
Sterol Composition of Clinically Relevant Mucorales and Changes Resulting from Posaconazole Treatment

 , and
, and
Abstract
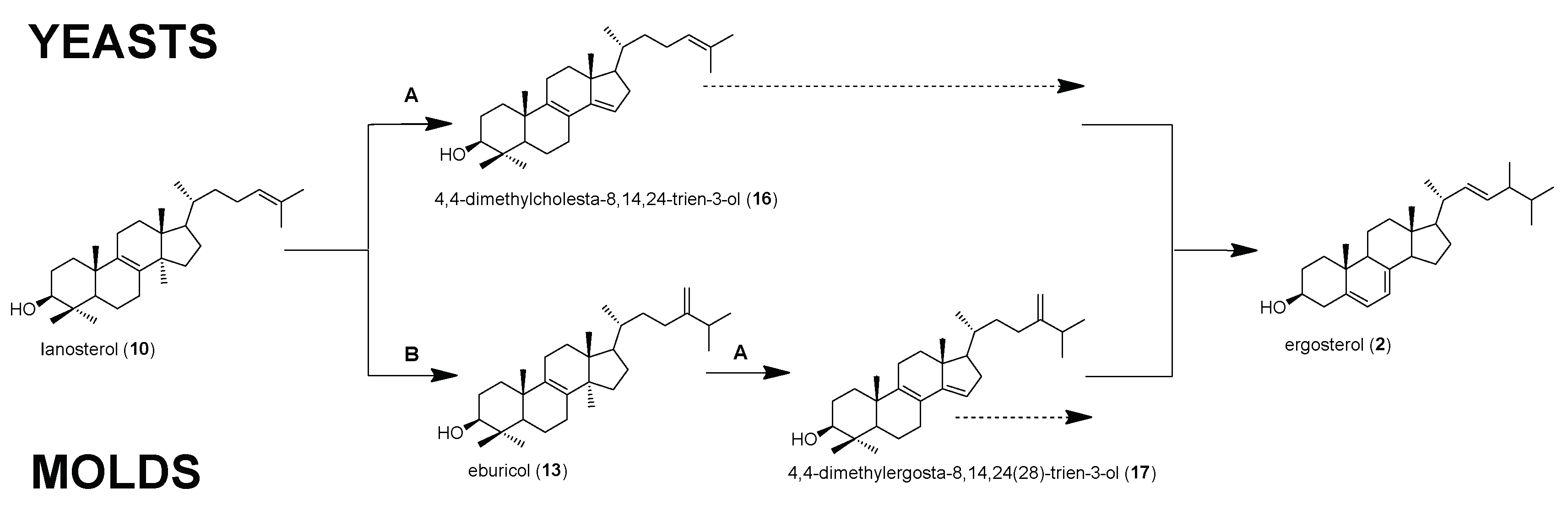
:1. Introduction
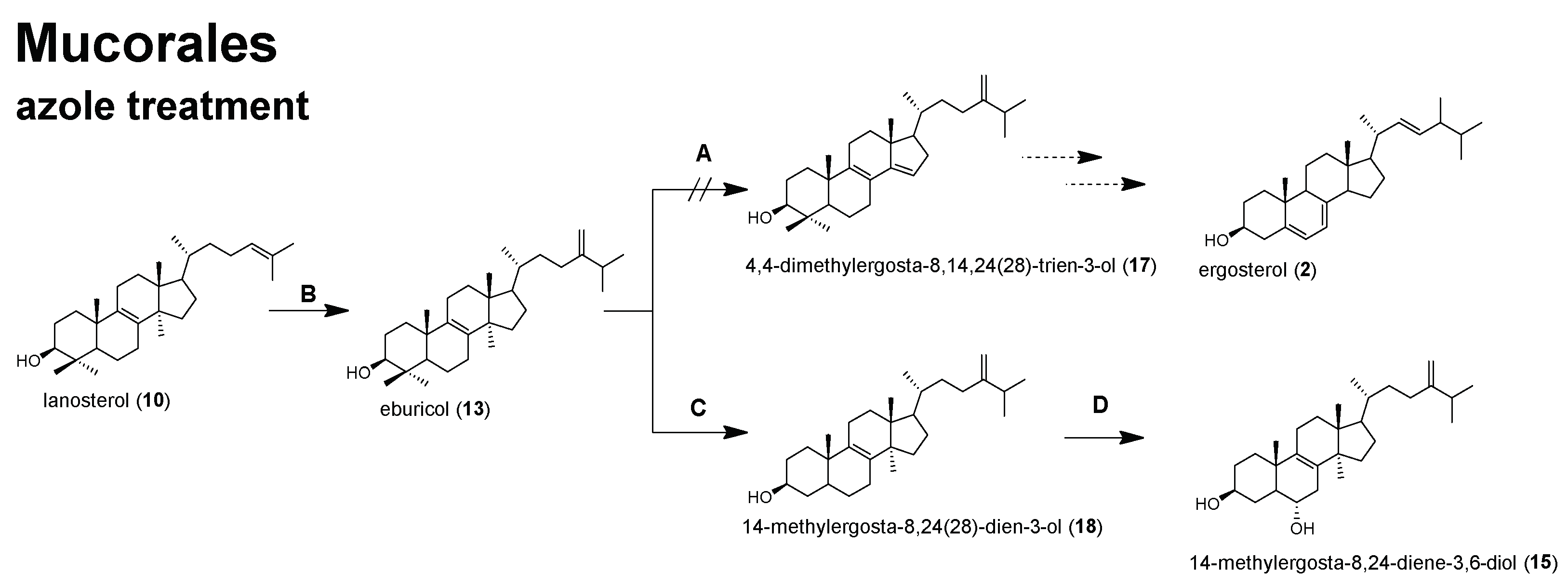
2. Results and Discussion
2.1. Relative and Absolute Amounts of Sterol Intermediates in Mucorales
2.2. Sterol Composition and Sterol Content of Posaconazole Treated R. arrhizus
3. Material and Methods
3.1. Fungal Strains Used in This Study
3.2. Fungal Growth and Culture Conditions
3.3. Sterol Extraction
3.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of Sterol TMS Ethers
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spellberg, B.; Edwards, J.; Ibrahim, A. Novel perspectives on mucormycosis: Pathophysiology, presentation and management. Clin. Microbiol. Rev. 2005, 18, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J. Taxonomy of fungi causing mucormycosis and entomophthoramycosis (zygomycosis) and nomenclature of the disease: Molecular mycologic perspectives. Clin. Infect. Dis. 2012, 54, S8–S15. [Google Scholar] [CrossRef] [PubMed]
- Morin-Sardin, S.; Nodet, P.; Coton, E.; Jany, J.-L. Mucor: A Janus-faced fungal genus with human health impact and industrial applications. Fungal Biol. Rev. 2017, 31, 12–32. [Google Scholar] [CrossRef]
- Dolatabadi, S.; Scherlach, K.; Figge, M.; Hertweck, C.; Dijksterhuis, J.; Menken, S.B.; de Hoog, G.S. Food preparation with mucoralean fungi: A potential biosafety issue. Fungal Biol. 2016, 120, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Kontoyiannis, D.P. Epidemiology and treatment of mucormycosis. Future Microbiol. 2013, 8, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.L.; Finley, P.J.; Mikkelson, D.R.; Tibbs, B. Mucormycosis: A rare fungal infection in tornado victims. J. Burn Care Res. 2014, 35, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.J.; Hiemenz, J.W.; Seibel, N.L.; Perfect, J.R.; Horwith, G.; Lee, L.; Silber, J.L.; DiNubile, M.J.; Reboli, A.; Bow, E.; et al. Ampothericin B lipid complex for invasive fungal infections: Analysis of safety and efficacy in 556 cases. Clin. Infect. Dis. 1998, 26, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, D.; Gagne, L.S.; Hammond, S.P.; Gilmore, E.T.; Joyce, A.C.; Soiffer, R.J.; Marty, F.M. Isavuconazole treatment of a patient with disseminated mucormycosis. J. Clin. Microbiol. 2014, 52, 1016–1019. