
Tiansi Liquid Modulates Gut Microbiota Composition and Tryptophan–Kynurenine Metabolism in Rats with Hydrocortisone-Induced Depression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Model Validation and Behavioral Changes
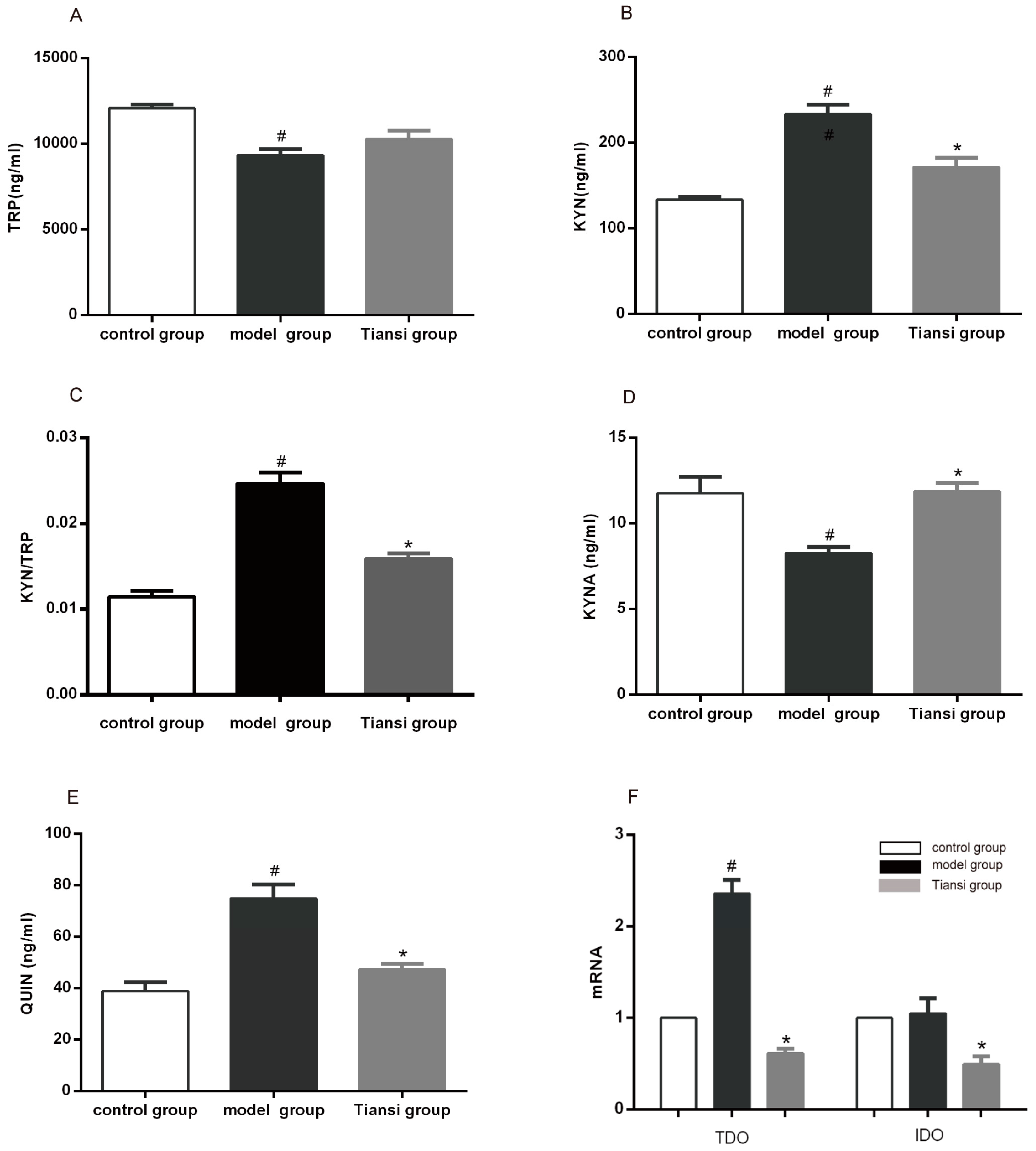
2.2. Regulation of the TRP–KYN Metabolic Pathway
2.2.1. Effects on TRP–KYN Pathway Metabolites
2.2.2. Effects on the Expression of Rate-Limiting Enzymes in the TRP–KYN Pathway
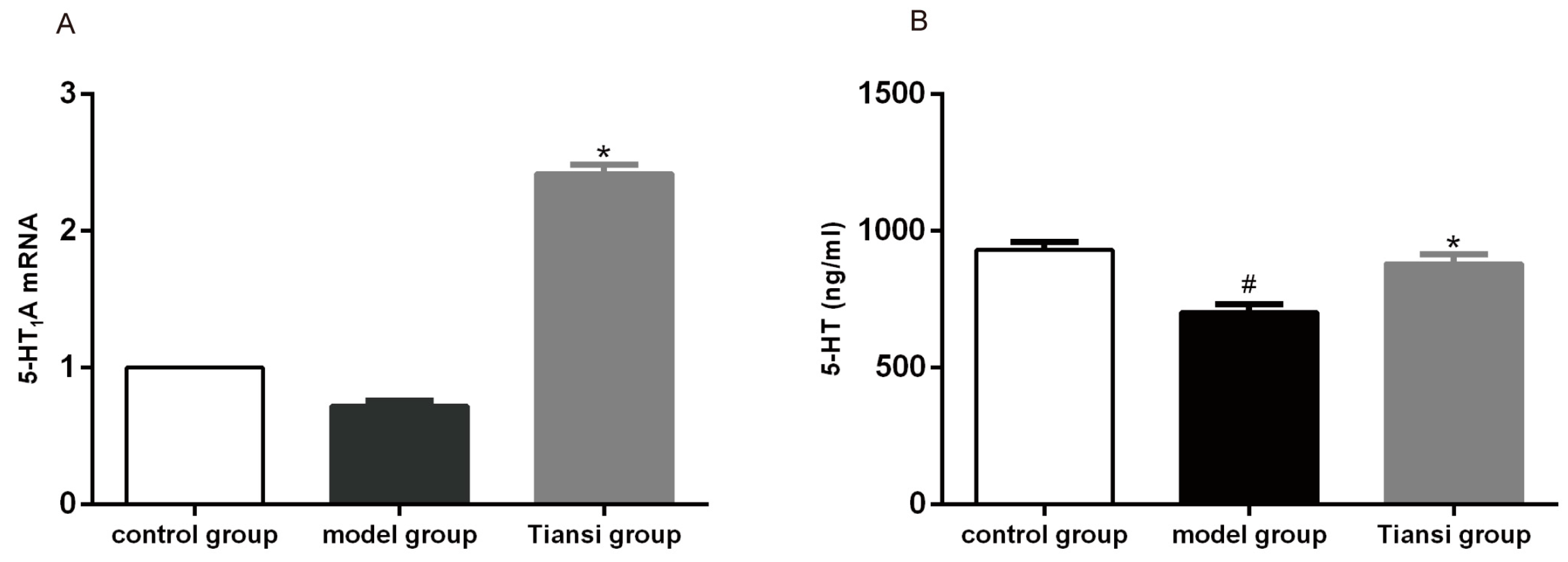
2.3. Effects on 5-HT1A Receptor Expression and 5-HT Content
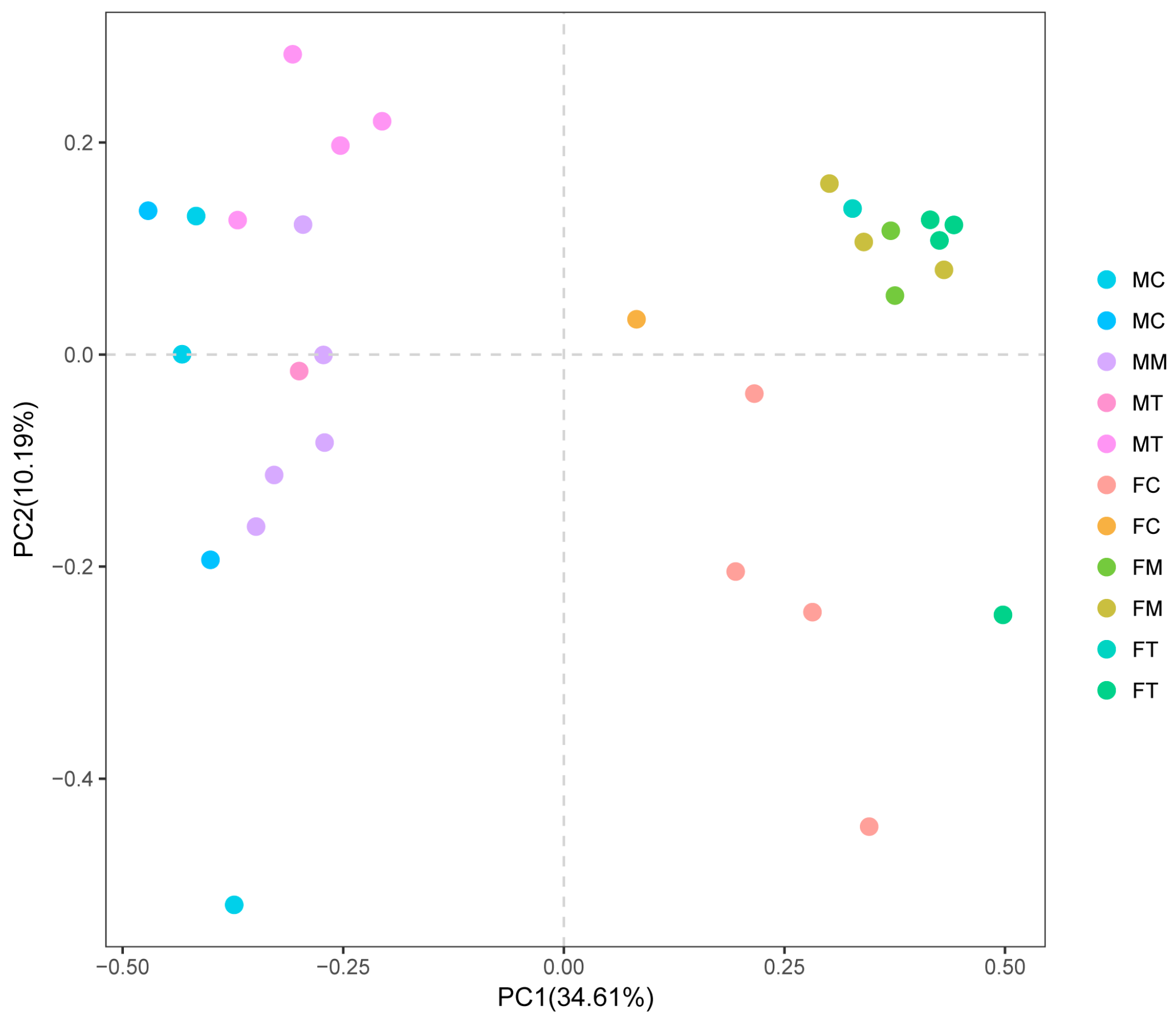
2.4. The Effect of Tiansi Liquid on Microbiota Composition
2.4.1. Tiansi Liquid Alters Microbiota Diversity and Richness in Rats
2.4.2. Altered Microbiota Composition in the Depression Model
Microbiota Composition of Fecal Samples
Microbiota Composition of Intestinal Mucosa Samples
2.5. The Associations among Microbiota and Tryptophan Metabolism
3. Discussion
4. Materials and Methods
4.1. Animals and Treatments
4.2. Generation of the Rat Model of Depression
4.3. Preparation of the Extract
4.4. Sample Preparation
4.5. Behavior Testing
4.5.1. Sucrose Preference Test (SPT)
4.5.2. Open Field Test (OFT)
4.6. Plasma TRP–KYN Pathway Analysis
4.6.1. HPLC-MS/MS Conditions
4.6.2. PCR Analysis
4.7. Sequence Analysis of 16S rRNA
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Moussavi, S.; Chatterji, S.; Verds, E.; Tandon, A.; Patel, V.; Ustun, B. Depression, chronic diseases, and decrements in health: Results from the World Health Surveys. Lancet 2007, 370, 851–858. [Google Scholar] [CrossRef]
- O’Keane, V.; Frodl, T.; Dinan, T.G. A review of atypical depression in relation to the course of depression and changes in HPA axis organization. Psychoneuroendocrinology 2012, 37, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Monje, M.L.; Toda, H.; Palmar, T.D. Inflammatory blockade restores adult hippocampal neurogenesis. Science 2003, 302, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Iosif, R.E.; Ekdahl, C.T.; Ahlenius, H.; Pronk, C.J.; Bonde, S.; Kokaia, Z.; Jacobsen, S.E.; Lindvall, O. Tumor necrosis factor receptor 1 is a negative regulator of progenitor proliferation in adult hippocampal neurogenesis. J. Neurosci. 2006, 26, 9703–9712. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal relationship between stressful life events and the onset of major depression. Am. J. Psychiatry 1999, 156, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.O.