Antiproliferative Activity of Combined Biochanin A and Ginsenoside Rh2 on MDA-MB-231 and MCF-7 Human Breast Cancer Cells

Abstract
:1. Introduction
2. Results and Discussion
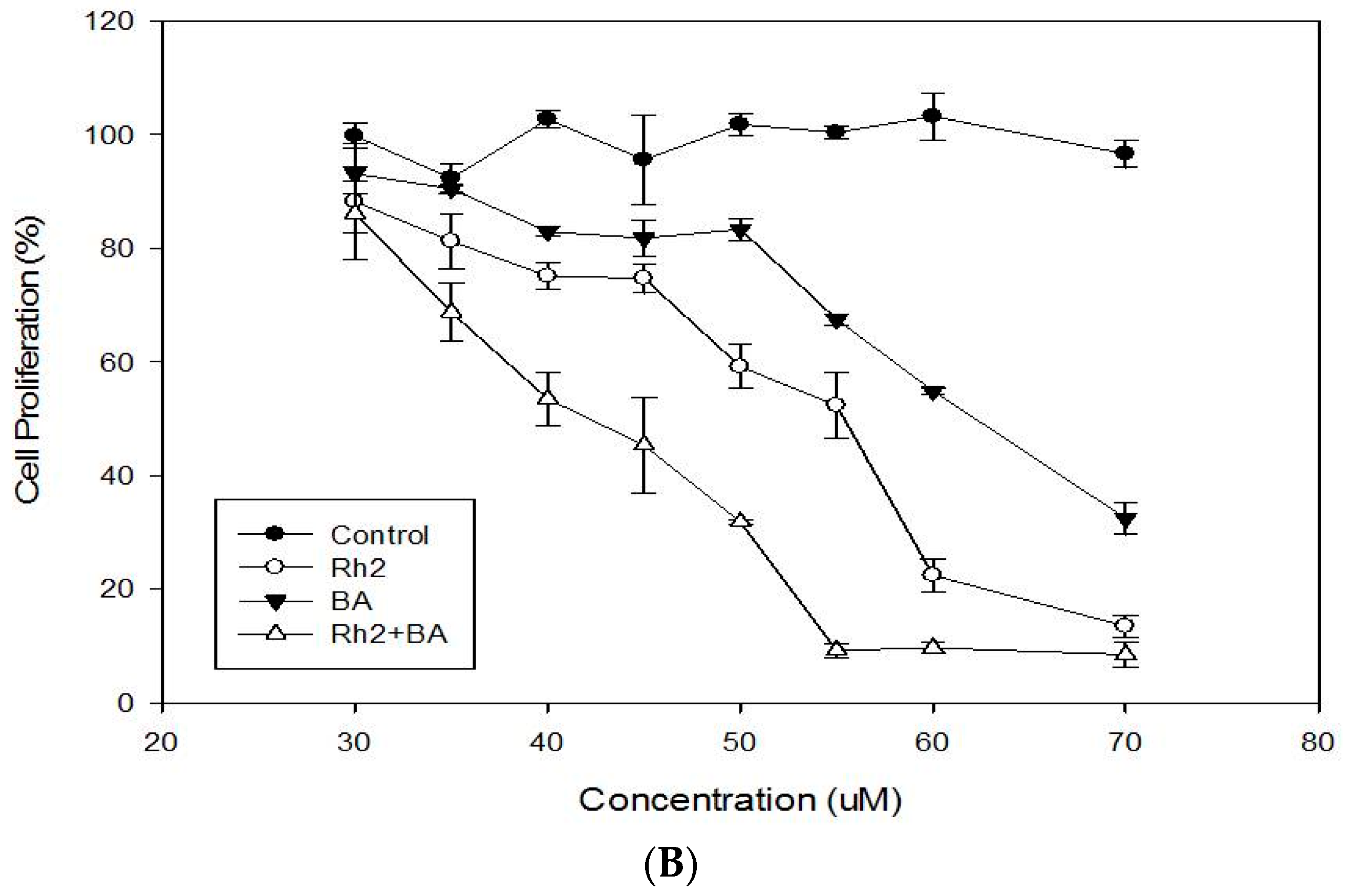
2.1. Cytotoxicity and Antiproliferative Activities of MDA-MB-231 and MCF-7
2.2. Enhanced Inhibition of Cell Migration
2.3. Modulations of Protein Expression and Signalling Pathways
3. Materials and Methods
3.1. Chemicals
3.2. Cytotoxicity Activity
3.3. Antiproliferative Activity
3.4. Combination Study
3.5. Wound-Healing Assay
3.6. Invasion Assay
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dutta, P.; Sarkissyan, M.; Paico, K.; Wu, Y.Y.; Vadgama, J.V. MCP-1 is overexpressed in triple-negative breast cancers and drives cancer invasiveness and metastasis. Breast Cancer Res. Treat. 2018, 3, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Kreike, B.; van Kouwenhove, M.; Horlings, H.; Weigelt, B.; Peterse, H.; Bartelink, H.; van de Vijver, M.J. Gene expression profiling and histopathological characterization of triple-negative/basal-like breast carcinomas. Breast Cancer. Res. 2007, 9, R65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, S.; Shyam, H.; Sharma, R.; Balapure, A.K. Genistein synergizes centchroman action in human breast cancer cells. Indian J. Pharmacol. 2016, 48, 637. [Google Scholar] [PubMed]
- Gan, D.; Zeng, X.; Liu, R.H.; Ye, H. Potential mechanism of mycelium polysaccharide from pholiota dinghuensis bi in regulating the proliferation and apoptosis of human breast cancer mcf-7 cells through p38/mapk pathway. J. Funct. Foods 2015, 12, 375–388. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479–3485. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, R. HSynergistic effect of apple extracts and quercetin 3-beta-d-glucoside combination on antiproliferative activity in MCF-7 human breast cancer cells in vitro. J. Agric. Food Chem. 2009, 57, 8581–8586. [Google Scholar] [CrossRef] [PubMed]
- Cirone, M.; Zompetta, C.; Tarasi, D.; Frati, L.; Faggioni, A. Infection of human T lymphoid cells by human herpesvirus 6 is blocked by two unrelated protein tyrosine kinase inhibitors, biochanin A and herbimycin. AIDS Res. Hum. Retrovir. 1996, 12, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Rufer, C.E.; Kulling, S.E. Antioxidant activity of isoflavones and their major metabolites using different in vitro assays. J. Agric. Food Chem. 2006, 54, 2926–2931. [Google Scholar] [CrossRef] [PubMed]
- Puli, S.; Lai, J.C.; Bhushan, A. Inhibition of matrix degrading enzymes and invasion in human glioblastoma (U87MG) cells by isoflavones. J. Neurooncol. 2006, 79, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Kalayciyan, A. Nicotine and biochanin A, but not cigarette smoke, induce anti-inflammatory effects on keratinocytes and endothelial cells in patients with Behcet’s disease. J. Investig. Dermatol. 2007, 127, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Schrepfer, S. The selective estrogen receptor-beta agonist biochanin A shows vasculo protective effects without uterotrophic activity. Menopause 2006, 13, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ge, B.; Wang, Y.; Ye, Y.; Zeng, S.; Huang, Z.Q. Biochanin A promotes proliferation that involves feedback loop of microRNA-375 and estrogen receptor alpha in breast cancercells. Cell. Phys. Biochem. 2015, 2, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Choi, Y.H.; Choi, S.; Chung, H.; Kim, K.; Kim, S.I.; Kim, N.D. Anti-proliferating effects of ginsenoside Rh2 on MCF-7 human breast cancer cells. Int. J. Oncol. 1999, 14, 869–944. [Google Scholar] [CrossRef] [PubMed]