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Arikan-Akdagli, S.; Dannaoui, E.; Groll, A.H.; Lagrou, K.; Chakrabarti, A.; Lanternier, F.; Pagano, L.; Skiada, A.; Akova, M.; et al. ESCMID and ECMM joint clinical guidelines for the diagnosis and management of mucormycosis 2013. Clin. Microbiol. Infect. 2014, 20, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Binder, U.; Sparber, M.; Lackner, M.; Caramalho, R.; Lass-Flörl, C. Susceptibility profiles of amphotericin B and posaconazole against clinical relevant Mucorales species under hypoxic conditions. Antimicrob. Agents Chemother. 2015, 59, 1344–1346. [Google Scholar] [CrossRef] [PubMed]
- Marty, F.M.; Ostrosky-Zeichner, L.; Cornely, O.A.; Mullane, K.M.; Perfect, J.R.; Thompson, G.R.; Alangaden, G.J.; Brown, J.M.; Fredricks, D.N.; Heinz, W.J.; et al. Isavuconazole treatment for mucormycosis: A single-arm open-label trial and case-control analysis. Lancet Infect. Dis. 2016, 16, 828–837. [Google Scholar] [CrossRef]
- McCreary, E.K.; Schulz, L.T.; Lepak, A.J. Isavuconazole: Has it saved us? A pharmacotherapy review and update on clinical experience. Curr. Treat. Options Infect. Dis. 2017, 9, 356–370. [Google Scholar] [CrossRef]
- Gray, K.C.; Palacios, D.S.; Dailey, I.; Endo, M.M.; Uno, B.E.; Wilcock, B.C.; Burke, M.D. Amphotericin primarily kills yeast by simply binding ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, C.; Trevijano-Contador, N.; Román, E.; Sánchez-Fresneda, R.; Casas, C.; Herrero, E.; Argüelles, J.C.; Pla, J.; Cuenca-Estrella, M.; Zaragoza, O. The production of reactive oxygen species is a universal action mechanism of amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drugs. Antimicrob. Agents Chemother. 2014, 58, 6627–6638. [Google Scholar] [CrossRef] [PubMed]
- Munayyer, H.K.; Mann, P.A.; Chau, A.S.; Yarosh-Tomaine, T.; Greene, J.R.; Hare, R.S.; Heimark, L.; Palermo, R.E.; Loenberg, D.; McNicholas, P.M. Posaconazole is a potent inhibitor of sterol 14α-demethylation in yeasts and molds. Antimicrob. Agents Chemother. 2004, 48, 3690–3696. [Google Scholar] [CrossRef] [PubMed]
- Miceli, M.H.; Kauffman, C.A. Isavconazole: A new broad-spectrum triazole antifungal agent. Clin. Infect. Dis. 2015, 61, 1558–1565. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Castelli, M.V.; Cuesta, I.; Monzon, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Activity of posaconazole and other antifungal agents against Mucorales strains identified by sequencing of internal transcribed spacers. Antimicrob. Agents Chemother. 