method. Tobromycin (10 μg), augmentin (30 μg), norfloxacin (10 μg), streptomycin (10 μg), ofloxacin (5 μg), cefuroxime (30 μg), and cefotaxime (30 μg) were tested as standard antibiotics for comparison. O. cernua showed inhibitory effects on A. tumefaciens isolates with MIC of 12,500 µg/mL, which is equal to streptomycin (10 μg), ofloxacin (5 mg) norfloxacin (10 μg), and cefotaxime (30 μg) activity against Agrobacterium. Erwinia isolates were less sensitive and higher concentrations of Orobanche extracts were needed for growth inhibition. O. aegyptiaca did not show antimicrobial activities in this study [97]. Nada et al. evaluated the antimicrobial activity of O. crenata extract against three gram positive bacteria (Staphylococcus aureus Rosenbach, Bacillus subtilis (Ehrenberg) Cohn, and Streptococcus faecalis (Andrewes & Horder) Schleifer & Kilpper-Bälz) and three gram negative bacteria Escherichia coli (Migula) Castellani & Chalmers, Pseudomonas aeruginosa Migula, and Neisseria gonorrhoeae (Zopf) Trevisan) using the disc diffusion assay. O. crenata showed moderate antibacterial activity against the investigated bacteria [35]. Abbes et al. tested antimicrobial activities of O. crenata and O. foetida methanolic and aqueous extracts using the disc diffusion method. Bacteria tested were P. aeruginosa, E. coli, Enterococcus faecalis (Andrewes & Horder) Schleifer & Kilpper-Bälz, Enterobacter cloacae (Jordan) Hormaeche & Edwards, Salmonella enterica (ex Kauffmann & Edwards) Le Minor & Popoff (subspecies: Salmonella typhi (Salmonella enterica subsp. enterica Serovar Typhi), Salmonella enteritidis (Salmonella enterica subsp. enterica Serovar Enteritidis), Salmonella salamae (Salmonella enterica subsp. salamae), Shigella flexneri Castellani & Chalmer, S. aureus, Streptococcus pyogenes Rosenbach, Listeria monocytogenes (Murray et al.) Pirie, Yersinia enterocolitica (Schleifstein & Coleman) Frederiksen, Proteus mirabilis Hauser, Bacillus cereus Frankland & Frankland, and B. subtilis. O. crenata methanolic extract was active against all tested bacteria except for S. aureus, O. foetida methanolic extract inhibited only S. enteritidis and L. monocytogenes. Aqueous extracts were not active againts the tested bacteria [37]. Antibacterial activity of an ethanolic extract of O. cernua at a concentration of 100 mg/mL in distilled water against five different bacteria species, four gram positive bacteria (S. aureus, B. cereus, S. pyogenes, Streptococcus sp.), and one gram negative bacterium (E. coli), was tested by Saadoun et al. For antimicrobial activity determination the hole-plate diffusion method, the dilution method, and the Bauer–Kirby method were applied. For evaluation of the minimum inhibitory concentration (MIC) the dilution method was used. Tobromycin (10 µg), nalidixic acid (30 µg), amoxicillin (30 µg), and cefotaxime (30 µg) were tested as standard antibiotics. O. cernua extract showed inhibitory activity against all tested bacteria with MIC of 1527, 3125, 25,000 and 50,000 µg/mL for S. aureus, Streptococcus sp., S. pyogenes and both for B. cereus and E. coli, respectively. In comparison with standard antibiotics an MIC of 3125 µg/mL is equal to cefotaxime (30 µg) and tobromycin (10 µg) activity against Streptococcus sp. and S. aureus, respectively; 25,000 and 50,000 µg/mL is equal to cefotaxime (30 µg) activity against S. pyogenes and B. cereus, respectively, and 50,000 µg/mL is equal to nalidixic acid (30 µg) activity against E. coli [98]. Antibacterial and antifungal activities in vitro of caffeic acid (34) and its derivatives including verbascoside (10) and orobanchoside (29) isolated from O. rapum-genistae, and poliumoside (11) (naturally occurring in O. crenata [14,36], P. aegyptiaca [56], and P. ramosa [14]) against two plant-pathogenic fungi (Sclerotinia sclerotiorum (Lib.) de Bary, Botrytis cinerea Pers. ex Nocca & Balb.) and seven plant-pathogenic bacteria (gram positive: Corynebacterium rathayi (Smith) Dowson, Corynebacterium fascians (Tilford) Dowson, Corynebacterium sepedonicum (Spieckermann & Kotthoff) Skaptason & Burkholder; gram negative: A. tumefaciens, Erwinia carotovora var. carotovora (Jones) Dye, Xanthomonas pelargonii (Brown) Starr & Burkholder, Pseudomonas syringae van Hall) were tested. The other tested substances were caffeic acid (34), ferulic acid (36), esculine, esculetin, rosmarinic acid, gallic acid methylester, chlorogenic acid (37), plantamajoside, and neomyricoside. Additionally Forsythia intermedia var. spectablis Koehne extract was tested. Ferulic acid (36) and chlorogenic acid (37) had been identified in O. cernua by other authors [17]. E. coli and S. aureus were used as references. For fungi MIC was evaluated in solid media using five different concentrations (0.12, 0.25, 0.50, 1.00, 2.00 mg/mL) of solutions of the tested substances, and for bacteria an agar diffusion method with four different concentrations of solutions of the tested substances (12.5, 25, 50, 100 mg/mL) together with MIC measurement in liquid media with six different concentrations of the tested substances (0.1, 0.5, 1.0, 1.5, 2.0, 2.5 mg/mL) were used. Orobanchoside (29) and caffeic acid (34) showed pronounced antifungal activities with MIC values of 2.00 mg/mL and 0.25 mg/mL, respectively, against S. sclerotiorum in media with pH 5. Orobanchoside (29) furthermore had an MIC value of 0.25 mg/mL against S. sclerotiorum and 1.00 mg/mL against B. cinera in media with pH 7, while caffeic acid (34) showed an MIC of 0.25 mg/mL against B. cinera in pH 7 medium. Verbascoside (10) and poliumoside (11) were both able to reduce growth of S. sclerotiorum and B. cinera in pH 5 and pH 7 media, but complete inhibition was not observed. Ferulic acid (36) showed MIC values of 0.13 mg/mL against S. scleretorium and B. cinerea in pH 5 and pH 7 media. Chlorogenic acid (37) was able to reduce growth of S. sclerotiorum and B. cinerea in pH 5 and pH 7 media, but again, complete inhibition was not observed. Against the tested bacteria MIC values were as follows: MIC of caffeic acid (34) against C. rathayi (1.0 mg/mL), C. fascians (1.0 mg/mL), C. sepedonicum (0.1 mg/mL), A. tumefaciens (1.0 mg/mL), E.carotovora var. carotovora (1.0 mg/mL), X. pelargonii (1.0 mg/mL), P. syringae (not determined), S. aureus (1.5 mg/mL), E. coli (1.5 mg/mL); MIC of ferulic acid (36) against C. rathayi (0.5 mg/mL), C. fascians (0.5 mg/mL), C. sepedonicum (0.5 mg/mL), A. tumefaciens (1.0 mg/mL), E.carotovora var. carotovora (1.0 mg/mL), X. pelargonii (0.5 mg/mL), P. syringae (1.0 mg/mL), S. aureus (1.0 mg/mL), E. coli (1.5 mg/mL); MIC of chlorogenic acid (37) against C. rathayi (1.5 mg/mL), C. fascians (1.0 mg/mL), C. sepedonicum (1.0 mg/mL), A. tumefaciens (2.0 mg/mL), E.carotovora var. carotovora (2.5 mg/mL), X. pelargonii (1.5 mg/mL), P. syringae (2.0 mg/mL), S. aureus (>2.5 mg/mL), E. coli (2.5 mg/mL); MIC values of verbascoside (10), poliumoside (11), and orobanchoside (29) were not investigated. Caffeic acid (34) and its derivatives are potential natural plant protective agents against some plant-pathogenic fungi and bacteria as demonstrated in this work. Streptomycin, tested along with the caffeic acid derivatives, was a much more potent bacterial growth inhibitor than the other tested compounds with MIC values of <0.1 mg/mL (C. rathayi, C. fascians, C. sepedonicum, A. tumefaciens, E.carotovora var. carotovora, S. aureus, E. coli), with an exception for X. pelargonii (>2.5 mg/mL) (P. syringae MIC not determined) [99].
2.5.3. Antioxidant Activities as Food Preservative
O. crenata ethanolic extract total antioxidant activity was tested using the phosphomolybdenum method with ascorbic acid as standard. The antioxidant activity was expressed as ascorbic acid equivalents (AE) (mg/g of extract). The two investigated individual Orobanche plants showed good total antioxidant activity 619 ± 9 mg AE/g extract and 561 ± 9 mg AE/g extract [35].
2.5.4. Antioxidative Effects, Anti-Inflammatory Activity in Human Leucocytes, Effects on Production of Reactive Oxygen Species (ROS)
Phenylpropanoid glycosides isolated from O. coerulescens were tested for their antioxidative effects on human low-density lipoprotein. For evaluation of their antioxidant activity dialyzed LDL obtained from human blood samples was diluted with PBS to 100 µg/mL, pre-incubated with the test compounds at 37 °C for 30 min, and then incubated with CuSO4 at 37 °C to induce lipid peroxidation. Resveratrol, a natural phenolic antioxidant e.g., from red wine, was used as a positive control. Conjugated diene formation was monitored and prolonged lag phase (min) used as an index of antioxidant activity when an antioxidant was present in LDL oxidation with Cu2+. All seven isolated phytochemical compounds, phenylpropanoid glycosides desrhamnosyl acteoside (9), acteoside (10), caerulescenoside (13), campneoside II (17), isoacteoside (23), oraposide (29), and 3′-methyl crenatoside (30) suppressed conjugated diene formation with IC50 values of 0.64 ± 0.03, 0.31 ± 0.01, 1.25 ± 0.06, 1.15 ± 0.04, 1.01 ± 0.05, 1.69 ± 0.15, and 2.97 ± 0.31 µM, respectively while resveratrol had an IC50 value of 6.75 ± 1.05 µM. This showed that all isolated compounds from O. coerulescens were more effective antioxidants than the positive control, resveratrol [23]. Phenylpropanoid glycosides acteoside (10), rossicaside B (28), and oraposide (29), isolated from O. coerulescens, were tested for their inflammation-modulating activity in human leucocytes. Peripheral human neutrophils (PMNs) and mononuclear cells were exposed to phorbol-12-myristate-13-acetate (PMA), a direct proetin kinase C (PKC) activator, and N-formyl-methionyl-leucocyl-phenylalanine (fMLP), a receptor mediated and G protein coupled activator, for the induction of production of reactive oxygen species (ROS) and upregulation of β2 integrin in an in-vitro model. For the prevention of PMA-induced ROS production, acteoside (10), rossicaside B (28), and oraposide (29) showed IC50 values of 12.8 ± 7.2 µM, 5.6 ± 2.8 µM, and 6.8 ± 2.3 µM respectively in PMNs, and IC50 values of 9.6 ± 3.2 µM, 23.9 ± 2.9 µM, and 10.0 ± 4.3 µM respectively in mononuclear cells. IC50 values for prevention of fMLP-induced ROS production were 3.5 ± 0.6 µM, 3.0 ± 0.1 µM, and 3.0 ± 0.2 µM for acteoside (10), rossicaside B (28), and oraposide (29) in PMNs, respectively, and 8.8 ± 3.2 µM, 3.5 ± 0.2 µM, and 3.5 ± 0.2 µM in mononuclear cells, respectively. Furthermore, the inhibition of NADPH oxidase (NOX) activity in cell lysate by phenylpropanoid glycosides was tested since NOX is the major ROS producing enzyme in activated leucocytes. Acteoside (10), rossicaside B (28), and oraposide (29) were more potent in NOX inhibition than the positive control, diphenyleneiodonium (DPI, a NOX inhibitor). Moreover, these