Comparison of Strategies to Overcome Drug Resistance: Learning from Various Kingdoms

1
HO Bio Institute, Yushima-2, Bunkyo-ku, Tokyo 113-0034, Japan
2
Department of Biochemistry, Meiji Pharmaceutical University, Noshio-2, Kiyose, Tokyo 204-8588, Japan
Molecules 2018, 23(6), 1476; https://doi.org/10.3390/molecules23061476
Submission received: 4 May 2018
/
Revised: 13 June 2018
/
Accepted: 15 June 2018
/
Published: 18 June 2018
(This article belongs to the Section Natural Products Chemistry)
Abstract
:Drug resistance, especially antibiotic resistance, is a growing threat to human health. To overcome this problem, it is significant to know precisely the mechanisms of drug resistance and/or self-resistance in various kingdoms, from bacteria through plants to animals, once more. This review compares the molecular mechanisms of the resistance against phycotoxins, toxins from marine and terrestrial animals, plants and fungi, and antibiotics. The results reveal that each kingdom possesses the characteristic features. The main mechanisms in each kingdom are transporters/efflux pumps in phycotoxins, mutation and modification of targets and sequestration in marine and terrestrial animal toxins, ABC transporters and sequestration in plant toxins, transporters in fungal toxins, and various or mixed mechanisms in antibiotics. Antibiotic producers in particular make tremendous efforts for avoiding suicide, and are more flexible and adaptable to the changes of environments. With these features in mind, potential alternative strategies to overcome these resistance problems are discussed. This paper will provide clues for solving the issues of drug resistance.
1. Introduction
Antimicrobial agents, including antibiotics, once eliminated the serious infectious diseases almost completely from the Earth [1]. However, the emergence of multidrug resistant bacteria has revived bacterial pathogens once again, and has made the infectious diseases difficult-to-treat or untreatable now [2,3]. So, finding strategies for the rapidly increasing prevalence of antibiotic resistance is a major global challenge for the life science and public health sectors [4,5,6,7,8].
Living organisms produce a wide range of low molecular weight, natural organic compounds, including phycotoxins, marine toxins, toxins from terrestrial animals, phytotoxins, toxins from fungi, and antibiotics, and other kinds of bacterial toxins. These toxins have been produced and diversified evolutionally for interspecies arms races between offensive predators and defensive prey [9,10,11,12,13]. Here, predators and preys are not necessarily higher organisms but microorganisms. The producers of these toxins need to have strategies to prevent themselves from suicide [14]. These protective strategies can oscillate and evolve, depending on natural environments and the different kingdoms of the producers [15,16,17,18,19,20]. On the other hand, the nature and chemistry of the toxins themselves evolves in both offensive and defensive contexts [21,22,23,24]. It is interesting, therefore, to compare the protective and/or defensive strategies of bacteria through fungi, algae, plants, and animals. In addition, the clarification in the differences of these complex strategies may provide clues to solve the growing problems of antibiotic resistance.
2. Phycotoxins
Phycotoxins are toxic secondary metabolites that are produced by prokaryotic and eukaryotic algae or seaweeds [25,26,27]. These are molecules of small to medium (300~3500 Da) mass belonging to diverse groups of chemical compounds. Most phycotoxins are produced by flagellates, especially dinoflagellates. However, they are also produced by diatoms, haptophytes (prymnesiophytes) [28], raphidophytes [29], and cyanobacteria [30]. These phycotoxins are known to accumulate in seafood as a result of the marine food chain. Food poisoning in humans occurs because of the ingestion of seafood that is contaminated with these toxins. Different phycotoxins cause distinct poisoning events. These poisoning events are grouped into six classes, that is, neurotoxic shellfish poisoning (NSP), diarrheic shellfish poisoning (DSP), azaspiracid poisoning (AZP), ciguatera fish poisoning (CFP), amnesic shellfish poisoning (ASP), and paralytic shellfish poisoning (PSP) [31]. Four of the six poisonings are induced by dinoflagellate-derived polyketide toxins [32]. The two others are ASP and PSP. ASP is caused by domoic acid, a kainic acid analog that is produced by diatoms in the Pseudo-nitschia genus, and PSP is caused by the saxitoxins, a group of cyclic tetrahydropurine compounds that are produced by cyanobacteria, such as Anabaena circinalis, Aphanizomenon sp., and Nostocales sp. as well as by dinoflagellates, such as Alexandrium catenella, Gymnodinium catenatum, and Pyrodinium bahamense [33,34,35].
The majority of dinoflagellate toxins are polyketide in origin. Thus, polyketide compounds are discussed at first, focusing mainly on their self-resistance to phycotoxins. Polyketides are biosynthesized via the sequential condensations of small carboxylic acid subunits with an acyl starter in a fashion that is reminiscent of fatty acid biosynthesis. Both polyketide synthases (PKS) and fatty acid synthases (FAS) possess a similar set of functional domains, namely, ketoacyl synthase (KS), acyl transferase (AT), ketoacyl reductase (KR), dehydratase (DH), enoyl reductase (ER), acyl carrier protein (ACP), and thioesterase (TE). PKS are traditionally classified into three types, namely, type I, type II, and type III [36,37]. Type I PKS are large multifunctional proteins that combine several domains in one protein. Two subclasses are known for Type I PKS. Fungal iterative Type I PKS use the same set of catalytic domains on one protein several times for chain extension, analogously to vertebrate FAS. In contrast, modular Type I PKS function in a conveyor belt-like manner, in that the different catalytic domains are organized in modules comprising all of the required enzymatic functions. Each module is used only once during the polyketide assembly. Based on their size, functionalities, and complex structures, it is predicted that the dinoflagellate-derived polyketides are biosynthesized by Type I modular PKS. However, recent genome sequencing and transcriptome analysis, combined with blast analysis, indicate that monofunctional Type I PKS are present in brevetoxin-producing dinoflagellates Karenia brevis [38,39], Alexandrium ostenfeldii [40], and Heterocapsa triqueta [41]. In any case, these Type I PKS genes are distributed patchily in phytoplankton; they are present in Cryptosporidium and Emiliania but not in Thalassiosira nor in Cyanidioschyzon [42].
The mechanisms of polyketide biosynthesis in phycotoxins have been investigated both in eukaryotic (mainly dinoflagellates) and in prokaryotic organisms (cyanobacteria). However, only a few PKS have been analyzed at a molecular level in eukaryotic organisms and major research has been performed in prokaryotic organisms, because in eukaryotic organisms, their genome sizes range from 15 Gbp to 150 Gbp [43]; chromosome copy numbers vary markedly from 4 to 220 [44]; and genomes are very complex as a result of gene duplication, lateral gene transfer, endosymbiotic gene transfer events [45], and so on. Cylindrospermopsin is produced by cyanobacterial species such as Cylindrospermopsis raciborskii, Aphanizomenon ovalisporum, Umezakia natans, Raphidiopsis curvata, and Anabaena bergii. It has hepatoxic and neurotoxic effects and is a potential carcinogen. Its toxicity is due to the inhibition of glutathione and protein synthesis, as well as the inhibition of cytochrome P450. The toxin is a polyketide-derived alkaloid with a central functional guanidine moiety and a hydroxymethyluracil. Feeding experiments with isotope-labeled precursors have shown that guanidinoacetate is the starter unit for cylindrospermopsin biosynthesis, and successive additions of five intact acetate units onto guanidinoacetate yield the carbon backbone of cylindrospermopsin [46]. Afterward, cylindrospermopsin biosynthetic gene clusters were cloned from three producing species, Cylindrospermopsis raciborskii AWT205 (43 kb), Aphanizomenon sp. strain 10E6 (57 kb), and Oscillatoria sp. Strain PCC 6506 (45 kb) [47,48,49] (GenBank accession Nos. EU140798, GQ385961, and FJ418586; GB No. hereafter). A comparison of these gene clusters indicates that they are homologous and evolutionarily related, and are diverged from a common ancestor, but a substantial shuffling occurred in these organisms. It is interesting that the multidrug exporter gene, cyrK (GB No. ABX60156), exists within the gene clusters, indicating that it functions as a strategy of self-resistance against cylindrospermopsin.
Jamaicamide A is produced by filamentous cyanobacterium, Lyngbya majuscula. It is a highly functionalized lipopeptide and shows sodium channel blocking activity. Feeding experiments with labeled precursor have mapped out series of acetate and amino acid residues on the structure. The major metabolic pathway employs two modular biosynthetic systems, nonribosomal peptide synthetases (NRPS), which are responsible for assembling amino acids; and polyketide synthases (PKS), for linking together acetate as the primary building block. Edwards et al. cloned the jamaicamide-producing gene cluster as a 58 kb DNA fragment composed of 17 open reading frames [50] (GB No. AY522504). They show exact collinearity with their expected utilization, form the operon jamABCDEFGHIJKLMNOP, and are transcribed in the same direction, except for the last gene, jamQ. The last ORF (Open reading frame) JamQ, which is thought to be involved in the cyclization of the pyrrolinone ring of the molecule, is transcribed in the reverse direction. The gene cluster is preceded by a long untranslated leader region (at least 844 bp), but its exact function is not clear yet [50,51]. No resistance-related gene has been found within the gene cluster.
Hectochlorin was also isolated from Lyngbya majuscula. It is a cyclic lipopeptide and exhibits antifungal activity against Candida albicans and antiproliferative activity because of the stimulation of actin assembly [52]. The structure of hectochlorin indicates that it is derived from a mixed PKS/NRPS pathway. The cloning of the biosynthetic gene cluster supports this suggestion [53]. It consists of eight open reading frames spanning 38 kb (GB No. AY974560). All of the eight genes are transcribed in the same direction. However, no resistance-related gene has been found within the gene cluster.
Curacin A was also obtained from Lyngbya majuscula. It has a unique structure containing the sequential positioning of a thiazoline and cyclopropyl ring that have been biosynthesized through the PKS/NRPS pathways. It is a cancer cell toxin as a result of the blocking of the cell cycle progression, by interacting with the colchicine binding site on tubulin and inhibiting microtubule polymerization. The biosynthetic gene cluster was cloned as a 64 kb DNA fragment and the metabolic system shows a very high level of collinearity between the genes in the cluster and the predicted biochemical steps [54,55] (GB Nos. AY652953 and HQ696500). All of the 14 genes are transcribed in the same direction. However, no resistance-related gene has been found within the cluster. As some ABC type transporter genes are found in the Lyngbya majuscula 3L genome [55], these transporters may be involved in the excretion of the toxins, similar to the case of cylindrospermopsin (GB No. GL890825). Amphidinolides and amphidinols from the genus Amphidinium dinoflagellates have similar structures to curacins [32,56].
Apratoxin A was isolated from Lyngbya (Moorea) bouillonii and has a structure that is composed of a polyketide section that is fused with a modified pentapetide to form a cyclic lipopeptide. Apratoxin A inhibits signal transducer and activator of transcription (STAT) 3 phosphorylation in various cell types, and induces pronounced G1 cell cycle arrest and apoptosis [57]. The cloned 58 kb biosynthetic gene cluster is composed of 12 open reading frames and has a Type I modular mixed PKS-NRPS organization [58]. No resistance-related gene has been found within the gene cluster. However, adjacent to the polyketide synthase genes, many ABC transporter genes are present, indicating that these ABC transporters may excrete the toxin from the cells for self-resistance (For example: GB Nos. OLT63032 and WP_075905632).
Lyngbyatoxin is an indole alkaloid first identified from a Moorea producens bloom having a tumor promoter activity due to the activation of protein kinase C. The 11.3 kb biosynthetic gene cluster contains four open reading frames encoding a bimodular nonribosomal peptide synthetase, cytochrome P450 monooxygenase, and a protein that is related to an oxidase/reductase [59] (GB No. AY588942). All of these fragments are transcribed in the same direction. The entire lyngbyatoxin gene cluster was attempted, in order to express heterologously in Streptomyces coelicolor A3(2) and Anabaena sp. strain PCC7120 [60,61]. Although the expression of the entire gene clusters were unsuccessful, cytochrome P45, monooxygenase LtxB, and the reverse prenyltransferase LtxC genes were accomplished, so as to express in S. coelicolor A3(2). In this gene cluster, no transporter nor resistance-related gene has been identified.
Teleocidin B, a protein kinase C activator that is produced by Streptomyces blastmyceticus NBRC 12747, is an analogue of lyngbyatoxin. The biosynthetic gene cluster of teleocidin B, composed of 23.2 kb of DNA fragments, was cloned and sequenced [62] (GB No. AB937114). It contains 15 open reading frames, including tleA for a nonribosomal peptide synthetase, tleB for a P-450 monooxygenase, tleC for an aromatic prenyltransferase, and three genes for ABC transporter. Interestingly, Streptomyces lividans TK21, containing the cluster, produces lyngbyatoxin but not teleocidin B. The essential gene for the biosynthesis of teleocidin B, tleD, is located outside of the tle cluster (GB No. AB937726). Furthermore, three ABC transporter genes are present, adjacent to the tleABC genes. These ABC transporters may excrete teleocidin B as well as lyngbyatoxin from the cells.
Hepatotoxic microcystins are a family of heptapeptides that are produced by bloom-forming freshwater cyanobacteria, such as Mycrocystis, Planktothrix, and Anabaena [63]. The microcystins contain a number of unusual amino acid residues, including 3-amino-9-methoxy-2,6,8-trimethyl-10-phenyl-4,6-decadieniic acid, 3-methylaspartic acid, and N-methyl-dehydroalanine. The closely related pentapeptide nodularin is found frequently in the cyanobacteria of the species of Nodularia spumigena [64]. Microcystins and nodularin inhibit eukaryotic protein phosphatases of Type 1 and Type 2a, and are able to penetrate the liver cells via active transport. Biochemical and genetic studies, including feeding experiments with labelled precursors, suggest that the microcystins are biosynthesized by a mixed PKS/NRPS pathway. The biosynthetic gene cluster for microcystin spanning 55 kb was cloned (GB No. AF183408). It is composed of 10 bidirectionally transcribed open reading frames that are arranged in two putative operons, mcyA-mcyC and mcyD-mcyJ. The mcyD-mcyJ gene cluster contains seven open reading frames, all of which are transcribed in the opposite direction to the putative mcyABC operon. Among them, the 1617 bp open reading frame mcyH encodes a putative 37,000 Da transmembrane protein, belonging to the ABC transporter. Although no obvious function can be assigned to McyH, it is possible to speculate that McyH may play a role in the excretion of the toxin [65,66]. This speculation is supported by phylogenetic analysis [67]. Additionally, it was reported that cyanobacterial phosphoprotein phosphatase (PPP) family protein phosphatases, such as PP1-cyano1 and PP1-cyano2 from Microcystis aeruginosa PCC 7820, are resistant to microcystin-LR [68]. Thus, the targets are also resistant to microcystin-LR in the producer organism.
The microcystin-related cyclic pentapeptide, nodularin, is produced by Nodularia spumigena. The 48 kb gene cluster of nodularin consists of nine open reading frames, ndaA to ndaI (GB No. AY210783). Similar to the case of microcystin, they are transcribed from a bidirectional regulatory promoter region and encode the nonribosomal peptide synthetase modules, polyketide synthase modules, and tailoring enzymes. NdaI consisting of 601 amino acid residues is an ABC transporter. The comparison of the gene clusters for microcystin and nodularin and of the condensation domains of NdaA and McyA/McyB revealed that extensive gene arrangements occurred between the two clusters, and that the gene cluster of nodularin evolved from a microcystin synthetase progenitor [64].
Aplysiatoxin, isolated from sea hare Stylocheilus longicauda and cyanobacteria, such as Lyngbya majuscula, Schizothrix calcicola, and Trichodesmium erythraeum, is composed of a 14-membered bis-macrocyclic ring and a side chain containing an aromatic ring. Like lyngbyatoxin, aplysiatoxin induces dermatitis through the activation of protein kinase C. It is also a tumor promoter. Nhatrangin, possessing many structural similarities to aplysiatoxin, is suggested to be putative starter units for the aplysiatoxin biosynthetic pathway [69]. However, the biosynthetic gene clusters of these metabolites have not been cloned.
Saxitoxins are the most renowned molecules, known as the paralytic shellfish toxins. However, they are originally biosynthesized by cyanobacteria, such as Anabaena circinales, Aphanizomenon grazile, Cylindrospermopsis raciborskii, and Lyngbya wollei, and by eukaryotic dinoflagellates, such as Alexandrium, Gymnodinium, and Pyrodinium [34,70], and, subsequently, are transferred to various invertebrate and vertebrate species through the aquatic freshwater and marine food chains. Saxitoxins are a group of carbamate alkaloid toxins consisting of a tetrahydropurine group and two guanidinium moieties. Intoxication with saxitoxins in humans may result in the severe and occasionally fatal illness known as paralytic shellfish poisoning. This illness is caused by the binding of saxitoxins to the α-subunit of voltage-gated Na+ channels (Nav). This is mediated by the interaction between the positively charged guanidinium groups of saxitoxins, with the negatively charged carboxyl groups at site 1 of the Na+ channel, thereby obstructing the entry of sodium ions through the pore and blocking nerve and muscle action potentials [71]. Interestingly, softshell clams (Mya arenaria) from areas that are exposed to red tides are more resistant to saxitoxins and accumulate saxitoxins at greater rates than the sensitive clams from unexposed areas. The resistance in the clams to saxitoxins is caused by a natural mutation of only one amino acid residue, which causes a 1000-fold decrease in affinity at the saxitoxin-binding site in the sodium channel pore. Thus, paralytic shellfish toxins like saxitoxins may act as natural selection agents, leading to a greater toxin resistance in the clam populations and an increased risk of paralytic shellfish poisoning to humans [72,73].
Saxitoxins are the most renowned molecules, known as the paralytic shellfish toxins. However, they are originally biosynthesized by cyanobacteria, such as Anabaena circinales, Aphanizomenon grazile, Cylindrospermopsis raciborskii, and Lyngbya wollei, and by eukaryotic dinoflagellates, such as Alexandrium, Gymnodinium, and Pyrodinium [34,70], and, subsequently, are transferred to various invertebrate and vertebrate species through the aquatic freshwater and marine food chains. Saxitoxins are a group of carbamate alkaloid toxins consisting of a tetrahydropurine group and two guanidinium moieties. Intoxication with saxitoxins in humans may result in the severe and occasionally fatal illness known as paralytic shellfish poisoning. This illness is caused by the binding of saxitoxins to the α-subunit of voltage-gated Na+ channels (Nav). This is mediated by the interaction between the positively charged guanidinium groups of saxitoxins, with the negatively charged carboxyl groups at site 1 of the Na+ channel, thereby obstructing the entry of sodium ions through the pore and blocking nerve and muscle action potentials [71]. Interestingly, softshell clams (Mya arenaria) from areas that are exposed to red tides are more resistant to saxitoxins and accumulate saxitoxins at greater rates than the sensitive clams from unexposed areas. The resistance in the clams to saxitoxins is caused by a natural mutation of only one amino acid residue, which causes a 1000-fold decrease in affinity at the saxitoxin-binding site in the sodium channel pore. Thus, paralytic shellfish toxins like saxitoxins may act as natural selection agents, leading to a greater toxin resistance in the clam populations and an increased risk of paralytic shellfish poisoning to humans [72,73].
The biosynthetic gene clusters of saxitoxins were cloned from cyanobacteria [34,74,75,76,77], and dinoflagellates [78]. The comparative analysis of the saxitoxin gene clusters in five species of cyanobacteria, that is, Cylindrospermopsis raciborskii T3 (GB No. DQ787200), Anabaena circinalis AWQC131C (GB No. DQ787201), Aphanizomenon sp. NH5 (GB No. EU603710), Lyngbya wollei (GB No. EU603711), and Raphidiopsis brookii D9 (GB No. ACYB00000000), indicates that the extensive shuffling of the genes that are involved in the biosynthesis of saxitoxins occurred among these species. Saxitoxins may be excreted through SxtF and SxtM, two multidrug and toxic compound extrusion (MATE) family transporters. Intriguingly, sxtM is present in all five sxt gene clusters, but sxtF is only present in C. raciborskii T3 and R. brookii D9. The two domains thata are involved in Na+ and drug recognition from NorM proteins (MATE family proteins [79]) of Vibrio parahaemolyticus and V. cholera are present in SxtF and SxtM [79]. In L. wollei, three sxtM genes are present. Therefore, these exporters may function in the resistant mechanisms of saxitoxin-producing bacteria and/or Nav themselves in the toxin producers that are resistant to saxitoxins or the mutations of the Nav result in the resistance. It is known that the structures of Nav is completely different in bacteria from those in eukaryotic organisms [80,81,82]. In two saxitoxin-producing dinoflagellate strains, Alexandrium fundyense CCMP1719 and A. minutum CCMP113, the analysis of sxtA, the starting gene of saxitoxin synthesis, showed that the dinoflagellate transcripts of sxtA have the same domain structure as the cyanobacterial sxtA genes, but the dinoflagellate transcripts are monocistronic, have a higher GC content, and contain typical dinoflagellate spliced-leader sequences and eukaryotic polyA-tails. Interestingly, in these eukaryotic dinoflagellate strains, two transporter genes sxtF and sxtM, were conserved [78].