2009, 53, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Pagano, L.; Cornely, O.A.; Busca, A.; Caira, M.; Cesaro, S.; Gasbarrino, C.; Girmenia, C.; Heinz, W.J.; Herbrecht, R.; Lass-Flörl, C.; et al. Combined antifungal approach for the treatment of invasive mucormycosis in patients with hematologic diseases: A report from the SEIFEM and FUNGISCOPE registries. Haematologica 2013, 98, e127–e129. [Google Scholar] [CrossRef] [PubMed]
- Caramalho, R.; Tyndall, J.D.A.; Monk, B.C.; Larentis, T.; Lass-Flörl, C.; Lackner, M. Intrinsic short-tailed azole resistance in mucormycetes s due to an evolutionary conserved aminoacid substitution of the lanosterol 14α-demethylase. Sci. Rep. 2017, 7, 15898. [Google Scholar] [CrossRef] [PubMed]
- Fryberg, M.; Oehlschlager, A.C.; Unrau, A.M. Biosynthesis of ergosterol in yeast. Evidence for multiple pathways. J. Am. Chem. Soc. 1973, 95, 5747–5757. [Google Scholar] [CrossRef] [PubMed]
- Osumi, T.; Taketani, S.; Katsuki, H.; Kuhara, T.; Matsumoto, I. Ergosterol biosynthesis in yeast: Pathways in the late stages and their variation under various conditions. J. Biochem. 1978, 83, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Parks, L.P. Physiological implications of sterol biosynthesis in yeast. Annu. Rev. Microbiol. 1995, 49, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Mellado, E.; Garcia-Effron, G.; Lopez, J.F.; Grimalt, O.J.; Cuenca-Estrella, J.M.; Rodriguez-Tudela, J.L. Ergosterol biosynthesis pathway in Aspergillus fumigatus. Steroids 2008, 73, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Binder, U.; Bracher, F.; Giera, M. Antifungal drug testing: Combining minimal inhibitory concentration testing with target identification by the use of gas chromatography—Mass spectrometry. Nat. Protoc. 2017, 12, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Weete, J.D.; Wise, M.L. Effects of triazoles on fungi. Exp. Mycol. 1987, 11, 214–222. [Google Scholar] [CrossRef]
- Alcazar-Fuoli, L.; Mellado, E. Ergosterol biosynthesis in Aspergillus fumigatus: Its relevance as an antifungal target and role in antifungal drug resistance. Front. Microbiol. 2013, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. A clinical isolate of Candida albicans with mutations in ERG11 (encoding sterol 14α-demethylase) and ERG5 (encoding C22-desturase) is cross resistant to azoles and amphotericin B. Antimicrob. Agents Chemother. 2010, 54, 3578–3583. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.; Müller, C.; Engelhardt, I.; Hiller, E.; Lemuth, K.; Eickhoff, H.; Wiesmüller, K.H.; Burger-Kentischer, A.; Bracher, F.; Rupp, S. An antifungal benzimidazole derivative inhibits ergosterol biosynthesis and reveals novel sterols. Antimicrob. Agents Chemother. 2015, 59, 6296–6307. [Google Scholar] [CrossRef] [PubMed]
