Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Chemical and Spectroscopic Characterization of AA309
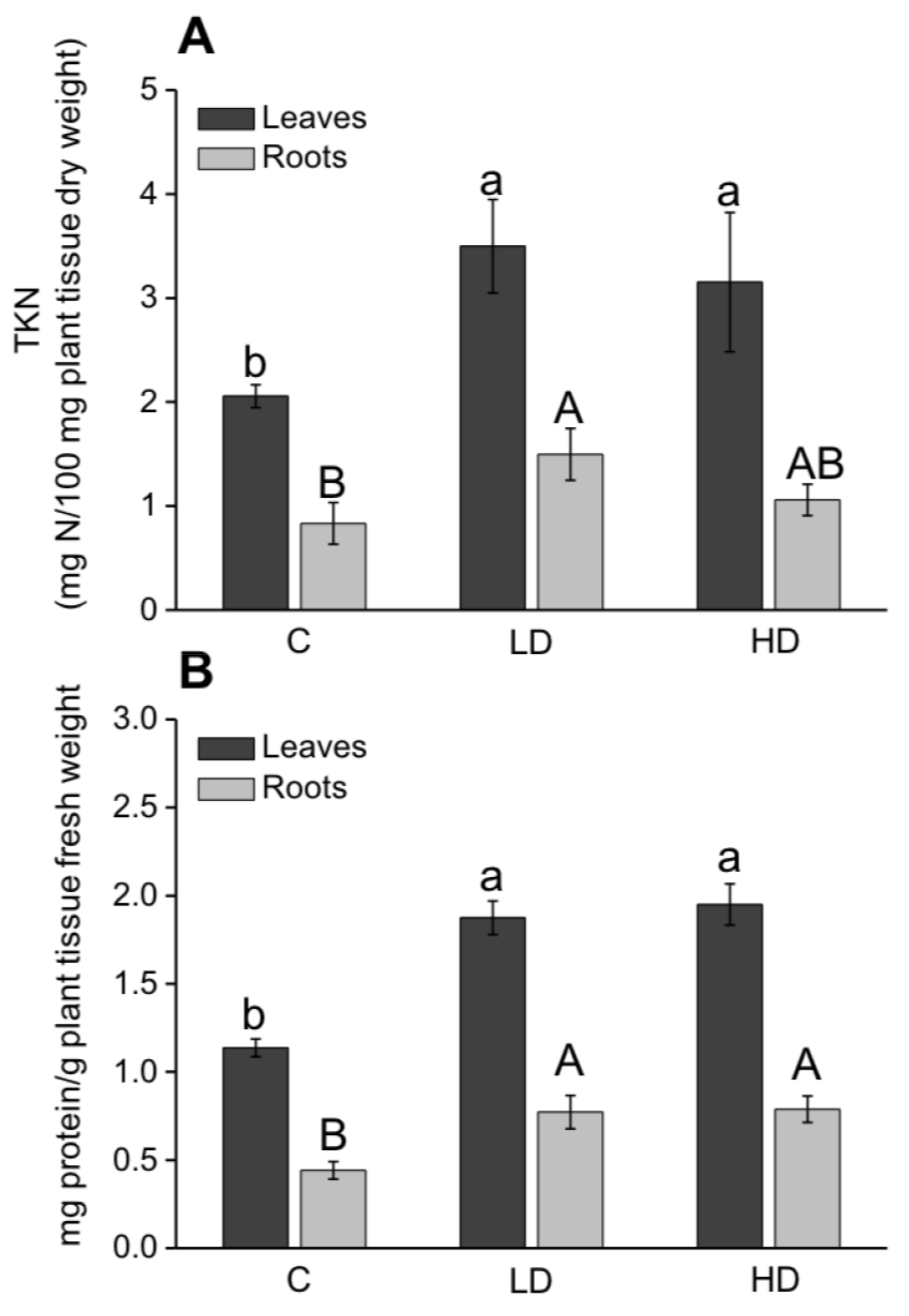
2.2. Effect on Leaf and Root Dry Weight
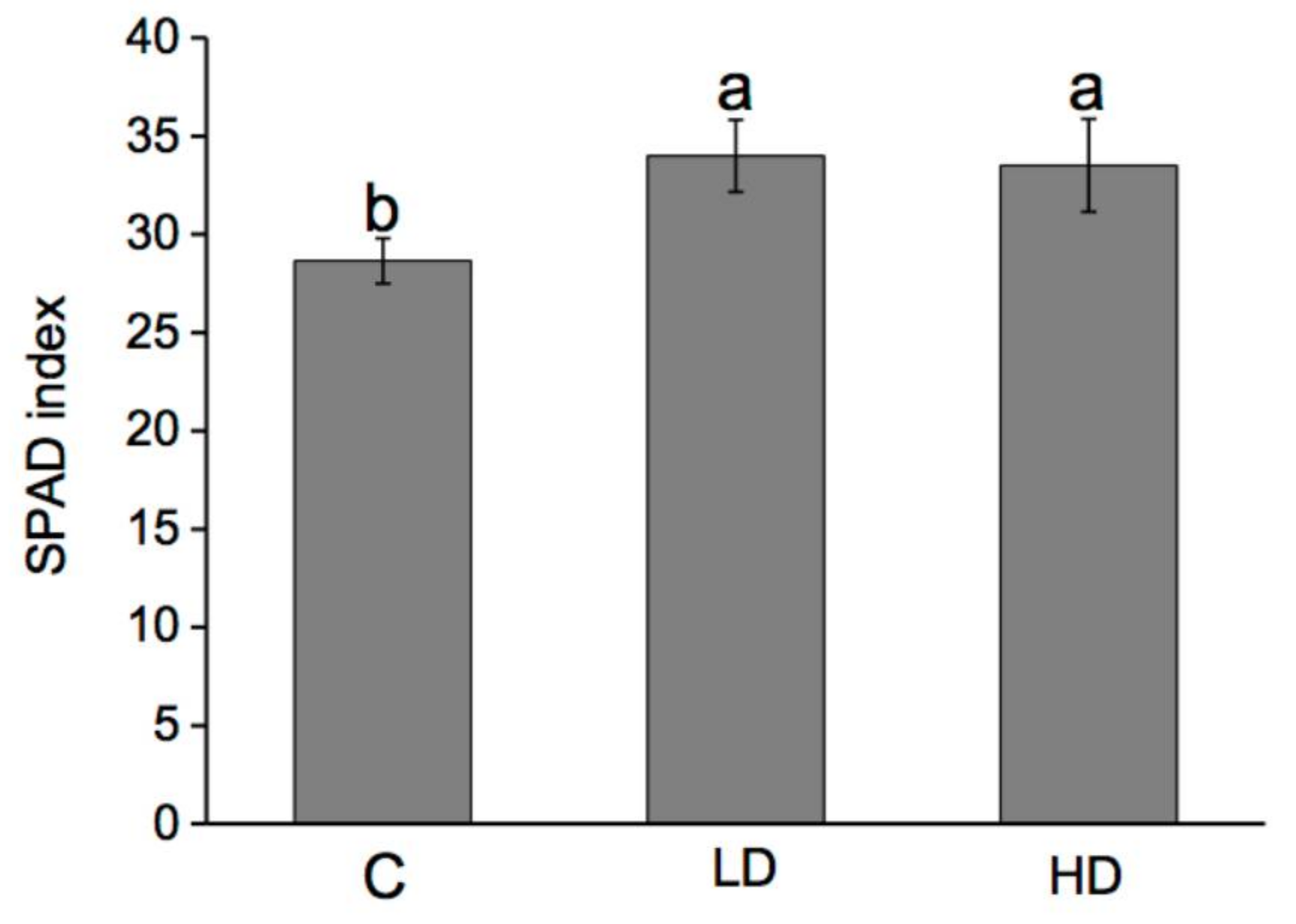
2.3. Effect on SPAD Index, Nitrogen, and Protein Content
2.4. Analyses of Phenolic Acids
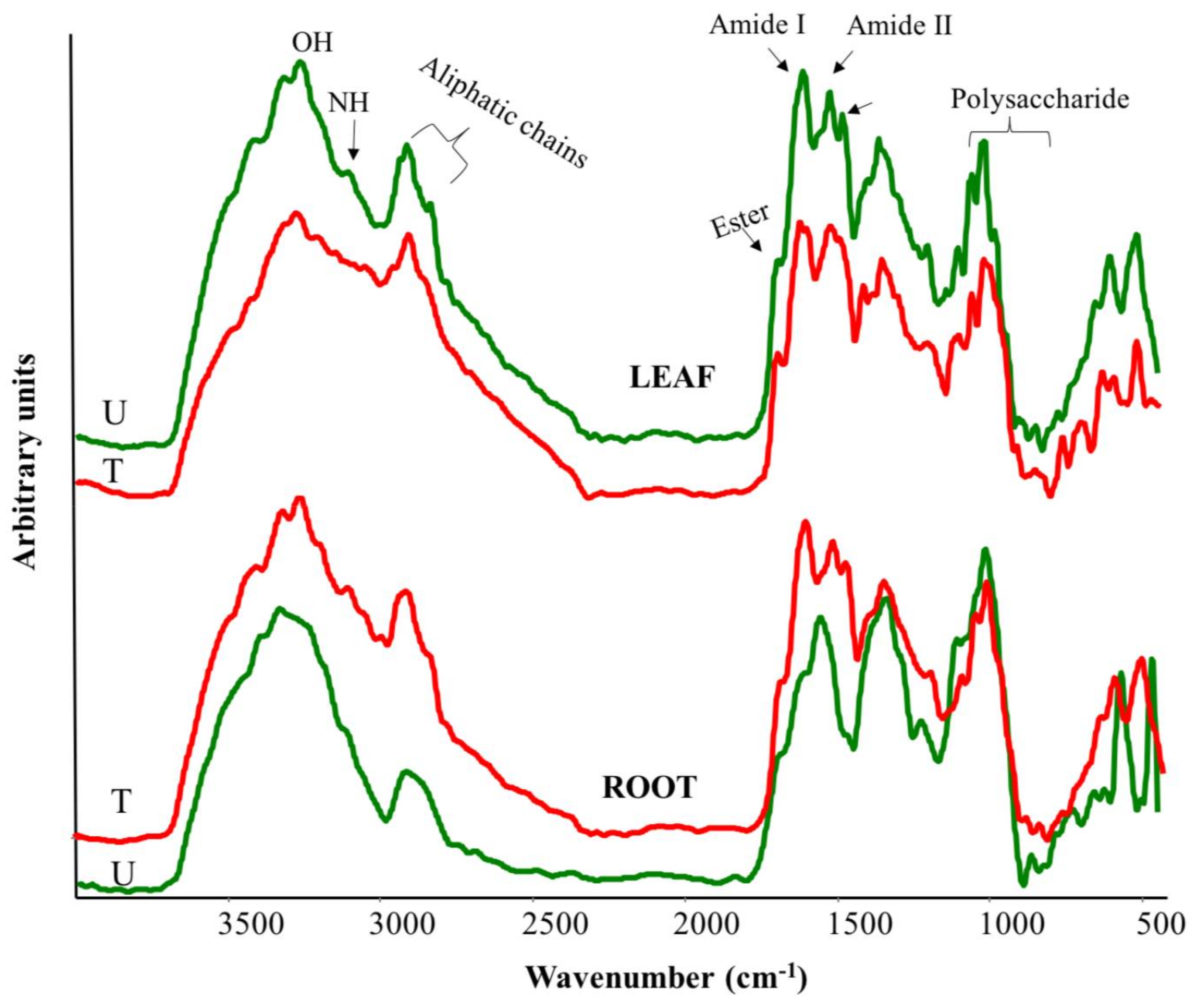
2.5. FTIR Spectroscopy on Plant Material
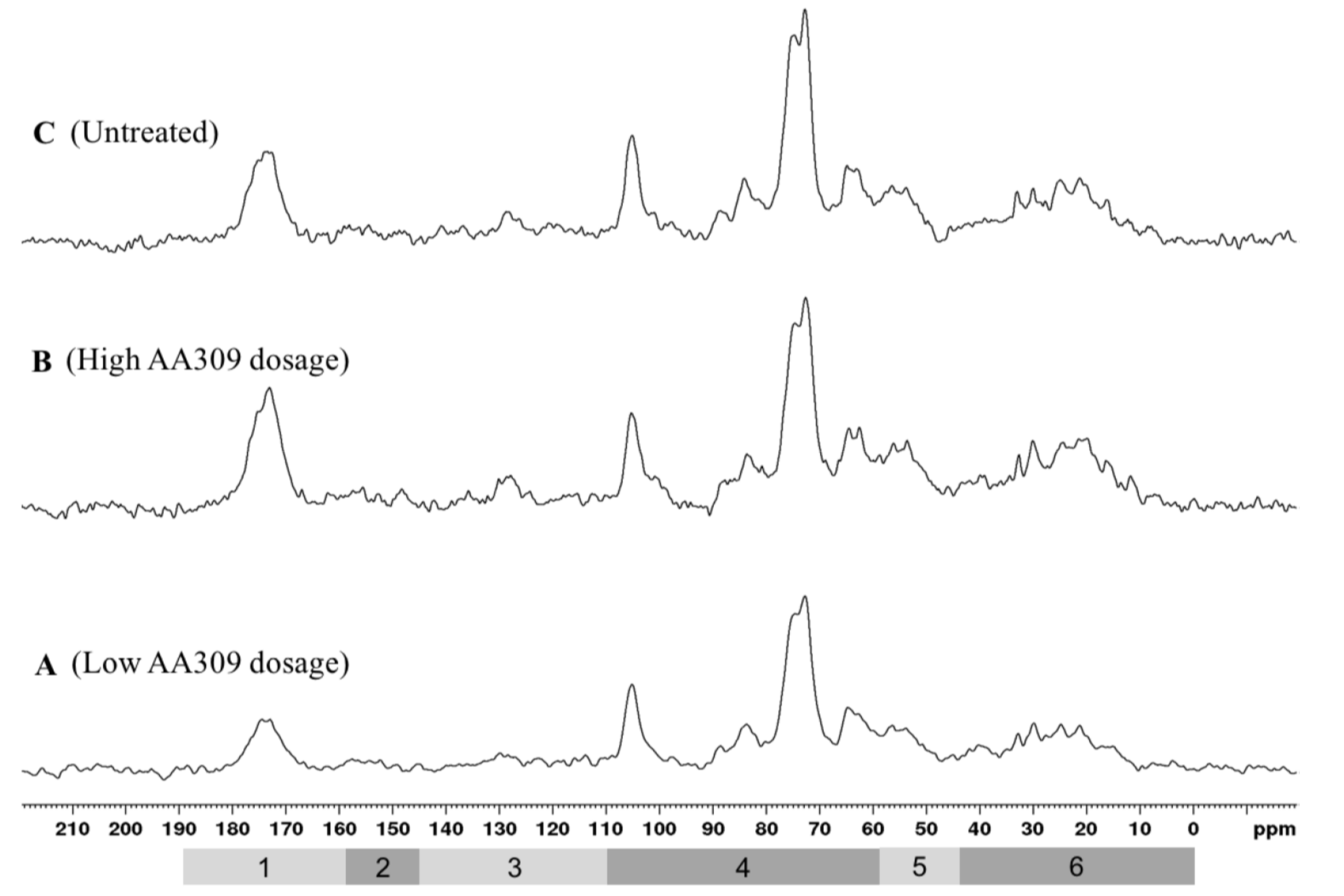
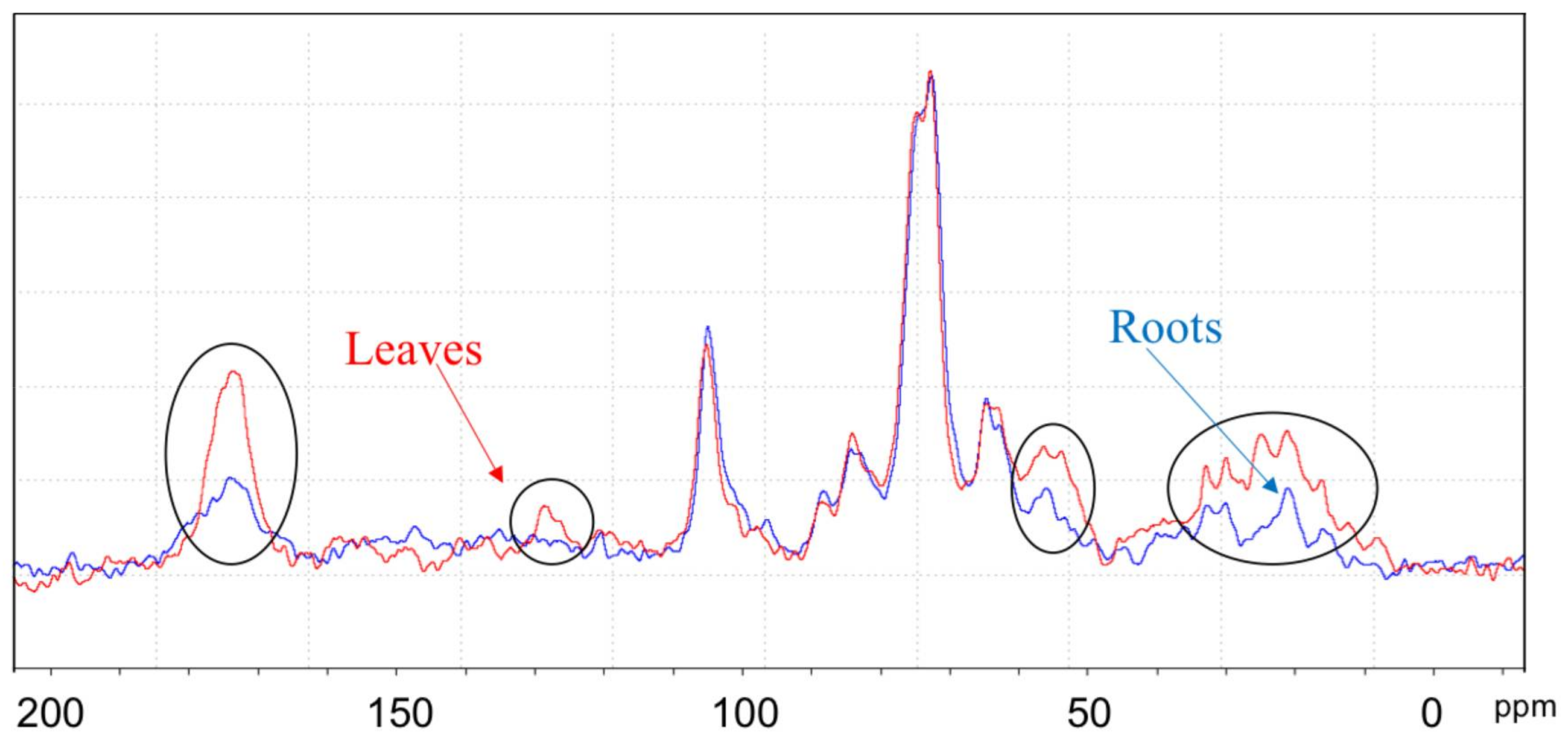
2.6. NMR Spectroscopy
3. Discussion
4. Material and Methods
4.1. Chemical Characterization of the Protein Hydrolysate
FTIR Analysis of AA309
4.2. Plant Material
4.2.1. Growth Conditions
4.2.2. SPAD Index, Nitrogen, Protein, and Phenolics
4.2.3. FITR and NMR Spectroscopies
Solid-State MAS NMR
Semi-Solid-State HR MAS NMR
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Aremu, M.O.; Olaofe, O.; Basu, S.K.; Abdulazeez, G.; Acharya, S.N. Processed cranberry bean (Phaseolus coccineus L.) seed flour for the African diet. Can. J. Plant Sci. 2010, 90, 719–728. [Google Scholar] [CrossRef]
- Lal, R. Constraints to adopting no-till farming in developing countries. Soil Tillage Res. 2007, 94, 1–3. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Tognoni, F.; Serra, G.; Ferrante, A.; Piaggesi, A. Use of biostimulants for reducing nutrient solution concentration in floating system. Acta Hortic. 2006, 718, 477–484. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Trentin, A.; Malagoli, M.; Nardi, S. Effect of an alfalfa plant-derived biostimulant on sulfur nutrition in tomato plants. In Proceedings of the 9th International Workshop on Sulfur Metabolism in Plants: Molecular Physiology and Ecophysiology of Sulfur; International Plant Sulfur Workshop Book Series; Springer: Berlin, Germany, 2015; pp. 215–220. [Google Scholar]
- Sharma, H.S.S.; Fleming, C.; Selby, C.J.R.; Trevor, R.; Martin, G. Plant biostimulants: A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 2014, 26, 465. [Google Scholar] [CrossRef]
- Zalabák, D.; Pospíšilová, H.; Šmehilová, M.; Mrízová, K.; Frébort, I.; Galuszka, P. Genetic engineering of cytokinin metabolism: Prospective way to improve agricultural traits of crop plants. Biotechnol. Adv. 2013, 31, 97–117. [Google Scholar] [CrossRef] [PubMed]
- European Biostimulants Industry Council (EBIC). Biostimulants in Brief; EBIC: Cambridge, UK, 2012. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 22, 353–360. [Google Scholar] [CrossRef]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange and leaf anatomy of zucchini squash grown under saline conditions. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Altissimo, A.; Franceschi, A.; Nardi, S. Phenol-containing organic substances stimulate phenylpropanoid metabolism in Zea mays. J. Plant Nutr. Soil Sci. 2011, 174, 496–503. [Google Scholar] [CrossRef]
