
Tracing Actin Filament Bundles in Three-Dimensional Electron Tomography Density Maps of Hair Cell Stereocilia

Abstract
:1. Introduction
2. Results
2.1. Longitudinal Average along an Estimated Direction of the Bundle
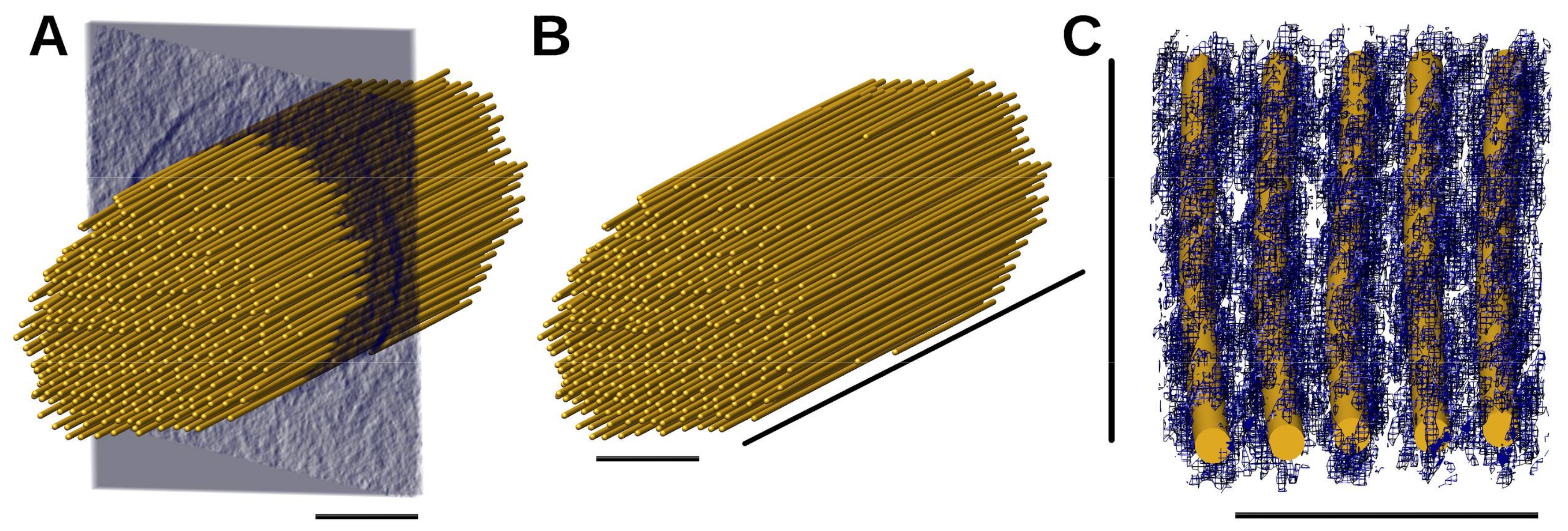
2.2. The Manually Built Model of Actin Filaments
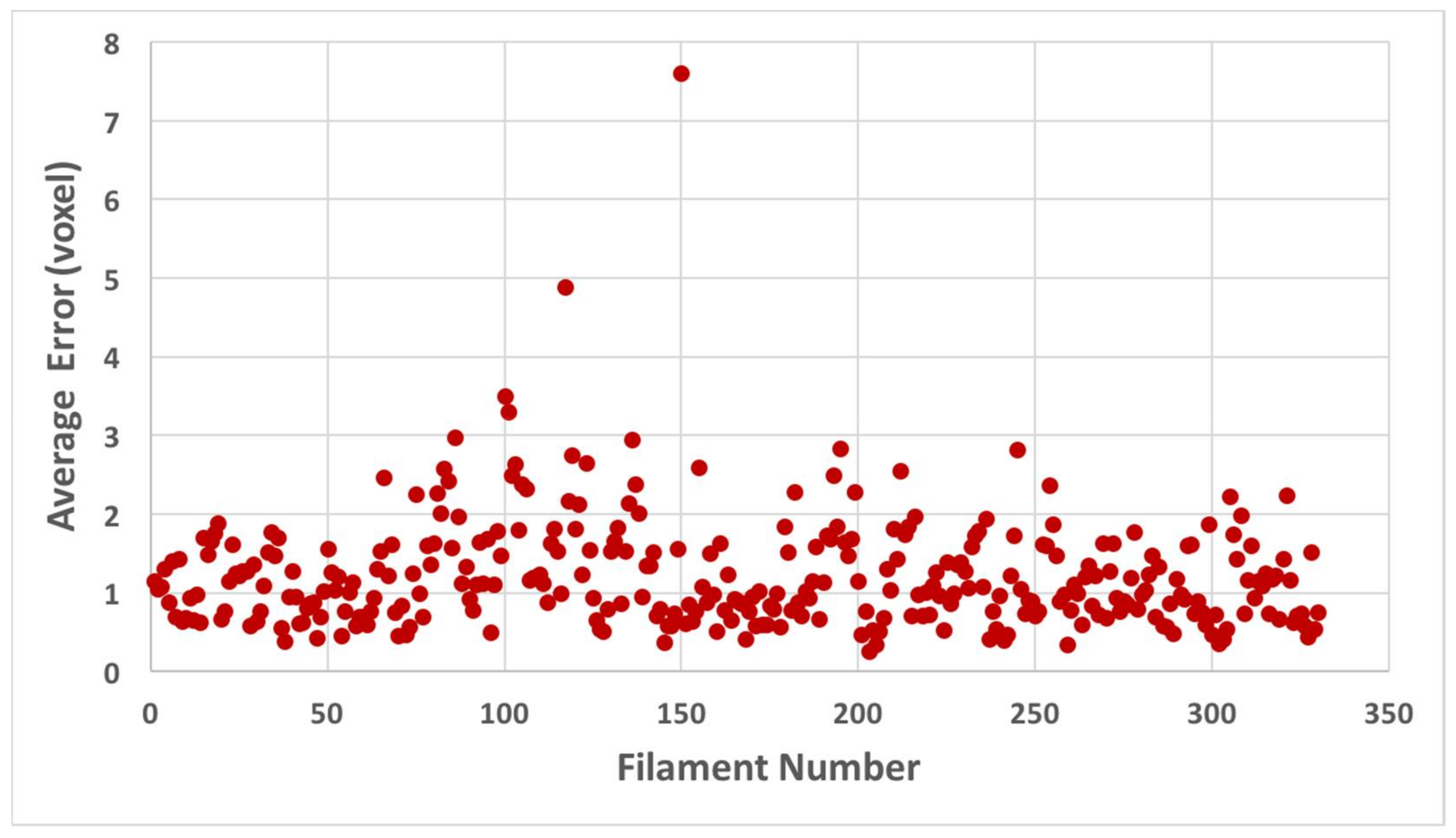
2.3. Filaments Detected Using BundleTrac
2.4. The Effect of Longitudinal Averaging and Seven-Peak Convolution
2.5. Local Polynomial Regression for Denoising
3. Materials and Methods
3.1. Cryo-Tomography of Inner Ear Sensory Epithelial Hair Cell Stereocilia
3.2. Manual Filament Tracing
3.3. BundleTrac
3.4. Quantification of the Cross-Distance between Two Sets of Filaments
3.5. A More Specialized Denoising Method
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hudspeth, A.J. How hearing happens. Neuron 1997, 19, 947–950. [Google Scholar] [CrossRef]
- LeMasurier, M.; Gillespie, P.G. Hair-cell mechanotransduction and cochlear amplification. Neuron 2005, 48, 403–415. [Google Scholar] [PubMed]
- Kwan, T.; White, P.M.; Segil, N. Development and regeneration of the inner ear. Ann. N. Y. Acad. Sci. 2009, 1170, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Frolenkov, G.I.; Belyantseva, I.A.; Friedman, T.B.; Griffith, A.J. Genetic insights into the morphogenesis of inner ear hair cells. Nat. Rev. Genet. 2004, 5, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.; Richardson, G.P. Linking genes underlying deafness to hair-bundle development and function. Nat. Neurosci. 2009, 12, 703–710. [Google Scholar] [PubMed]
- Groves, A.K.; Zhang, K.D.; Fekete, D.M. The genetics of hair cell development and regeneration. Annu. Rev. Neurosci. 2013, 36, 361–381. [Google Scholar] [CrossRef] [PubMed]
- Barr-Gillespie, P.G. Assembly of hair bundles, an amazing problem for cell biology. Mol. Biol. Cell 2015, 26, 2727–2732. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.B.; Krey, J.F.; Hassan, A.; Metlagel, Z.; Tauscher, A.N.; Pagana, J.M.; Sherman, N.E.; Jeffery, E.D.; Spinelli, K.J.; Zhao, H.; et al. Molecular architecture of the chick vestibular hair bundle. Nat. Neurosci. 2013, 16, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Metlagel, Z.; Krey, J. Cryo-Electron Tomography of Hair Cell Stereocilia in their Unstained, Frozen-Hydrated Stat. 2018; Manuscript in preparation. [Google Scholar]