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat. Communat. 2015. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Regulation of the stress response by the gut microbiota: Implications for psychoneuroendocrinology. Psychoneuroendocrinology 2012, 37, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Reber, S.O.; Siebler, P.H.; Donner, N.C.; Morton, J.T.; Smith, D.G.; Kopelman, J.M.; Lowe, K.R.; Wheeler, K.J.; Fox, J.H.; Hassell, J.E., Jr.; et al. Immunization with a heat-killed preparation of the environmental bacterium Mycobacterium vaccae promotes stress resilience in mice. Proc. Natl. Acad. Sci. USA 2012, 113, E3130–E3139. [Google Scholar] [CrossRef] [PubMed]
- Bharwani, A.; Mian, M.F.; Foster, J.A.; Surette, M.G.; Bienenstock, J.; Forsythe, P. Structural & functional consequences of chronic psychosocial stress on the microbiome & host. Psychoneuroendocrinology 2016, 63, 217–227. [Google Scholar] [PubMed] [Green Version]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Kuc, D.; Zgrajka, W.; Parada, T.J.; Urbanik, S.T.; Turski, W.A. Micromolar concentration of kynurenic acid in rat small intestine. Amino Acids 2008, 35, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Begley, T.P.; Kinsland, C.; Mehl, R.A.; Osterman, A.; Dorrestein, P. The biosynthesis of nicotinamide adenine dinucleotides in bacteria. Vitam. Horm. 2001, 61, 103–119. [Google Scholar] [PubMed]
- Blokland, A.; Lieben, C.; Deutz, N.E.P. Anxiogenic and depressive like effects, but no cognitive deficits, after repeated moderate tryptophan depletion in the rat. J. Psychopharmacol. 2002, 16, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M.; Kim, Y.K.; Verkerk, R.; Scharpe, S.; Steinbusch, H.; Leonard, B. Kynurenine pathway in major depression: Evidence of impaired neuroprotection. J. Affect. Disord. 2007, 98, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Le Floc’h, N.; Otten, W.; Merlot, E. Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids 2011, 41, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Recinella, L.; Locatelli, M.; Guglielmi, P.; Secci, D.; Leporini, L.; Chiavaroli, A.; Leone, S.; Martinotti, S.; Brunetti, L.; et al. Protective Effects Induced by Microwave-Assisted Aqueous Harpagophytum Extract on Rat Cortex Synaptosomes Challenged with Amyloid β-Peptide. Phytother. Res. 2017, 31, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Bienenstock, J.; Dinan, T.G. The probiotic Bifidobacteria infantis: An assessmentof potential antidepressant properties in the rat. J. Psychiatr. Res. 2008, 43, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Shu, L.; Luo, Z.; Liu, P.; Yang, M.; Tan, C. Preliminary clinical effectiveness of Morinda offcinalis water extract in the treatment of depression. China J. China Mater. Med. 2002, 27, 75–78. [Google Scholar]