- Aremu, A.O.; Masondo, N.A.; Rengasamy, K.R.R.; Amoo, S.O.; Gruz, J.; Bíba, O.; Šubrtová, M.; Pěnčík, A.; Novák, O.; Doležal, K.; et al. Physiological role of phenolic biostimulants isolated from brown seaweed Ecklonia maxima on plant growth and development. Planta 2015, 241, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of TCA cycle and N metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef] [PubMed]
- Baglieri, A.; Cadili, V.; Mozzetti, C.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Utilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil. 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Ertani, A.; Francioso, O.; Tugnoli, V.; Righi, V.; Nardi, S. Effect of commercial Lignosulfonate-Humate on Zea mays L. metabolism. J. Agric. Food Chem. 2011, 59, 11940–11948. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Pizzeghello, D.; Baglieri, A.; Cadili, V.; Tambone, F.; Gennari, M.; Nardi, S. Humic-like substances from agro-industrial residues affect growth and nitrogen assimilation in maize (Zea mays L.) plantlets. J. Geochem. Exp 2013, 129, 103–111. [Google Scholar] [CrossRef]
- Guinan, K.J.; Sujeeth, N.; Copeland, R.B.; Jones, P.W.; O’Brien, N.M.; Sharma, H.S.S.; Prouteau, P.F.J.; O’Sullivan, J.T. Discrete roles for extracts of Ascophyllum nodosum in enhancing plant growth and tolerance to abiotic and biotic stresses. Acta Hortic. 2013, 1–12. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. Hortic. Sci. 