- Irobalieva, R.N.; Martins, B.; Medalia, O. Cellular structural biology as revealed by cryo-electron tomography. J. Cell Sci. 2016, 129, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.; Starosolski, Z.; Wahle, M.; Rigort, A.; Wriggers, W. Automated tracing of filaments in 3D electron tomography reconstructions using Sculptor and Situs. J. Struct. Biol. 2012, 178, 121–128. [Google Scholar] [PubMed]
- Elosegui-Artola, A.; Jorge-Penas, A.; Moreno-Arotzena, O.; Oregi, A.; Lasa, M.; Garcia-Aznar, J.M.; De Juan-Pardo, E.M.; Aldabe, R. Image analysis for the quantitative comparison of stress fibers and focal adhesions. PLoS ONE 2014, 9, e107393. [Google Scholar] [CrossRef] [PubMed]
- Eltzner, B.; Wollnik, C.; Gottschlich, C.; Huckemann, S.; Rehfeldt, F. The filament sensor for near real-time detection of cytoskeletal fiber structures. PLoS ONE 2015, 10, e0126346. [Google Scholar] [CrossRef] [PubMed]
- Herberich, G.W.T.; Sechi, A.; Windoffer, R.; Leube, R.; Aach, T. Fluorescence microscopic imaging and image analysis of the cytoskeleton. In Proceedings of the Asilomar Conference on Signals, Systems and Computers, Pacific Grove, CA, USA, 7–10 November 2010; pp. 1359–1363. [Google Scholar]
- Nguyen, U.T.; Bhuiyan, A.; Park, L.A.; Ramamohanarao, K. An effective retinal blood vessel segmentation method using multi-scale line detection. Pattern Recognit. 2013, 46, 703–715. [Google Scholar] [CrossRef]
- Weichsel, J.; Urban, E.; Small, J.V.; Schwarz, U.S. Reconstructing the orientation distribution of actin filaments in the lamellipodium of migrating keratocytes from electron microscopy tomography data. Cytometry A 2012, 81, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Haralick, R.M. Ridges and valleys on digital images. Comput. Vis. Gr. Image Process. 1983, 22, 28–38. [Google Scholar] [CrossRef]
- Kirbas, C.; Quek, F. A review of vessel extraction techniques and algorithms. ACM Comput. Surv. 2004, 36, 81–121. [Google Scholar] [CrossRef]
- Alioscha-Perez, M.; Benadiba, C.; Goossens, K.; Kasas, S.; Dietler, G.; Willaert, R.; Sahli, H. A Robust Actin Filaments Image Analysis Framework. PLoS Comput. Biol. 2016, 12, e1005063. [Google Scholar] [CrossRef] [PubMed]