- Kong, Q.M.; Shu, L.; Zhang, H.Y.; Jiao, F.Y.; Han, Z.C.; Wang, J.; Du, B.; Shi, J.G.; Wang, X.P.; Ai, C.S.; et al. Efficacy and safety of Morinda officinalis oligose capsule in the treatment of depression. China J. Clin. Pharmacol. 2011, 27, 170–173. [Google Scholar]
- Sharma, B.; Gupta, V.K. Modulations of Mammalian Brain Functions by Antidepressant Drugs: Role of Some Phytochemicals as Prospective Antidepressants. Evid. Based Med. Pract. 2011, 2, 1–12. [Google Scholar]
- Donnapee, S.; Li, J.; Yang, X.; Ge, A.H.; Donkor, P.O.; Gao, X.M.; Chang, Y.X. Cuscuta chinensis Lam: A systematic review on ethnopharmacology, phytochemistry and pharmacology of an important traditional herbal medicine. J. Ethnopharmacol. 2014, 157, 292–308. [Google Scholar] [CrossRef] [PubMed]
- Mokhtarifar, N.; Sharif, B.; Naderi, N.; Mosaddegh, M.; Faizi, M. Evaluation of anti-depressant effects of Cuscuta chinensis in experimental models. Res. Pharm. Sci. 2012, 5, S826. [Google Scholar]
- Zhou, J.Y.; Lu, Y.; Xu, X.Q.; Zhang, J.; Li, H.N.; Chang, H.S. Effect of antidepression and mechanism of regulation on IDO of Tiansi Liquid. J. Beijing Univ. Tradit. Chin. Med. 2015, 38, 182–185. [Google Scholar]
- Moran, C. The small bowel microbiota. Curr. Opin. Gastroenterol. 2015, 30, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.X.; Wu, W.C. Effects of psychological stress on small intestinal motility and bacteria and mucosa in mice. World J. Gastroenerol. 2005, 11, 2016–2021. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; von Wright, A.; Vilpponen-Salmela, T.; Ben-Amor, K.; Akkermans, A.D.L.; de Vos, W.M. Mucosaassociated bacteria in the human gastrointestinal tract are uniformly distributed along the colon and differ from the community recovered from feces. Appl. Environ. Microbiol. 2002, 68, 3401–3407. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Rase, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendler, K.S.; Hettema, J.M.; Butera, F.; Gardner, C.O.; Prescott, C.A. Life event dimensions of loss humiliation entrapment and danger in the prediction of onsets of major depression and generalized anxiety. Arch. Gen. Psychiatry 2003, 60, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.J.; Wu, A.Q. Chronic restraint stress and glucocorticoid stress in an experimental model of depression in rat. Shanghai Arch. Psychiatry 2009, 21, 71–73. [Google Scholar]