- Park, H.M.; Kim, S.J.; Kim, J.S.; Kang, H.S. Reactive oxygen species mediated ginsenoside Rg3-and Rh2-induced apoptosis in hepatoma cells through mitochondrial signaling pathways. Food Chem. Toxicol. 2012, 50, 2736–2741. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Wu, G.C.; Hu, R.; Li, M.; Feng, H. Ginsenoside Rh2 inhibits glioma cell proliferation by targeting microRNA-128. Acta Pharmacol. Sin. 2011, 32, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Lee, E.H.; Ko, S.R.; Choi, K.J.; Park, J.H.; Im, D.S. Effects of ginsenosides Rg3 and Rh2 on the proliferation of prostate cancer cells. Arch. Pharmacol. Res. 2004, 27, 429. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, H.; Hou, J. Effects of 20(S)-ginsenosideRh2 and 20(R)-ginsenoside Rh2 on proliferation and apoptosis of human lung adenocarcinoma A549 cells. Chin. J. Chin. Mater. Med. 2011, 36, 1670–1674. [Google Scholar]
- Tan, J.W.; Kim, M.K. Neuro protective effects of Biochanin A against β-amyloid-induced neurotoxicity in PC12 cells via a mitochondrial-dependent apoptosis pathway. Molecules 2016, 5, 548. [Google Scholar] [CrossRef] [PubMed]
- Quan, K.; Liu, Q.; Wan, J.Y.; Zhao, Y.J.; Guo, R.Z.; Alolga, R.N.; Qi, L.W. Rapid preparation of rare ginsenosides by acid transformation and their structure-activity relationships against cancer cells. Sci. Rep. 2015, 5, 8598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.T.; Hung, H.C.; Chen, C.J.; Hsu, W.L.; Ying, C. Effects of the dietary phytoestrogen biochanin A on cell growth in the mammary carcinoma cell line MCF-7. J. Nutr. Biochem. 1999, 10, 510–517. [Google Scholar] [CrossRef]
- Moon, Y.J.; Shin, B.S.; An, G.; Morris, M.E. Morris, Biochanin A inhibits breast cancer tumor growth in a murine xenograft model. Pharm. Res. 2008, 9, 2158–2163. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Kim, T.W.; Singh, S.V. Ginsenoside Rh2-mediated G1phase cell cycle arrest in human breast cancer cells is caused by p15Ink4B and p27Kip1-dependent inhibition of cyclin-dependent kinases. Pharm. Res. 2009, 26, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Morris, M.E. Pharmacokinetics and bioavailability of the bioflavonoid biochanin A: Effects of quercetin and EGCG on biochanin A disposition in rats. Mol. Pharm. 2007, 4, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Eberding, A.; Madera, C.; Fazli, L.; Jia, W.; Goldenberg, L.; Gleave, M.; Guns, E.S. Rh2 synergistically enhances paclitaxel or mitoxantrone in prostate cancer models. J. Urol. 2006, 175, 1926–1931. [Google Scholar] [CrossRef]
- Xiao, P.; Zheng, B.W.; Sun, J.M.; Yang, J. Biochanin A induces anticancer effects in SK-Mel-28 human malignant melanoma cells via induction of apoptosis, inhibition of cell invasion and modulation of NF-kappa B and MAPK signaling pathways. Oncol. Lett. 2017, 5, 5989–5993. [Google Scholar] [CrossRef] [PubMed]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.Y.; Koh, M.S.; Moon, A. The p38 MAPK inhibitors for the treatment of inflammatory diseases and cancer. Expert Opin. Investig. Drug 2009, 18, 1893–1905. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Li, J.; Xia, J.; Jiang, R.; Zuo, G.W.; Li, X.P.; Chen, Y.; Xiong, W.; Chen, D.L. Ginsenoside 20(s)-Rh2 as potent natural histone deacetylase inhibitors suppressing the growth of human leukemia cells. Chem. Biol. Interact. 