- Colley, T.; Alanio, A.; Kelly, S.L.; Sehra, G.; Kizawa, Y.; Warilow, A.G.S.; Parker, J.E.; Kelly, D.E.; Kimura, G.; Anderson-Dring, L.; et al. In vitro and in vivo antifungal profile of a novel and long acting inhaled azole PC945, on Aspergillus fumigatus infection. Antimicrob. Agents Chemother. 2017, 61, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Georgopapadakou, N.H. Antifungals: Mechanism of action and resistance, established and novel drugs. Curr. Opin. Microbiol. 1998, 1, 547–557. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [PubMed]
- Lupetti, A.; Danesi, R.; Campa, M.; del Tacca, M.; Kelly, S. Molecular basis of resistance to azole antifungals. Trends Mol. Med. 2002, 8, 76–81. [Google Scholar] [CrossRef]
- McCorkindale, N.J.; Hutchinson, S.A.; Pursey, B.A.; Scott, W.T.; Wheeler, R. A comparison of the types of sterol found in species of the Saprolegniales and Leptomitales with those found in some other Phycomycetes. Phytochemistry 1969, 8, 861–867. [Google Scholar] [CrossRef]
- Gordon, P.A.; Stewart, P.R.; Clark-Walker, G.D. Fatty acids and sterol composition of Mucor genevensis in relation to dimorphism and anaerobic growth. J. Bacteriol. 1971, 107, 114–120. [Google Scholar] [PubMed]
- Safe, S. The effect of environment on the free and hydrosoluble sterols of Mucor rouxii. Biochim. Biophys. Acta 1973, 326, 471–475. [Google Scholar] [CrossRef]
- Weete, J.D.; Lawler, G.C.; Laseter, J.L. Total lipid and sterol components of Rhizopus arrhizus: Identification and metabolism. Arch. Biochem. Biophys. 1973, 155, 411–419. [Google Scholar] [CrossRef]
- Weete, J.D.; Gandhi, S.R. Sterols of the phylum Zygomycota: Pylogenetic implications. Lipids 1997, 32, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic distribution of fungal sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef] [PubMed]
- Dannaoui, E.; Persat, F.; Borel, E.; Piens, M.-A.; Picot, S. Sterol composition of itraconazole-resistant and itraconazole-susceptible isolates of Aspergillus fumigatus. Can. J. Microbiol. 2001, 47, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Gsaller, F.; Hortschansky, P.; Furukawa, T.; Carr, P.D.; Rash, B.; Capilla, J.; Müller, C.; Bracher, F.; Bowyer, P.; Haas, H.; et al. Sterol biosynthesis and azole tolerance is governed by the opposing actions of SrbA and the CCAAT binding complex. PloS Pathog. 2016, 12, e1005775. [Google Scholar]