The sxtA encodes a polyketide synthase in saxitoxin-producing cyanobacterium Anabaena circinalis. It is interesting evolutionally that SxtA is comprised of two distinct regions, namely, the N-terminal region of about 800 amino acids and the C-terminal region of about 390 amino acids; the former contains an acyl-CoA N-acyltransferase and a phosphopantetheine binding domain, which are homologous to those from proteobacteria, such as Myxococcus xanthus and Burkholderia ambifaria; and the latter shares a significant identity to a class I and II aminotransferase from actinobacteria, such as Frankia alni and Catenulispora acidiphila. In dinoflagellate Alexandrium tamarense, SxtA is split into two proteins corresponding to the N-terminal portion containing the methyltransferase and acyl carrier protein domains, and a C-terminal portion with the aminotransferase domain. The evolutional relationships of the saxitoxin biosynthetic genes in cyanobacteria and dinoflagellates were also analyzed [83,84,85,86].
Anatoxin A is a neurotoxic alkaloid and an agonist of the nicotinic acetylcholine receptor. Anatoxin A induces a neuromuscular blockade, resulting from muscle membrane depolarization and desensitization; impairs blood pressure, heart rate, and gas exchange causing hypoxia, muscle spasm, paralysis, and respiratory arrest; and finally death [87]. The gene cluster responsible for the biosynthesis of anatoxin A was identified in Oscillatoria sp., Anabaena flos-aquae 37, and Cylindrospermum stagnale PCC7417 [88,89,90]. The gene clusters from Oscillatoria sp. PCC6506 and Oscillatoria sp. PCC 6407 are identical, and those from Anabaena flos-aquae 37 and Cylindrospermum stagnale PCC7417 are similar to that of Oscillatoria sp. PCC6506, but they are slightly rearranged. The clusters contain three polyketide synthase genes, one acyl carrier protein gene, and one transporter gene. On the basis of the clusters, the biosynthetic route of anatoxin A was proposed [89]. The transporter AnaI may be responsible for the excretion of anatoxin A from the cells.
The hapalindole-type family of natural products is a group of lipophilic indole alkaloids that are produced by members of the cyanobacterial species of the order Stigonematales. This family includes hapalindoles, fisherindoles, ambiguines, and welwitindolinones [91]. These alkaloids show insecticidal, fungicidal, phytotoxic, and antialgal properties. Welwitindolinone A isonitrile shows antibacterial, antifungal, and antimycobacterial activities, and hapalindole A, fisherindole L, and N-methyl-welwitindolinone C isothiocyanate display cytotoxic activity against various cancer cells [92]. Interestingly, N-methyl-welwitindolinone C isothiocyanate attenuates the resistance of human breast carcinoma MCF-7/ADR cells to anticancer drugs, including vinblastine, taxol, actinomycin D, daunomycin, and colchicine, without affecting the cytotoxicity of cisplatin [93].
The biosynthetic gene clusters of the hapalindole-type alkaloids were cloned from the cyanobacterial strains Fischerella sp. ATCC 43239, Fischerella sp. PCC 9339, Fischerella ambigua UTEX 1903, Hapalosiphon welwitschii UH IC-52-3, Hapalosiphon welwitschii UTEX B1830, and Westiella intricate UH HT-29-1 [94,95,96]. There are three drug efflux pump or ABC transporter genes in the hapalindole gene cluster of Fischerella sp. PCC 9339 [IMG Gene IDs: 2517064622, 2517064623 and 2517064634], in the fisherindole gene cluster of Fischerella muscicola UTEX 1829 (GB Nos. APZ79543, APZ79544 and APZ79545), and in the ambiguine gene cluster of Fischerella ambigua UTEX 1903 (GB Nos. KJ742065 and KF664586). In the welwitindolinone biosynthetic gene cluster, on the other hand, there is one multidrug transporter of 105 amino acid residues of EmrE family in Westiella intricata UH HT-29-1 (GB No. AIH14815) and in Hapalosiphon welwitschii UH IC-52-3 (GB No. AIH14769). These transporters and/or efflux pumps may play an important role in the exclusion of hapalindole-type alkaloids.
Ciguatera fish poisoning is a food-borne disease that is endemic to tropical and subtropical coral reef regions of the world. However, as a result of the recent global warming, international trade, and increased nutrient loading, ciguatera is now emerging as a significant issue in Asia, America, and Europe [97]. The ciguatera fish poisoning is caused by the consumption of fish that are contaminated with ciguatoxins. Ciguatoxins are produced by benthic dinoflagellates of the genus Gambierdiscus and are concentrated in commonly consumed fish in the tropical and subtropical regions of the world, through the marine food chain. They are heat-stable, lipophilic polycyclic ethers of complex structures, and their molecular weights are 1000~1500 Da. The pharmacology of ciguatoxins is characterized by their ability to cause the persistent activation of Nav, to increase neuronal excitability and neurotransmitter release, and to cause cell swelling, leading to a complex array of gastrointestinal, neurological, and cardiovascular symptoms [98,99].
Remarkable structural similarities between polyether ladder toxins, like ciguatoxins, brevetoxins, maitotoxin, yessotoxin, okadaic acid, and gambierol, which are derived from the marine eukaryotes dinoflagellates, and monensin, a polyether-type antibiotic that is isolated from Streptomyces cinnamonensis, suggest that these toxins are biosynthesized through the polyketide route in a manner that is analogous to that of monensin assembly [31,32,100,101,102]. Monensin is shown to be biosynthesized by the modular type I PKS genes [103,104]. This is confirmed by the isotope incorporation experiments. However, the detailed biosynthetic mechanisms of ciguatoxins have not been explored at the genetic level, although similar biosynthetic pathways may also be employed in dinoflagellates. Through this connection, Monroe and Van Dolah [38] identified eight polyketide synthase transcripts in brevetoxin-producing Karenia brevis, by a high throughput cDNA library screening. Although there is no direct linking of these transcripts to brevetoxin biosynthesis, some transcripts contain polyadenylation, 3′-untranslated regions (UTRs), and an identical dinoflagellate-specific spliced leader domains at the 5′ end of PKS synthase transcripts. In addition, Kohli et al. [105] reported two gene clusters that were unique to maitotoxin-producing dinoflagellate species Gambierdiscus australes and G. belizeanus, suggesting that these clusters may be associated with maitotoxin biosynthesis. However, no transporter-related gene has been described. Other polyether ladder toxins are palytoxin [106] from dinoflagellates, soft corals, and cyanobacteria; and ostreocin from dinoflagellate Ostreopsis siamensis [107]. Predators such as a starfish (Acanthaster planci) and fish (Chaetodon species) feed on the Palythoa colonies and accumulate high toxin concentrations in their organs in its active form. The predators can tolerate high toxin concentrations by sequestration [108]. However, the biosynthetic genes have not been cloned. Okadaic acid, a cytotoxic polyether, is biosynthesized by marine dinoflagellates of the genus Prorocentrum and is a causative toxin of diarrhetic shellfish poisoning. It is an inhibitor of the eukaryotic serine/threonine protein phosphatase Type 1 and 2a, and is a promotor of tumors [100]. Interestingly, non-toxic sulfated diesters of okadaic acid and dinophysis toxin DTX-1, a derivative of okadaic acid, are initially biosynthesized in the dinoflagellate cells, indicating that these sulfated diesters make the producer resistant to okadaic acid [109].
Domoic acid is a neurotoxin and is biosynthesized by the marine diatom Pseudo-nitzschia australis, and related species [110]. The toxin targets ionotropic glutamate receptors that are present in various vital organs, inducing memory impairment, coma, recurrent seizures, and epilepsy. Kainic acid isolated from the red alga Digenia simplex and acromelic acid derived from the toxic fungus Paralepistopsis acromelalga are analogues of domoic acid [111,112,113]. To date, the domoic acid biosynthetic genes and the biosynthetic reactions have not been described. Examining the labeling patterns of domoic acid tht is produced in Pseudo-nitzschia cultures, it was proposed that domoic acid arises from the condensation of the C10 isoprenoid with glutamic acid, an activated C5 product of the TCA cycle [114,115]. In addition, Boissonneault et al. [116] identified some genes that were up-regulated under domoic acid-producing conditions, using microarray and RT-qPCR methods. These include a cycloisomerase, an SLC6 transporter [117], phosphoenolpyruvate carboxykinase, glutamate dehydrogenase, a small heat shock protein, and an aldo-keto reductase. Interestingly, the cycloisomerase, the SLC6 transporter, and the aldo-keto reductase genes had a statistically significant increase in accord with the increase in the domoic acid production. Thus, the SLC6 transporter may play an important role in the movement of domoic acid, into or out of cells.
Cyanobactins are defined as ribosomally synthesized peptides with post-translational modifications, which are produced by cyanobacteria [118,119]. Previously, they were thought to be biosynthesized in the tunicate Lissoclinum patella. It is now demonstrated that the cyanobacterium, Prochloron, a symbiont of the tunicate, is in fact responsible for the production of cyanobactins, through a post-ribosomal peptide synthesis pathway.
Patellamides are members of cyanobactin-group compounds. They are cytotoxic cyclic peptides and have reverse multidrug resistance in human cancer cells [120,121,122]. In 2005, Schmidt et al. cloned a 11 kb DNA fragment comprising patA-patG genes, which are responsible for the biosynthesis of patellamide A and patellamide C [123] (GB No. AY986476). The patE gene encodes a patellamide A and C precursor peptide of 71 amino acid residues, the first 37 of which serve as a leader sequence for processing. Of the remaining 34 amino acid residues, 16 amino acids constitute directly the patellamide A and patellamide C sequences, whereas the remaining 18 amino acids make up the motifs directing the cyclization of patellamides. Other gene products, such as PatA, PatD, and PatG, may be involved in the post-translational modification, leading to the biosynthesis of patellamide A and patellamide C [124,125]. ABC transporters are found in these gene clusters, indicating that these transporters may function as an excretion of these toxic substances from the cells [118]. Similar type of gene clusters are found in biosyntheses of microcins that are produced by Gram-negative bacteria [126], bacteriocins in Gram-positive bacteria [127], microviridin in cyanobacteria [128], and goadsporin in Streptomyces [129].
Microcins are gene-encoded antimicrobial peptides that are produced by Gram-negative bacteria, especially Enterobacteria [130]. They belong to a large family of bacteriocins and are involved in microbial competition. Recently, the complete genome sequence of a microcin B-producing Pseudomonas antarctica, PAMC 27494, was determined [131] (GB No. CP015600). The microcin B precursor that is encoded by mcbA is post-translationally processed to the mature form by McbBCD. The mcbE and mcbF genes encode the microcin ABC transporter system (GB Nos. ANF87043 and ANF87042), indicating that the processed microcin B is exported through this system outside of the cells. Another microcin-group antibiotic microcin C7 acts as a bactericide by inhibiting the aspartyl-tRNA synthetase and stalling the protein translation machinery. The biosynthetic gene cluster for microcin C7 on a plasmid was cloned and sequenced [132] (GB No. X57583). The cloned biosynthetic gene cluster consists of six open reading frames, namely, mccA, mccB, mccC, mccD, mccE, and mccF. The 21 bp mccA gene encodes the heptapeptide precursor, and mccC and mccE encode an efflux pump and acetyltransferase, respectively. Thus, at least two proteins, MccC and MccE, are implicated in the self-resistance of the producing strains to microcin C. Furthermore, MccF also involves in self-immunity [133,134].
The biosynthetic gene cluster of goadsporin was cloned from Streptomyces sp. TP-A0584 [129] (GB No. AB205012). The cluster contains a structural gene, godA, and nine god genes that are implicated in post-translation modification, immunity, and transcriptional regulation. GodB and GodC show a sequence similarity to the members of the ABC transporter family and may be responsible for the translocation of goadsporin to the cell membrane, and the excretion of goadsporin to outside of the producing cells [135]. Table 1 shows the resistance-related genes in the biosynthetic gene clusters of phycotoxins and related compounds. Summarizing these results, it is apparent that transporters, exporters, and efflux pumps play a major role in the self-resistance against phycotoxins in the producer organisms. Although the modification of toxins is also observed, like okadaic acid, it is only a rare case.
3. Marine Toxins
Electrical signaling across lipid membranes is essential for communication within and between cells. Ion-channels can pass the rapid and selective movements of one or several species of ions across the cell membrane. Voltage-gated ion channels are activated by changes in the local membrane potential. Voltage-gated sodium channels (Nav) play an essential role in the initiation and propagation of action potentials in neurons and other electrically excitable cells, such as myocytes and endocrine cells [71,136,137,138]. The Nav of human and mouse consist of α-subunit of 260 kDa and β-subunit of 30~40 kDa. Among them, the α-subunit is sufficient for functional Nav. The α-subunits of Nav are encoded by 10 genes, which are expressed in different excitable tissues [139]. Nav1.1, Nav1.2, Nav1.3, and Nav1.6 are the primary Nav in the central nervous system; Nav1.7, Nav1.8, and Nav1.9 are the primary Nav in the peripheral nervous system; Nav1.4 is the primary Nav in skeletal muscle; and Nav1.5 is the primary Nav in heart. The 10th sodium channel protein is not voltage-gated. Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.6, and Nav1.7 are tetrodotoxin-sensitive, and their IC50 are less than 10 nM. Nav1.5, Nav1.8, and Nav1.9 are tetrodotoxin-insensitive, and their IC50 are 1~10 μM. The α-subunits are large, single-chain polypeptides that are organized in four homologous domains, designated DI to DIV. Each domain consists of six trans-membrane helical segments, named S1 to S6. Segments S1 to S4 from each domain form the voltage-sensing domain (VSD). The four voltage-sensing domains are arranged around a central aqueous channel that is formed by the pore domain (PD). The pore domain (PD) includes the selectivity filter (SF). The selectivity filter (SF) is composed of aspartate (D) in DI, glutamate (E) in DII, lysine (K) in DIII, and alanine (A) in DIV (DEKA). The ring playing an important role in Na+ permeation is composed of two glutamates in DI and DII and two aspartate residues in DIII and DIV (EEDD). These amino acid residues are located just three residues downstream from those in the DEKA ring (Figure 1).
Tetrodotoxin is a deadly neurotoxin that selectively blocks Nav. Although tetrodotoxin is popularly known in Japan as the toxin in pufferfish, it is present in a diverse group of animals, including gobies, newts, frogs, horseshoe crabs, blur-ringed octopus, starfish, and red alga, dinoflagellates, and bacteria [140,141,142]. A general hypothesis is that a symbiotic or commensal bacterium living within these organisms is responsible for tetrodotoxin production. This hypothesis is supported by the fact that when pufferfish were fed a tetrodotoxin-free diet in an environment, they became nontoxic [143]. Interestingly, a comparison of the protein sequences of the skeletal muscle Nav shows that the tyrosine residue (Y407) in the pore loop of DI is substituted by the non-aromatic amino acid residue asparagine (N) or cysteine (C) in the tetrodotoxin-resistant fugu and Tetraodon channels (Figure 1). Furthermore, in the tetrodotoxin-insensitive human Nav1.5 channel from the heart muscle, it is replaced by cysteine (C). Some garter snake populations from different geographical locations are resistant to tetrodotoxin, however they conserve the aromatic amino acid residue (Y) at position 407. Instead, substitutions of several amino acids in the pore loop of DIV are responsible for tetrodotoxin resistance. Thus, tetrodotoxin attains a defensive role that protects the prey species from predation. However, some predators, like snakes, prey on tetrodotoxin-bearing animals, such as newts [10,11,144,145,146,147,148]. As described above, the saxitoxins bind to the same amino acid residues in the pore loop region on Nav as the tetrodotoxin. Neuronal Nav from the saxitoxin-resistant softshell clams (Mya arenaria) have the aromatic amino acid residue at position 407 intact, but glutamate (E) at position 764 in the pore loop of DII is substituted by aspartate (D) [72,73,149].
Tetrodotoxin is known to be biosynthesized by various bacteria, including actinobacteria, bacteroides, firmicutes, and proteobacteria [140,141]. However, its biosynthetic mechanism has not been clarified. Therefore, the self-resistance mechanism in these microorganisms remains to be defined. Tarichatoxin that has been isolated from the Taricha newts of California [150] and maculotoxin that has been isolated from Hapalochlaena maculosa (the blue-ringed octopus; [151]) have the same chemical structures as tetrodotoxin.
Cone snails, which are predatory marine gastropods feeding on fish, worms, or snails, produce a cocktail of venoms that are used for predation, defense, and competition. The major venom components are conotoxins or conopeptides. They are remarkably diverse in terms of structure and function [152,153,154]. Over 10,000 conotoxins or conopeptides are identified. They are biosynthesized as propeptides and are subject to extensive post-translational modifications in order to form mature peptides. The propeptides are cleaved by specialized venom endopeptidases belonging to the pathogenesis-related protein superfamily [155]. The mature peptides are comprised of 12~50 amino acid residues and 1~5 disulfide bridges [156]. Once they are injected into the prey or predators (fish, molluscs, or worms) [157], they act as fast-acting paralytics [158]. Depending on the chemical species of conotoxins, they function as inhibitors of voltage-gated calcium channels (e.g., ω-conotoxins), Nav, nicotinic acetylcholine receptors, serotonin receptors (e.g., σ-conotoxins), NMDA receptors (e.g., conantokins), G-protein-coupled receptors (e.g., ρ-conopeptides), and neurotransmitter transporters (e.g., χ-conopeptides), and so on [159]. For example, μ-conotoxins elicit a sodium channel inhibition through the direct pore block overlapping with tetrodotoxin at site 1, whereas ι-conotoxins enhance the channel opening by shifting the voltage dependence of the sodium channel activation to more hyperpolarized potentials. While α-conotoxins are selective antagonists of the nicotinic acetylcholine receptors, the conopeptide ρ-TIA with 19 amino acid residues inhibits α1-adrenoceptors. It is interesting that unpaired cysteine residues in conotoxins undergo posttranslational modifications, such as ADP-ribosylation [160], lipidations [161], nitrosylation [162], or cysteinylation [163]. These modifications may be involved in additional functionality, stabilization, subcellular localization, and detoxication.
μ-Conotoxins are peptides that are composed of 16~26 amino acid residues, structured by three disulfide bridges. They belong to the M superfamily of conopeptides (six cysteine residues, organized as CC–C–C–CC). The μ-Conotoxin bind to the extracellular S5–S6 loop of Nav, like tetrodotoxin and saxitoxin, although they discriminate further between Nav subtypes, having a higher affinity to the mammalian brain subtype Nav1.2 and the skeletal subtype Nav1.4 than to Nav1.7 and Nav1.8. This difference indicates that the binding sites of tetrodotoxin and μ-conotoxins only partially overlap and involve multiple Nav residues in the case of the larger μ-conotoxins, whereas the tetrodotoxin binding is crucially defined by relatively few residues in the pore of the Nav. The μO-conotoxins possess three disulfide bridges and belong to the O superfamily (six cysteine residues, organized as C–C–CC–C–C). The μO-conotoxins act as inhibitors of sodium channel conductance. Although the binding site of μO-conotoxins remains yet to be fully defined, it overlaps at least partially with those of the δ-conotoxin at the DIV of Nav (binding site 6). Another site is the voltage sensor of DII, which is shared with scorpion β-toxin (binding site 4). It is suggested, therefore, that the interaction of different toxins with a single region of the channel could be responsible for the opposite effects on the conductance. μO-conotoxins function as inhibitors of Nav, while δ-conotoxins and scorpion β-toxins function as activators of Nav [159]. Considering these facts, the Conus species protect themselves from the attack of the conotoxins through the mutation of the target sites and the sequestration and/or post-translational modification of the toxins [157,164].