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Schaafsma, G. Safety of protein hydrolysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Pecha, J.; Fürst, T.; Kolomazník, K.; Friebrová, V.; Svoboda, P. Protein biostimulant foliar uptake modeling: The impact of climatic conditions. AIChE J. 2012, 58, 2010–2019. [Google Scholar] [CrossRef]
- Rentsch, D.; Schmidt, S.; Tegeder, M. Transporters for uptake and allocation of organic nitrogen compounds in plants. FEBS Lett. 2007, 581, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Apone, F.; Tito, A.; Carola, A.; Arciello, S.; Tortora, A.; Filippini, L.; Monoli, I.; Cucchiara, M.; Gibertoni, S.; Chrispeels, M.J.; et al. A mixture of peptides and sugars derived from plant cell walls increases plant defense responses to stress and attenuates age in molecular changes in cultured skin cells. J. Biotechnol. 2010, 145, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-wide identification of differentially expressed genes in Solanum lycopersicon L. in response to an alfalfa-protein hydrolysate using microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [PubMed]
- Santi, C.; Zamboni, A.; Varanini, Z.; Pandolfini, T. Growth stimulatory effects and genome-wide transcriptional changes produced by protein hydrolysates in maize seedlings front. Plant Sci. 2017, 8, 433. [Google Scholar]
- Sánchez-Gómez, R.; Garde-Cerdán, T.; Zalacain, A.; Garcia, R.; Cabrita, M.J. Salinas Vine-shoot waste aqueous extract applied as foliar fertilizer to grapevines: Effect on amino acids and fermentative volatile content. J. Agric. Food Chem. 2016, 197 Pt A, 132–140. [Google Scholar]