compounds showed effective free radical-scavenging activity in a cell-free DPPH (2,2-diphenyl-1-picrylhydrazyl) assaying system. Acteoside (10) and oraposide (29) also significantly inhibited PMA- and fMLP-induced β2 integrin expression in human peripheral leucocytes. These effects make O. coerulescens and other drugs containing acteoside (10), rossicaside B (28), and oraposide (29) interesting as potential anti-inflammatory agents for the treatment of oxidative-stress-related diseases [24]. Antioxidative potential was also investigated by Kidachi et al. for phenylpropanoids acteoside (10), cistanoside D (20), isoacteoside (23), oraposide (29), 3‴-O-methyl crenatoside (30), and isocrenatoside (32) from a methanolic O. minor extract, two synthetic derivatives acteoside-tetramethylether, oraposide-tetramethylether, as well as caffeic acid (34) and hydroxytyrosol using the DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging activity assay. Strong activities were observed for acteoside (10) (IC50 15.2 µM), isoacteoside (23) (IC50 20.0 µM), oraposide (29) (IC50 24.5 µM), and isocrenatoside (32) (IC50 29.0 µM), whereas moderate activities were observed for 3‴-O-methyl crenatoside (30) (IC50 54.2 µM), caffeic acid (34) (IC50 38.7 µM), and hydroxytyrosol (IC50 44.6 µM), and no antioxidant activities were observed for acteoside-tetramethylether (IC50 > 100 μM), oraposide-tetramethylether (IC50 > 100 μM), and cistanoside D (20) (IC50 > 100 μM). Epigallocatechin gallate used as positive control showed an IC50 value of 13.5 µM. No standard deviations of the measured values were indicated [46]. Gao et al. found O. cernua extract and acteoside (10) to exhibit strong scavenging effects with IC50 values of 56.3 µg/mL and 20.6 µg/mL, respectively. No standard deviations of the measured values were given [100]. Antioxidant activities of O. crenata and O. foetida methanolic and aqueous extracts was tested by Abbes et al. using DPPH and ABTS (2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid) radical scavenging activity assays. Synthetic antioxidants BHT (2,6-di-tert-butyl-4-methylphenol) and AA (ascorbic acid) were used as positive controls. At 1.00 µg/mL DPPH radical scavenging activities of O. crenata methanolic extract, O.crenata water extract, O. foetida methanolic extract, O. foetida water extract, BHT, and AA were 19.5, 18.3, 5.86, 14.7, 13.4, and 54.0%, respectively. At 200 µg/mL O. crenata methanolic extract, O.crenata water extract, O. foetida methanolic extract, O. foetida water extract, BHT, and AA showed DPPH radical scavenging activities of 88.1, 77.0, 92.0 86.1, 85.0, and 86.3%, respectively. The highest activity against DPPH radicals was observed for O. foetida methanolic extract with an IC50 value of 7.19 ± 1.75 µg/mL (BHT: IC50 65.5 ± 1.4 µg/mL; AA IC50 0.93 ± 0.07 µg/mL). Antioxidant activities in the ABTS test, expressed in % inhibition at 0.5 µg/mL, were 4.04 (O. crenata methanolic extract), 1.64 (O. crenata aqueous extract), 1.28 (O. foetida methanolic extract), and 2.34 (O. foetida aqueous extract) (BHT: 9.98%; AA: 23.1%). Activities of nearly 100% for O. crenata methanolic extract, O.crenata water extract, O. foetida methanolic extract, O. foetida water extract, BHT, and AA, respectively, were observed at concentrations of 200 µg/mL [37].
2.5.5. Blood Pressure and Blood Platelet Aggregation
Intravenous injection of the glycosidic fraction of O. crenata 70% ethanolic extract into rats in doses up to 20 mg/100 g led to a temporary lowering of the arterial blood pressure of the treated animals. Higher doses caused slight, persistent lowering of the arterial blood pressure [29]. Hypotensive activity of O. aegyptiaca 30% aqueous extract and of the alkaloid containing chloroform fraction (further fractionation of the extract with different solvents gave hexane, ether, chloroform, alcohol, and water fractions) after i.v. injection into dogs was also evaluated. The alkaloidal fraction showed strong hypotensive effects. (Hypertension was artificially induced using the Goldblatt technique.) 10 mg i.v. lowered the blood pressure by about 48 mmHg for three hours [57]. A mixture of verbascoside (10) and orobanchoside (29) extracted from O. hederae was tested for its effect on ADP-induced (10–15 µM) blood platelet aggregation and blood pressure in New Zealand male rabbits and Wistar male rats. A dose-dependent inhibition of ADP-induced platelet aggregation of 12.9 ± 4.0%, 43.7 ± 7.8%, 49.4 ± 6.4%, 59.4 ± 6.9%, and 73.7 ± 8.3% at concentrations of 0.2 mg/mL, 0.4 mg/mL, 0.6 mg/mL, 0.8 mg/mL, and 1.0 mg/mL phenylpropanoid glycosides respectively was observed using an aggregometer. Blood pressure was not affected by phenylpropanoid glycosides injected i.v. into the test animals [42].
2.5.6. Contractions of Toad and Rabbit Hearts and Rat Intestines
O. aegyptiaca 30% aqueous extract was further fractionated with different solvents to give hexane, ether, chloroform, alcohol, and water fractions. The extract and fractions were tested for different biological activities. Effects on toad (Bufo regularis Reuss) and rabbit hearts were investigated. Doses of 1, 2, 3, and 4 mL of the aqueous extract were added to 50 mL bath (Ringer´s solution for toad hearts, Lock´s solution for rabbit hearts) and the amplitude or heart rate (toad hearts) as well as the volume of Lock´s solution perfused by the heart (rabbit hearts) were recorded. Contractions of toad hearts and rabbits’ hearts perfused by the extract were stimulated. Also contractions of isolated rats intestines were stimulated whereas uterine contractions in rats were inhibited [57].
2.5.7. Diuretic Effects
Oral application of the phenylpropanoid containing fraction of O. crenata extract in rats had strong diuretic effects. O. crenata extract doses of 100 mg/100 g body weight and 200 mg/100 g body weight were orally applied. Rats were put in diuresis cages and the volume of the collected urine was measured after 1, 3, 6, and 24 h. The untreated control group produced 0, 2.65 ± 0.22, 6.25 ± 0.05, and 11.2 ± 0.1 mL urine after 1, 3, 6, and 24 h, respectively. After application of O. crenata extract doses of 100 mg/100 g b.w. 0, 3.27 ± 0.05, 6.95 ± 0.05, and 12.9 ± 0.1 mL urine were collected after 1, 3, 6, and 24 h, respectively. O. crenata extract doses of 200 mg/100 g b.w. led to 0.37 ± 0.05, 3.95 ± 0.05, 8.25 ± 0.12, and 14.3 ± 0.2 mL of urine after 1, 3, 6, and 24 h, respectively, showing increasing diuresis with higher O. cernua extract doses [29]. O. aegyptiaca 30% aqueous extract was further fractionated with different solvents to give hexane, ether, chloroform, alcohol, and water fractions. The extract and fractions were tested for different biological activities. Diuretic effects of the 20% alcoholic extract were observed in rabbits. Urine volumes of treated animals were measured after 0.5, 1, 2, 3, and 24 h and compared to urine volumes of the animals after 24 h without any treatment. The average urine volume of treated animals (average dose of 9.5 mL of 20% extract/kg b.w.) after 24 h was 107 ± 51 in comparison with 73 ± 21.2 mL for the untreated animals [57].
2.5.8. Inhibition of Amyloid β-Aggregation
Acteoside (10) and oraposide (29) isolated from O. minor were tested for their inhibitory effects on aggregation of human 42-mer amyloid β-protein (Aβ-42), which is believed to play an important role in the pathogenesis of Alzheimer´s disease. Thioflavin-T (Th-T) fluorescence assays, transmission electron microscopy (TEM), and circular dichroism (CD) spectroscopy were used to investigate the inhibitory effects. Acteoside (10) and oraposide (29) showed potent inhibitory effects on the aggregation of Aβ-42 with IC50 values of 8.9 µM and 3.6 µM respectively. IC50 values were calculated from the inhibition rate (%) of each compound towards Aβ-42 aggregation after 24 h by using the Th-T assay. Furthermore, an anti-aggregating effect was suggested by the significant reduction of Aβ fibril formation by 50 µM acteoside (10). β-Sheet formation in Aβ-42 was also inhibited [45]. Kidachi et al. also tested inhibition of amyloid β-42 (Aβ-42) aggregation by phenylpropanoids acteoside (10) and oraposide (29) from O. minor methanolic extract, their synthetic derivatives acteoside-tetramethylether, oraposide-tetramethylether, as well as cistanoside D (20), isoacteoside (23), 3‴-O-methyl crenatoside (30), isocrenatoside (32), caffeic acid (34), and hydroxytyrosol. The IC50 values were calculated from the inhibitory rate (%) of each compound towards Aβ-42 aggregation after 48 h by using the thioflavin-T (Th-T) fluorescence assay. Aβ-42 aggregation was inhibited by acteoside (10) and oraposide (29) with IC50 values of 11.3 µM and 8.2 µM, respectively. Moderate inhibitory activity was observed for 3‴-O-methyl crenatoside (30) (IC50 28.0 µM), very weak inhibitory activity was observed for caffeic acid (34) (IC50 93.8 μM) and hydroxytyrosol (IC50 92.0 μM), and no inhibitory activity was observed for acteoside-tetramethylether (IC50 > 100 μM), oraposide-tetramethylether (IC50 > 100 μM), and cistanoside D (20) (IC50 > 100 μM). 3,4-Di-O-caffeoylquinic acid used as positive control for Aβ-42 aggregation showed an IC50 value of 30.2 µM. No standard deviations of the measured values were indicated [46]. The observed anti-amyloidal effects make acteoside (10) a potential agent for treating or preventing Alzheimer’s disease [45].
2.5.9. Memory Enhancing Effects
Acteoside (10) showed memory enhancing effects and increased significantly the expression of nerve growth factor (NGF) and tropomycin receptor kinase A (TrkA) mRNA and protein in the hippocampus in mice [13]. NGF and TrkA are closely associated with cognitive function and a decrease thereof is related to Alzheimer´s disease. Acteoside (10) treatment resulted in an improvement of learning and memory deficits via promotion of NGF and TrkA expression in the brain. The authors used a senescent mouse model induced by a combination of chronic intraperitoneal administration of D-gal (60 mg/kg/day) and oral administration of AlCl3 (5 mg/kg/day) once daily for 90 days. After 60 days mice in three different groups were treated intragastrically with acteoside (10) (30, 60, and 120 mg/kg/day) for 30 days. Learning ability and memory of the mice were tested using the Morris water maze test. Afterwards mice brains were removed and the hippocampus CA1 region studied immunohistochemically. Reverse transcription polymerase chain reactions (RT-PCR) and western blot analyses were performed to investigate the expression of NGF mRNA and TrkA mRNA [13].
2.5.10. Muscle Relaxant and Antispasmodic Effects