- Kelly, J. The Gut Microbiota in Depression. Ph.D. Thesis, University College Cork, Cork, Ireland, October 2016. [Google Scholar]
- Lu, Y.; Chen, J.; Zheng, J.; Hu, G.; Wang, J.; Huang, C.; Lou, L.; Wang, X.; Zeng, Y. Mucosal adherent bacterial dysbiosis in patients with colorectal adenomas. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Naik, S. Compartmentalized and systemic control of tissue immunity by commensals. Nat. Immunol. 2013, 14, 646–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the hosts metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Jia, H.; Zhou, C.; Yang, Y.; Zhao, Y.; Yang, M.; Zou, Z. Variations in gut microbiota and fecal metabolic phenotype associated with depression by 16S rRNA gene sequencing and LC/MS-based metabolomics. J. Pharm. Biomed. Anal. 2017, 14, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Wang, X.; Wang, Z.; Zhang, J.; Jiang, R.; Wang, X.; Liu, Z.; Xia, Z.; Xu, Z. Similar Fecal Microbiota Signatures in Patients with Diarrhea-Predominant Irritable Bowel Syndrome and Patients with Depression. Clin. Gastroenterol. Hepatol. 2016, 14, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Ding, B.; Feng, C.; Yin, S.; Zhang, T.; Qi, X.; Lv, H.; Guo, X.; Dong, K.; Zhu, Y.; et al. Prevotella and Klebsiella proportions in fecal microbial communities are potential characteristic parameters for patients with major depressive disorder. J. Affect. Disord. 2017, 1, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.H.; Aa, J.; Zha, W.; Wang, G.; Wu, X.; Liu, L. Effect of metronidazole on metabolism of hyperlipidaemia hamster studied by metabonomics. J. China Pharm. Univ. 2012, 43, 443–448. [Google Scholar]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Chuang, H.L.; Huang, Y.T.; Wu, C.C.; Chou, G.T.; Wang, S.; Tsai, Y.C. Alteration of behavior and monoamine levels attributable to Lactobacillus plantarum PS128 in germ-free mice. Behav. Brain. Res. 2016, 1, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Nakano, V.; Ignacio, A.; Fernandes, M.R.; Fukugaiti, M.H.; Avila-campos, M.J. Intestinal Bacteroides and Parabacteroides species producing antagonistic substances. Microbiology 2006, 1, 61–64. [Google Scholar]