- Herberich, G.; Windoffer, R.; Leube, R.; Aach, T. 3D segmentation of keratin intermediate filaments in confocal laser scanning microscopy. In Conference Proceedings: Annual International Conference of the IEEE Engineering in Medicine and Biology Society, IEEE Engineering in Medicine and Biology Society Annual Conference; IEEE: Piscataway Township, NJ, USA, 2011; pp. 7751–7754. [Google Scholar]
- Mukherjee, S.; Condron, B.; Acton, S.T. Tubularity flow field—A technique for automatic neuron segmentation. IEEE Trans. Image Process. 2015, 24, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Condron, B.; Aksel, A.; Acton, S. Segmentation and tracing of single neurons from 3D confocal microscope images. IEEE J. Biomed. Health Inform. 2013, 17, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Sampo, J.A.; Takalo, J.J.; Siltanen, S.; Miettinen, A.; Lassas, M.; Timonen, J. (Eds.) Curvelet-based method for orientation estimation of particles from optical images. Opt. Eng. 2014, 53, 033109. [Google Scholar]
- Möller, B.; Piltz, E.; Bley, N. (Eds.) Quantification of Actin Structures Using Unsupervised Pattern Analysis Techniques. In Proceedings of the 2014 22nd International Conference on Pattern Recognition, Stockholm, Sweden, 24–28 August 2014. [Google Scholar]
- Moch, M.; Herberich, G.; Aach, T.; Leube, R.E.; Windoffer, R. Measuring the regulation of keratin filament network dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 10664–10669. [Google Scholar] [CrossRef] [PubMed]
- Weichsel, J.; Schwarz, U.S. Two competing orientation patterns explain experimentally observed anomalies in growing actin networks. Proc. Natl. Acad. Sci. USA 2010, 107, 6304–6309. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Si, D.; Zeng, T.; Ji, S.; He, J. (Eds.) Deep convolutional neural networks for detecting secondary structures in protein density maps from cryo-electron microscopy. In Proceedings of the 2016 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Shenzhen, China, 15–16 December 2016. [Google Scholar]
- Si, D.; Ji, S.; Al Nasr, K.; He, J. A machine learning approach for the identification of protein secondary structure elements from electron cryo-microscopy density maps. Biopolymers 2012, 97, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.; Wriggers, W. Evolutionary bidirectional expansion for the tracing of alpha helices in cryo-electron microscopy reconstructions. J. Struct. Biol. 