Dose dependent smooth muscle relaxant effects (phenylpropanoid containing fraction of O. crenata extract) were observed when testing different doses on the peristaltic movements of isolated perfused rabbit’s intestine. Doses of 50, 80, 100, 150, and 200 mg/50 mL bath were tested. Movement inhibitions of 31.9 ± 6.8, 38.4 ± 10.1, 44.2 ± 4.3, 51.2 ± 10.3, and 95.4 ± 2.7% were obsereved, respectively [29]. Potent antispasmodic effects on isolated perfused guinea-pig ileum were observed (phenylpropanoid fraction of O. crenata extract). Contractions were induced by acetylcholine application and afterwards different doses of O. crenata extract were tested for their antispasmodic potential. Extract doses tested were 200, 400, 600, and 800 mg/50 mL bath. Inhibition of spasmodic contractions ranged from 22.8 ± 2.1, 55.6 ± 3.7, and 67.3 ± 10.1 to 94.3 ± 6.4%, respectively, showing a dose dependent antispasmodic potential [29].
2.5.11. Nutrient Source
O. crenata was found to be a good source of nutrients. It contained a low moisture level (<8%), a high amount of protein (7.30%), ash contents of 9.20–10.1%, a crude fiber content ranging from 22.1 to 23.5%, and a nutritive value of 244–247 kcal/100 g plant dry weight [35].
2.5.12. Photoprotective Effects
O. cernua ethanolic extract and its principal component, acteoside (10), were studied for their photoprotective effects on UVB-induced photoaging as well as for the underlying molecular mechanisms in normal human dermal fibroblasts (NHDFs). UV radiation causes excessive reactive oxygen species (ROS) generation, which triggers matrix metalloproteinase (MMPs) production, collagen degradation, and premature aging (photoaging). Cell viability of UVB-irradiated NHDFs and the effects of O. cernua extract and acteoside (10) on cell viability were tested using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. O. cernua extract (100 µg/mL) and acteoside (10) (10 µM) were able to recover cell viability by 24.7% and 26.9%, respectively. For effects of O. cernua extract and acteoside (10) on intracellular ROS generation cells were first exposed to UVB irradiation, which increased the ROS level by 282% (ROS levels of normal group were set to 100%). The following treatment with O. cernua extract (100 µg/mL) and acteoside (10) (10 µM) reduced ROS levels by 73.0% and 42.3%, respectively. Furthermore, O. cernua extract and acteoside (10) significantly reduced MMP-1 and IL-6 (Interleukin) levels in NHDFs exposed to UVB-irradiation. O. cernua extract (100 µg/mL) and acteoside (10) (10 µM)-treated groups suppressed UVB-induced MMP-1 levels by 49.0% and 57.1%, respectively. Additionally, the secretion of IL-6 was lowered by 79.4% and 57.1%, by O. cernua extract (100 µg/mL) and acteoside (10) (10 µM). O. cernua extract and acteoside (10) could reverse a UVB induced decrease in type-I procollagen mRNA, with an increased rate of 52.7% and 25.7%, respectively. Furthermore, the UVB-induced increased production of MMP-1 and MMP-3 mRNA levels were strongly inhibited by O. cernua extract (100 µg/mL) and acteoside (10) (10 µM). O. cernua extract decreased the expression of MMP-1 and MMP-3 mRNA by 42.5% and 28.3%, respectively, while acteoside (10) decreased the expression by 44.4% and 66.7%, respectively. Carried with the AP-1 binding sites, the promoters of MMPs were transactivated by AP-1 transcription factor. The expression of phosphorylated c-fos and c-jun, the major components of AP-1 was measured and inhibitory effects on UVB-induced p-c-fos and p-c-jun expression in a dose-dependent manner by acteoside (10) treatments were observed. O. cernua extract (100 µg/mL) reduced the levels of p-c-fos and p-c-jun by 56.0% and 75.6%, respectively, and acteoside (10) (10 µM) reduced the levels by 93.0% and 65.3%, respectively. The mitigen-activated protein kinase (MAPK) signaling pathway, as the upstream of AP-1 transcription factor, has been reported to be activated by UVB-elevated ROS. Biological effects of O. cernua extract and acteoside (10) on the MAPKs family were further studied in UVB-irradiated NHDFs. UVB radiation elevated the phosphorylated forms of MAPKs molecules including ERK, JNK and p38. O. cernua extract and acteoside (10) suppressed the phosphorylation of ERK, JNK and p38 caused by UVB. Levels of p-ERK, p-JNK and p-38 were decreased by O. cernua (100 µg/mL) extract by 46.4%, 58.8% and 84.8%, respectively, and decreased by acteoside (10) (10 µM) treatment by 47.9%, 75.5%, and 77.4%, respectively. The effects of O. cernua extract and acteoside (10) on Nrf2 nuclear translocation and antioxidant enzyme expression were investigated in Western blots in UVB-irradiated NHDFs. Data showed that the nuclear levels already raised by UVB-irradiation were further elevated by O. cernua extract and acteoside (10). The expression of Nrf2 was increased by 56.0% and 69.0% by O. cernua extract (100 µg/mL) and acteoside (10) (10 µM), respectively. Moreover, HO-1 and NQO-1 levels were increased by O. cernua extract (100 µg/mL) by 76.4% and 120%, and by acteoside (10) (10 µM) by 103% and 110%, respectively. Furthermore, O. cernua extract and acteoside (10) were able to reverse the downregulation of TGF-β1 and p-Smad2/3 expression in UVB-irradiated NHDFs. TGF-β1and p-Smad2/3 protein expressions were recovered by 71.6% and 70.7% by O. cernua extract (100 µg/mL), and by 53.7% and 182%, respectively, by acteoside (10) (10 µM), compared with the UVB radiation group. Also O. cernua extract (100 µg/mL) and acteoside (10) (10 µM) inhibited the UVB-induced Smad7 expression by 48.9% and 57.1%, respectively, in comparison with the UVC group. The antiphotoaging effects of O. cernua extract and acteoside (10) were investigated and it was detected that O. cernua extract and acteoside (10) inhibited UVB-irradiated MMP-1 and MMP-3 mRNA upregulation and IL-6 secretion. Moreover, O. cernua extract and acteoside (10) reduced UVB-induced MMP-1 protein secretion, and enhanced type-I procollagen synthesis in NHDFs. O. cernua extract and acteoside (10) treatment furthermore led to the inhibition of the UVB-activated MAPK/AP-1 pathway by inhibiting the UVB-induced phosphorylation of ERK, JNK, and p38 and the expression of p-c-fos and p-c-jun. Levels of cytoprotective agents HO-1 and NQO-1 were increased by O. cernua extract and acteoside, hereby increasing protection against UVB-induced oxidative stress through activation of the cutaneous endogenous antioxidant system. UVB-induced enhacement of Smad7 expression and decrease of Smad2/phosphorylation were reversed by O. cernua extract and acteoside (10), and TGF-β1 expression was enhanced, hereby repairing the TGF-β/Smad signaling pathway and enhancing type-I procollagen synthesis [100]. (No standard deviations of the measured values described above were indicated.)
2.5.13. Summary of Bioactivities
In conclusion, Orobanche extracts and isolated Orobanche natural products were positively tested for a variety of biological activities including anti-hypertensive, anti-platelet aggregating, and memory enhancing effects. UV protecting and anti-photoaging effects open an interesting field of study and antioxidant activities on human LDL, inflammation modulating effects in human leucocytes, ROS production and amyloid β-aggregation inhibiting effects make the species containing the responsible substances potential agents for treatment of Alzheimer’s disease and oxidation related diseases. Furthermore, Orobanche extracts are active against a wide variety of pathogenic fungi and bacteria and can be potential alternatives to synthetic antibiotics and plant protecting agents. The by far best investigated compound is the phenylpropanoid glycoside acteoside (10) which is responsible for a large part of the observed effects, such as antioxidant, anti-inflammatory, radical scavenging, amyloid β-aggregation inhibiting, memory enhancing, antimicrobial, and photoprotective effects and also oraposide (29) was shown to have several interesting effects. However, the occurrence of acteoside (10) is not restricted to Orobanche or Orobanchaceae but the compound is widely distributed in the plant kingdom. It is found in over 200 species belonging to 23 plant families, most of them belonging to the order Lamiales [101]. Thus, even though Orobanche extracts and substances extracted thereof show the above stated biological activities, there might be better and easier accessible sources for the bioactive compounds than the holoparasitic taxa of the genus Orobanche.
3. Discussion
Most of the natural products found in the genus Orobanche (n = 70) have so far been reported only from one source (n = 51), and only three compounds from more than four taxa: acteoside 10 (from 13 source taxa), oraposide 29 (from 12 source taxa), and orobanone 45 (also from 12 source taxa). While most of the literature on Orobanche is about strigolactones, seed germination stimulants, parasitic weed management, and host-parasite interaction (SciFinder, last accessed first of October, 2018), publications on secondary metabolism of Orobanche species are relatively rare. Of the more than 200 species belonging to Orobanche s.l. only 27 species have been investigated for secondary metabolites. Compound classes detected in the analyzed species comprise aromatic aldehydes, ketones and phenylmethanoids (Figure 1), phenylethanoids (Figure 2), phenylethanoid glycosides (Figure 3), phenylpropanoid glycosides (Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8), phenolic acids (Figure 9), lignans (Figure 10 and Figure 11), flavonoids (Figure 12 and Figure 13), a tropone derivative (Figure 14), and sterols (Figure 15, Figure 16, Figure 17, Figure 18, Figure 19 and Figure 20). Investigations on biological activities of Orobanche extracts and isolated pure secondary metabolites from Orobanche species show a wide variety of effects, e.g., antibacterial and antifungal activities [35,46,98], inhibition of amyloid-β-aggregation [45,46] or photoprotection against UVB-irradiation [100]. Orobanche are not only destructive weeds, but might also be a source of active agents against several diseases, in particular against fungal and bacterial, and inflammatory diseases, correlated with ROS production. Nevertheless, it has to be considered, that phenylpropanoids in general and e.g., acteoside, one of the best investigated compounds of Orobanche in particular, are not restricted to Orobanche species but are widely distributed in the plant kingdom, possibly making other species more interesting sources of these compounds [101,102]. An aspect that deserves more research and could be a challenging subject for future studies is the idea that natural products sequestered by Orobanche species from their host species could be further metabolized by the parasites. Metabolization of host plant natural products could result in new, formerly undescribed hybrid compounds not synthesized by a single species. To study this phenomenon, more analytical studies of the secondary metabolism of Orobanche species and their host plants are warranted.