The phylum of Cnidaria is the oldest animal venomous lineage. Its venom is a complex mixture of toxic compounds, including enzymes, pore-forming toxins, and neurotoxins. Actinoporins are the most abundant cnidarian pore-forming toxins, with a molecular weight of about 20 kDa, lacking in an intramolecular disulfide bridge. They specifically bind to sphingomyelin in the lipid membrane, and form oligomeric transmembrane pore, causing an osmotic imbalance and cell death. Most sea anemone species produce different isoforms of a specific actinoporins, which differ in isoelectric point, molecular weight, and cytolytic activity [165,166,167]. Actinoporins are biosynthesized as their prepropeptides, comprising of about 34 amino acid residues, which include the signal peptides of 19~21 amino acid residues. Intriguingly enough, although the actinoporins specifically target the sphingomyelin in the cell membrane, this lipid in sea anemones is replaced by its phosphono analogue. That is, the sphingomyelin possesses a phosphonocholine head group, to which the actinoporins cannot bind, and consequently, makes the sea anemones resistant to their own toxin [168].
Aerolysin-like pore-forming toxins are found mainly in pathogenic bacteria, but also in sea anemones and hydra. Hydralysins, pore-forming proteinous toxins from hydra, show paralytic, cytolytic, and hemolytic activities. They are secreted into the gastrovascular cavity immediately after the engulfment of prey, where they are bound to membranes of the ingested prey. However, the hydra itself is protected from the effect of its own lytic toxins, because hydralysins do not bind to hydra membranes, probably because of the lack of the receptor [169].
4. Toxins from Terrestrial Animals
Snake venoms are complex mixtures of organic and inorganic compounds that act on a variety of specific metabolic and physiological targets of prey, victims, and predators, assisting in feeding and defense [170,171]. The organic compounds are proteins/peptides in nature, including acetylcholine esterases, complements, disintegrins, defensins, growth factors, nucleases, nucleotidases, metalloproteinases, phospholipase A2, proteinase inhibitors, and others [172,173].
α-Bungarotoxin is an α-neurotoxin consisting of 74 amino acid residues with five disulfide bridges (GB No. P60615). It is isolated from the snake venom of Bungarus multicinctus and binds to the postsynaptic nicotinic acetylcholine receptor at the neuromuscular junction, almost irreversibly. The specific high-affinity binding of α-bungarotoxin to the acetylcholine receptor requiresfive amino acid residues of the C-terminal, and several amino acids that are located near the end of loop II, such as Trp28, Asp30, Arg36, and Lys52 (Figure 2A) [174,175].
Acetylcholine receptors are divided into two types, nicotinic and muscarinic receptors. Nicotinic acetylcholine receptors are pentameric structures consisting of five subunits that are arranged to create a cylindrical complex, forming an ion channel [176]. There are 12 neuronal specific subunits, that is, α2 to α10, and β2 to β4. Depending on the combination of subunits, the structural and functional diversities arise. All of the subunits have a conserved extracellular large N-terminal domain of about 200 amino acids, distinct and conserved; three transmembrane domains; a cytoplasmic loop of various size and amino acid sequence; and a fourth transmembrane domain with a variable extracellular C-terminal sequence. Snake toxins, such as α-cobra toxin and α-bungarotoxin, are bound only to the α-type subunit of acetylcholine receptor. These toxins bind to a hydrophobic pocket that is formed at the interface between the α-subunit and the adjacent subunit. For the ligand-binding, the disulfide bridge (Cys-loop) that is formed between Cys128 and Cys142, and the Cys–Cys pair at 192 and 193, are required. In addition, hydrophobic aromatic amino acids, including Tyr93, Trp149, Tyr190, and Tyr198, are involved in ligand binding. Glu45 and Arg209 are present in every member of the Cys-loop receptor family and they form a common link between the ligand-binding site and channel, including Val46, Ser269, and Pro272 (Figure 2B) [177,178,179].
All of the acetylcholine receptors from species that are sensitive to α-bungarotoxin, such as mice, have a tryptophan at position 187 and an aromatic amino acid residue at position 189. In those from the species that are less sensitive, such as snakes, mongooses, hedgehogs, and humans, these two amino acids are replaced by non-aromatic residues (Figure 2C). In addition, the acetylcholine receptor of the venom-resistant mongooses that feed on snakes has several other mutations, such as Ser191 to Ala, Pro194 to Leu, and Pro197, to His (Figure 2C) [180]. Furthermore, the Asn187 in mongooses and Asn189 in snakes are glycosylated [181]. However, although these mutated acetylcholine receptors do not bind α-bungarotoxin, they still retain their cholinergic properties. Resistance to α-bungarotoxin is believed to have evolved at least four times in mammals, as a consequence of the changes to the nicotinic acetylcholine receptor molecule to which the toxin binds. It is concluded, therefore, that snakes and some mammals, such as honey badgers, hedgehogs, mongooses, pigs, and humans, that are resistant to the attack of snake venom, have mutated and/or modified the acetylcholine receptors as the strategy of the resistance against snake venom [182,183,184]. α-Bungarotoxin acts also as an antagonist to the GABAA receptor [185].
Bothropstoxin-II comprising of 138 amino acid residues (GB No. P45881) is a phospholipase A2 that is isolated from Bothrops jararacussu snake venom, which induces platelet aggregation and ATP release reaction. The induction is shown to be evoked through multiple signal transduction pathways using several specific inhibitors, including genistein and staurosporine [186].
Atrolysin A is a zinc metalloproteinase that is isolated from the venom of the western diamondback rattlesnake, Crotalus atrox. It consists of 419 amino acid residues (GB No. Q92043) and shows proteolytic and hemorrhagic activities. Interestingly, both the proteolytic and hemorrhagic activities are partially inhibited by the opossum serum oprin, and completely inhibited by the opossum serum [187]. However, both the proteolytic and hemorrhagic activities of atrolysin B are completely inhibited by oprin. Oprin is homologous to human α-1B-glycoprotein. Atrolysin B is a zinc metalloproteinase that is isolated from the venom of the western diamondback rattlesnake, Crotalus atrox. It consists of 414 amino acid residues (GB No. Q90391). The inhibitory activity of the opossum serum may be related to the protection against snake venom [188,189]. In this connection, it is interesting that some opossums belonging to the family Didelphidae can eat pitvipers with impunity [190]. Botrocetin, one of the components in snake venom, is a non-enzymatic protein that causes the von Willebrand factor-dependent aggregation of platelets. Intriguingly, some amino acid residues within the botrocetin-binding regions in the von Willebrand factors are substituted in opossums. The prevention of the binding of botrocetin to the von Willebrand factor in opossums may be one of the reasons for the resistance. However, as snake venom contain dozens of toxic compounds, the evolution of the resistance requires adaptive changes at multiple loci.
Micrurotoxin 1 and micrurotoxin 2 are two toxins that are present in the Costa Rican coral snake’s venom that bind tightly to GABAA receptors [191]. Both of them consist of 64 amino acid residues, with five disulfide bridges (GB Nos. C0HJR1 and C0HJR2). The GABAA receptors belong to the pentameric Cys-loop superfamily of ligand-gated ion channel receptors, which encompasses the nicotinic acetylcholine, glycine, and serotonin receptors [178]. The mutation of His33 in micrurotoxin 2 to serine, impairs its function, indicating that this locus is vital for toxin activity. On the other hand, the micrurotoxin 1 function is influenced by mutations in the loop-C [192,193,194] of the α1 subunit of the GABAA receptor [191], indicating that loop-C is involved in the interaction between micrurotoxin and the GABAA receptor, as in the cases of toxins and nicotinic acetylcholine receptors.
Scorpion venom are highly complex mixtures of small peptides, proteins, mucoproteins, amino acids, biogenic amines, lipids, carbohydrates, and inorganic salts. Among them, non-disulfide-bridged peptides are attractive compounds, because they show antimicrobial, antimalarial, immunosuppressing, and anticancer activities, and may be relevant for the development of pharmaceutical drugs [195]. Another toxin is neurotoxins, and they are disulfide-bridged peptides with a significantly constrained structure. They act on various ion channels in excitable membranes, including sodium channels, potassium channels, calcium channels, and chloride channels [196,197]. This process is thought to have developed in response to the extended positive selection via predator-prey interactions.
Scorpion neurotoxins affecting Nav are functionally divided into α- and β-toxins, according to their primary actions on these channels. α-Toxins target the Nav receptor site 3, inhibiting channel inactivation, while β-toxins bind to site 4 of the Nav receptors [71,137,138,198]. Both toxins contain 60~80 amino acids that are linked by four disulfide bridges.
LqhII and LqhIII are α-toxins that are isolated from the venom of Leiurus quinquestriatus hebraeus, consisting of 64 amino acids with 4 disulfide bonds and 67 amino acids with 4 disulfide bonds, respectively (GB Nos. P59355 and P56678, respectively). The LqhII sequence reveals only one of each substitution of N-terminal and C-terminal amino acid, as compared to AaHII (GB No. P01484), which is isolated from Androctonus australis. LqhII and sea anemone toxins are shown to bind to the overlapping region comprising receptor site 3 on the rat brain and insect sodium channels (DIV S3-S4). The mutation of some amino acids in this region makes Nav resistant to scorpion α-toxin LqTx [199,200]. Interestingly, LqhII shows toxicity to mice, comparable to that of AaHII, while LqhII shows a 3.2-fold higher toxicity to cockroaches, as compared to AaHII, indicating that the N-terminal and C-terminal amino acids determine the species specificity of toxicity of the two toxins. LqhIII has an 80% sequence identity with the α-like toxin BomIII (GB No. P13488). LqhIII shows about a 2-fold lower toxicity to mice than BomIII, but is about 2-fold more toxic to cockroaches than BomIII. Thus, relatively minor changes in the sequence of scorpion toxins affect their relative species selectivity [201]. Moreover, it is shown that the tolerance of insects to a scorpion toxin AaIT occurs at both the pharmacokinetic and pharmacodynamics levels [202]. The CssIV from Centruroides suffuses belongs to the class of scorpion β-toxins (GB No. P60266), and shifts the voltage-dependent activation to more negative membrane potentials, leading to repetitive firing in muscles and nerves. This activity depends on the binding to DI S5-S6, DII S1-S2 and DII S3-S4, and DIII S5-S6. The mutations of the ritical amino acids in these regions result in a reduction of voltage-sensor trapping activity [203].
As for the resistance to scorpion toxins, several papers were reported. Rowe et al. reported that bark scorpion toxin induces pain in many mammals, including house mice and humans, by activating Nav1.7, but it has no effect on Nav1.8 [204]. On the other hand, for grasshopper mice, Onychomys torridus, Nav1.8 has several amino acid mutations, which bind bark scorpion toxins and inhibit Na+ currents, inducing analgesia. Especially, the mutations of amino acid residues in the DII SS2-S5 linker region of Nav1.8 are involved in this phenomena. Thus, by using a toxin that is bound to a non-target Nav, the resistance in grasshopper mice is aided by enhancing the interaction between toxin and receptor, such that the physiological consequences of the toxin binding are altered to the benefit of the targeted animal [204]. The long-eared bat (Otonycteris hemprichii) and pallid bat (Antrozous pallidus) can eat scorpions without harmful effects, although the exact resistance mechanisms to the toxins have not been clarified [205,206]. Legros et al. reported that the venom from the scorpion, Androctonus australis, is pharmacologically inactive on K+ channels and on the Nav from this scorpion [207].
Spider venom is made up of complex mixtures of polyamines; lectins; defensins; enzymes, such as proteinases; phospholipases and hyaluronidases; neurotoxins; and others. They act as receptor and/or ion channel toxins [208,209,210,211,212], antibacterial substances [213], and potentiators of erectile function [214], and so on. On the level of sequence identity and inter-cysteine spacing, spider toxins that target Nav channels are divided into 12 families [211]. Huwentoxin-IV (GB No. AAP33074) is a sodium channel inhibitor that is isolated from the venom of the Chinese tarantula, Ornithoctonus huwena, and is composed of 35 amino acid residues with three disulfide bridges. It belongs to the family 1. It preferentially inhibits the neuronal subtype Nav1.7 and is docked at the receptor site 4, which is located at the extra-surface DII S3-S4 linker region [215]. From the analysis of the mutants of huwentoxin-IV, it is suggested that the polar residues threonine-28, arginine-29, and glutamine-34 in the C-terminal play crucial roles in the interaction of huwentoxin-IV and Nav [216]. On the subesophageal ganglion neurons from the tarantula, at least three types of voltage-gated ion channels are co-expressed, namely, calcium channels, two types of potassium channels, and tetrodotoxin-sensitive sodium channels. Interestingly, these ion-channels are relatively insensitive to their own toxins. As for the sodium channels, huwentoxin-IV preferentially inhibits Nav1.7. However, the affinity of huwentoxin-IV for the tarantula tetrodotoxin-sensitive sodium channel is over 120-fold lower than for the human Nav1.7. A comparison of the amino acid sequences in the site 4 regions of the Nav1.7 of human and tarantula reveals that two crucial residues (Asp837 and Glu839) are substituted by two neutral residues (Gly837 and Ser839; GB No. ABH12275; Figure 3). This indicates that the substitutions of the acidic amino acids in the critical region with neutral amino acids may cause the self-resistance to their own toxin [216,217]. Jingzhaotoxin-I is a 33 amino acid residue inhibitor cysteine knot motif peptide that has been separated from tarantula, Chilobrachys jingzhao, venom. It inhibits the tetrodotoxin-resistant Nav that is expressed in mammalian cardiac myocytes and tetrodotoxin-sensitive Nav in mammalian sensory neurons, but does not inhibit the tetrodotoxin-resistant Nav in mammalian sensory neurons [218]. The toxin interacts with site 3, located at the extracellular S3–S4 linker of DIV. Interestingly, the single mutation of two amino acid residues (Asp1609 and Lys1613) in the S3–S4 loop of the DIV decreases the sensitivity of the toxin for the human Nav channel [219]. However, it is not clear how these mutations are related to the resistance to the toxin in the producing tarantula.
Amphibians like poisonous frogs are protected by an exocrine defense system that is composed of cutaneous poison glands, which are specialized cells secreting a variety of defense chemicals [220]. The defensive chemicals include alkaloids, biogenic amines, bufadienolides, and so on [221]. The poisonous frogs, such as Bufoniae and Dendrobatidae, sequester the lipophilic alkaloids, such as indolizidines, from dietary sources like mites and ants [222,223,224]. However, the resistance mechanism in poisonous frogs is only poorly understood. The resistance to batrachotoxin is a modification of the target, voltage-gated sodium channels [225,226]. Table 2 shows the toxin resistance-related genes in marine and terrestrial animals. The self-defense mechanisms in marine and terrestrial animals are mostly due to the modification or mutation of the targets. However, the toxins that have been discussed here are mainly peptides or proteins in nature, except for tetrodotoxin, saxitoxin, lipophilic alkaloids, and batrachotoxin. Producer animals also sequester the toxins or the defensive chemicals in specialized cells [221]. This is another important strategy for defense against themselves (self-defense) and intruders, and may be essential for a predator–prey struggle. In saxitoxin, the producer microorganisms use transporters as a defense mechanism, while the exposed animal (Mya arenaria) uses the mutation of the target, indicating that the defense mechanisms show inter-species and inter-population variation [72] (Table 1 and Table 2).
5. Plant Toxins
Plants produce a vast variety of secondary metabolites, differing in chemical structures and functions. They act as defense, signaling, and immunological compounds in plants, among many others. Camptothecin (CPT) is a water-insoluble tryptophan-derived quinolone alkaloid and is a lead compound for two FDA (Food and Drug Administration) approved antineoplastic drugs, irinotecan and topotecan [227,228]. It was originally isolated from Camptotheca acuminata more than 50 years ago [229]. However, it is now known that it is found in at least 16 different plant species, belonging to 13 unrelated genera. Moreover, it is produced not only by endophytic fungi, including Entrophosphora infrequens, Neurospora sp., and Fusarium solani [230,231], but also by endophytic bacteria from Pyrenacantha volubilis (Icacinaceae) [232]. The endophytic bacteria belong to the Bacillus species, and a 5 kb plasmid was isolated from one of the bacteria. It is speculated that the biosynthetic genes are present on the 5 kb plasmid, because the elimination of the plasmid by the treatment with acriflavine results in the loss of the production of CPT [232]. The reason for this remains to be clarified, because the plasmid is too small to cover the biosynthetic gene cluster for CPT.
CPT is an inhibitor of eukaryotic topoisomerase I. It is also toxic to most higher-plants. So, it is necessary for the producing plants to protect themselves from the attack of their own toxin. A comparison of the amino acid sequences of topoisomerase Is of the CPT-producing and CPT-nonproducing plants revealed that asparagine at 722 (numbered according to human topoisomerase I) in the nonproducing plants, such as Ophiorrhiza japonica, and humans is substituted by serine in the CPT-producing plants, such as Ophiorrhiza pumila, O. liukiuensis, and Camptotheca acuminata (Figure 4), suggesting that the mutation of Asn722Ser is responsible for the self-resistance to CPT in the producing plants [233,234].
Sclareol is a labdane-type dipertene that is detected on the leaf surface of the Nicotiana species. It shows antifungal activity and also inhibits plant growth. It is reported that the drug resistance-type ATP-binding cassette (ABC) transporter, NpPDR1, is involved in extracellular sclareol secretion in Nicotiana plumbaginifolia, and the expression of NpPDR1 makes it resistant to sclareol. Interestingly, NpPDR1 is constitutively expressed in the trichome, where the biosynthesis of sclareol occurs essentially [235,236,237]. After biosynthesis, phytotoxins are often stored in specialized organs [238].
ABC transporters are involved in the transportation of defense molecules, sequestration of xenobiotics and intracellular metabolites to the vacuole, and many others [239,240,241]. Berberine is a benzylisoquinoline alkaloid and is used as an antidiarrhetic and antimalarial drug. Coptis japonica accumulates berberine exclusively in the vacuoles (sequestration), whereas Thalictrum minus secretes biosynthesized berberine outside of the cells (excretion). The ABC transporter CjABCB1/CjMDR1 and a P-glycoprotein-like ABC transporter are involved in these processes [242,243]. However, a proton-antiporter may also mediate the membrane transport of berberine [244]. Vinblastine and vincristine are indole alkaloids that are isolated from Catharanthus roseus and are used as anticancer drugs that disrupt the microtubule formation, and interfere with amino acid and glutathione metabolisms, and nucleic acid and lipid biosynthesis [245,246]. In C. roseus, an ABC transporter CrTPT2 functions as an efflux transporter of catharanthine to the cell surface. Catharanthine is a biosynthetic precursor of vinblastine and vincristine [247,248]. Flavonoids are a major class of plant secondary metabolites. Some flavonoids show anti-oxidation, anti-inflammatory, and antitumor activities [249]. To avoid their toxic effects, producing plants sequester flavonoids within vacuoles by transporting them with the multidrug resistance-related protein subfamily of ABC transporters [250].
Sanguinarine is a benzophenanthridine alkaloid with cytotoxic properties, such as the induction of oxidative DNA damage and rapid apoptosis [251]. It also inhibits the growth of cultured cells of Nicotiana and Arabidopsis. So it is either accumulated in the vacuole or excreted into the cell wall for self-protection [252,253]. In addition, it is reduced to the less toxic dihydrosanguinarine [254,255,256]. Coniine is a piperidine alkaloid isolated from Conium maculatum and is known as the killer of Socrates in 399 BC [257]. It is a nicotinic acetylcholine receptor antagonist, which leads to the inhibition of the nervous system and the antinociceptive effect [258]. Interestingly, the cabbage looper (Trichoplusia ni) larvae that are raised on coniine and γ-coniceine-enriched diets do not show any effect on the growth and only a slight reduction of development time. The three reasons are proposed to explain these responses, namely: (1) a decreased consumption rate, (2) efficient excretion of ingested alkaloids unmetabolized in frass, and (3) partial detoxification of alkaloids by cytochrome P450 [259]. This proposal is supported by the fact that the larval growth is decreased in the presence of piperonyl butoxide, a cytochrome P450 inhibitor.