2012, 177, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.L.; Ju, T.; Chiu, W. Identification of secondary structure elements in intermediate-resolution density maps. Structure 2007, 15, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Dal Palu, A.; He, J.; Pontelli, E.; Lu, Y. Identification of Alpha-Helices from Low Resolution Protein Density Maps. In Proceedings of the Computational Systems Bioinformatics Conference (CSB), Stanford, CA, USA, 14–18 August 2006. [Google Scholar]
- Jiang, W.; Baker, M.L.; Ludtke, S.J.; Chiu, W. Bridging the information gap: Computational tools for intermediate resolution structure interpretation. J. Mol. Biol. 2001, 308, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Birmanns, S.; Rusu, M.; Wriggers, W. Using Sculptor and Situs for simultaneous assembly of atomic components into low-resolution shapes. J. Struct. Biol. 2011, 173, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Redemann, S.; Weber, B.; Möller, M.; Verbavatz, J.M.; Hyman, A.A.; Baum, D.; Prohaska, S.; Müller-Reichert, T. The Segmentation of Microtubules in Electron Tomograms Using Amira. In Mitosis: Methods and Protocols; Humana Press: New York, NY, USA, 2014; pp. 261–278. [Google Scholar]
- Weber, B.; Greenan, G.; Prohaska, S.; Baum, D.; Hege, H.C.; Müller-Reichert, T.; Hyman, A.A.; Verbavatz, J.M. Automated tracing of microtubules in electron tomograms of plastic embedded samples of Caenorhabditis elegans embryos. J. Struct. Biol. 2012, 178, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Redemann, S.; Baumgart, J.; Lindow, N.; Shelley, M.; Nazockdast, E.; Kratz, A.; Prohaska, S.; Brugués, J.; Fürthauer, S.; Müller-Reichert, T. C. elegans chromosomes connect to centrosomes by anchoring into the spindle network. Nat. Commun. 2017, 8, 15288. [Google Scholar] [CrossRef] [PubMed]
- Rigort, A.; Günther, D.; Hegerl, R.; Baum, D.; Weber, B.; Prohaska, S.; Medalia, O.; Baumeister, W.; Hege, H.C. Automated segmentation of electron tomograms for a quantitative description of actin filament networks. J. Struct. Biol. 2012, 177, 135–144. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Schmid, M.F.; Zhou, Z.H.; Rixon, F.; Chiu, W. Finding and using local symmetry in identifying lower domain movements in hexon subunits of the herpes simplex virus type 1 B capsid. J. Mol. Biol. 2001, 309, 903–914. [Google Scholar] [PubMed]