- Nashy, E. ; H.A.; Osman, O.; Mahmoud, A.A.; Ibrahim, M. Molecular spectroscopic study for suggested mechanism of chrome tanned leather. Spectrochim. Acta Part A 2012, 88, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yen, H.C. Early salt stress effects on the changes in chemical composition in leaves of ice plant and Arabidopsis. A fourier transform infrared spectroscopy study. Plant Physiol. 2002, 130, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Rao, C.N.R. Chemical Application of Infrared Spectroscopy 1963; Academic Press: New York, NY, USA; London, UK, 1964; 1441p. [Google Scholar]
- Conte, P.C.; Novotny, E.H.; Caponetto, G.; Laudicina, V.A.; Ciofalo, M.; Panno, M.; Palazzolo, E.; Badalucco, L.; Alonzo, G. CPMAS 13C NMR Characterization of leaves and litters from the reafforestated area of mustigarufi in Sicily (Italy). Open Magn. Res. J. 2010, 3, 89–95. [Google Scholar] [CrossRef]
- De Marco, A.; Spaccini, R.; Vittozzi, P.; Esposito, F.; Berg, B.; De Santo, A.V. Decomposition of black locust and black pine leaf litter in two coeval forest stands on Mount Vesuvius and dynamics of organic components assessed through proximate analysis and NMR spectroscopy. Soil Biol. Biochem. 2012, 51, 1–15. [Google Scholar] [CrossRef]
- Matsumiya, Y.; Kubo, M. Soybean peptide: Novel plant growth promoting peptide from soybean. In Soybean and Nutrition; El-Shemy, H., Ed.; InTech Europe Publisher: Rijeka, Croatia, 2011; pp. 215–230. [Google Scholar]