The pyrrolizidine alkaloids represent a class of plant secondary metabolites [260,261,262], and are strongly hepatotoxic, pneumotoxic, and teratogenic to most vertebrates and invertebrates. They are most likely produced as a chemical defense mechanism against herbivores. Some pyrrolizidine alkaloids, such as clazamycins and jenamidines, are biosynthesized by Gram-positive and Gram-negative bacteria [263]. Senecio species biosynthesize pyrrolizidine alkaloids in unique patterns, as senecionine N-oxide, as a common intermediate in the roots. Then, species-specific alkaloids are thought to be produced in shoots. These alkaloids are stored in vacuoles in the form of mainly their N-oxides [264]. On the other hand, a protective role of plant pyrrolizidine alkaloids is observed in specialized herbivorous insects. A number of insect herbivores have evolved adaptations not only to overcome the defense barrier of pyrrolizidine alkaloid-protected plants, but also to sequester and utilize the alkaloids for their own defense against predators. For example, the larvae of the European cinnabar moth, Tyria jacobaeae, sequester the alkaloids from their larval host plant Senecio jacobaea. Larvae raised on a pyrrolizidine alkaloid-free diet prove consistently palatable to wolf spiders, whereas the larvae and adults containing the alkaloids were rejected [265,266]. Leaf beetles of the genus Oreina are another example of pyrrolizidine alkaloid sequestration for insect defense. Leaf beetles release their defense compounds from special exocrine glands that are located in the elytra and pronotum. Most insects with the alkaloid-sequestering species store and maintain the alkaloids as N-oxide. The N-oxides are less toxic than the parent alkaloids, so it is more convenient for self-protection. In arctiids, the sequestered pyrrolizidine alkaloid N-oxide are found in all of the tissues, but preferentially in the integument [267,268].
Steroidal glycoalkaloids, such as α-solanine and α-chaconine, found in solanaceous food plants like potato and tomato, are antinutritional factors for humans. They cause gastrointestinal and neurological disorders and are lethal for humans at high concentrations. They disrupt membranes and inhibit acetylcholinesterase activity [269]. The steroidal glycoalkaloids consist of two structural components, the aglycone, a cholesterol-derived compound; and a carbohydrate side-chain. The biosynthetic gene clusters of α-tomatine and α-solanine were cloned [270,271]. The endogenous enzymes with glycosidase activity remove sugar molecules from the saccharide moiety of α-tomatine, creating a less cytotoxic compound, α-tomatidine. This detoxification mechanism was also observed in Fusarium oxysporum and Cladosporium fulvum [272,273], indicating that the presence of glycosidases in the plants and fungi are associated with modulation of the toxicity of steroidal alkaloids in the defense response. This indication is supported by the fact that the aglycones solanidine and tomatidine produce only a slight to negligible inhibition of acetylcholinesterase activity [269,274], and that the glycosylation by GAME1, a gene that is involved in the biosynthesis, is crucial to prevent the toxic effect of the alkaloids to the plant cells [275]. On the other hand, benzoxazinoids are stored as biologically inactive glycosides that are cleaved by β-glucosidase upon attack, releasing the active aglycones [276]. The aglycones are active against bacteria, fungi, and herbivores. The iridoid glucosides, such as aucubin and catalpol, are other examples. After the attack of herbivores, iridoid glucosides are cleaved by glucosidases to the toxic terpenoid aglycones [277,278].
More than 3000 plants species, such as almond and sweet cherry, use hydrogen cyanide (HCN) as a fast-acting, powerful toxin to protect their seeds and leaves against attack from herbivores. The cyanogenic glucosides, prunasin and amygdalin, release HCN upon cell wounding. Tissue disruption brings together both cyanogenic glycosides and the HCN-releasing enzymes, β-glucosidase and hydroxynitrile lyase, which are stored in separate compartments in the intact plant cells [279,280]. Glucosinolates are produced by Brassicaceae, such as cabbage, rapeseed, and radish. Upon tissue disruption, glucosinolates are cleaved by myrosinase, a glucosidase, to form toxic isothiocyanates [281]. These binary glycoside and glycosidase systems are referred to as two-component plant chemical defense [278].
Cardenolides are composed of aglycones of steroid structures that are derived from terpenoids and sugars [282]. Although mostly recognized as plant compounds, cardenolides are produced via the cholesterol pathway in animal tissues. Ouabain and digoxin are the typical endogenous cardiac glycosides. They are strong inhibitors of the Na+/K+-ATPases. Ouabain is toxic to locusts or cockroaches, while to caterpillars of the tobacco hornworm ouabain is tolerated. This insensitivity is explained by the high concentration of K+ in the lepidopteran hemolymph. K+ has an antagonistic effect to the ouabain binding to the Na+/K+-ATPase. On the other hand, the caterpillars of the monarch butterfly sequester cardenolides from its apocynaceous host plants [283]. In addition, mutations of critical amino acid residues of the target (Na+/K+-ATPase) lower the sensitivity of the monarch butterfly (Danaus plexippus) and the milkweed bug (Oncopeltus fasciatus) to cardenolides. For example, the milkweed bug has three copies of the Na+/K+-ATPase α1 subunit gene, α1A, α1B, and α1C. The α1C knockdowns with RNAi cause difficulties in motor function and have a reduced survival rate, indicating that the α1C gene is the most important for survival. On the other hand, although the α1A or α1B knockdowns with RNAi sustain the normal survival rate, they are no longer able to tolerate cardenolides. The comparison of amino acid residues that are involved in the binding of ouabain [284] indicates that substitutions of Gln111Thr, Asn122His, and Phe786Asn mediate insensitivity to cardenolides (Figure 5) [285,286,287]. Furthermore, it is reported that the oleander hawk moth (Daphnis nerii) uses the perineurium as a diffusion barrier (restriction) for polar cardenolides like ouabain and efflux transporters (exclusion), for non-polar cardenolides like digoxin. As quinidine and verapamil inhibit the barrier, P-glycoproteins-like transporters are suggested to be involved in the barrier. These results suggest that the lepidopteran perineurium functions as a diffusion barrier for polar cardenolides and forms an active barrier for non-polar cardenolides [288].
Nicotine, an alkaloid derived from the leaves of tobacco plants (Nicotiana tabacum, Nicotiana attenuate, and other Nicotiana species), is the primary addictive agent in tobacco products and binds to nicotinic acetylcholine receptors [289]. Nicotine is stored in the trichomes of tobacco leaves (sequestration). N. attenuate, a species of wild tobacco, is attacked by larvae of both specialist (Manduca sexta) and generalist (Spodoptera exigua) lepidopteran herbivores. M. sexta is highly tolerant to nicotine. Interestingly, ingestion of nicotine and its N-oxides to M. sexta larvae induces cytochrome P-450 CYP6B46. Nicotine-induced CYP6B46 is used to efflux midgut-nicotine into the hemolymph and it facilitates nicotine exhalation. Nicotine, but not nicotine-N-oxide, deters predatory wolf spiders. On the other hand, the S. exigua larvae oxidizes nicotine and are more susceptible to predation by wolf spiders [290].
The defenses of plants to insects and pathogens are initiated by the recognition of insect oral secretion and signals from injured plant cells. These early events include damage-induced ion imbalance, variations in membrane potentials, Ca+-signaling, production of reactive oxygen species, kinase activities, and phytohormones [291,292,293,294,295]. N. attenuate α-DIOXYGENASE1 is an oxylipin-forming gene that is elicited during herbivory by fatty acid-amino acid conjugates, which are contained in oral secretion of M. sexta. N. attenuate specifically accumulates 2-hydroxylinolenic acid during feeding by M. sexta larvae. α-DIOXYGENASE1-silenced plants are less resistant to a M. sexta attack, indicating that 2-hydroxylinolenic acid, produced from linolenic acid by attack-activated-α-DIOXYGENASE1, participates in defense activation during insect feeding [296].
Menthol is a cyclic monoterpene alcohol, which possesses cooling characteristics and is a major constituent in the essential oil of Mentha canadensis L. It acts upon the transient receptor potential melastatin family member 8 (TRPM8) receptors by rapidly increasing the intracellular calcium and mobilizing the calcium flux. Aside from its cold-inducing sensation capabilities, menthol exhibits cytotoxic effects in cancer cells, induces reduction in malignant cell growth, and engages in synergistic excitation of GABA receptors and sodium ion channels, resulting in analgesia. It is often stored in trichomes from which it is released upon cell rupture [297,298].
Lupins produce quinolizidine alkaloids, such as albine, lupanine, and multiflorine, in leaf chloroplasts, export them via the phloem all over the plant, and the accumulate in epidermal tissues, especially in reproductive organs. Quinolizidine alkaloids are known to interfere with the nervous systems of animals. As for lupins, alkaloid-rich and alkaloid-free varieties (sweet lupins) are known [299,300]. It is shown that aphid generalists such as Myzus persicae only suck on sweet lupins, and not on alkaloid-rich varieties with high alkaloid contents in the phloem. On the other hand, specialist aphids, such as Macrosiphum albifrons, live on lupins, sequester the dietary alkaloids, and use them as a defense against predators [301]. Many other animals show a similar discrimination. Table 3 shows the toxin resistance-related genes in plant. In plants, sequestration in specialized organs like vacuoles and the related transporters are the major mechanisms of the resistance. These mechanisms are used as the defense, not only in producer plants, but also in herbivore animals such as insects. In addition, the mutation of targets such as topoisomerases and chemical conversion of glycoside to aglycones or reverse are also observed.
6. Toxins from Fungi
Fungi, in particular the fruiting bodies of higher fungi, are potential victims of attack by fungivores and microorganisms. The fruiting bodies of mushrooms are a rich source of secondary metabolites with unusual chemical structures. Aflatoxins are among such metabolites. They are produced mainly by Aspergillus flavus and A. parasiticus. The four major natural aflatoxins are known as aflatoxins B1, B2, G1, and G2. Aflatoxin B1 and aflatoxin B2 are hydroxylated and excreted in the milk as less toxic aflatoxins, M1 and M2. Aflatoxin B1 is metabolized and activated in the intestine and liver by cytochrome P450 to aflatoxin B1-8,9-epoxide. Aflatoxin epoxide is highly electrophilic and reacts with the DNA guanine moiety to form covalent bonds at the N-7 guanine residue, leading to depurination and carcinogenesis. Aflatoxin epoxide also attacks mitochondrial DNA and disrupts ATP production. These damages lead to hepatic fibrosis, decreased liver function, and cancer. Differences of sensitivity to toxicity of aflatoxin B1 are due to the differences in its metabolism [302,303]. The aflatoxin biosynthetic gene clusters ranging 82 kb were cloned from Aspergillus flavus [304,305,306] and A. parasiticus [307]. Although aflT codes a membrane-bound protein with homology to antibiotic efflux pumps and is presumed to be involved in aflatoxin secretion, the disruption of this gene does not affect theaflatoxin formation [306,307]. Other genes that are related to self-resistance are not detected, although numerous genes for ABC and MFS transporters are present in the genomes [304,308]. Sterigmatocystin, a mycotoxin that is produced by the Aspergillus fungi, and causing a carcinogenic, mutagenic, and teratogenic effect as aflatoxins, is biosynthesized as a precursor of aflatoxin A [304,309,310].
Ochratoxins and citrinin are produced by several species of the genera Aspergillus and Penicillium. The fungi producing ochratoxins and citrinin are commonly encountered in animal feed and human food. Ochratoxins are pentaketide-derived dihydroisocoumarin moieties that are peptide-bonded to phenylalanine derivatives. Three ochratoxins are known and the order of toxicity is ochratoxin A, ochratoxin B, and ochratoxin C. The most sensitive effects of ochratoxin A are on the kidney, causing nephropathy and urinary tumors. The ochratoxin biosynthetic gene clusters were cloned from five species, namely, Aspergillus steynii, A. westerdijkiae, A. niger, A. carbonarius, and Penicillium nordicum [311,312,313,314,315,316]. A comparison of the five clusters revealed that the central part of the clusters consists of five ORFs, namely, halogenase, bZIP transcription factor, cytochrome P450, non-ribosomal peptide synthetase (NRPS), and polyketide synthase, although the genes in the flanking regions are different [314]. Interestingly, the gene cluster from P. nordicum contains ORFs that are homologous to an organic anion transporter and a nitrate transporter [312,313]. The former transporter was reported to be responsible for the transport of ochratoxin into the outside of the cell. Citrinin is a mycotoxin that is produced by genera Penicillium, Aspergillus, and Monascus, and shows a nephrotoxic activity. The biosynthetic gene cluster was cloned as a 43kb DNA fragment from M. aurantiacus and M. purpureus [317,318]. The Orf5 codes for a putative membrane transport protein.
Fusarium head blight is a serious fungal disease of grains that is caused by the infection of a range of Fusarium fungi. Fusarium-infected grains are often contaminated with mycotoxins such as trichothecenes, like nivalenol and deoxynivalenol (vomitoxin); fumonisins; and zearalenones. These contaminated toxins are also hazardous to humans and livestock. Fusarium species produce both sexual and asexual spores. These spores are resistant to environmental stresses, and play important roles in the development and propagation of the Fusarium species. The determination of the whole genome sequences of F. graminearum [319] and various omics analyses [320] clarify the penetration and invasion strategies of F. graminearum, and hence, the defensive strategies in the host plants, such as wheat and barley, at the genetic level. As a result, signaling molecules such as salicylic acid, jasmonic acid, and ethylene are elucidated to initiate the signal transduction systems in the defense and pathogenesis systems [321,322]. Furthermore, plant proteins, such as ABC transporters, uridine diphosphate-glucosyltransferases, cytochrome P450s, and glutathione-S-transferases, are shown to be involved in deoxynivalenol detoxication [323]. On the other hand, the trichothecene biosynthetic gene cluster was cloned from F. graminearum, where one efflux pump gene (Tri12) was identified [324,325], indicating that the efflux pump excretes the trichothecene into the outside of the cells in the trichothecene-producing organisms. F. graminearum produces a red pigment, aurofusarin, which is a polyketide derivative. Interestingly, the biosynthetic gene cluster contains an efflux pump gene, although aurofusarin is not a mycotoxin. However, the aurofusarin deficient mutants increase the level of the mycotoxin zearalenone [326]. Zearalenone is a nonsteroidal estrogenic mycotoxin that is produced by several species of Fusarium fungi. Zearalenone has major effects on reproduction in females, but it affects the male reproductive system as well [327]. The biosynthetic gene cluster of zearalenone was cloned from Fusarium graminearum as a 50 kb DNA fragment [328,329,330]. A gene for monocarboxylate transporter-like protein (GzMCT) is located adjacent to the gene cluster [329], indicating that the transporter may be involved in the excretion of zearalenone from the inside of the cells to the outside. Clonostachys rosea is a soil-borne ascomycete and is known to be a potential biological control agent against various plant pathogens, including zearalenone-producing Fusarium culmorum. The enzyme zearalenone hydrolase in C. rosea is shown to be responsible for the transformation of zearalenone to the far less product [331,332]. Fumonisins are a group of mycotoxins that are produced by Fusarium species and have been shown to cause liver damage in various species, including primates. The genetic and biochemical analyses of Fusarium identified a fumonisin biosynthetic gene cluster, including two transporter proteins [333,334]. The gene clusters were also identified in the Aspergillus species, where a gene for the transporter protein was present [335,336].
Penitrem A is an indole diterpene mycotoxin that is produced by the Penicillium species. It is biosynthesized through paxilline and secopenitrem. The biosynthetic genes for the penitrems cloned from Penicillium crustosum consist of two separate clusters, which contain the transporter gene ptmT [337]. The genes ptmGAQMBCP are highly homologous to those of the paxilline gene cluster. Roquefortine C is a mycotoxin belonging to a class of naturally occurring diketopiperazines that are produced by the Penicillium species. It shows bacteriostatic and neurotoxic activities. Roquefortine C was proposed to be the precursor of meleagrin and neoxaline [338]. The roquefortine/meleagrin biosynthetic gene cluster was cloned from Penicillium chrysogenum, where a facilitator superfamily transporter gene was present [339]. Aphidicolin, a fungal diterpene that is isolated from Cephalosporium aphidicola, is a specific inhibitor of the DNA polymerase α. The gene cluster for the aphidicolin of the 15.6 kb DNA fragment was cloned and sequenced, which includes the ABC transporter protein composed of 564 amino acid residues [340].
Sirodesmin PL is a phytotoxin that is produced by the fungus, Leptosphaeris maculans. It causes a chlorotic lesion on plant leaves, and has antibacterial and antiviral activities. Sirodesmin PL is a member of the epipolythiodioxopiperazine (ETP) class of fungal secondary metabolites. The biosynthetic gene cluster of the 68 kb DNA fragment was cloned and sequenced, containing MDR1 type ABC transporter, SirA. It is supposed to be involved in toxin export and self-protection [341,342,343]. Gliotoxin is produced by several fungi, including Gliocladium fimbriatum, Aspergillus fumigatus, Trichoderma, Penicillium, and some Candida species. It has an immunosuppressive activity and is a virulent factor of the human fungal pathogens. It is also a member of the epipolythiodioxopiperazine (ETP) mycotoxins. The biosynthetic gene cluster was cloned from A. fumigatus, containing a major facilitator superfamily type transporter [342]. In addition, GliT, a gliotoxin reductase, plays an important role in the self-resistance [344,345].
The ergot alkaloids are a family of secondary metabolites that are produced by several orders of fungi in the phylum Ascomycota, and particularly in plant pathogens and plant symbionts of the family, Clavicipitaceae. The alkaloid profiles are also diverse. Their activities are derived from their affinity for receptors for the monoamine neurotransmitters [346]. The activities of ergot alkaloids include vasoconstriction or vasodilation, stimulation of uncontrolled muscle contraction and hallucination, and other effects on the central nervous system. The gene cluster for ergot alkaloids was cloned from Claviceps purpurea, extending over 68.5 kb, containing four different nonribosomal peptide synthetase genes [347,348,349]. Now, the biosynthetic gene clusters were cloned in several species of fungi [350,351,352,353,354]. So, 14 genes, that is, dmaW, easF, easC, easE, easD, easA, easG, cloA, lpsB, lpsA, lpsC, easH, easP, and easO, have been shown to direct steps in the ergot alkaloid biosynthetic pathway of Clavicipitaceae, although only Periglandula ipomoeae is known to have all of them. However, no transporter gene was present in these regions [351]. Interestingly, the genes that are common to the clusters encode the enzymes catalyzing early shared biosynthetic steps, whereas those that are unique to the clusters of specific fungi, encode later, lineage-specific steps [347,350,355,356]. It is speculated, therefore, that the responsible genes have been evolved through multiple events of gene duplication, gene gain, and gene loss.
Loline alkaloids are produced by endophytic fungi, protecting host grasses by affecting a large range of insects, providing resistance to vertebrate and invertebrate herbivores, and pathogens and parasites [357]. The two biosynthetic gene clusters (LOL-1 and LOL-2) were detected in the fungal symbiont Neotyphodium uncinatum. Nine genes were identified in a 25 kb region of LOL-1, and LOL-2 contained the homologs lolC-2 through lolE-2 in the same order and orientation [356,357,358,359,360]. The biosynthetic gene clusters of indole-diterpenes such as paspalinine, paspalitrem A, terpendole C and E, and lolitrem B and E, and of peramine in epichloae and other Clavicipitaceae, were cloned and compared [356,361,362,363,364]. The gene cluster for peramine contains two putative members of the facilitator superfamily of transporter proteins, although it is not clear if these proteins are involved in the peramine excretion. Interestingly, these proteins are conserved in Fusarium graminearum, Neurospora crassa, Magnaporthe grisea, and Aspergillus nidulans [362].
Swainsonine is an indolizidine alkaloid that is produced by insect and plant pathogens and symbionts belonging to the order Hypocreales, Chaetothyriales, Onygenales, Pleosporales, and Leotiomycetes. It inhibits, specifically, α-mannnosidase II in the Golgi apparatus, disrupting the endomembrane system of the cells. The biosynthetic gene clusters were cloned from six species, which contained a transmembrane transporter SwnT, except the two endophytes, Ipomoea carnea and Alternaria oxytropis [365]. Table 4 summarizes the toxin resistance-related genes in the fungi. As for the self-resistance against toxins from the fungi, the predominant strategies are transporters. The transformation of the toxins, like zearalenone, to less toxic derivatives is also reported. However, it seems that much more analyses are needed to clarify the detailed self-resistance strategies in fungi.