- Misslinger, M.; Gsaller, F.; Hortschansky, P.; Müller, C.; Bracher, F.; Bromley, M.J.; Haas, H. The cytochrome b5 CybE is regulated by iron availability and is crucial for azole resistance in A. fumigatus. Metallomics 2017, 9, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Staudacher, V.; Krauss, J.; Giera, M.; Bracher, F. A convenient cellular assay for the identification of the molecular target of ergosterol biosynthesis inhibitors and quantification of their effects on total ergosterol biosynthesis. Steroids 2013, 78, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Binder, U.; Maurer, E.; Grimm, C.; Giera, M.; Bracher, F. Fungal sterol C22-desaturase is not an antimycotic target as shown by selective inhibitors and testing on clinical isolates. Steroids 2015, 101, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nemec, T.; Jernejec, K.; Cimerman, A. Sterols and fatty acids of different Aspergillus species. FEMS Microbiol. Lett. 1997, 149, 201–205. [Google Scholar] [CrossRef]
- Kelly, S.L.; Lamb, D.C.; Corran, A.J.; Baldwin, B.C.; Kelly, D.E. Mode of action and resistance to azole antifungals associated with the formation of 14α-methylergosta-8,24(28)-dien-3β,6α-diol. Biochem. Biohys. Res. Commun. 1995, 207, 910–915. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Relative [%] and Absolute [µg/mg] Amount of Sterols in Each Species | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. | IUPAC Name | Common Name | L. corymbifera | L. ramosa | M. circinelloides | Rh. pusillus | R. arrhizus | R. microsporus | A. fumigatus |
| 1 | ergosta-5,8,22-trien-3β-ol | Lichesterol | 0.2 0.02 (± 0.00) | 0.4 0.07 ± 0.02) | 0.3 0.07 (± 0.00) | 0.2 0.05 (± 0.00) | 0.8 0.07 (± 0.01 | 0.9 0.13 (± 0.02 | 1.2 0.12 (± 0.03) |
| 2 | ergosta-5,7,22-trien-3β-ol | Ergosterol | 57.5 6.45 (± 0.66) | 69.2 13.69 (± 3.76) | 65.8 12.93 (± 0.37) | 38.0 8.13 (± 0.33) | 76.3 7.07 (± 0.04) | 70.3 9.91 (± 1.85) | 95.0 9.29 (± 2.28) |
| 3 | ergosta-7,22-dien-3β-ol | 3.5 0.39 (± 0.04) | i.t.** | n.d.* | 1.9 0.41 (± 0.02) | 0.2 0.02 (± 0.01) | 0.2 0.03 (± 0.02) | n.d. | |
| 4 | ergosta-5,7,22,24(28)-tetraen-3β-ol | 0.4 0.04 (± 0.00) | 0.6 0.11 (± 0.00) | 0.5 0.10 (± 0.01) | 0.3 0.06 (± 0.00) | 1.0 0.09± 0.02) | 1.2 0.17 (± 0.03) | 0.7 0.06 (± 0.01) | |