Supplementary Materials
The following are available online. Table S1: Overview Natural Products synthesized by Orobanche species. Table S2: Natural Products sequestered by Orobanche species from host species. Text S1: Literature search strategy & key words.
Author Contributions
Conceptualization, C.Z.; Investigation, F.S.; Writing-Original Draft Preparation, F.S.; Writing-Review & Editing, C.Z.; Supervision, C.Z.
Funding
This research received no external funding.
Acknowledgments
The authors wish to thank Jürgen Pusch (Sondershausen) for fruitful discussions and for help in obtaining relevant literature.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kojić, M.; Maširević, S.; Jovanović, D. Distribution and biodiversity of broomrape (Orobanche L.) worldwide and in Serbia. Helia 2001, 24, 73–92. [Google Scholar]
- Pusch, J.; Günther, K.-F. Orobanchaceae. In Gustav Hegi—Illustrierte Flora von Mitteleuropa, Band VI Teil 1A, Dez. 2009//Orobanchaceae (Sommerwurzgewächse), Scrophulariaceae (Rachenblütler), 3rd ed.; Wagenitz, G., Ed.; Weissdorn-Verlag: Jena, Germany, 2009. [Google Scholar]
- Park, J.-M.; Manen, J.-F.; Colwell, A.E.; Schneeweiss, G.M. A plastid gene phylogeny of the non-photosynthetic parasitic Orobanche (Orobanchaceae) and related genera. J. Plant Res. 2008, 121, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Schneeweiss, G.M. Phylogenetic relationships and evolutionary trends in Orobanchaceae. In Parasitic Orobanchaceae—Parasitic Mechanisms and Control Strategies; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 243–265. [Google Scholar]
- Schneider, A.C. Resurrection of the genus Aphyllon for New World broomrapes (Orobanche s.l., Orobanchaceae). PhytoKeys 2016, 75, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Westwood, J.; Uludag, A. Proceedings IPPS International Parasitic Plant Society 10th World Congress of Parasitic Plants. 2009. Available online: http://parasiticplants.org/docs/IPPS_10th_Congress_Abstracts_Kusadasi_Turkey.pdf (accessed on 29 October 2018).
- Joel, D.M. The new nomenclature of Orobanche and Phelipanche. Weed Res. 2009, 49, 6–7. [Google Scholar] [CrossRef]
- Bennett, J.R.; Mathews, S. Phylogeny of the parasitic plant family Orobanchaceae inferred from phytochrome A. Am. J. Bot. 2006, 93, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneeweiss, G.M.; Colwell, A.; Park, J.-M.; Jang, C.-G.; Stuessy, T.F. Phylogeny of holoparasitic Orobanche (Orobanchaceae) inferred from nuclear ITS sequences. Mol. Phylogenet. Evol. 2004, 30, 465–478. [Google Scholar] [CrossRef]
- The Plant List, a Working List of All Plant Species. Scientific Plant Names of Vascular Plants and Bryophytes. Collaboration between the Royal Botanic Gardens, Kew and Missouri Botanical Garden. Available online: http://www.theplantlist.org (accessed on 23 July 2018).
- Roudbaraki, S.J.; Nori-Shargh, D. The volatile constituent analysis of Orobanche alba Stephan from Iran. Curr. Anal. Chem. 2016, 12, 496–499. [Google Scholar] [CrossRef]
- Fruchier, A.; Rascol, J.-P.; Andary, C.; Privat, G. A tropone derivative from Orobanche rapum-genistae. Phytochemistry 1981, 20, 777–779. [Google Scholar] [CrossRef]
- Gao, L.; Peng, X.-M.; Huo, S.-X.; Liu, X.-M.; Yan, M. Memory enhancement of acteoside (verbascoside) in a senescent mice model induced by a combination of D-gal and AlCl3. Phytother. Res. 2015, 29, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Di Fabio, A.; Foddai, S.; Ballero, M.; Poli, F. The occurrence of phenylpropanoid glycosides in Italian Orobanche spp. Biochem. Syst. Ecol. 1995, 23, 855–858. [Google Scholar] [CrossRef]
- Aynehchi, Y.; Salehi Sormaghi, M.H.; Amin, G.H.; Soltani, A.; Qumehr, N. Survey of Iranian plants for saponins, alkaloids, flavonoids and tannins. II. Int. J. Crude Drug Res. 1982, 20, 61–70. [Google Scholar] [CrossRef]
- Mølgaard, P.; Ravn, H. Evolutionary aspects of caffeoyl ester distribution in dicotyledons. Phytochemistry 1988, 27, 2411–2421. [Google Scholar] [CrossRef]
- Qu, Z.-Y.; Zhang, Y.-W.; Yao, C.-L.; Jin, Y.-P.; Zheng, P.-H.; Sun, C.-H.; Liu, J.-X.; Wang, Y.-S.; Wang, Y.-P. Chemical constituents from Orobanche cernua Loefling. Biochem. Syst. Ecol. 2015, 60, 199–203. [Google Scholar] [CrossRef]
- Qu, Z.-Y.; Zhang, Y.-W.; Zheng, S.-W.; Yao, C.-L.; Jin, Y.-P.; Zheng, P.-H.; Sun, C.-H.; Wang, Y.-P. A new phenylethanoid glycoside from Orobanche cernua Loefling. Nat. Prod. Res. 2016, 30, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-Z.; Wang, X.-Q.; Li, C. Chemical constituents from Orobanche cernua. Zhongcaoyao 2014, 45, 2447–2452. [Google Scholar]
- Zhao, J.; Liu, T.; Ma, L.; Yan, M.; Zhao, Y.; Gu, Z.; Huang, Y. Protective effect of acteoside on immunological liver injury induced by Bacillus Calmette-Guerin plus lipopolysaccharide. Planta Med. 2009, 75, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yan, M.; Huang, Y.; Liu, T.; Zhao, Y. Study on water soluble constituents of Orobanche coerulescens. Tianran Chanwu Yanjiu Yu Kaifa 2009, 21, 619–621. [Google Scholar]
- Murayama, T.; Yanagisawa, Y.; Kasahara, A.; Onodera, K.-I.; Kurimoto, M.; Ikeda, M. A novel phenylethanoid, isocrenatoside isolated from the whole plant of Orobanche coerulescens. J. Nat. Med. 1998, 52, 455–458. [Google Scholar]
- Lin, L.-C.; Chiou, W.-F.; Chou, C.-J. Phenylpropanoid glycosides from Orobanche caerulescens. Planta Med. 2004, 70, 50–53. [Google Scholar] [PubMed]
- Lin, L.-C.; Wang, Y.-H.; Hou, Y.-C.; Chang, S.; Liou, K.-T.; Chou, Y.-C.; Wang, W.-Y.; Shen, Y.-C. The inhibitory effect of phenylpropanoid glycosides and iridoid glucosides on free radical production and beta2 integrin expression in human leucocytes. J. Pharm. Pharmacol. 2006, 58, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-J.; Yang, Q.; Wang, F.; Zhu, P. A new phenethyl alcohol glycoside from Orobanche coerulescens. Zhongcaoyao 2016, 47, 1269–1271. [Google Scholar]
- Zhao, J.; Yan, M.; Huang, Y.; He, W.-Y.; Zhao, Y. Study on chemical constituents of Orobanche coerulescens. Zhongyaocai 2007, 30, 1255–1257. [Google Scholar] [PubMed]
- Shao, H.-X.; Yang, J.-Y.; Ju, A.-H. Studies on chemical constituents of Mongolian medicine Orobanche coerulescens. Zhonghua Zhongyiyao Zazhi 2011, 26, 129–131. [Google Scholar]
- Zhang, Q.-R. A new phenethyl alcohol glycoside from Orobanche coerulescens. Zhongguo Zhong Yao Za Zhi 2017, 42, 1136–1139. [Google Scholar] [PubMed]
- El-Shabrawy, O.A.; Melek, F.R.; Ibrahim, M.; Radwan, A.S. Pharmacological evaluation of the glycosidated phenylpropanoids containing fraction from Orobanche crenata. Arch. Pharm. Res. 1989, 12, 22–25. [Google Scholar] [CrossRef]
- Afifi, M.S.; Lahloub, M.F.; El-Khayaat, S.A.; Anklin, C.G.; Rüegger, H.; Sticher, O. Crenatoside: A novel phenylpropanoid glycoside from Orobanche crenata. Planta Med. 1993, 59, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Gatto, M.A.; Sanzani, S.M.; Tardia, P.; Linsalata, V.; Pieralice, M.; Sergio, L.; Di Venere, D. Antifungal activity of total and fractionated phenolic extracts from two wild edible herbs. Nat. Sci. 2013, 5, 895–902. [Google Scholar] [CrossRef]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; Di Venere, D. Activity of extracts from wild edible herbs against postharvest fungal diseases of fruit and vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]
- Gatto, M.A.; Sergio, L.; Ippolito, A.; Di Venere, D. Phenolic extracts from wild edible plants to control postharvest diseases of sweet cherry fruit. Postharvest Biol. Technol. 2016, 120, 180–187. [Google Scholar] [CrossRef]
- Nishibe, S.; Tamayama, Y.; Sasahara, M.; Andary, C. A phenylethanoid glycoside from Plantago asiatica. Phytochemistry 1995, 38, 741–743. [Google Scholar] [CrossRef]
- Nada, S.A.; El-Chaghaby, G.A. Nutritional evaluation, phytoconstituents analysis and biological activity of the parasitic plant Orobanche crenata. J. Chem. Biol. Sci. Sect. A 2015, 5, 171–180. [Google Scholar]
- Dini, I.; Iodice, C.; Ramundo, E. Phenolic metabolites from Orobanche speciosa. Planta Med. 1995, 61, 389–390. [Google Scholar] [CrossRef] [PubMed]
- Abbes, Z.; El Abed, N.; Amri, M.; Kharrat, M.; Ben Hadj Ahmed, S. Antioxidant and antibacterial activities of the parasitic plants Orobanche foetida and Orobanche crenata on faba bean in Tunisia. J. Anim. Plant Sci. 2014, 24, 310–314. [Google Scholar]
- Andary, C.; Wylde, R.; Maury, L.; Heitz, A.; Dubourg, A.; Nishibe, S. X-ray analysis and extended NMR study of oraposide. Phytochemistry 1994, 37, 855–857. [Google Scholar] [CrossRef]
- Andary, C.; Wylde, R.; Laffite, C.; Privat, G.; Winternitz, F. Structures of verbascoside and orobanchoside, caffeic acid sugar esters from Orobanche rapum-genistae. Phytochemistry 1982, 21, 1123–1127. [Google Scholar] [CrossRef]
- Aynilian, G.H.; Abou-Char, C.I.; Edgecombe, W. Screening of herbarium specimens of native plants from the families Amaranthaceae, Dipsaceae and Orobanchaceae for alkaloids, saponins and tannins. Planta Med. 1971, 19, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Pieretti, S.; Di Giannuario, A.; Capasso, A.; Nicoletti, M. Pharmacological effects of phenylpropanoid glycosides from Orobanche hederae. Phytother. Res. 1992, 6, 89–93. [Google Scholar] [CrossRef]
- Capasso, A.; Pieretti, S.; Di Giannuario, A.; Nicoletti, M. Pharmacological study of phenylpropanoid glycosides: Platelet aggregation and blood pressure studies in rabbits and rats. Phytother. Res. 1993, 7, 81–83. [Google Scholar] [CrossRef]
- Baccarini, A.; Melandri, B.A. Studies on Orobanche hederae physiology: Pigments and CO2 fixation. Physiol. Plant. 1967, 20, 245–250. [Google Scholar] [CrossRef]
- Rohmer, M.; Ourisson, G.; Benveniste, P.; Bimpson, T. Sterol biosynthesis in heterotrophic plant parasites. Phytochemistry 1975, 14, 727–730. [Google Scholar] [CrossRef]
- Kurisu, M.; Miyamae, Y.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Inhibition of amyloid β aggregation by acteoside, a phenylethanoid glycoside. Biosci. Biotechnol. Biochem. 2013, 77, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Kidachi, E.; Kurisu, M.; Miyamae, Y.; Hanaki, M.; Murakami, K.; Irie, K.; Shigemori, H. Structure-activity relationship of phenylethanoid glycosides on the inhibition of amyloid β aggregation. Heterocycles 2016, 92, 1976–1982. [Google Scholar]
- Dzhumyrko, S.F.; Sergeeva, N.V. Carotenoid pigments from Orobanche owerinii. Chem. Nat. Compd. 1985, 21, 712–713. [Google Scholar] [CrossRef]
- Han, J.-X.; Yang, J.-Y.; Shao, H.-X.; Ju, A.-H. A study on the chemical constituents of Orobanche pycnostachya Hance. Neimenggu Daxue Xuebao Ziran Kexueban 2010, 41, 669–672. [Google Scholar]
- Li, C.-F.; Wen, A.-P.; Wang, X.-Q.; Han, G.-Q. HPLC simultaneous determination of three phenylethanoid glycosides in Orobanche pycnostachya. Yaowu Fenxi Zazhi 2016, 36, 291–295. [Google Scholar]