7. Antibiotic Resistance
Drug resistance, especially antibiotic resistance, is one of the most prevalent and threatening events in public health. The genes of antibiotic resistance are hypothesized to be derived from the antibiotic-producing bacteria, such as Streptomyces [366,367,368]. However, thanks to the metagenomic and high-throughput sequencing technologies, the current knowledge on resistome, the collection of the resistance genes [369,370], has expanded tremendously [371,372,373]. The aminoglycoside antibiotics, such as streptomycin, kanamycin, and gentamicin, interfere with protein synthesis by acting on the smaller 30S subunit of the bacterial ribosome, causing bactericidal effects against the pathogens [374]. In addition, the possibility for the treatment of the human immunodeficiency virus infection has been demonstrated [375]. Streptomycin is the first aminoglycoside antibiotic [376]. The aminoglycoside antibiotics are classified into two groups, those whose activities are affected and those not affected by methylation of 16S rRNA [374]. Streptomycin belongs to the latter group. The biosynthetic gene cluster was cloned as a 90kb DNA fragment [377,378] (GB No. AJ862840). It contains two phosphotransferases (StrA and StrK) and the EamA/RhaT family transporter, located adjacent to the cluster. These proteins are supposed to be involved in the self-resistance in Streptomyces griseus [379] (GB No. NC_010572).
Kanamycin was isolated from Streptomyces kanamyceticus [380] and is the 4,6-disubstituted 2-deoxystreptamine-containing amino glycoside antibiotic, together with gentamicins, tobramycin, and amikacin. Their activities are compromised by methylation of 16S rSNA. The kanamycin biosynthetic gene clusters were cloned [381,382,383,384] (GB Nos. AJ582817, AB164642, and AB254080). They contain genes for aminoglycoside 6′-N-acetyltransferase (kanM) and 16S rRNA methyltransferase (kmr), indicating that they are involved in self-resistance. Furthermore, there are several efflux (KanO and KanN) and ABC transporter proteins (KanS, Kan R and KanQ). Interestingly, kanamycin A and kanamycins B and C are biosynthesized in two different routes [385]. The gentamicin biosynthetic gene clusters were cloned from Micromonospora echinospora [381] (GB Nos. AJ575934, AJ628149, and AY524043). Three proteins within the clusters (GtmL/GmrB/GrmO, GtmF/GmrA/GrmA, and GtmK/GenV/GntO) were proposed to be involved in self-resistance. GtmL/GmrB/GrmO and GtmF/GmrA/GrmA are rRNA methyltransferases, and GtmK/GenV/GntO are transmembrane efflux proteins. Tobramycin is 3′-deoxykanamycin B. The gene clusters were cloned from Streptoalloteichus hindustanus [381,386] (GB Nos. AB103327, AJ579650 and AJ810851). Acetyltransferases and phosphotransferases, which may be involved in the self-resistance, are present outside of the clusters. Furthermore, two transporter proteins are present (TobU and TobT; GB Nos. CAH18564, and CAH18551, respectively).
Neomycin (fradiomycin), paromomycin, and lividomycin belong to 4,5-disubstituted 2-deoxystreptamine-containing aminoglycoside antibiotics. Their activities are compromised by methylation of 16S rRNA. The neomycin biosynthetic gene cluster was cloned from Streptomyces fradiae as a 50 kb DNA fragment (GB No. AJ629247). Two proteins, AphA [387] (GB No. CAF33306) and AacC8 (GB No. CAF33325), were proposed to be involved in the self-resistance. They are aminoglycoside 3′-phosphotransferase and aminoglycoside 3-acetyltransferase, respectively. In addition, two ABC transporters (GB Nos. CAF33314 and CAF33315) were detected within the cluster. The paromomycin biosynthetic gene cluster was cloned from Streptomyces rimosus as a 48 kb DNA fragment (GB No. AJ628955). The aminoglycoside 3′-phosphotransferase (ParR) and ABC transporters (ParT and ParU) that are located within the cluster are homologous to those in the neomycin biosynthetic gene cluster. Interestingly, two acetyltransferases (GB Nos. CAG44462 and CAG44463) that are involved in self-resistance are present in other locations [388] (GB No. AJ749845). The lividomycin biosynthetic gene cluster was cloned from Streptomyces lividus as a 40 kb DNA fragment (GB No. AJ748832). No resistance-related gene was detected within the cluster, except two ABC transporter genes (GB Nos. CAG38699 and CAG38700). Hygromycin B is an aminocyclitol antibiotic that inhibits the protein synthesis and 30S ribosomal subunit assembly. The producer Streptomyces hygroscopicus is highly resistant to hygromycin B due to the presence of hygromycin B phosphotransferase activity [389]. Hygromycin A, structurally unrelated to hygromycin B, inhibits the peptidyltranferase reaction of protein synthesis. The hygromycin A biosynthetic gene cluster of 31.5 kb DNA fragment was cloned [390]. O-phosphotransferase Hyg21 is involved in self-resistance [391]. Istamycin that is produced by Streptomyces tenjimariensis is an aminoglycoside antibiotic [392]. FmrT consisting of 211 amino acid residues was proposed to be the rRNA methyltransferase involved in self-resistance [393]. Istamycin is also acetylated by kasugamycin-producing Streptomyces kasugaensis. [394]. Kasugamycin is another aminoglycoside antibiotic that is produced by S. kasugaensis. It is used mainly for the prevention of the growth of a fungus causing rice blast disease. The ABC transporter genes, kasKLM, are responsible for the self-resistance of a kasugamycin-producer strain [395] (GB No. AB033992). Fortimicin (astromicin) is an aminoglycoside antibiotic that is produced by Micromonospora olivasterospora, and fmrO encoding 16S rRNA methyltransferase, plays a role in self-resistance [396]. The streptothricin group antibiotics show a broad antibacterial spectrum. However, their characteristic delayed toxicity prevents their clinical application. The biosynthetic gene cluster contains genes for an acetyltransferase and two ABC transporters, which may play a role in self-resistance [397,398,399] (GB Nos. AB684620 and AB684619).
The macrolide antibiotics are a class of natural products that consist of a large macrocyclic lactone ring, to which one or more deoxy sugars are attached [400,401]. The lactone rings are usually 14-, 15-, or 16-membered. Erythromycin and oleandomycin belong to 14-membered macrolides. The erythromycin biosynthetic gene clusters were cloned from Saccharopolyspora erythraea and Actinopolyspora erythraea [402,403,404] (GB No. AM420293). Within the clusters, ermE encoding N-6-aminoadenine-N-methyltransferase is involved in self-resistance. Esterases, efflux proteins, phosphotransferases, acetyltransferases, glycosyltransferases, and dioxygenases are also proposed to be involved in self-resistance [404]. Two ABC transporters (OleB and OleC) and a glycosyltransferase (OleD) are proposed to be involved in the self-resistance of oleandomycin-producing Streptomyces antibioticus [405,406]. Tylosin belongs to a 16-membered macrolide antibiotic. The tylosin biosynthetic gene cluster was cloned from Streptomyces fradiae [407]. The resistance genes encoding rRNA methyltransferase (tlrB; GB No. AAD12162) and the ABC transporter (tlrC; GB No. AAA26832) are located at both ends of the cluster. Mycinamicin, which is produced by Micromonospora griseorubida, is a 16-membered macrolide antibiotic. The biosynthetic gene cluster was cloned [408]. The myrB encoding rRNA, methyltransferase, is involved in self-resistance [409]. Methymycin is a 12-membered macrolide that is isolated from Streptomyces venezuelae. The modification of 23S rRNA by PikR1 and PikR2, and glycosylation/deglycosylation play the self-resistance in the producing strain [410].
Tetracyclines are members of the polyketide family natural products, and are characterized by their tetracyclic ring structure [411,412]. The gene cluster for oxytetracycline biosynthesis was cloned from Streptomyces rimosus as a 25 kb DNA fragment, including bacterial Type II polyketide synthases genes [413,414] (GB No. DQ143963). Two resistance-related genes, otrA and otrB, are present at both ends. The otrA encodes the TetM-like ribosome protection protein [415] (GB No. CAA37477) and the otrB encodes an efflux MFS transporter (GB No. AOR83343). The gene cluster for chlortetracycline biosynthesis was cloned from Streptomyces (Kitasatospora) aureofaciens (GB Nos. CP020567 and HM627755). Two resistance-related proteins (GB Nos. ARF80631 and ARF80644) are detected within the cluster. They are the MFS efflux transporter and the GTP-binding ribosomal protection protein [416,417], respectively. In addition, OtrC proteins (ABC transporters, GB Nos. AAR96051 and AAR96052) are suggested to be concerned with the self-resistance [418,419].
Chloramphenicol is an antibiotic that is produced by Streptomyces venezuelae and other Streptomyces species. It behaves primarily by inhibiting protein synthesis and is used for the treatment of Gram-positive and Gram-negative bacterial infections. However, the side effects, such as bone marrow suppression, nausea, and diarrhea, restrict its common use. The whole DNA sequence of the S. venezuelae genome was determined, including the chloramphenicol biosynthetic gene cluster [420] (GB Nos. FR845719 and AF262220). Two transporters [421] (GB Nos. CCA54203 and CCA57351), acetyltransferase [422], and a phosphotransferase [423,424] (GB No. CCA57350) were reported to be involved in self-resistance. In addition, the chloramphenicol hydrolase that removes the dichloroacetyl moiety from chloramphenicol may be involved in the resistance [425]. However, chloramphenicol acetyltransferase activities, which are responsible for the inactivation of chloramphenicol in various bacteria, including Streptomyces coelicolor Mueller, S. acrimycini, and S. griseus, are not detectable in chloramphenicol-producing S. venezuelae [426,427].
The glycopeptide antibiotic, vancomycin, and the structurally related antibiotics are supposed to be the last lines of defense against a variety of serious infections that are caused by Gram-positive bacteria. These antibiotics cannot penetrate the peptidoglycan layer and do not act against Gram-negative bacteria. The gene cluster for vancomycin biosynthesis was cloned from Amycolatopsis orientalis (GB No. HE589771). Within the cluster, the ABC transporter (GB No. CCD33134) is located. Besides this, VanHAX resistance cassette exist [428]. These enzymes participate in the resistance mechanism in vancomycin-resistant enterococci strains by redirecting a portion of the peptidoglycan pathway [429,430,431]. Interestingly, the VanHAX is detectable not only in vancomycin-related glycopeptide-producing Actinoplanes teichomyceticus [432] and Streptomyces toyokaensis [433], but also in non-producing Streptomyces coelicolor (GB No. AL939117). The biosynthetic gene clusters for vancomycin-related glycopeptide antibiotics, such as balhimycin, chloroeremomycin, A40926, A47934, and teicoplanin were cloned [432,433,434,435,436,437,438,439,440,441]. The ABC transporters are present in all of these clusters. It is reported that the glycopeptide antibiotic A40926 producer Nonomuraea species possesses a novel d,d-peptidase/d,d-carboxypeptidase, which is involved in self-resistance and peptidoglycan maturation [442]. Moenomycins, which are produced by Streptomyces ghanaensis and related organisms, are phosphoglycolipid antibiotics that target peptidoglycan glycosyltransferases that are involved in bacterial cell wall biosynthesis. Transporter proteins are present within the cluster [443,444]. Friulimicin is a lipopeptide antibiotic that is produced by Actinoplanes friuliensis. Transporter proteins within the cluster were reported to be involved in the self-resistance [445]. Bleomycin that is produced by Streptomyces verticillus is a DNA synthesis inhibitor and has been used for cancer chemotherapy. Two resistant determinants, blmA and blmB, were isolated encoding an acetyltransferase and the bleomycin-binding protein, respectively [446]. Zorbamycin is a member of the bleomycin family glycopeptide anti-tumor antibiotic. In contrast to those of bleomycin and tallysomycin, another bleomycin family antibiotic, zorbamycin producer S. flavoviris, lacks the N-acetyltransferase, and the zorbamycin-binding protein is sufficient to confer resistance in the producing bacteria [447].
β-Lactam antibiotics, including penicillins and cephalosporins, are the most commonly used antibiotics, although they have been used for almost one century. β-Lactam antibiotics are classified into five groups, namely: penicillins, cephalosporins/cephamycins, clavulanic acid, thienamycin, and nocardicin A and sulfazecin. Penicillin is the first antibiotic that has been isolated as a natural secondary metabolite [448]. Penicillins and cephalosporins/cephamycins are produced by bacteria, as well as fungi, while others are produced by bacteria [449,450]. The gene clusters for the biosyntheses of penicillins/cephalosporins/cephamycins were cloned from Streptomyces clavuligerus [451,452] (GB No. CM000913), S. cattleya [453] (GB No. FQ859185), Nocardia lactamdurans [454,455], Lysobacter lactamgenus [456] (GB No. X56660), Penicillium chrysogenum [457] (GB No. AM920436), and Aspergillus nidulans [458] (GB Nos. AH000059 and X54853). The genes for β-lactamases and penicillin-binding proteins, which are involved in the self-resistance in bacteria [459,460,461], are present within these clusters of bacteria, whereas they are absent in those of the fungi. Clavulanic acid is an inhibitor of various kinds of β-lactamases from pathogenic bacteria and was isolated from S. clavuligerus [462]. It is used in combination with β-lactam antibiotics. The gene cluster for the biosynthesis of clavulanic acid is located between the cephamycin gene cluster and penicillin-binding protein, and β-lactamase genes [452,463,464]. A comparison of the cephamycin gene clusters of S. clavuligerus and S. cattleya, a clavulanic acid-non-producer, indicates that the clavulanic acid gene cluster is inserted between the cephamycin gene cluster and penicillin-binding protein/β-lactamase genes, without affecting the presence of the penicillin-binding protein and β-lactamase genes, suggesting that the penicillin-binding proteins and the β-lactamases play important roles in the protection from cephamycin, but not from clavulanic acid in the producer. However, the precise role of penicillin-binding protein and β-lactamase genes in clavulanic acid biosynthesis remains to be elucidated [465].
Thienamycin is the progenitor natural product of the broad-spectrum carbapenem antibiotics [466]. The gene cluster for the biosynthesis of thienamycin is located in the plasmid of S. cattleya [467,468] (GB No. AJ421798). There are three genes that are involved in self-resistance within the cluster, that is, thnF, thnJ, and thnS, by encoding N-acetyltransferase, transport protein, and β-lactamase, respectively [467]. In addition, the thnC encoding efflux pump may be implicated in the resistance. Nocardicin A is a monocyclic β-lactam antibiotic monobactam, and was isolated from Nocardia uniformis [469] and other actinomycetes. It shows moderate activity against Gram-negative bacteria and exhibits some β-lactamase resistance. The biosynthetic gene cluster of nocardicin A was cloned [470] (GB No. AY541063). Acetyltransferase (NocD) and the transporter protein (NocH) were proposed to be involved in self-resistance. In accord with the β-lactamase resistance, the β-lactamase gene was deficient. Another monobactam antibiotic sulfazecin was isolated from Pseudomonas acidophila. It is active against Gram-negative bacteria [471] and is not inactivated by metallo-β-lactamases, which renders bacteria with extended-spectrum β-lactam resistance. The gene cluster contains several transporter genes, a β-lactamase gene, and the multidrug transporter gene mdtB, which may be involved in the self-resistance [472] (GB No. KX757706). The exact role of β-lactamase remains to be clarified.
Viomycin is a member of the tuberactinomycin family of antibiotics, which are peptide antibiotics containing nonprotenogenic amino acids. They are essential drugs against Mycobacterium tuberculosis. The viomycin biosynthetic gene cluster was cloned and sequenced. The vph gene encoding viomycin phosphotransferase is involved in the self-resistance [473]. Pristinamycin that is produced by Streptomyces pristinaespiralis is a streptogramin group antibiotic. The DNA fragment of 120 kb covers the pristinamycin-specific genes for the biosynthesis, regulation, and resistance of pristinamycin, although multidrug resistance gene ptr is located outside the 210 kb supercluster [474,475]. Lincomycin and celesticetin are lincosamide antibiotics. Resistance is usually encountered in the form of the MLS phenotype, which includes macrolides, lincosamides, and streptogramin B type antibiotics [476]. The clr gene product, rRNA methyltransferase, is involved in self-resistance [477]. Daunorubisin and doxorubin are clinically important anthracycline antitumor antibiotics that are isolated from Streptomyces peucetius. Four proteins were reported to be involved in self-resistance. DrrA and DrrB proteins form an ATP cassette transporter/antiporter system [478]. The DrrC protein is a DNA-binding protein, like an UvrA-like protein [479]. The DrrD protein may function as oxygen oxidoreductase, like McrA in mitomycin C resistance [480]. Chromomycin A3 is an aureolic acid group antitumor antibiotic that is produced by Streptomyces griseus. The biosynthetic gene cluster contains three genes that are involved in the self-resistance. The cmrA and cmrB genes encode the ABC transporters, and cmrX encodes a UvrA-like protein of UV repair nuclease [481]. Mithramycin is another aureolic acid type antitumor antibiotic that is produced by the Streptomyces species [482]. The biosynthetic gene cluster of mithramycin was cloned [483] (GB No. X89899). The three genes (mtrX, mtrA and mtrB) that are located at the end of the cluster were proposed to be involved in the self-resistance, two of which encode the ABC transporters. Novobiocin is a member of the aminocoumarin type of antibiotics, which include coumermycin A1 and clorobiocin. The novobiocin biosynthetic gene cluster contains two self-resistance genes, novA and gyrB. The former encodes the ABC transporter and the latter encodes novobiocin-resistant gyrase subunit B [484] (GB No. AF170880). The biosynthetic gene clusters of coumermycin A1 and clorobiocin were also cloned and sequenced [485,486]. Mitomycin C is an antitumor antibiotic that is produced by Streptomyces lavendulae. Two self-resistance proteins were reported, one is oxygen oxidoreductase (Mcr) and the other is mitomycin-binding protein (Mrd) [487,488]. Yatakemycin is an antitumor antibiotic belonging to the family of CC-1065, and duocarmycin, which are produced by Streptomyces species. They are DNA-alkylating agents. The biosynthetic gene cluster contains five self-resistance-related genes including DNA glycosylase, DNase, and the transporter [489,490]. Natamycin is a polyene macrolide antifungal antibiotic. ABC transporters are involved in self-resistance [491]. Capuramycin is a nucleoside antibiotic that is isolated from the Amycolatopsis species, which inhibits bacterial translocase I that is involved in peptidoglycan cell wall biosynthesis. The gene cluster for the biosynthesis of capuramycin was cloned [492] (GB No. KP995196). The phosphotransferase gene capP, located at the end of the cluster, is implicated in the self-resistance. A-500359s are nucleoside antibiotics inhibiting phosphor-N-acetylmuramyl-pentapeptide translocase. A phospho-transferase is involved in the self-resistance [493]. Laspartomycin is a lipopeptide antibiotic that is produced by Streptomyces viridochromogenes [494]. The gene cluster was cloned [495]. Three transporters were proposed to be involved in the self-resistance. Platensimycin (PTM) and platencin (PTN) are bacterial fatty acid synthase inhibitors that are produced by Streptomyces platensis. PtmP3/PtnP3 and FabF proteins confer PTM and PTN self-resistance by target replacement and target modification [496]. d-Cycloserine is an anti-tubercular antibiotic that is produced by Streptomyces lavendulae. The biosynthetic gene cluster was cloned [497]. The d-alanyl-d-alanine ligase DcsI and a membrane protein, DcsJ, are involved in self-resistance. Fosfomycin has a unique chemical structure, containing a carbon-phosphorus and an epoxide. It inhibits peptidoglycan biosynthesis. The FomA and FomB proteins confer self-resistance on the producer organism by the phosphorylation of fosfomycin and fosfomycin monophosphate, respectively [498]. The rifamycins are broad-spectrum antibiotics that inhibit bacterial RNA polymerase. Several self-resistance mechanisms have been reported, including RNA polymerase modification, glycosylation, phosphorylation, and transporters [499,500]. Thiostrepton, micrococcin, nosiheptide, promothiocin, and promoinducin are ribosomally produced thiopeptide antibiotics that are produced by the Streptomyces, Bacillus, and Micrococcus species. The mutation of the ribosomal protein L11 is the mechanism of self-resistance [501], and rRNA methylation may be also involved [502].