| 5 | ergosta-7,22,24(28)-trien-3β-ol | 0.2 0.02 (± 0.01) | 0.2 0.04 (± 0.03) | 0.2 0.04 (± 0.00) | 0.2 0.03 (± 0.02) | 0.2 0.03 ± 0.02) | 0.2 0.03 (± 0.02) | 0.2 0.02 (± 0.00) | |
| 6 | ergosta-5,7,24(28)-trien-3β-ol | n.d. * | 7.4 1.50 (± 1.94) | 10.4 2.05 (± 0.37) | n.d. ** | 2.6 0.24 ± 0.22) | 3.3 0.46 (± 0.39) | 0.6 0.06 (± 0.01) | |
| 7 | ergosta-5,7-dien-3β-ol | 28.5 3.19 (± 0.36) | 12.3 2.48 (± 0.11) | 10.6 2.07 (± 0.26) | 37.2 9.96 (± 0.07) | 10.6 0.98 (± 0.19) | 12.9 1.82 (± 0.43) | 0.3 0.03 (± 0.00) | |
| 8 | ergosta-7,24(28)-dien-3β-ol | Episterol | 2.4 0.27 (± 0.04) | 1.9 0.38 (± 0.13) | 1.8 0.35 (± 0.01) | 1.4 0.29 (± 0.01) | 2.1 0.20 (± 0.13) | 2.4 0.34 (± 0.19) | 0.8 0.08 (± 0.01) |
| 9 | ergost-7-en-3β-ol | 3.7 0.41 (± 0.02) | 0.1 0.02 (± 0.03) | i.t. ** | 3.0 0.64 (± 0.02) | 0.3 0.03 (± 0.02) | 0.5 0.07 (± 0.03) | n.d. * | |
| 10 | 4,4,14-trimethylcholesta-8,24-dien-3β-ol | Lanosterol | 0.3 0.03 (± 0.00) | 1.1 0.21 (± 0.24) | 1.3 0.26 (± 0.04) | 0.7 0.07 (± 0.03) | 0.7 0.07 (± 0.03) | 1.0 0.14 (± 0.05) | 0.2 0.02 (± 0.00) |
| 11 | 4,4-dimethylcholesta-8,24-dien-3β-ol | T-MAS | 0.2 0.04 (± 0.00) | 0.6 0.12 (± 0.13) | 0.7 0.15 (± 0.01) | 0.6 0.13 (± 0.01) | 0.2 0.02 (± 0.00) | 0.2 0.03 (± 0.00) | 0.4 0.04 (± 0.00) |
| 12 | 4-methylergost-8-en-3β-ol | 0.4 0.05 (± 0.00) | 0.1 0.01 (± 0.02) | i.t. ** | 0.9 0.18 (± 0.00) | 0.2 0.02 (± 0.02) | 0.2 0.02 (± 0.02) | n.d. * | |
| 13 | 4,4,14-trimethylergosta-8,24(28)-dien-3β-ol | Eburicol | 2.0 0.22 (± 0.01) | 5.3 1.07 (± 1.33) | 7.2 1.41 (± 1.16) | 15.4 3.29 (± 0.18) | 3.7 0.34 (± 0.12) | 5.5 0.77 (± 0.15) | n.d. * |
| 14 | 4,4-dimethylergosta-8,24(28)-dien-3β-ol | 0.7 0.08 (± 0.01) | 0.9 0.17 (± 0.19) | 1.1 0.21 (± 0.01) | 0.7 0.15 (± 0.00) | 0.3 0.03 (± 0.01) | 0.3 0.05 (± 0.01) | 0.6 0.06 (± 0.00) | |
| total sterol content | 100 11.22 (± 1.16) | 100 20.17 (± 7.83) | 100 19.64 (± 0.45) | 100 21.39 (± 0.61) | 100 9.26 (± 0.66) | 100 14.09 (± 2.96) | 100 9.78 (± 2.36) | ||
| Compound | Relative [%] and Absolute [µg/mg] Amount of Sterols | |||||
|---|---|---|---|---|---|---|
| R. arrhizus | A. fumigatus | |||||
| No. | IUPAC Name | Common Name | POS Treated | Untreated | POS Treated | Untreated |
| 1 | ergosta-5,8,22-trien-3β-ol | Lichesterol | 0.7 0.09 (± 0.01) | 0.8 0.07(± 0.01) | 1.2 0.10 (± 0.00) | 1.2 0.12 (± 0.03) |
| 2 | ergosta-5,7,22-trien-3β-ol | Ergosterol | 58.5 7.52 (± 0.20) | 76.3 7.07(SD ± 0.04) | 84.1 7.41 (± 1.05) | 95.0 9.29 (± 2.28) |