- Biddle, A.S.; Black, S.J.; Blanchard, J.L. An in vitro model of the horse gut microbiome enables identification of lactate-utilizing bacteria that differentially respond to starch induction. PLoS ONE 2013, 8, e77599. [Google Scholar] [CrossRef] [PubMed]
- Sabina, F. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar]
- Molina, M.A.; Díaz, A.M.; Hesse, C.; Ginter, W.; Gentilini, M.V.; Nuñez, G.G.; Canellada, A.M.; Sparwasser, T.; Berod, L.; Castro, M.S.; et al. Immunostimulatory effects triggered by Enterococcus faecalis CECT7121 probiotic strain involve activation of dendritic cells and interferon-gamma production. PLoS ONE 2015, 10, e0127262. [Google Scholar] [CrossRef] [PubMed]
- Bangsgaard Bendtsen, K.M.; Krych, L.; Sørensen, D.B.; Pang, W.; Nielsen, D.S.; Josefsen, K.; Hansen, L.H.; Sørensen, S.J.; Hansen, A.K. Gut microbiota composition is correlated to grid floor induced stress and behavior in the BALB/c mouse. PLoS ONE 2012, 7, e46231. [Google Scholar] [CrossRef] [PubMed]
- Ruhe, H.G.; Mason, N.S.; Schene, A.H. Mood is indirectly related to serotonin, norepinephrine and dopamine levels in humans: A meta-analysis of monoamine depletion studies. Mol. Psychiatry 2007, 12, 331–359. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.H.; Jia, H.M.; Zhang, H.W.; Feng, Y.F.; An, L.; Zou, Z.M. Hippocampus and serum metabolomic studies to explore the regulation of Chaihu-Shu-Gan-San on metabolic network disturbances of rats exposed to chronic variable stress. Mol. Biosyst. 2014, 10, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Comings, D.E.; Muhleman, D.; Dietz, G.; Sherman, M.; Forest, G.L. Sequence of human tryptophan 2,3-dioxygenase (TDO2): Presence of a glucocorticoid response like element composed of a GTT repeat and an intronic CCCCT repeat. Genomics 1995, 29, 390–396. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, K.; Harkin, A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology 2015, 112, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R.T. Adrenal cortical activity changes in manic-depressive illness: Influence on intermediary metabolism of tryptophan. Arch. Gen. Psychiatry 1967, 17, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Young, S.N. Mechanism of decline in rat brain 5-hydroxytryptamine after induction of liver tryptophan pyrrolase by hydrocortisone: Roles of tryptophan catabolism and kynurenine synthesis. Br. J. Pharmacol. 1981, 74, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Fernstrom, J.D. Tryptophan availability and serotonin synthesis in rat brain: Effects of experimental diabetes. Adv. Biochem. Psychopharmacol. 1982, 34, 85–106. [Google Scholar] [PubMed]