Neocarzinostatin is an enediyne type antitumor antibiotic that is synthesized by Streptomyces carzinostaticus in the form of a chromoprotein complex. The amino acid residues D33 and D99 of the apoprotein play significant roles for self-protection. In addition, the neocarzinostatn carrier protein and mycothiol-dependent cellular detoxication are also important [503]. Calicheamicin is the non-chromoprotein enediyne type of antitumor antibiotic that is produced by Micromonospora echinospora. The biosynthetic gene cluster was cloned [504]. The non-heme iron metalloprotein CalC within the cluster is participated in the self-resistance [505] (GB No. AF497482). CalU16 and CalU19 are reported to be structural homologues of CalC. Kedarcidin is another enediyne type chromoprotein antitumor antibiotic that is isolated from the Streptoalloteichus species [506]. The kedA, kedX, and kedX2 genes are involved in self-resistance, encoding apoprotein, efflux pump, and efflux pump, respectively [507]. Cyanosporasides, sporolides, and fijiolides are postulated to represent spontaneous enediyne degradation products. Cyanosporasides were isolated from the marine actinomycetes, Salinispora pacifica and Streptomyces, species. The biosynthetic gene clusters were cloned. It contains a couple of transporters and resistance proteins [508]. Salinosporamide A is a proteasome inhibitor that is isolated from the marine bacterium, Salinispora tropica. It shows anti-leukemic activity. The mutation of the 20S proteasome β-subunit confers self-resistance on the producer bacterium [509] (GB Nos. CP000667; EF397502). Miklamicin is a spirotetronate type of antibiotic that is produced by the endophytic Micromonospora sp. Three transporter proteins are present within the biosynthetic gene cluster [510] (GB No. LC021382). Microbisporicin is a lantibiotic antibiotic that is produced by Microbispora coralline. The biosynthetic gene cluster was cloned, which contains a couple of transporter proteins [511] (GB No. HM536998).
The self-resistance genes are also detected in Gram-positive Bacillus and the related species producing surfactin [512,513], subtilin [514], sublancin [515], zwittermicin [516,517], bacitracin [518], polymyxin [519] (GB No. EU371992), and edeine [520] (GB No. KC771276), and Gram-negative Alcaligenes, Pseudomonas, and fish pathogen Yersinia species, producing kalimantacin [521,522,523] (GB No. GU479979), pseudomonic acid [524], and holomycin [525], respectively. Holomycin is also produced by Streptomyces clavuligerus [526,527]. Interestingly, the resistance mechanisms are different between the Gram-negative Yersinia species and the Gram-positive Streptomyces species [525,527]. The biocontrol Agrobacterium radiobacter K84 secretes the antibiotic 84 that is selectively transported into the plant pathogen, A. tumefaciens. The mutation of the leucyl-tRNA synthetase (LeuRS) is responsible for self-resistance [528]. The old antifungal antibiotic griseofulvin and an immunosuppressant drug, mycophenolic acid, were isolated from the Penicillium species. The transporter and IMP dehydrogenase are involved in self-resistance [529,530,531] (GB No. HQ731031). From these results, it is concluded that the antibiotic producers use more sophisticated resistance mechanisms than other organisms, such as rRNA methylation (modification of target), modification and detoxication of antibiotics by acetylation; phosphorylation and adenylylation; chemical degradation by, for example, β-lactamase; excretion of antibiotics by efflux pump; modification of target (e.g., tRNA synthetase); and so on. Table 5 shows the antibiotic resistance-related strategies in bacteria and fungi. The early stage and more recent researches on the antibiotic self-resistance were reviewed by Demain [532], Vining [533], Cundliffe [534], and Cundliffe and Demain [535].
8. Conclusions
Drug resistance, especially antibiotic resistance, is getting worse and worse. Antibiotic resistance genes (ARGs) are complex mixtures of the genes of intrinsic antibiotic resistance [536,537] and acquired resistance [538], and constitute resistome [369,370,371,372]. Resistome is composed of the ARGs from antibiotic producers and pathogenic bacteria, and cryptic genes and precursor genes. These genes interconnect with each other very closely by horizontal gene transfer and adaptive mutations. Moreover, resistome is now identified in almost every microbial community, including soil, activated sludge, human gut and oral microbiomes, and animal gut microbiomes [539,540]. On the other hand, it is now extremely difficult to find an effective antibiotic against resistant pathogenic bacteria, although every effort has been taken to discover new antibiotics [541,542,543]. One of the steps to solve this problem is to know precisely the mechanisms of drug resistance in the various kingdoms once more. This review compares the molecular mechanisms underpinning the self-resistance against phycotoxins, toxins from marine and terrestrial animals, plants, and fungi and antibiotics. The results show that each kingdom possesses the characteristic features, as follows: transporters/efflux pumps in phycotoxins, mutation of targets and sequestration in marine and terrestrial animal toxins, ABC transporters and sequestration in plant toxins, transporters in fungal toxins, and various mechanism in antibiotics, indicating that antibiotic producers make tremendous efforts for avoiding suicide by using an enormous array of strategies and are more flexible and adaptable to the changes of environments (Table 1, Table 2, Table 3, Table 4 and Table 5).
Self-resistance or self-defense is one of the conclusions after the long history of evolution and adaptation to the environment, where desperate struggles were experienced between predators and prey species for survival. The typical example is seen in tetrodotoxin. It is distributed in taxonomically diverse groups, from bacteria such as actinobacteria, bacteroides, firmicutes, and proteobacteria [140,141], to pufferfish, snakes, newts, and other animals [9,73]. Although the self-resistance mechanism to tetrodotoxin has not been explored in bacteriawhich, in animals, is as a result of the mutation of only a few restricted residues of the target, Nav (Figure 1). Similarly, the self-resistance mechanisms to toxins in plants are also limited to a few numbers, that is, sequestration and the related transporters. On the other hand, the mechanisms of antibiotic resistance are tremendously complex (Table 5). Antibiotics are thought to be toxins for pathogenic bacteria. For their survival, it is necessary for the pathogenic bacteria to behave as defensive prey against natural secondary metabolites, including antibiotics, as well as synthetic chemicals. Furthermore, compared to animals and plants, bacteria are more flexible to genetic variations, as described above. So it is extremely challenging to manage and overcome each of the mechanisms of the antibiotic resistance.
The relation between pathogenic bacteria and humans/livestock is able to speak, figuratively, as that between predators and prey, or herbivores and plants (Figure 6). Arbuckle et al. described three types of gene-product based resistance, namely, toxin scavenging, target-site insensitivity, and off-target repurposing [9]. Immunological molecules correspond to toxin scavenging, and the pseudo-receptors to pathogenic bacteria play similar roles in target-site insensitivity, and the receptors with higher affinity to pathogenic bacteria, but no signaling ability, are candidates as off-target reprograming. These molecules, if possible, may act as significant functions for overcoming the drug resistance. As for the relationship between herbivores and plants, adaptation and sequestration in specific organs can be considered. To attain the adaptability to pathogenic bacteria in humans/livestock, some immunological activation is needed. Sequestration in specific organs of toxins, their non-toxic precursors, and enzymes to activate precursors, may be a dream, but I would expect these situations, considering recent technological developments.
With these features in mind, potential alternative strategies to overcome these resistance mechanisms are investigated in the following paragraph. Firstly, improvement of the directions of antibiotic usage. It is necessary for physicians to use narrow-spectrum, but not broad-spectrum, appropriate antibiotics in the right amounts, at the right time, and to identify the viral or bacterial pathogen precisely in a strain level, but not in the species level. Secondly, the innovation not to disseminate the resistance bacteria and resistance genes. Antibiotics are able to be prescribed coupled with bacteriophage [544,545], monoclonal antibody [546,547], and vaccines [548,549], or to be replaced with these approaches, if possible. Toxins are pivotal for preventing bacterial infection, when the activity of antitoxin is properly controlled [550,551]. The innate immune system, consisting of the immediate activation of the pathogen non-specific innate immunity, and following the activation of adaptive immune responses, is the last line of defense against the infectious diseases [552,553]. Lastly, the exploration of new compounds. Transporters or efflux pumps are the most fundamental and prevailing strategies for self-resistance and self-defense in every kingdom. So, inhibitors of transporters/efflux pumps are hopeful candidates for the prevention of infectious diseases. Together with the activation of silent genes, the combinatorial application of synthetic biological technology with genomic, metagenomics, and functional analyses of marine and terrestrial invertebrates, plants, and microbes will open the revival of the new golden era of natural products [554]. In fact, discoveries of teixobactin [555], compound 10 [556], and others [557], offer promising possibilities for a bright future.
Founding
APC was sonspred by MDPI.
Acknowledgments
I would like to devote this review paper to the late Prof. Hamao Umezawa, the Former director of the Department of Antibiotics, at the National Institute of Health of Japan (now reorganized to the National Institute of Infectious Diseases), for introducing me to the antibiotic field.
Conflicts of Interest
The author declares no conflict of interest.
References
- Walsh, C.T.; Wencewicz, T.A. Prospects for new antibiotics: A molecule-centered perspective. J. Antibiot. 2014, 67, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Courvalin, P. Why is antibiotic resistance a deadly emerging disease? Clin. Microbiol. Infect. 2016, 22, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L. General principles of antibiotic resistance in bacteria. Drug Discov. Today Technol. 2014, 11, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Gomes, K.M.; Duarte, R.S.; de Freire Bastos, M.D. Lantibiotics produced by Actinobacteria and their potential applications (a review). Microbiology 2017, 163, 109–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaou, K.C.; Rigol, S. A brief history of antibiotics and select advances in their synthesis. J. Antibiot. 2018, 71, 153–184. [Google Scholar] [CrossRef] [PubMed]
- Simpkin, V.L.; Renwick, M.J.; Kelly, R.; Mossialos, E. Incentivising innovation in antibiotic drug discovery and development: Progress, challenges and next steps. J. Antibiot. 2017, 70, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Young, K.; Silver, L.L. What is an “ideal” antibiotic? Discovery challenges and path forward. Biochem. Pharmacol. 2017, 133, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xue, Y.; Gao, W.; Li, J.; Zu, X.; Fu, D.; Feng, S.; Bai, X.; Zuo, Y.; Li, P. Actinobacteria-derived peptide antibiotics since 2000. Peptides 2018, 103, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, K.; Rodriguez de la Vega, R.C.; Casewell, N.R. Coevolution takes the sting out of it: Evolutionary biology and mechanisms of toxin resistance in animals. Toxicon 2017, 140, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Brodie, E.D., III; Brodie, E.D., Jr. Predictably convergent evolution of sodium channels in the arms race between predators and prey. Brain Behav. Evol. 2015, 86, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C.R.; Durso, A.M.; Hanifin, C.T.; Pfrender, M.E.; Ducey, P.K.; Stokes, A.N.; Barnett, K.E.; Brodie, E.D., III; Brodie, E.D., Jr. Is there more than one way to skin a newt? Convergent toxin resistance in snakes is not due to a common genetic mechanism. Heredity 2016, 116, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Hollis, K.L. Ants and antlions: The impact of ecology, coevolution and learning on an insect predator-prey relationship. Behav. Processes 2017, 139, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Johannessen, T. From Antagonistic to a Synergistic Predator Prey Perspective: Bifurcations in Marine Ecosystems; Academic Press: Cambridge, MA, USA, 2014; ISBN 978-0-12-417016-2. [Google Scholar]
- Breed, M.D.; Moore, J. Self-Defense. In Animal Behavior, 2nd ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 326–355. ISBN 978-0-12-801532-2. [Google Scholar]
- Cory, J.S. Evolution of host resistance to insect pathogens. Curr. Opin. Insect Sci. 2017, 21, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Morehouse, N.I.; Relyea, R. Pesticide tolerance in amphibians: Induced tolerance in susceptible populations, constitutive tolerance in tolerant populations. Evol. Appl. 2013, 6, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Weinberger, H.; Sullivan, J.C.; Reitzel, A.M.; Finnerty, J.R.; Gurevitz, M. Concerted evolution of sea anemone neurotoxin genes is revealed through analysis of the Nematostella vectensis genome. Mol. Biol. Evol. 2008, 25, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Sirikantaramas, S.; Yamazaki, M.; Saito, K. Mechanisms of resistance to self-produced toxic secondary metabolites in plants. Phytochem. Rev. 2008, 7, 467–477. [Google Scholar] [CrossRef]
- Tarvin, R.D.; Santos, J.C.; O’Connell, L.A.; Zakon, H.H.; Cannatella, D.C. Convergent substitutions in a sodium channel suggest multiple origins of toxin resistance in poison frogs. Mol. Biol. Evol. 2016, 33, 1068–1081. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K.; Sugiyama, A.; Morita, M.; Shitan, N. Secondary transport as an efficient membrane transport mechanism for plant secondary metabolites. Phytochem. Rev. 2008, 7, 513–524. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wuester, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Dettner, K. Chemical defense and toxins of lower terrestrial and freshwater animals. In Comprehensive Natural Products II; Elsevier: New York, NY, USA, 2010; Volume 4, pp. 387–410. ISBN 978-0-12-409547-2. [Google Scholar]
- Sahebi, M.; Hanafi, M.M.; van Wijnen, A.J.; Akmar, A.S.N.; Azizi, P.; Idris, A.S.; Taheri, S.; Foroughi, M. Profiling secondary metabolites of plant defence mechanisms and oil palm in response to Ganoderma boninense attack. Internat. Biodeterior. Biodegrad. 2017, 122, 151–164. [Google Scholar] [CrossRef]
- Walton, K.; Berry, J.P. Indole alkaloids of the Stigonematales (Cyanophyta): Chemical diversity, biosynthesis and biological activity. Mar. Drugs 2016, 14, 73. [Google Scholar] [CrossRef] [PubMed]
- Ajani, P.; Harwood, D.T.; Murray, S.A. Recent trends in marine phycotoxins from Australian coastal waters. Mar. Drugs 2017, 15, E33. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.A.; Andersen, A.J.; Andersen, N.G.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Chemical diversity, origin, and analysis of phycotoxins. J. Nat. Prod. 2016, 79, 662–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, L.A.; Gautam, S.; Kumar, S. Phycotoxins. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: New York, NY, USA, 2014; pp. 25–29. ISBN 978-0-08-100596-5. [Google Scholar]
- Nedbalova, L.; Strizek, A.; Sigler, K.; Rezanka, T. Effect of salinity on the fatty acid and triacylglycerol composition of five haptophyte algae from the genera Coccolithophora, Isochrysis and Prymnesium determined by LC-MS/APCI. Phytochemistry 2016, 130, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Higashi, A.; Fujitani, Y.; Nakayama, N.; Tani, A.; Ueki, S. Selective growth promotion of bloom-forming raphidophyte Heterosigma akashiwo by a marine bacterial strain. Harmful Algae 2016, 60, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Luckas, B.; Krueger, K.; Roeder, K. Phycotoxins and food safety. In Chemical Contaminants and Residues in Food, 2nd ed.; Schrenk, D., Cartus, A., Eds.; Elsevier: New York, NY, USA, 2017; pp. 337–378. ISBN 978-0-08-100674-0. [Google Scholar]
- Kalaitzis, J.A.; Chau, R.; Kohli, G.S.; Murray, S.A.; Neilan, B.A. Biosynthesis of toxic naturally-occurring seafood contaminants. Toxicon 2010, 56, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Rein, K.S.; Snyder, R.V. The biosynthesis of polyketide metabolites by dinoflagellates. Adv. Appl. Microbiol. 2006, 59, 93–123. [Google Scholar] [PubMed]
- O’Neill, K.; Musgrave, I.F.; Humpage, A. Low dose extended exposure to saxitoxin and its potential neurodevelopmental effects: A review. Environ. Toxicol. Pharm. 2016, 48, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef] [PubMed]
- Monroe, E.A.; Van Dolah, F.M. The toxic dinoflagellate Karenia brevis encodes novel type I-like polyketide synthases containing discrete catalytic domains. Protist 2008, 159, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Monroe, E.A.; Johnson, J.G.; Wang, Z.; Pierce, R.K.; Van Dolah, F.M. Characterization and expression of nuclear-encoded polyketide synthases in the brevetoxin-producing dinoflagellate. J. Phycol. 2010, 46, 541–552. [Google Scholar] [CrossRef]
- Suikkanen, S.; Kremp, A.; Hautala, H.; Krock, B. Paralytic shellfish toxins or spirolides? The role of environmental and genetic factors in toxin production of the Alexandrium ostenfeldii complex. Harmful Algae 2013, 26, 52–59. [Google Scholar] [CrossRef]
- Eichholz, K.; Beszteri, B.; John, U. Putative monofunctional type I polyketide synthase units: A dinoflagellate-specific feature? PLoS ONE 2012, 7, e48624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, U.; Beszteri, B.; Derelle, E.; Van de Peer, Y.; Read, B.; Moreau, H.; Cembella, A. Novel insights into evolution of protistan polyketide synthases through phylogenomic analysis. Protist 2008, 159, 21–30. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Lambert, G.; Andersen, R.A.; Coffroth, M.A.; Galbraith, D.W. Symbiodinium (pyrrhophyta) genome sizes (DNA content) are smallest among dinoflagellates. J. Phycol. 2005, 41, 880–886. [Google Scholar] [CrossRef]
- Spector, D.L. Dinoflagellate nuclei. In Dinoflagellates; Spector, D.L., Ed.; Academic Press: Cambridge, MA, USA, 1984; pp. 107–147. ISBN 0-12-656520-1. [Google Scholar]