| 3 | ergosta-7,22-dien-3β-ol | 0.1 0.01 (± 0.01) | 0.2 0.02 (± 0.01) | 0.1 0.01 (± 0.01) | n.d. * | |
| 4 | ergosta-5,7,22,24(28)-tetraen-3β-ol | 0.7 0.09 (± 0.02) | 1.0 0.09 (± 0.02) | 0.8 0.07 (± 0.01) | 0.7 0.06 (± 0.01) | |
| 5 | ergosta-7,22,24(28)-trien-3β-ol | 0.1 0.01 (± 0.00) | 0.2 0.03 (± 0.02) | 0.2 0.01 (± 0.01) | 0.2 0.02 (± 0.00) | |
| 6 | ergosta-5,7,24(28)-trien-3β-ol | 0.9 0.12 (± 0.12) | 2.6 0.24 (± 0.22) | 0.6 0.05 (± 0.04) | 0.6 0.06 (± 0.01) | |
| 7 | ergosta-5,7-dien-3β-ol | 5.1 0.66 (± 0.33) | 10.6 0.98 (± 0.19) | 1.9 0.17 (± 0.10) | 0.3 0.03 (± 0.00) | |
| 8 | ergosta-7,24(28)-dien-3β-ol | Episterol | 0.8 0.10 (± 0.00) | 2.1 0.20 (± 0.13) | 1.0 0.09 (± 0.02) | 0.8 0.08 (± 0.01) |
| 9 | ergost-7-en-3β-ol | 0.1 0.01(± 0.01) | 0.3 0.03 (± 0.02) | i.t. ** | n.d. * | |
| 10 | 4,4,14-trimethylcholesta-8,24-dien-3β-ol | Lanosterol | 2.5 0.33 (± 0.26) | 0.7 0.07 (± 0.03) | 1.3 0.12 (± 0.12) | 0.2 0.02 (± 0.00) |
| 11 | 4,4-dimethylcholesta-8,24-dien-3β-ol | T-MAS | i.t. ** | 0.2 0.02 (± 0.00) | 0.4 0.04 (± 0.00) | 0.4 0.04 (± 0.00) |
| 12 | 4-methylergost-8-en-3β-ol | 0.1 0.01(± 0.01) | 0.2 0.02 (± 0.02) | n.d. * | n.d. * | |
| 13 | 4,4,14-trimethylergosta-8,24(28)-dien-3β-ol | Eburicol | 29.1 3.74 (± 1.58) | 3.7 0.34 (± 0.12) | 6.9 0.61 (± 0.80) | n.d.* |
| 14 | 4,4-dimethylergosta-8,24(28)-dien-3β-ol | i.t. ** | 0.3 0.03 (± 0.01) | 0.7 0.06 (± 0.02) | 0.6 0.06 (SD ± 0.00) | |
| 15 | 14-methylergosta-8,24(28)-diene-3β,6α-diol | 0.7 0.09 (± 0.11) | n.d. * | n.d. * | n.d.* | |
| total sterol content | 100 12.86 (± 1.30) | 100 9.26 (± 0.66) | 100 8.81 (± 0.08) | 100 9.78 (± 2.36) | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, C.; Neugebauer, T.; Zill, P.; Lass-Flörl, C.; Bracher, F.; Binder, U. Sterol Composition of Clinically Relevant Mucorales and Changes Resulting from Posaconazole Treatment. Molecules 2018, 23, 1218. https://doi.org/10.3390/molecules23051218
Müller C, Neugebauer T, Zill P, Lass-Flörl C, Bracher F, Binder U. Sterol Composition of Clinically Relevant Mucorales and Changes Resulting from Posaconazole Treatment. Molecules. 2018; 23(5):1218. https://doi.org/10.3390/molecules23051218
Chicago/Turabian StyleMüller, Christoph, Thomas Neugebauer, Patrizia Zill, Cornelia Lass-Flörl, Franz Bracher, and Ulrike Binder. 2018. "Sterol Composition of Clinically Relevant Mucorales and Changes Resulting from Posaconazole Treatment" Molecules 23, no. 5: 1218. https://doi.org/10.3390/molecules23051218