- Schruers, K.; Griez, E. The effects of tryptophan depletion on mood and psychiatric symptoms. J. Affect. Disord. 2003, 64, 107–119. [Google Scholar] [CrossRef]
- Morgese, M.G.; Schiavone, S.; Trabace, L. Emerging role of amyloid beta in stress response: Implication for depression and diabetes. Eur. J. Pharmacol. 2017, 15, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Morgese, M.G.; Schiavone, S.; Mhillaj, E.; Bove, M.; Tucci, P.; Trabace, L. N-3 PUFA diet enrichment prevents amyloid beta-induced depressive-like phenotype. Pharmacol. Res. 2018, 129, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, S.; Tucci, P.; Mhillaj, E.; Bove, M.; Trabace, L.; Morgese, M.G. Antidepressant drugs for beta amyloid-induced depression: A new standpoint? Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 78, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Y.; Li, B.; Song, L.; Chen, M.; Lu, Y. Impacts of Tiansi Liquid on Alzheimer’s Disease Inducedby Aβ25–35 in the Model Rats. World J. Int. Tradit. West. Med. 2013, 11, 1103–1106. [Google Scholar]
- Dylan, D.; Matthew, H.S.; William, V.T.; Bryan, D.M.; Andrew, J.H.; Steven, K.H.; Anthony, L.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 2017, 551, 648–652. [Google Scholar]
- Snell, E.E. Tryptophanase: Structure, catalytic activities, and mechanism of action. Adv. Enzymol. Relat. Areas Mol. Biol. 1975, 42, 287–333. [Google Scholar] [PubMed]
- Takako, S.I.; Yasuo, Y.; Kyosuke, S.; Fuminobu, Y.; Hirohisa, K. Molecular basis of indole production catalyzed by tryptophanase in the genus Prevotella. FEMS Microbiol. Lett. 2011, 322, 51–59. [Google Scholar] [Green Version]
- Williams, B.B.; Van Benschoten, A.H.; Cimermancic, P.; Donia, M.S.; Zimmermann, M.; Taketani, M.; Ishihara, A.; Kashyap, P.C.; Fraser, J.S.; Fischbach, M.A. Discovery and Characterization of Gut Microbiota Decarboxylases that Can Produce the Neurotransmitter Tryptamine. Cell Host Microbe 2014, 16, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chu, S.F.; Li, J.; Chen, N.H. FK506 binding protein 51 is involved in the depression-like behaviors induced by glucocorticoids. Chin. Pharmacol. Bull. 2014, 30, 407–412. [Google Scholar]