- Casabianca, S.; Cornetti, L.; Capellacci, S.; Vernesi, C.; Penna, A. Genome complexity of harmful microalgae. Harmful Algae 2017, 63, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, D.L.; Hemscheidt, T.K.; Moore, R.E.; Runnegar, M.T.C. Biosynthesis of cylindrospermopsin. J. Org. Chem. 2000, 65, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Mejean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-eicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar] [CrossRef] [PubMed]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Stueken, A.; Jakobsen, K.S. The cylindrospermopsin gene cluster of Aphanizomenon sp. strain 10E6: Organization and recombination. Microbiology 2010, 156, 2438–2451. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.J.; Marquez, B.L.; Nogle, L.M.; McPhail, K.; Goeger, D.E.; Roberts, M.A.; Gerwick, W.H. Structure and biosynthesis of the jamaicamides, new mixed polyketide-peptide neurotoxins from the marine cyanobacterium Lyngbya majuscule. Chem. Biol. 2004, 11, 817–833. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.C.; Gerwick, L.; Gonzalez, D.; Dorrestein, P.C.; Gerwick, W.H. Transcriptional analysis of the jamaicamide gene cluster from the marine cyanobacterium Lyngbya majuscula and identification of possible regulatory proteins. BMC Microbiol. 2009, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Marquez, B.L.; Watts, K.S.; Yokochi, A.; Roberts, M.A.; Verdier-Pinard, P.; Jimenez, J.I.; Hamel, E.; Scheuer, P.J.; Gerwick, W.H. Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J. Nat. Prod. 2002, 65, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, A.V.; Sorrels, C.M.; Gerwick, W.H. Cloning and biochemical characterization of the hectochlorin biosynthetic gene cluster from the marine cyanobacterium Lyngbya majuscule. J. Nat. Prod. 2007, 70, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Sitachitta, N.; Rossi, J.V.; Roberts, M.A.; Flatt, P.M.; Jia, J.; Sherman, D.H.; Gerwick, W.H. Biosynthetic pathway and gene cluster analysis of curacin A, an antitubulin natural product from the tropical marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2004, 67, 1356–1367. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.C.; Monroe, E.A.; Podell, S.; Hess, W.R.; Klages, S.; Esquenazi, E.; Niessen, S.; Hoover, H.; Rothmann, M.; Lasken, R.S.; et al. Genomic insights into the physiology and ecology of the marine filamentous cyanobacterium Lyngbya majuscula. Proc. Natl. Acad. Sci. USA 2011, 24, 8815–8820. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Iinuma, Y.; Kobayashi, J. Cloning of polyketide synthase genes from amphidinolide-producing dinoflagellate amphidinium sp. Biol. Pharm. Bull. 2006, 29, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Law, B.K.; Luesch, H. Apratoxin A reversibly inhibits the secretory pathway by preventing cotranslational translocation. Mol. Pharmacol. 2009, 76, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Grindberg, R.V.; Ishoey, T.; Brinza, D.; Esquenazi, E.; Coates, R.C.; Liu, W.; Gerwick, L.; Dorrestein, P.C.; Pevzner, P.; Lasken, R.; et al. Single cell genome amplification accelerates identification of the apratoxin biosynthetic pathway from a complex microbial assemblage. PLoS ONE 2011, 6, e18565. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.J.; Gerwick, W.H. Lyngbyatoxin biosynthesis: Sequence of biosynthetic gene cluster and identification of a novel aromatic prenyltransferase. J. Am. Chem. Soc. 2004, 126, 11432–11433. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Ottilie, S.; Eustaquio, A.S.; Edwards, D.J.; Gerwick, L.; Moore, B.S.; Gerwick, W.H. Evaluation of Streptomyces coelicolor A3(2) as a heterologous expression host for the cyanobacterial protein kinase C activator lyngbyatoxin A. FEBS J. 2012, 279, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Wells, K.N.; Singh, A.J.; Gerwick, W.H.; Philmus, B. Assessment of Anabaena sp. strain PCC 7120 as a heterologous expression host for cyanobacterial natural products: Production of lyngbyatoxin A. ACS Synth. Biol. 2016, 5, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Zhang, L.; Wakimoto, T.; Hoshino, S.; Mori, T.; Ito, T.; Ishikawa, J.; Tanner, M.E.; Abe, I. A methyltransferase initiates terpene cyclization in teleocidin B biosynthesis. J. Am. Chem. Soc. 2014, 136, 9910–9913. [Google Scholar] [CrossRef] [PubMed]
- Fontanillo, M.; Koehn, M. Microcystins: Synthesis and structure–activity relationship studies toward PP1 and PP2A. Bioorg. Med. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef] [PubMed]
- Kaebernick, M.; Dittmann, E.; Boerner, T.; Neilan, B.A. Multiple alternate transcripts direct the biosynthesis of microcystin, a cyanobacterial nonribosomal peptide. Appl. Environ. Microbiol. 2002, 68, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Doehren, H.; Boerner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Pearson, L.A.; Hisbergues, M.; Boerner, T.; Dittmann, E.; Neilan, B.A. Inactivation of an ABC transporter gene, mcyH, results in loss of microcystin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Appl. Environ. Microbiol. 2004, 70, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Carmichae, W.W.; Kennelly, P.J. Cyanobacterial PPP family protein phosphatases possess multifunctional capabilities and are resistant to microcystin-LR. J. Biol. Chem. 1999, 274, 10039–10046. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Tri, P.H.; Hung, N.V.; Krunic, A.; Shim, S.H.; Soejarto, D.D.; Orjala, J. Nhatrangins A and B, aplysiatoxin-related metabolites from the marine cyanobacterium Lyngbya majuscula from Vietnam. J. Nat. Prod. 2010, 73, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Soto-Liebe, K.; Murillo, A.A.; Krock, B.; Stucken, K.; Fuentes-Valdes, J.J.; Trefault, N.; Cembella, A.; Vasquez, M. Reassessment of the toxin profile of Cylindrospermopsis raciborskii T3 and function of putative sulfotransferases in synthesis of sulfated and sulfonated PSP toxins. Toxicon 2010, 56, 1350–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lera Ruiz, M.; Kraus, R.L. Voltage-gated sodium channels: Structure, function, pharmacology, and clinical indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef] [PubMed]
- Bricelj, V.M.; Connell, L.; Konoki, K.; MacQuarrie, S.P.; Scheuer, T.; Catterall, W.A.; Trainer, V.L. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Soong, T.W.; Venkatesh, B. Adaptive evolution of tetrodotoxin resistance in animals. Trends Genet. 2006, 22, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [PubMed]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Liebe, K.; Lopez-Cortes, X.A.; Fuentes-Valdes, J.J.; Stucken, K.; Gonzalez-Nilo, F.; Vasquez, M. In silico analysis of putative paralytic shellfish poisoning toxins export proteins in cyanobacteria. PLoS ONE 2013, 7, e55664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stucken, K.; John, U.; Cembella, A.; Murillo, A.A.; Soto-Liebe, K.; Fuentes-Valdes, J.J.; Friedel, M.; Plominsky, A.M.; Vasquez, M.; Gloeckner, G. The smallest known genomes of multicellular and toxic cyanobacteria: Comparison, minimal gene sets for linked traits and the evolutionary implications. PLoS ONE 2010, 5, e9235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stueken, A.; Orr, R.J.S.; Kellmann, R.; Murray, S.A.; Neilan, B.A.; Jakobsen, K.S. Discovery of nuclear-encoded genes for the neurotoxin saxitoxin in dinoflagellates. PLoS ONE 2011, 6, e20096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, T.; Tsuchiya, T. Multidrug efflux transporters in the MATE family. Biochim. Biophys. Acta 2009, 1794, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, J.; Minor, D.L., Jr. Bacterial voltage-gated sodium channels (BacNaVs) from the soil, sea, and salt lakes enlighten molecular mechanisms of electrical signaling and pharmacology in the brain and heart. J. Mol. Biol. 2015, 427, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Navarro, B.; Xu, H.; Yue, L.; Shi, Q.; Clapham, D.E. A prokaryotic voltage-gated sodium channel. Science 2001, 294, 2372–2375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ren, W.; DeCaen, P.; Yan, C.; Tao, X.; Tang, L.; Wang, J.; Hasegawa, K.; Kumasaka, T.; He, J.; et al. Crystal structure of an orthologue of the NaChBac voltage-gated sodium channel. Nature 2012, 486, 130–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, J.D.; Wisecaver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of saxitoxin synthesis in cyanobacteria and dinoflagella. Mol. Biol. Evol. 2013, 30, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS ONE 2009, 4, e5758. [Google Scholar] [CrossRef] [PubMed]
- Orr, R.J.S.; Stuken, A.; Murray, S.A.; Jakobsen, K.S. Evolutionary acquisition and loss of saxitoxin biosynthesis in dinoflagellates: The second “core” gene, sxtG. Appl. Environ. Microbiol. 2013, 79, 2128–2136. [Google Scholar] [CrossRef] [PubMed]
- Orr, R.J.S.; Stuken, A.; Murray, S.A.; Jakobsen, K.S. Evolution and distribution of saxitoxin biosynthesis in dinoflagellates. Mar. Drugs 2013, 11, 2814–2828. [Google Scholar] [CrossRef] [PubMed]
- Araoz, R.; Molgo, J.; de Marsac, N.T. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Mejean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence that biosynthesis of the neurotoxic alkaloids anatoxin-a and homoanatoxin-a in the cyanobacterium Oscillatoria PCC 6506 occurs on a modular polyketide synthase initiated by L-proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [Google Scholar] [CrossRef] [PubMed]
- Mejean, A.; Paci, G.; Gautier, V.; Ploux, O. Biosynthesis of anatoxin-a and analogues (anatoxins) in cyanobacteria. Toxicon 2014, 91, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Rantala-Ylinen, A.; Kana, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.; Dave, A.; MacKay, J.A.; Rawal, V.H. The chemistry of hapalindoles, fischerindoles, ambiguines, and welwitindolinones. In The Alkaloids: Chemistry and Biology; Knoelker, H.-J., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 73, pp. 65–160. ISBN 978-0-12-411565-1. [Google Scholar]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Zilfou, J.T.; Stratmann, K.; Patterson, G.M.L.; Moore, R.E. Welwitindolinone analogues that reverse P-glycoprotein-mediated multiple drug resistance. Mol. Pharmacol. 1995, 47, 241–247. [Google Scholar] [PubMed]
- Hillwig, M.L.; Fuhrman, H.A.; Ittiamornkul, K.; Sevco, T.J.; Kwak, D.H.; Liu, H. Identification and characterization of a welwitindolinone alkaloid biosynthetic gene cluster in stigonematalean cyanobacterium Hapalosiphon welwitschii. ChemBioChem 2014, 15, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Hillwig, M.L.; Zhu, Q.; Liu, X. Biosynthesis of ambiguine indole alkaloids in cyanobacterium Fischerella ambigua. ACS Chem. Biol. 2014, 9, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.L.; Sharma, D.; Bunn, B.M.; Gerwick, L.; Viswanathan, R.; Moffitt, M.C. Comparative analysis of hapalindole, ambiguine and welwitindolinone gene clusters and reconstitution of indole-isonitrile biosynthesis from cyanobacteria. BMC Microbiol. 2014, 14, 213. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Vetter, I.; Eisenblaetter, A.; Krock, B.; Ebbecke, M.; Desel, H.; Zimmermann, K. Ciguatera fish poisoning: A first epidemic in Germany highlights an increasing risk for European countries. Toxicon 2014, 91, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cataldi, M. Ciguatoxin. Ref. Modul. Biomed. Sci. 2016, 1. [Google Scholar] [CrossRef]
- Shmukler, Y.B.; Nikishin, D.A. Ladder-shaped ion channel ligands: Current state of knowledge. Mar. Drugs 2017, 15, 232. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xu, J.; Sinkkonen, J.; Wu, J. Polyketides from marine dinoflagellates of the genus Prorocentrum, biosynthetic origin and bioactivity of their okadaic Acid Analogues. Mini-Rev. Med. Chem. 2010, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Sattely, E.S.; Fischbach, M.A.; Walsh, C.T. Total biosynthesis: In vitro reconstitution of polyketide and nonribosomal peptide pathways. Nat. Prod. Rep. 2008, 25, 757–793. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Satake, M.; Wright, J.L.C. Polyketide biosynthesis in dinoflagellates: What makes it different? Nat. Prod. Rep. 2014, 31, 1101–1137. [Google Scholar] [CrossRef] [PubMed]
- Leadlay, P.F.; Staunton, J.; Oliynyk, M.; Bisang, C.; Cortes, J.; Frost, E.; Hughes-Thomas, Z.A.; Jones, M.A.; Kendrew, S.G.; Lester, J.B.; et al. Engineering of complex polyketide biosynthesis—Insights from sequencing of the monensin biosynthetic gene cluster. J. Ind. Microbiol. Biotechnol. 2001, 27, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Oliynyk, M.; Stark, C.B.W.; Bhatt, A.; Jones, M.A.; Hughes-Thomas, Z.A.; Wilkinson, C.; Oliynyk, Z.; Demydchuk, Y.; Staunton, J.; Leadlay, P.F. Analysis of the biosynthetic gene cluster for the polyether antibiotic monensin in Streptomyces cinnamonensis and evidence for the role of monB and monC genes in oxidative cyclization. Mol. Microbiol. 2003, 49, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, C.J.S.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genom. 2015, 16, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelin, M.; Brovedani, V.; Sosa, S.; Tubaro, A. Palytoxin-containing aquarium soft corals as an emerging sanitary problem. Mar. Drugs 2016, 14, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Gleibs, S.; Mebs, D. Distribution and sequestration of palytoxin in coral reef animals. Toxicon 1999, 37, 1521–1527. [Google Scholar] [CrossRef]
- Hu, T.; LeBlanc, P.; Burton, I.W.; Walter, J.A.; McCarron, P.; Melanson, J.E.; Strangman, W.K.; Wright, J.L.C. Sulfated diesters of okadaic acid and DTX-1: Self-protective precursors of diarrhetic shellfish poisoning (DSP) toxins. Harmful Algae 2017, 63, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.F.; Awan, S.A.; Ling, S.; Wang, R.; Wang, S. Domoic acid: Attributes, exposure risks, innovative detection techniques and therapeutics. Algal Res. 2017, 24, 97–110. [Google Scholar] [CrossRef]
- Clayden, J.; Read, B.; Hebditch, K.R. Chemistry of domoic acid, isodomoic acids, and their analogues. Tetrahedron 2005, 61, 5713–5724. [Google Scholar] [CrossRef]
- Garcia-Altares, M. Structural diversity of microalgal marine toxins. Comprehen. Anal. Chem. 2017, 78, 35–88. [Google Scholar]
- Hodgson, E. Toxins and venoms. Prog. Mol. Biol. Transl. Biol. 2012, 112, 373–415. [Google Scholar]
- Ramsey, U.P.; Douglas, D.J.; Walter, J.A.; Wright, J.L.C. Biosynthesis of domoic acid by the diatom Pseudo-nitzschia multiseries. Nat.Toxins 1998, 6, 137–146. [Google Scholar] [CrossRef]
- Savage, T.J.; Smith, G.J.; Clark, A.T.; Saucedo, P.N. Condensation of the isoprenoid and amino precursors in the biosynthesis of domoic acid. Toxicon 2012, 59, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Boissonneault, K.R.; Henningsen, B.M.; Bates, S.S.; Robertson, D.L.; Milton, S.; Pelletier, J.; Hogan, D.A.; Housman, D.E. Gene expression studies for the analysis of domoic acid production in the marine diatom Pseudo-nitzschia multiseries. BMC Mol. Biol. 2013, 14, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pramod, A.B.; Foster, J.; Carvelli, L.; Henry, L.K. SLC6 transporters: Structure, function, regulation, disease association and therapeutics. Mol. Aspects Med. 2013, 34, 197–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.; Vasconcelos, V. Cyanobactins from cyanobacteria: Current genetic and cemical state of knowledge. Mar. Drugs 2015, 13, 6910–6946. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.A.; Ireland, C.M. Patellamide E: A new cyclic peptide from the ascidian Lissoclinum patella. J. Nat. Prod. 1992, 55, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.A.; Gustafson, K.R.; Cardellina, J.H.; Boyd, M.R. Patellamide F, a new cytotoxic cyclic peptide from the colonial ascidian Lissoclinum patella. J. Nat. Prod. 1995, 58, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.B.; Jacobs, R.S. A marine natural product, patellamide D, reverses multidrug resistance in a human leukemic cell line. Cancer Lett. 1993, 71, 97–102. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [PubMed]
- Koehnke, J.; Bent, A.F.; Houssen, W.E.; Mann, G.; Jaspars, M.; Naismith, J.H. The structural biology of patellamide biosynthesis. Curr. Opin. Struct. Biol. 2014, 29, 112–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czekster, C.M.; Ge, Y.; Naismith, J.H. Mechanisms of cyanobactin biosynthesis. Curr. Opin. Chem. Biol. 2016, 35, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madison, L.L.; Vivas, E.I.; Li, Y.M.; Walsh, C.T.; Kolter, R. The leader peptide is essential for the post-translational modification of the DNA-gyrase inhibitor microcin B17. Mol. Microbiol. 1997, 23, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, C.; Brede, D.A.; Nes, I.F.; Diep, D.B. Circular bacteriocins: Biosynthesis and mode of action. Appl. Environ. Microbiol. 2014, 80, 6854–6862. [Google Scholar] [CrossRef] [PubMed]
- Gatte-Picchi, D.; Weiz, A.; Ishida, K.; Hertweck, C.; Dittmann, E. Functional analysis of environmental DNA-derived microviridins provides new insights into the diversity of the tricyclic peptide family. Appl. Environ. Microbiol. 2014, 80, 1380–1387. [Google Scholar] [CrossRef] [PubMed]
- Onaka, H.; Nakaho, M.; Hayashi, K.; Igarashi, Y.; Furumai, T. Cloning and characterization of the goadsporin biosynthetic gene cluster from Streptomyces sp. TP-A0584. Microbiology 2005, 151, 3923–3933. [Google Scholar] [CrossRef] [PubMed]
- Rebuffat, S. Microcins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 129–137. ISBN 978-0-12-385095-9. [Google Scholar]
- Lee, J.; Cho, Y.-J.; Yang, J.Y.; Jung, Y.-J.; Hong, S.G.; Kim, O.-S. Complete genome sequence of Pseudomonas antarctica PAMC 27494, a bacteriocin-producing psychrophile isolated from Antarctica. J. Biotech. 2017, 259, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-pastor, J.E.; Millan, J.L.S.; Castilla, M.A.; Moreno, F. Structure and organization of plasmid genes required to produce the translation inhibitor microcin C7. J. Bacteriol. 1995, 177, 7131–7140. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Metlitskaya, A.; Severinov, K.; Nair, S.K. Structural basis for microcin C7 inactivation by the Mcce acetyltransferase. J. Biol. Chem. 2011, 286, 21295–21303. [Google Scholar] [CrossRef] [PubMed]
- Severinov, K.; Nair, S.K. Microcin C: Biosynthesis and mechanisms of bacterial resistance. Future Microbiol. 2012, 7, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Yamashita, K.; Goto, Y.; Shimomura, M.; Hayashi, S.; Asamizu, A.; Sugai, Y.; Ikeda, H.; Suga, H.; Onaka, H. Dissection of goadsporin biosynthesis by in vitro reconstitution leading to designer analogues expressed in vivo. Nat. Commun. 2017, 8, 14207. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, V.; Klein, M.L. Small molecule modulation of voltage gated sodium channels. Curr. Opin. Struct. Biol. 2017, 43, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A. Voltage-gated sodium channels at 60: Structure, function and pathophysiology. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef] [PubMed]
- Weir, C.J. Ion channels, receptors, agonists and antagonists. Anaesth. Intensive Care Med. 2016, 17, 645–651. [Google Scholar] [CrossRef]
- Goldin, A.L. Resurgence of sodium channel research. Annu. Rev. Physiol. 2001, 63, 871–894. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neila, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Jal, S.; Khora, S.S. An overview on the origin and production of tetrodotoxin, a potent neurotoxin. J. Appl. Microbiol. 2015, 119, 907–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moczydlowski, E.D. The molecular mystique of tetrodotoxin. Toxicon 2013, 63, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Geffeney, S.; Brodie, E.D., Jr.; Ruben, P.C.; Brodie, E.D., III. Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels. Science 2002, 297, 1336–1339. [Google Scholar] [CrossRef] [PubMed]
- Geffeney, S.L.; Fujimoto, E.; Brodie, E.D., 3rd; Brodie, E.D., Jr.; Ruben, P.C. Evolutionary diversification of TTX-resistant sodium channels in a predator-prey interaction. Nature 2005, 434, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Hanifin, C.T.; Gilly, W.F. Evolutionary history of a complex adaptation: Tetrodotoxin resistance in salamanders. Evolution 2015, 69, 232–244. [Google Scholar] [CrossRef] [PubMed]
- McGlothlin, J.W.; Kobiela, M.E.; Feldman, C.R.; Castoe, T.A.; Geffeney, S.L.; Hanifin, C.T.; Toledo, G.; Vonk, F.J.; Richardson, M.K.; Brodie, E.D., Jr.; et al. Historical contingency in a multigene family facilitates adaptive evolution of toxin resistance. Curr. Biol. 2016, 26, 1616–1621. [Google Scholar] [CrossRef] [PubMed]
- Toledo, G.; Hanifin, C.; Geffeney, S.; Brodie, E.D., III. Convergent evolution of tetrodotoxin-resistant sodium channels in predators and prey. Curr. Top. Membr. 2016, 78, 87–113. [Google Scholar] [PubMed]
- Venkatesh, B.; Lu, S.Q.; Dandona, N.; See, S.L.; Brenner, S.; Soong, T.W. Genetic basis of tetrodotoxin resistance in pufferfishes. Curr. Biol. 2005, 15, 2069–2072. [Google Scholar] [CrossRef] [PubMed]
- Mosher, H.S.; Fuhrman, F.A.; Buchwald, H.D.; Fischer, H.G. Tarichatoxin-tetrodotoxin: A potent neurotoxin. Science 1964, 144, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Croft, J.A.; Howden, M.E.H. Chemistry of maculotoxin: A potent neurotoxin isolated from Hapalochlaena maculosa. Toxicon 1972, 10, 645–651. [Google Scholar] [CrossRef]
- Dao, F.-Y.; Yang, H.; Su, Z.-D.; Yang, W.; Wu, Y.; Ding, H.; Chen, W.; Tang, H.; Lin, H. Recent advances in conotoxin classification by using machine learning methods. Molecules 2017, 22, 1057. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta 2018, 1862, 81–196. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Olivera, B.M. Venom peptides from cone snails: Pharmacological probes for voltage-gated sodium channels. Curr. Top. Membr. 2016, 78, 65–86. [Google Scholar] [PubMed]