- Furuya, S.; Umemiya, Y. The influence of chemical forms on foliar-appliednitrogen absorption foreach trees. Horticulture 2002, 594, 97–103. [Google Scholar]
- Chen, L.; Bush, D.R. LHT1, a lysine- and histidine-specific amino acid transporter in Arabidopsis. Plant Physiol. 1997, 115, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Huggins, D.; Grant, G.H.; Knight, M.R.; Campbell, M.M. A role for glycine in the gating of plant NMDA-like receptors. Plant J. 2003, 35, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Cavani, L.; Ter Halle, A.; Richard, C.; Ciavatta, C. Photosensitizing properties of protein hydrolysate-based fertilizers. J. Agric. Food Chem. 2006, 54, 9160–9167. [Google Scholar] [CrossRef] [PubMed]
- Nardi, S.; Ertani, A.; Francioso, O. Soil–root cross-talking: The role of humic substances. J. Plant Nutr. Soil Sci. 2017, 180, 5–13. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ildebrandt, T.M.; Nunes-Nesi, A.; Araújo, W.L.N. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Tinti, A.; Nardi, S. Biological activity of vegetal extracts containing phenols on plant metabolism. Molecules 2016, 21, 205. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; De Stefani, D.; Bononi, A.; Rizzuto, R.; Pinton, P. Structural and functional link between the mitochondrial network and the endoplasmic reticulum. Int. J. Biochem. Cell Biol. 2009, 41, 1817–1827. [Google Scholar] [CrossRef] [PubMed]