- Andary, C.; Privat, G.; Chevallet, P.; Orzalesi, H.; Serrano, J.J.; Boucard, M. Chemical and pharmacodynamic study of heteroside esters of caffeic acid, isolated from Orobanche rapum-genistae. Farm. Sci. 1980, 35, 3–30. [Google Scholar]
- Andary, C.; Rascol, J.P.; Privat, G. Two new ecotypes of Orobanche rapum-genistae Thuill. Trav. Soc. Pharm. Montp. 1980, 40, 293–295. [Google Scholar]
- Bridel, M.; Charaux, C. L’orobanchine, glucoside nouveau, retiré des tubercules de l’Orobanche rapum Thuill. Bull. Soc. Chim. Fr. 1924, 34, 1153–1600. [Google Scholar]
- Bridel, M.; Charaux, C. Sur le processus du noircissement des Orobanches au cours de leur dessiccation. C. R. Hebd. Seanc. Acad. Sci. 1925, 180, 387–388. [Google Scholar]
- Viron, C.; Lhermite, S.; André, P.; Lafosse, M. Isolation of phenylpropanoid glycosides from Orobanche rapum by high speed countercurrent chromatography. Phytochem. Anal. 1998, 9, 39–43. [Google Scholar] [CrossRef]
- Viron, C.; Pennanec, R.; André, P.; Lafosse, M. Large scale centrifugal partition chromatography in purification of polyphenols from Orobanche rapum. J. Liq. Chrom. 2000, 23, 1681–1688. [Google Scholar] [CrossRef]
- Afifi, M.S.A.; Lahloub, M.F.; Zaghloul, A.M.; El-Khayaat, S.A. Phenylpropanoid glycosides from Orobanche aegyptiaca and Orobanche ramosa. J. Pharm. Sci. 1993, 9, 225–233. [Google Scholar]
- Sharaf, A.; Youssef, M. Pharmacologic investigation on Orobanche egyptiaca with a special study on its hypotensive action. Qual. Plant. Mater. Veg. 1971, 20, 255–269. [Google Scholar] [CrossRef]
- Lahloub, M.F.; Zaghloul, A.M.; El-Khayaat, S.A.; Afifi, M.S.; Sticher, O. 2′-O-Acetylpoliumoside: A new phenylpropanoid glycoside from Orobanche ramosa. Planta Med. 1991, 57, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. Identification of the flavone pigment of Phelipaea ramosa. Chem. Ind. 1958, 48, 1590–1591. [Google Scholar]
- Melek, F.R.; Aboutabl, E.A.; Elsehrawy, H. Tricin from Orobanche ramosa L. Egypt. J. Pharm. Sci. 1992, 33, 753–756. [Google Scholar]
- Andary, C.; Privat, G.; Wylde, R.; Heitz, A. Pheliposide et arenarioside, deux nouveaux esters hétérosidiques de l’acide caféique isolés de Orobanche arenaria. J. Nat. Prod. 1985, 48, 778–783. [Google Scholar] [CrossRef]
- Smith, J.D.; Mescher, M.C.; de Moraes, C.M. Implications of bioactive solute transfer from hosts to parasitic plants. Curr. Opin. Plant Biol. 2013, 16, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Lotti, G.; Paradossi, C. Host-parasite mineral composition in plants infected with Orobanchaceae. Agric. Ital. 1977, 77, 153–165. [Google Scholar]
- Lotti, G.; Paradossi, C. Absorption of petroselinic acid by Orobanche hederae on Hedera helix. Agrochimica 1987, 31, 484–488. [Google Scholar]
- Sareedenchai, V.; Zidorn, C. Sequestration of polyacetylenes by the parasite Orobanche hederae (Orobanchaceae) from its host Hedera helix (Araliaceae). Biochem. Syst. Ecol. 2008, 36, 772–776. [Google Scholar] [CrossRef]
- Wink, M.; Witte, L.; Hartmann, T. Quinolizidine alkaloid composition of plants and of photomixotrophic cell suspension cultures of Sarothamnus scoparius and Orobanche rapum-genistae. Planta Med. 1981, 43, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Rascol, J.P.; Andary, C.; Roussel, J.L.; Privat, G. Alkaloids from Orobanche rapum-genistae. I. Isolation and identification of three major alkaloids, (−)-sparteine, (+)-lupanine, and (+)-13-hydroxylupanine. Plantes Med. Phytother. 1978, 12, 287–295. [Google Scholar]
- Rascol, J.P.; Andary, C.; Privat, G. Alkaloids of Orobanche rapum-genistae. II. Variation of the amount of sparteine and lupanine in the host-parasite relations. Trav. Soc. Pharm. Montp. 1980, 40, 261–268. [Google Scholar]
- Fournier, G.; Paris, M. Mise en évidence de cannabinoïds chez Phelipaea ramosa, Orobanchacées, parasitant le chanvre, Cannabis sativa, Cannabinacées. Planta Med. 1983, 49, 250–251. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D. Toxicity in animals. Trends in evolution? Toxicon 2001, 39, 87–96. [Google Scholar] [CrossRef]
- Bornancin, L.; Bonnard, I.; Mills, S.C.; Banaigs, B. Chemical mediation as a structuring element in marine gastropod predator-prey interactions. Nat. Prod. Rep. 2017, 34, 644–676. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Robert, C.A.M. Sequestration of plant secondary metabolites by insect herbivores: Molecular mechanisms and ecological consequences. Curr. Opin. Insect Sci. 2016, 14, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Opitz, S.E.W.; Müller, C. Plant chemistry and insect sequestration. Chemoecology 2009, 19, 117–154. [Google Scholar] [CrossRef]
- Harborne, J.B. Twenty-five years of chemical ecology. Nat. Prod. Rep. 2001, 18, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.S.; Rank, C.; Jensen, S.R. Transfer of iridoid glucosides from host plant Galium verum to hemiparasitic Euphrasia stricta. Biochem. Syst. Ecol. 2006, 34, 763–765. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Foderaro, T.A.; Li, Y.-X. Iridoid glycoside uptake by Castilleja integra via root parasitism on Penstemon teucrioides. Phytochemistry 1993, 32, 1151–1153. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Harris, G.H. Transfer of pyrrolizidine and quinolizidine alkaloids to Castilleja (Scrophulariaceae) hemiparasites from composite and legume host plants. J. Chem. Ecol. 1987, 13, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Arslanian, R.L.; Harris, G.H.; Stermitz, F.R. New quinolizidine alkaloids from Lupinus argenteus and its hosted root parasite Castilleja sulphurea. Stereochemistry and conformation of some naturally occurring cyclic carbinolamides. J. Org. Chem. 1990, 55, 1204–1210. [Google Scholar] [CrossRef]
- Adler, L.S.; Wink, M. Transfer of quinolizidine alkaloids from hosts to hemiparasites in two Castilleja-Lupinus associations: Analysis of floral and vegetative tissues. Biochem. Syst. Ecol. 2001, 29, 551–561. [Google Scholar] [CrossRef]
- Boros, C.A.; Marshall, D.R.; Caterino, C.R.; Stermitz, F.R. Iridoid and phenylpropanoid glycosides from Orthocarpus spp. Alkaloid content as a consequence of parasitism on Lupinus. J. Nat. Prod. 1991, 54, 506–513. [Google Scholar] [CrossRef]
- Wink, M.; Witte, L. Quinolizidine alkaloids in Genista acanthoclada and its holoparasite, Cuscuta palaestina. J. Chem. Ecol. 1993, 19, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Bäumel, P.; Witte, L.; Czygan, F.-C.; Proksch, P. Transfer of qinolizidine alkaloids from various host plants of the Fabaceae to parasitizing Cuscuta species. Biochem. Syst. Ecol. 1994, 22, 647–656. [Google Scholar] [CrossRef]
- Schneider, M.J.; Stermitz, F.R. Uptake of host plant alkaloids by root parasitic Pedicularis species. Phytochemistry 1990, 29, 1811–1814. [Google Scholar] [CrossRef]
- Cabezas, N.J.; Urzúa, A.M.; Niemeyer, H.M. Translocation of isoquinoline alkaloids to the hemiparasite, Tristerix verticillatus from its host, Berberis montana. Biochem. Syst. Ecol. 2009, 37, 225–227. [Google Scholar] [CrossRef]
- Marko, M.D.; Stermitz, F.R. Transfer of alkaloids from Delphinium to Castilleja via root parasitism. Norditerpenoid alkaloid analysis by electrospray mass spectrometry. Biochem. Syst. Ecol. 1997, 25, 279–285. [Google Scholar] [CrossRef]
- Martín-Cordero, C.; Ayuso, M.A.; Richomme, P.; Bruneton, J. Quinolizidine alkaloids from Viscum cruciatum, hemiparasitic shrub of Lygos sphaerocarpa. Planta Med. 1989, 55, 196. [Google Scholar] [CrossRef]
- Cordero, C.M.; Serrano, A.G.; Gonzalez, M.A. Transfer of bipiperidyl and quinolizidine alkaloids to Viscum cruciatum Sieber (Loranthaceae) hemiparasitic on Retama sphaerocarpa Boissier (Leguminosae). J. Chem. Ecol. 1993, 19, 2389–2393. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cordero, C.; Pedraza, M.A.; Gil, A.M.; Ayuso, M.J. Bipiperidyl and quinolizidine alkaloids in fruits of Viscum cruciatum hemiparasitic on Retama sphaerocarpa. J. Chem. Ecol. 1997, 23, 1913–1916. [Google Scholar] [CrossRef]
- Lehtonen, P.; Helander, M.; Wink, M.; Sporer, F.; Saikkonen, K. Transfer of endophyte-origin defensive alkaloids from a grass to a hemiparasitic plant. Ecol. Lett. 2005, 8, 1256–1263. [Google Scholar] [CrossRef]
- Smith, J.D.; Woldemariam, M.G.; Mescher, M.C.; Jander, G.; de Moraes, C.M. Glucosinolates from host plantsinfluence growth of the parasitic plant Cuscuta gronovii and its susceptibility to aphid feeding. Plant Physiol. 2016, 172, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Boonsong, C.; Wright, S.E. The cardiac glycosides present in mistletoes growing on Nerium oleander. Aust. J. Chem. 1961, 14, 449–457. [Google Scholar] [CrossRef]
- Rothe, K.; Diettrich, B.; Rahfeld, B.; Luckner, M. Uptake of phloem-specific cardenolides by Cuscuta sp. growing on Digitalis lanata and Digitalis purpurea. Phytochemistry 1999, 51, 357–361. [Google Scholar] [CrossRef]
- Nakamura, T.; Okuyama, E.; Tsukada, A.; Yamazaki, M.; Satake, M.; Nishibe, S.; Deyama, T.; Moriya, A.; Maruno, M.; Nishimura, H. Acteoside as the analgesic principle of cedron (Lippia triphylla), a Peruvian medicinal plant. Chem. Pharm. Bull. 1997, 45, 499–504. [Google Scholar] [CrossRef]
- Isacchi, B.; Iacopi, R.; Bergonzi, M.C.; Ghelardini, C.; Galeotti, N.; Norcini, M.; Vivoli, E.; Vincieri, F.F.; Bilia, A.R. Antihyperalgesic activity of verbascoside in two models of neuropathic pain. J. Pharm. Pharmacol. 2011, 63, 594–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraja, T.G.; Nare, R.B.; Laxmikant, V.; Patil, B. In vitro screening of antimicrobial activity of Orobanche aegyptiaca. J. Biopestic. 2010, 3, 548–549. [Google Scholar]
- Saadoun, I.; Hameed, K.M.; Al-Momani, F.; Ababneh, Q. Effect of three Orobanche spp. extracts on some local phytopathogens, Agrobacterium and Erwinia. Turk. J. Biol. 2008, 32, 113–117. [Google Scholar]
- Saadoun, I.; Hameed, K.M. Antibacterial activity of Orobanche cernua extract. J. Basic Microbiol. 1999, 39, 377–380. [Google Scholar] [CrossRef]
- Ravn, H.; Andary, C.; Kovács, G.; Mølgaard, P. Caffeic acid esters as in vitro inhibitors of plant pathogenic bacteria and fungi. Biochem. Syst. Ecol. 1989, 17, 175–184. [Google Scholar] [CrossRef]
- Gao, W.; Wang, Y.-S.; Qu, Z.-Y.; Hwang, E.; Ngo, H.T.T.; Wang, Y.-P.; Bae, J.; Yi, T.-H. Orobanche cernua Loefling attenuates ultraviolet B-mediated photoaging in human dermal fibroblasts. Photochem. Photobiol. 2018, 94, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Kurkin, V.A. Phenylpropanoids from medicinal plants: Distribution, classification, structural analysis, and biological activity. Chem. Nat. Compd. 2003, 39, 123–153. [Google Scholar] [CrossRef]
Sample Availability: There are no samples available from the authors. |
Figure 1.
Phenylmethanoids, aromatic aldehydes and ketones 1–5.