- Kremer, J.R.; Mastronarde, D.N.; McIntosh, J.R. Computer visualization of three-dimensional image data using IMOD. J. Struct. Biol. 1996, 116, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Mastronarde, D.N.; Held, S.R. Automated tilt series alignment and tomographic reconstruction in IMOD. J. Struct. Biol. 2017, 197, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Stephanie, Z.; Julio, K.; Willy, W.; Jing, H. Comparing an Atomic Model or Structure to a Corresponding Cryo-electron Microscopy Image at the Central Axis of a Helix. J. Comput. Biol. 2017, 24, 52–67. [Google Scholar]
- Starosolski, Z.; Szczepanski, M.; Wahle, M.; Rusu, M.; Wriggers, W. Developing a denoising filter for electron microscopy and tomography data in the cloud. Biophys. Rev. 2012, 4, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Wand, M.P.; Jones, M.C. Kernel Smoothing, 1st ed.; Chapman & Hall: London, UK, 1995. [Google Scholar]
Sample Availability: Not available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Row | Implementation | Avg_L a | Gauss b | 7 Peaks c | 1 Peak d | AvgError e |
|---|---|---|---|---|---|---|
| 1 | Trace_L_G_7 | ✔ | ✔ | ✔ | ✖ | 1.300 |
| 2 | Trace_L_7 | ✔ | ✖ | ✔ | ✖ | 1.345 |
| 3 | Trace_G_7 | ✖ | ✔ | ✔ | ✖ | 1.523 |
| 4 | Trace_L_1 | ✔ | ✖ | ✖ | ✔ | 2.157 |
| 5 | Trace_L_G_1 | ✔ | ✔ | ✖ | ✔ | 2.420 |
| Improvement in the seven-peak convolution f | 46.28% | |||||
| Improvement in the longitudinal average g | 14.62% | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sazzed, S.; Song, J.; Kovacs, J.A.; Wriggers, W.; Auer, M.; He, J. Tracing Actin Filament Bundles in Three-Dimensional Electron Tomography Density Maps of Hair Cell Stereocilia. Molecules 2018, 23, 882. https://doi.org/10.3390/molecules23040882
Sazzed S, Song J, Kovacs JA, Wriggers W, Auer M, He J. Tracing Actin Filament Bundles in Three-Dimensional Electron Tomography Density Maps of Hair Cell Stereocilia. Molecules. 2018; 23(4):882. https://doi.org/10.3390/molecules23040882
Chicago/Turabian StyleSazzed, Salim, Junha Song, Julio A. Kovacs, Willy Wriggers, Manfred Auer, and Jing He. 2018. "Tracing Actin Filament Bundles in Three-Dimensional Electron Tomography Density Maps of Hair Cell Stereocilia" Molecules 23, no. 4: 882. https://doi.org/10.3390/molecules23040882