- Nardi, S.; Tosoni, M.; Pizzeghello, D.; Provenzano, M.R.; Cilenti, A.; Sturaro, A.; Rella, R.; Vianello, A. Chemical characteristics and biological activity of organic substances extracted from soils by root exudates. Soil Sci. Soc. Am. J. 2005, 69, 2012–2019. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef]
- Bradford, M.A. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry; Freeman and Company: San Francisco, CA, USA, 1969; p. 776. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | ° | Unit |
|---|---|---|
| pH | 4 | |
| Electric conductivity | 1.08 | dS/m |
| Ash | 81.0 | g/kg |
| Organic N | 134.0 | g/kg |
| NH4+ | 2.0 | g/kg |
| Total N (TN) | 136.0 | g/kg |
| Total C (TC) | 400.0 | g/kg |
| Total SO3 | 47.0 | g/kg |
| Na2O | 10.0 | g/kg |
| CaO | 6.7 | g/kg |
| Cr | 2.6 | g/kg |
| K2O | 130 | mg/kg |
| Cu | 5.40 | mg/kg |
| Cd | <1 | mg/kg |
| Pb | <5 | mg/kg |
| Ni | 2.00 | mg/kg |
| Zn | 13.00 | mg/kg |
| Mn | 5.50 | mg/kg |
| Amino Acid | g/kg |
|---|---|
| Aspartate | 0.129 |
| Glutamate | 0.066 |
| Serine | 0.027 |
| Glycine | 0.099 |
| Threonine | 0.026 |
| Arginine | 0.053 |
| Alanine | 0.038 |
| Tyrosine | 0.033 |
| Cysteine | 0.075 |
| Valine | 0.048 |
| Methionine | 0.060 |
| Phenylalanine | 0.101 |
| Isoleucine | 0.093 |
| Leucine | 0.037 |
| Lysine | 0.170 |
| Caffeic | p-Coumaric | Ferulic | Hydroxybenzoic | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | µg/g dw | |||||||||||