| No | Name | R1 | R2 | R3 |
|---|---|---|---|---|
| 1 | p-Hydroxybenzaldehyde | H | OH | H |
| 2 | Protocatechuic aldehyde | OH | OH | H |
| 3 | Isovanillin | OH | OCH3 | H |
| 4 | Vanillin | OCH3 | OH | H |
| 5 | Syringaldehyde | OCH3 | OH | OCH3 |
Figure 2.
Phenylethanoid, p-Hydroxy acetophenone 6.

Figure 3.
Non-acylated phenylethanoid glycosides 7 and 8.

| No | Name | R1 | R2 |
|---|---|---|---|
| 7 | 2-Phenylethyl β-primeveroside | H | Xyl |
| 8 | Salidroside 1 | OH | H |
1 Syn. rhodioloside.
Figure 4.
Phenylpropanoid glycosides I, 9–22.

| No | Name | R1 | R2 | R3 | R4 | R5 | R6 |
|---|---|---|---|---|---|---|---|
| 9 | Desrhamnosyl acteoside | H | H | H | H | H | H |
| 10 | Acteoside 1 | H | H | H | H | Rha | H |
| 11 | Poliumoside | H | H | H | H | Rha | Rha |
| 12 | Arenarioside | H | H | H | H | Rha | Xyl |
| 13 | Caerulescenoside | H | H | H | H | (3-O-Glc)-Rha | H |
| 14 | 2′-O-Acetylacteoside | H | H | H | Ac | Rha | H |
| 15 | 2′-O-Acetylpoliumoside 2 | H | H | H | Ac | Rha | Rha |
| 16 | Pheliposide | H | H | H | Ac | Rha | Xyl |
| 17 | Campneoside II | H | H | OH | H | Rha | H |
| 18 | Campneoside I | H | H | OCH3 | H | Rha | H |
| 19 | Leucosceptoside A | CH3 | H | H | H | Rha | H |
| 20 | Cistanoside D | CH3 | CH3 | H | H | Rha | H |
| 21 | Orobancheoside B | CH3 | CH3 | H | H | Rha | feruloyl- |
| 22 | Orobancheoside A | CH3 | CH3 | H | H | (4-O-Ac)-Rha | H |
1 Syn. kusaginin [16], orobanchin, verbascoside; 2 syn. brandioside.
Figure 5.
Phenylpropanoid glycosides II, 23 and 24.