- Gregus, A.; Wintink, A.J.; Davis, A.C.; Kalynchuk, L.E. Effect of repeated corticosterone injections and restraint stress on anxiety and depression-like behavior in male rats. Behav. Brain Res. 2005, 156, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Gursoy, E.; Cardounel, A.; Kalimi, M. Biological effects of single and repeated swimming stress in male rats: Beneficial effects of glucocorticoids. Endocrine 2000, 13, 123–129. [Google Scholar] [CrossRef]
- Xu, L.Y.; Li, Z.M.; Yang, L.; Lv, Y.L.; Wang, D.; Li, X.R. Quantitative determination of polysaccharides in Tusizi(Semen Cuscutae). J. Beijing Univ. Tradit. Chin. Med. 2011, 34, 548–551. [Google Scholar]
- Liu, J.J. Effect of Polysaccharides from Morinda Offcin alis on the Oxidative Stress and Congitive-behaviors in an Experimental Depression Model of Rats. China Mod. Dr. 2011, 49, 1–2. [Google Scholar]
- Remus, J.L.; Stewart, L.T.; Camp, R.M.; Novak, C.M.; Johnson, J.D. Interaction of metabolic stress with chronic mild stress in altering brain cytokines and cucrose preference. Behav. Neurosci. 2015, 129, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Lavi-Avnon, Y.; Yadid, G.; Overstreet, D.H.; Weller, A. Abnormal patterns of maternal behavior in a genetic animal model of depression. Physiol. Behav. 2005, 84, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Khadrawy, Y.A.; Salem, A.M.; El-Shamy, K.A.; Ahmed, E.K.; Fadl, N.N.; Hosny, E.N. Neuroprotective and therapeutic effect of caffeine on the rat model of Parkinson’s Disease induced by rotenone. J. Diet. Suppl. 2017, 14, 553–572. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds Morinda officinalis How polysaccharides and Cuscuta chinensis polysaccharides are available from the authors. |








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, D.; Chang, H.; Ma, S.; Guo, J.; She, G.; Zhang, F.; Li, L.; Li, X.; Lu, Y. Tiansi Liquid Modulates Gut Microbiota Composition and Tryptophan–Kynurenine Metabolism in Rats with Hydrocortisone-Induced Depression. Molecules 2018, 23, 2832. https://doi.org/10.3390/molecules23112832
Cheng D, Chang H, Ma S, Guo J, She G, Zhang F, Li L, Li X, Lu Y. Tiansi Liquid Modulates Gut Microbiota Composition and Tryptophan–Kynurenine Metabolism in Rats with Hydrocortisone-Induced Depression. Molecules. 2018; 23(11):2832. https://doi.org/10.3390/molecules23112832
Chicago/Turabian StyleCheng, Dan, Hongsheng Chang, Suya Ma, Jian Guo, Gaimei She, Feilong Zhang, Lingling Li, Xinjie Li, and Yi Lu. 2018. "Tiansi Liquid Modulates Gut Microbiota Composition and Tryptophan–Kynurenine Metabolism in Rats with Hydrocortisone-Induced Depression" Molecules 23, no. 11: 2832. https://doi.org/10.3390/molecules23112832