| Leaves | ||||||||||||
| C | 26.91 | ±2.22 c* | 95.70 | ±5.23 a | 32.39 | ±8.22 a | 97.05 | ±21.54 a | ||||
| LD | 43.05 | ±1.54 b | 111.03 | ±13.52 a | 21.34 | ±8.56 a | 86.55 | ±19.02 a | ||||
| HD | 67.96 | ±5.23 a | 75.19 | ±11.23 a | 20.80 | ±9.21 a | 95.67 | ±16.63 a | ||||
| Roots | ||||||||||||
| C | 19.01 | ±3.12 a | 21.83 | ±5.23 b | 8.79 | ±0.56 a | 21.66 | ±2.77 b | ||||
| LD | 6.62 | ±0.98 c | 67.22 | ±12.88 a | 3.68 | ±0.12 b | 81.57 | ±12.03 a | ||||
| HD | 14.89 | ±1.05 b | 95.09 | ±15.26 a | 9.97 | ±0.65 a | 38.07 | ±8.28 b | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ertani, A.; Francioso, O.; Ferrari, E.; Schiavon, M.; Nardi, S. Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form. Molecules 2018, 23, 1031. https://doi.org/10.3390/molecules23051031
Ertani A, Francioso O, Ferrari E, Schiavon M, Nardi S. Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form. Molecules. 2018; 23(5):1031. https://doi.org/10.3390/molecules23051031
Chicago/Turabian StyleErtani, Andrea, Ornella Francioso, Erika Ferrari, Michela Schiavon, and Serenella Nardi. 2018. "Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form" Molecules 23, no. 5: 1031. https://doi.org/10.3390/molecules23051031
APA StyleErtani, A., Francioso, O., Ferrari, E., Schiavon, M., & Nardi, S. (2018). Spectroscopic-Chemical Fingerprint and Biostimulant Activity of a Protein-Based Product in Solid Form. Molecules, 23(5), 1031. https://doi.org/10.3390/molecules23051031