| No | Name | R |
|---|---|---|
| 23 | Isoacteoside 1 | H |
| 24 | Isocampneoside I 2 | OCH3 |
1 Syn. isoverbascoside, 2 stereochemistry not reported.
Figure 6.
Phenylpropanoid glycosides III, 25–28.

Figure 7.
Phenylpropanoid glycosides IV, 29 and 30. Note: According to Andary et al. [38] crenatoside and oraposide both describe the same structure. The authors report another isolated substance, orobanchoside (also mentioned in a former work from Andary et al. [39]). In a later work from Nishibe et al. [34] the structure of orobanchoside is being revised and it was established that orobanchoside is the same compound as crenatoside and oraposide. The name oraposide is used preferentially here, because it seems to be the oldest name for the compound at hand; however, crenatoside seems to have been used more often in the literature.
Figure 7.
Phenylpropanoid glycosides IV, 29 and 30. Note: According to Andary et al. [38] crenatoside and oraposide both describe the same structure. The authors report another isolated substance, orobanchoside (also mentioned in a former work from Andary et al. [39]). In a later work from Nishibe et al. [34] the structure of orobanchoside is being revised and it was established that orobanchoside is the same compound as crenatoside and oraposide. The name oraposide is used preferentially here, because it seems to be the oldest name for the compound at hand; however, crenatoside seems to have been used more often in the literature.