2015, 5, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jin, S.H. Ginsenoside Rh2 induces apoptosis via activation of caspase-1 and -3 and up-regulation of Bax in human neuroblastoma. Arch. Pharm. Res. 2004, 27, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, A.K. Anti-breast cancer activity of Fine Black ginseng (Panax ginseng Meyer) and ginsenoside Rg5. J. Ginseng Res. 2015, 39, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Laptenko, O.; Prives, C. Transcriptional regulation by p53: One protein, many possibilities. Cell Death Differ. 2006, 13, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.L.; Gu, W. Ubiquitination, phosphorylation and acetylation: The molecular basis for p53 regulation. Curr. Opin. Cell. Biol. 2003, 15, 164–171. [Google Scholar] [CrossRef]
- Felice, D.L.; Sun, J.; Liu, R.H. A modified methylene blue assay for accurate cell counting. J. Funct. Foods 2009, 1, 109–118. [Google Scholar] [CrossRef]
- Jiang, X.; Li, T.; Liu, R.H. 2α-Hydroxyursolic acid inhibited cell proliferation and induced apoptosis in MDA-MB-231 human breast cancer cells through the p38/MAPK signal transduction pathway. J. Agric. Food Chem. 2016, 64, 1806–1816. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer. Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, H.L.; Lee, M.H.; You, K.E.; Kwon, B.J.; Seo, H.J.; Park, J.C. Asiaticoside enhances normal human skin cell migration, attachment and growth in vitro wound healing model. Phytomedicine 2012, 19, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Tapia, P.A.; Argandona, F.; Palomino, W.A.; Devoto, L. Human chorionic gonadotropin (hCG) modulation of TIMP1 secretion by human endometrial stromal cells facilitates extravillous trophoblast invasion in vitro. Hum. Reprod. 2013, 28, 2215–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Zhu, Y.; Gao, Y.; Shi, Z.; Hu, Y.; Ren, G. Suppressive effects of saponin-enriched extracts from quinoa on 3T3-L1 adipocyte differentiation. Food Funct. 2015, 6, 3282–3290. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds Combined Biochanin A and Ginsenoside Rh2 are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | EC50 Value | |||
|---|---|---|---|---|
| MDA-MB-231 | MCF-7 | |||
| Single | Combined | Single | Combined | |
| Biochanin A | 63.76 μM | 25.20 μM | 59.76 μM | 27.68 μM |
| Rh2 | 57.53 μM | 22.75 μM | 52.53 μM | 25.41 μM |
| CI Values at Different Inhibition of Rates | |||
|---|---|---|---|
| 50% | 75% | 90% | 95% |
| 0.435 | 0.553 | 0.723 | 0.882 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, G.; Shi, Z.; Teng, C.; Yao, Y. Antiproliferative Activity of Combined Biochanin A and Ginsenoside Rh2 on MDA-MB-231 and MCF-7 Human Breast Cancer Cells. Molecules 2018, 23, 2908. https://doi.org/10.3390/molecules23112908
Ren G, Shi Z, Teng C, Yao Y. Antiproliferative Activity of Combined Biochanin A and Ginsenoside Rh2 on MDA-MB-231 and MCF-7 Human Breast Cancer Cells. Molecules. 2018; 23(11):2908. https://doi.org/10.3390/molecules23112908
Chicago/Turabian StyleRen, Guixing, Zhenxing Shi, Cong Teng, and Yang Yao. 2018. "Antiproliferative Activity of Combined Biochanin A and Ginsenoside Rh2 on MDA-MB-231 and MCF-7 Human Breast Cancer Cells" Molecules 23, no. 11: 2908. https://doi.org/10.3390/molecules23112908