| No | Name | R |
|---|---|---|
| 29 | Oraposide 1 | H |
| 30 | 3‴-O-Methyl crenatoside | CH3 |
1 Syn. crenatoside, orobanchoside.
Figure 8.
Phenylpropanoid glycosides V, 31 and 32, and phenylethanoid glycoside 33.

| No | Name | R |
|---|---|---|
| 31 | Descaffeoyl crenatoside | H |
| 32 | Isocrenatoside | caffeoyl- |
| 33 | 3′-O-Methyl isocrenatoside | ferulouyl- |
Figure 9.
Phenolic acids 34–36.

| No | Name | R1 | R2 |
|---|---|---|---|
| 34 | Caffeic acid | H | H |
| 35 | Methyl caffeate | H | CH3 |
| 36 | trans-Ferulic acid | CH3 | H |
Figure 10.
Chlorogenic acid (5-Caffeoyl quinic acid) 37.

Figure 11.
Dimethyl-6,9,10-trihydroxybenzol[kl]xanthene-1,2-dicarboxylate 38.

Figure 12.
Lignans 39–41.

| No | Name | R1 | R2 |
|---|---|---|---|
| 39 | (+)-Pinoresinol 4′-O-β-D-glucopyranoside | H | H |
| 40 | Isoeucommin A | H | OCH3 |
| 41 | (+)-Syringaresinol 4′-O-β-D-glucopyranoside | OCH3 | OCH3 |
Figure 13.
Flavones 42–44.

| No | Name | R1 | R2 |
|---|---|---|---|
| 42 | Apigenin | H | H |
| 43 | Luteolin | OH | H |
| 44 | Tricin | OCH3 | OCH3 |
Figure 14.
Tropone derivative—Orobanone 45.

Figure 15.
Sterols I: stigmastane derivative, stigmastanol 46.

Figure 16.
Sterols II: cholest-5-en derivatives, 47–54.

| No | Name | R1 | R2 |
|---|---|---|---|
| 47 | Cholesterol | H |  |
| 48 | Campesterol | H |  |
| 49 | 24-Methylene cholesterol | H |  |
| 50 | β-Sitosterol | H |  |
| 51 | Stigmasterol | H |  |
| 52 | Isofucosterol | H |  |
| 53 | Daucosterol | β-D-Glc |  |
| 54 | Stigmasterol-3-O-β-D-glucoside | β-D-Glc |  |
Figure 17.
Sterols III: cholest-7-en derivatives, 55–58.

| No | Name | R1 | R2 |
|---|---|---|---|
| 55 | ∆7-Campestenol | H |  |
| 56 | Episterol | OH |  |
| 57 | ∆7-Stigmastenol | OH |  |
| 58 | ∆7-Avenasterol | OH |  |
Figure 18.
Sterols IV: lophenol derivatives, 59–62.

| No | Name | R |
|---|---|---|
| 59 | 24-Methyl lophenol |  |
| 60 | 24-Methylene lophenol |  |
| 61 | 24-Ethyl lophenol |  |
| 62 | 24-Ethylidene lophenol |  |
Figure 19.
Sterols V, 63–66.

| No | Name | R1 | R2 |
|---|---|---|---|
| 63 | 4α-Methyl ∆8-campestenol | H |  |
| 64 | 4α-Methyl ∆8-stigmastenol | OH |  |
| 65 | 24,28-Dihydro obtusifoliol | OH |  |
| 66 | Obtusifoliol | OH |  |
Figure 20.
Sterols VI, 67–70.

| No | Name | R1 | R2 |
|---|---|---|---|
| 67 | 24,28-Dihydrocycloeucalenol | H |  |
| 68 | Cycloeucalenol | H |  |
| 69 | Cycloartenol | CH3 |  |
| 70 | 24-Methylene cycloartenol | CH3 |  |
Figure 21.
N-Methylangustifoline S1.

Figure 22.
Quinolizidine alkaloids I, S2–S10.

| No | Name | R1 | R2 | R3 | R4 |
|---|---|---|---|---|---|
| S2 | (−)-Sparteine | H, H | H | H | H, H |
| S3 | (−)-17-Oxosparteine | H, H | H | H | =O |
| S4 | (+)-Lupanine | =O | H | H | H, H |
| S5 | 17-Oxolupanine | =O | H | H | =O |
| S6 | (+)-13-Hydroxylupanine | =O | H | OH | H, H |
| S7 | 13-Tigloyloxylupanine 1 | =O | H |  | H, H |
| S8 | 13-Angeloyloxylupanine 1 | =O | H |  | H, H |
| S9 | 13-Benzoyloxylupanine 1 | =O | H |  | H, H |
| S10 | 4-Hydroxylupanine | =O | OH | H | H, H |
1 Stereochemistry not resolved.
Figure 23.
Quinolizidine alkaloids II, S11–S13.

Figure 24.
Piperidine alkaloid—Ammodendrine S14.

Figure 25.
Polyacetylenes S15–S17.

Figure 26.
Cannabinoids S18 and S19.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scharenberg, F.; Zidorn, C. Genuine and Sequestered Natural Products from the Genus Orobanche (Orobanchaceae, Lamiales). Molecules 2018, 23, 2821. https://doi.org/10.3390/molecules23112821
AMA Style
Scharenberg F, Zidorn C. Genuine and Sequestered Natural Products from the Genus Orobanche (Orobanchaceae, Lamiales). Molecules. 2018; 23(11):2821. https://doi.org/10.3390/molecules23112821
Chicago/Turabian StyleScharenberg, Friederike, and Christian Zidorn. 2018. "Genuine and Sequestered Natural Products from the Genus Orobanche (Orobanchaceae, Lamiales)" Molecules 23, no. 11: 2821. https://doi.org/10.3390/molecules23112821