- Milne, T.J.; Abbenante, G.; Tyndall, J.D.A.; Halliday, J.; Lewis, R.J. Isolation and characterization of a cone snail protease with homology to CRISP proteins of the pathogenesis-related protein superfamily. J. Biol. Chem. 2003, 278, 31105–31110. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, L.; Zhou, M.; You, Y.; Zhu, X.; Qiang, Y.; Qin, M.; Luo, S.; Ren, Z.; Xu, A. Molecular evolution and diversity of conus peptide toxins, as revealed by gene structure and intron sequence analyses. PLoS ONE 2013, 8, e82495. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Young, N.D.; Williamson, N.A.; Purcell, A.W. Proteomic interrogation of venom delivery in marine cone snails: Novel insights into the role of the venom bulb. J. Proteome Res. 2010, 9, 5610–5619. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, L.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2016, 112, E3782–E3791. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.J.; Moss, J. Enzymatic and nonenzymatic ADP-ribosylation of cysteine. Mol. Cell. Biochem. 1994, 138, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Tate, E.W.; Kalesh, K.A.; Lanyon-Hogg, T.L.; Storck, E.M.; Thinon, E. Global profiling of protein lipidation using chemical proteomic technologies. Curr. Opin. Chem. Biol. 2015, 24, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Dey, A.; Daaka, Y. Protein S-nitrosylation measurement. Methods Enzymol. 2013, 522, 409–425. [Google Scholar] [PubMed]
- Gajewiak, J.; Azam, L.; Imperiala, J.; Walewska, A.; Green, B.R.; Bandyopadhyay, P.K.; Raghuraman, S.; Ueberheide, B.; Bern, M.; Zhou, H.M.; et al. A disulfide tether stabilizes the block of sodium channels by the conotoxin μO§-GVIIJ. Proc. Natl. Acad. Sci. USA 2014, 111, 2758–2763. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.; Kelley, W.P.; Rubakhin, S.S.; Bingham, J.P.; Sweedler, J.V.; Gilly, W.F. Anatomical correlates of venom production in Conus californicus. Biol Bull. 2002, 203, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Frazao, B.; Vasconcelos, V.; Antunes, A. Sea anemone (Cnidaria, Anthozoa, Actiniaria) roxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: A review on Cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- Valle, A.; Alvarado-Mesen, J.; Lanio, M.E.; Alvarez, C.; Barbosa, J.A.R.G.; Pazos, I.F. The multigene families of actinoporins (part I): Isoforms and genetic structure. Toxicon 2015, 103, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Meinardi, E.; Florin-Christensen, M.; Paratcha, G.; Azcurra, J.M.; Florin-Christensen, J. The molecular basis of the self/nonself selectivity of a coelenterate toxin. Biochim. Biophys. Res. Commun. 1995, 216, 348–354. [Google Scholar] [CrossRef]
- Sher, D.; Zlotkin, E. A hydra with many heads: Protein and polypeptide toxins from hydra and their biological roles. Toxicon 2009, 54, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- De Queiroz, M.R.; de Sousa, B.B.; da Cunha Pereira, D.F.; Mamede, C.C.N.; Matias, M.S.; de Morais, N.C.G.; de Oliveira Costa, J.; de Oliveira, F. The role of platelets in hemostasis and the effects of snake venom toxins on platelet function. Toxicon 2017, 133, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.F. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Renome Res. 2005, 15, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; Bordon, K.C.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro, E.L., Jr.; et al. Minor snake venom proteins: Structure, function and potential applications. Biochim. Biophys. Acta 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, S. Animal toxins for channelopathy treatment. Neuropharmacology 2018, 132, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, F.A.; de Groat, W.C.; Artim, D.E. Bungarotoxins. In Botulinum Toxin. Therapeutic Clinical Practice and Science; Jankovic, J., Albanese, A., Atassi, M.Z., Dolly, J.O., Hallett, M., Mayer, N.H., Eds.; Elsevier: New York, NY, USA, 2009; pp. 425–445. ISBN 978-1-4160-4928-9. [Google Scholar]
- Nirthanan, S.; Gwee, M.C. Three-finger α-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dani, J.A. Neuronal nicotinic acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 2015, 124, 3–19. [Google Scholar]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Sine, S.M.; Engel, A.G. Recent advances in Cys-loop receptor structure and function. Nature 2006, 440, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Unwin, N. Refined structure of the nicotinic acetylcholine receptor at 4A resolution. J. Mol. Biol. 2005, 346, 967–989. [Google Scholar] [CrossRef] [PubMed]
- Barchan, D.; Kachalsky, S.; Neumann, D.; Vogel, Z.; Ovadia, M.; Kochva, E.; Fuchs, S. How the mongoose can fight the snake: The binding site of the mongoose acetylcholine receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 7717–7721. [Google Scholar] [CrossRef] [PubMed]
- Barchan, D.; Ovadia, M.; Kochva, E.; Fuchs, S. The binding site of the nicotinic acetylcholine receptor in animal species resistant to α-bungarotoxin. Biochemistry 1995, 34, 9172–9176. [Google Scholar] [CrossRef] [PubMed]
- Drabeck, D.H.; Dean, A.M.; Jansa, S.A. Why the honey badger don’t care: Convergent evolution of venom-targeted nicotinic acetylcholine receptors in mammals that survive venomous snake bites. Toxicon 2015, 99, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Cobra (Naja spp.) nicotinic acetylcholine receptor exhibits resistance to Erabu sea snake (Laticauda semifasciata) short-chain α-neurotoxin. J. Mol. Evol. 2004, 58, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Hannan, S.; Mortensen, M.; Smart, T.G. Snake neurotoxin α-bungarotoxin is an antagonist at native GABAA receptors. Neuropharmacology 2015, 93, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Fuly, A.L.; Soares, A.M.; Marcussi, S.; Giglio, J.R.; Guimarães, J.A. Signal transduction pathways involved in the platelet aggregation induced by a D-49 phospholipase A2 isolated from Bothrops jararacussu snake venom. Biochimie 2004, 86, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Catanese, J.J.; Kress, L.F. Isolation from opossum serum of a metalloproteinase inhibitor homologous to human α-1B-glycoprotein. Biochemistry 1992, 31, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Biardi, J.E.; Coss, R.G. Rock squirrel (Spermophilus variegatus) blood sera affects proteolytic and hemolytic activities of rattlesnake venoms. Toxicon 2011, 57, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Drabeck, D.H.; Jansa, S.A.; Gibbs, H.L. Venom resistance as a model for understanding the molecular basis of complex coevolutionary adaptations. Integr. Comp. Biol. 2016, 56, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Jansa, S.A.; Voss, R.S. Adaptive evolution of the venom-targeted vWF protein in opossums that eat pitvipers. PLoS ONE 2011, 6, e20997. [Google Scholar] [CrossRef] [PubMed]
- Rosso, J.P.; Schwarz, J.R.; Diaz-Bustamante, M.; Céard, B.; Gutiérrez, J.M.; Kneussel, M.; Pongs, O.; Bosmans, F.; Bougis, P.E. MmTX1 and MmTX2 from coral snake venom potently modulate GABAA receptor activity. Proc. Natl. Acad. Sci. USA 2015, 112, E891–E900. [Google Scholar] [CrossRef] [PubMed]
- Malany, S.; Osaka, H.; Sine, S.M.; Taylor, P. Orientation of α-neurotoxin at the subunit interfaces of the nicotinic acetylcholine receptor. Biochemistry 2000, 39, 15388–15398. [Google Scholar] [CrossRef] [PubMed]
- Sine, S.M. The nicotinic receptor ligand binding domain. J. Neurobiol. 2002, 53, 431–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.K.; Stokes, C.; Horenstein, N.A.; Papke, R.L. The effective opening of nicotinic acetylcholine receptors with single agonist binding sites. J. Gen. Physiol. 2011, 137, 369–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Housley, D.M.; Housley, G.D.; Liddell, M.J.; Jennings, E.A. Scorpion toxin peptide action at the ion channel subunit level. Neuropharmacology 2017, 127, 46–78. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Hernández, V.; Jiménez-Vargas, J.M.; Gurrola, G.B.; Valdivia, H.H.; Possani, L.D. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A.; Swanson, T.M. Structural basis for pharmacology of voltage-gated sodium and calcium channels. Mol. Pharmacol. 2015, 88, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Bosmans, F.; Tytgat, J. Voltage-gated sodium channel modulation by scorpion α-toxins. Toxicon 2007, 49, 142–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saab, C.Y.; Cummins, T.R.; Dib-Hajj, S.D.; Waxman, S.G. Molecular determinant of Nav1.8 sodium channel resistance to the venom from the scorpion Leiurus quinquestriatus hebraeus. Neurosci. Lett. 2002, 331, 79–82. [Google Scholar] [CrossRef]
- Sautiere, P.; Cestele, S.; Kopeyan, C.; Martinage, A.; Drobecq, H.; Doljansky, Y.; Gordon, D. New toxins acting on sodium channels from the scorpion Leiurus quinquestriatus hebraeus suggest a clue to mammalian vs insect selectivity. Toxicon 1998, 36, 1141–1154. [Google Scholar] [CrossRef]
- Fishman, L.; Herrmann, R.; Gordon, D.; Zlotkin, E. Insect tolerance to a neurotoxic polypeptide: Pharmacokinetic and pharmacodynamic aspects. J. Exp. Biol. 1997, 200, 1115–1123. [Google Scholar] [PubMed]
- Cestele, S.; Qu, Y.; Rogers, J.C.; Rochat, H.; Scheuer, T.; Catterall, W.A. Voltage sensor-trapping: Enhanced activation of sodium channels by β-scorpion toxin bound to the34 S–S loop in domain II. Neuron 1998, 21, 919–931. [Google Scholar] [CrossRef]
- Rowe, A.H.; Xiao, Y.; Rowe, M.P.; Cummins, T.R.; Zakon, H.H. Voltage-gated sodium channel in grasshopper mice defends against bark scorpion toxin. Science 2013, 342, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holderied, M.; Korine, C.; Moritz, T. Hemprich’s long-eared bat (Otonycteris hemprichii) as a predator of scorpions: Whispering echolocation, passive gleaning and prey selection. J. Comp. Physiol. A 2011, 197, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Hopp, B.H.; Arvidson, R.S.; Adams, M.E.; Razak, K.A. Arizona bark scorpion venom resistance in the pallid bat, Antrozous pallidus. PLoS ONE 2017, 12, e0183215. [Google Scholar] [CrossRef] [PubMed]
- Legros, C.; Martin-Eauclaire, M.F.; Cattaert, D. The myth of scorpion suicide: Are scorpions insensitive to their own venom? J. Exp. Biol. 1998, 201, 2625–2636. [Google Scholar] [PubMed]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beleboni, R.D.O.; Pizzo, A.B.; Fontana, A.C.; de OG Carolino, R.; Coutinho-Netto, J.; Dos Santos, W.F. Spider and wasp neurotoxins: Pharmacological and biochemical aspects. Eur. J. Pharmacol. 2004, 493, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bosmans, F.; Swartz, K.J. Targeting voltage sensors in sodium channels with spider toxins. Trends Pharmacol. Sci. 2010, 31, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klint, J.K.; Senff, S.; Rupasinghe, D.B.; Er, S.Y.; Herzig, V.; Nicholson, G.M.; King, G.F. Spider-venom peptides that target voltage-gated sodium channels: Pharmacological tools and potential therapeutic leads. Toxicon 2012, 60, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Rash, L.D.; Hodgson, W.C. Pharmacology and biochemistry of spider venoms. Toxicon 2002, 40, 225–254. [Google Scholar] [CrossRef]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Insects, arachnids and centipedes venom: A powerful weapon against bacteria. A literature review. Toxicon 2017, 130, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Nunes, K.P.; Torres, F.S.; Borges, M.H.; Matavel, A.; Pimenta, A.M.; De Lima, M.E. New insights on arthropod toxins that potentiate erectile function. Toxicon 2013, 69, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Blumenthal, K.; Jackson, J.O., II; Liang, S.; Cummins, T.R. The tarantula toxins ProTx-II and huwentoxin-IV differentially interact with human Nav1.7 voltage sensors to inhibit channel activation and inactivation. Mol. Pharmacol. 2010, 78, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Luo, X.; Jiang, L.; Chen, H.; Wang, J.; He, H.; Liang, S. Synthesis and biological characterization of synthetic analogs of Huwentoxin-IV (Mu-theraphotoxin-Hh2a), a neuronal tetrodotoxin-sensitive sodium channel inhibitor. Toxicon 2013, 71, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Hu, Z.; Cai, T.; Liu, K.; Wu, W.; Luo, X.; Jiang, L.; Wang, M.; Yang, J.; Xiao, Y.; et al. Characterization of ion channels on subesophageal ganglion neurons from Chinese tarantula Ornithoctonus huwena: Exploring the myth of the spider insensitive to its venom. Toxicon 2016, 120, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Tang, J.; Hu, W.; Xie, J.; Maertens, C.; Tytgat, J.; Liang, S. Jingzhaotoxin-I, a novel spider neurotoxin preferentially inhibiting cardiac sodium channel inactivation. J. Biol. Chem. 2005, 280, 12069–12076. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Chen, X.; Lu, M.; Wu, Y.; Deng, M.; Zeng, X.; Liu, Z.; Liang, S. Molecular determinant for the tarantula toxin jingzhaotoxin-I slowing the fast inactivation of voltage-gated sodium channels. Toxicon 2016, 111, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Prates, I.; Antoniazzi, M.M.; Sciani, J.M.; Pimenta, D.C.; Toledo, L.F.; Haddad, C.F.; Jared, C. Skin glands, poison and mimicry in dendrobatid and leptodactylid amphibians. J. Morphol. 2012, 273, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W.; Spande, T.F.; Garraffo, H.M. Alkaloids from amphibian skin: A tabulation of over eight-hundred compounds. J. Nat. Prod. 2005, 68, 1556–1575. [Google Scholar] [CrossRef] [PubMed]
- Clark, V.C.; Raxworthy, C.J.; Rakotomalala, V.; Sierwald, P.; Fisher, B.L. Convergent evolution of chemical defense in poison frogs and arthropod prey between Madagascar and the Neotropics. Proc. Natl. Acad. Sci. USA 2005, 102, 11617–11622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hantak, M.M.; Grant, T.; Reinsch, S.; McGinnity, D.; Loring, M.; Toyooka, N.; Saporito, R.A. Dietary alkaloid sequestration in a poison frog: An experimental test of alkaloid uptake in Melanophryniscus stelzneri (Bufonidae). J. Chem. Ecol. 2013, 39, 1400–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGugan, J.R.; Byrd, G.D.; Roland, A.B.; Caty, S.N.; Kabir, N.; Tapia, E.E.; Trauger, S.A.; Coloma, L.A.; O’Connell, L.A. Ant and mite diversity drives toxin vriation in the little devil poison frog. J. Chem. Ecol. 2016, 42, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.M.; Toma, T.; Thomas-Tran, R.; Du Bois, J. Asymmetric synthesis of batrachotoxin: Enantiomeric toxins show functional divergence against NaV. Science 2016, 354, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Y.; Wang, G.K. Single rat muscle Na+ channel mutation confers batrachotoxin autoresistance found in poison-dart frog Phyllobates terribilis. Proc. Natl. Acad. Sci. USA 2017, 114, 10491–10496. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Jiang, T.; Li, Q.; Ling, X. Camptothecin (CPT) and its derivatives are known to target topoisomerase I (Top1) as their mechanism of action: Did we miss something in CPT analogue molecular targets for treating human disease such as cancer? Am. J. Cancer Res. 2017, 7, 2350–2394. [Google Scholar] [PubMed]
- Sirikantaramas, S.; Yamazaki, M.; Saito, K. Camptothecin: Biosynthesis, biotechnological production and resistance mechanism(s). Adv. Bot. Res. 2013, 68, 139–161. [Google Scholar]
- Wall, M.E.; Wani, M.C.; Cook, C.E.; Palmer, K.H.; McPhail, A.T.; Sim, G.A. Plant antitumor agents. I. The isolation and structure of camptothecin, a novel alkaloidal leukemia and tumor inhibitor from Camptotheca acuminata. J. Am. Chem. Soc. 1966, 88, 3888–3890. [Google Scholar] [CrossRef]
- Puri, S.C.; Verma, V.; Amna, T.; Qazi, G.N.; Spiteller, M. An endophytic fungus from Nothapodytes foetida that produces camptothecin. J. Nat. Prod. 2005, 68, 1717–1719. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Shawl, A.S.; Verma, V.; Kour, A.; Athar, M.; Andrabi, R.; Sultan, P.; Qazi, G.N. An endophytic Neurospora sp. from Nothapodytes foetida producing camptothecin. Appl. Biochem. Microbiol. 2008, 44, 203–209. [Google Scholar] [CrossRef]
- Soujanya, K.N.; Siva, R.; Mohana Kumara, P.; Srimany, A.; Ravikanth, G.; Mulani, F.A.; Aarthy, T.; Thulasiram, H.V.; Santhoshkumar, T.R.; Nataraja, K.N.; et al. Camptothecin-producing endophytic bacteria from Pyrenacantha volubilis Hook. (Icacinaceae): A possible role of a plasmid in the production of camptothecin. Phytomedicine 2017, 36, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Sirikantaramas, S.; Yamazaki, M.; Saito, K. Mutations in topoisomerase I as a self-resistance mechanism coevolved with the production of the anticancer alkaloid camptothecin in plants. Proc. Natl. Acad. Sci. USA 2008, 105, 6782–6786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viraporn, V.; Yamazaki, M.; Saito, K.; Denduangboripant, J.; Chayamarit, K.; Chuanasa, T.; Sukrong, S. Correlation of camptothecin-producing ability and phylogenetic relationship in the genus Ophiorrhiza. Planta Med. 2011, 77, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, J.; Roland, J.; Peeters, E.; Trombik, T.; Ducos, E.; Nader, J.; Boutry, M. NtPDR1, a plasma membrane ABC transporter from Nicotiana tabacum, is involved in diterpene transport. Plant Mol. Biol. 2013, 82, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Stukkens, Y.; Bultreys, A.; Grec, S.; Trombik, T.; Vanham, D.; Boutry, M. NpPDR1, a pleiotropic drug resistance-type ATP-binding cassette transporter from Nicotiana plumbaginifolia, plays a major role in plant pathogen defense. Plant Physiol. 2005, 139, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Jirschitzka, J.; Mattern, D.J.; Gershenzon, J.; D’Auria, J.C. Learning from nature: New approaches to the metabolic engineering of plant defense pathways. Curr. Opin. Biotechnol. 2013, 24, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Theodoulou, F.L.; Carrier, D.J.; Schaedler, T.A.; Baldwin, S.A.; Baker, A. How to move an amphipathic molecule across a lipid bilayer: Different mechanisms for different ABC transporters? Biochem. Soc. Trans. 2016, 44, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Flavonoid transport mechanisms: How to go, and with whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Bazin, I.; Dan, K.; Obata, K.; Kigawa, K.; Ueda, K.; Sato, F.; Forestier, C.; Yazaki, K. Involvement of CjMDR1, a plant multidrug-resistance-type ATP-binding cassette protein, in alkaloid transport in Coptis japonica. Proc. Natl. Acad. Sci. USA 2003, 100, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, K.; Sakai, K.; Sato, F.; Yamamoto, H.; Yazaki, K. Thalictrum minus cell cultures and ABC-like transporter. Phytochemistry 2003, 62, 483–489. [Google Scholar] [CrossRef]
- Otani, M.; Shitan, N.; Sakai, K.; Martinoia, E.; Sato, F.; Yazaki, K. Characterization of vacuolar transport of the endogenous alkaloid berberine in Coptis japonica. Plant Physiol. 2005, 138, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, K. Vinblastine. Compr. Pharmacol. Ref. 2007, 1–4. [Google Scholar] [CrossRef]
- Davey, M. Secondary metabolism in plant cell cultures. Encycl. Appl. Plant Sci. 2017, 2, 462–467. [Google Scholar]
- Sharma, A.; Verma, P.; Mathur, A.; Mathur, A.K. Genetic engineering approach using early Vinca alkaloid biosynthesis genes led to increased tryptamine and terpenoid indole alkaloids biosynthesis in differentiating cultures of Catharanthus roseus. Protoplasma 2018, 255, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; De Luca, V. ATP-binding cassette transporter controls leaf surface secretion of anticancer drug components in Catharanthus roseus. Proc. Natl. Acad. Sci. USA 2013, 110, 15830–15835. [Google Scholar] [CrossRef] [PubMed]
- Raffa, D.; Maggio, B.; Raimondi, M.V.; Plescia, F.; Daidone, G. Recent discoveries of anticancer flavonoids. Eur. J. Med. Chem. 2017, 142, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Gaziano, R.; Moroni, G.; Buè, C.; Miele, M.T.; Sinibaldi-Vallebona, P.; Pica, F. Antitumor effects of the benzophenanthridine alkaloid sanguinarine: Evidence and perspectives. World J. Gastrointest. Oncol. 2016, 8, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Bird, D.A.; Franceschi, V.R.; Facchini, P.J. A tale of three cell types: Alkaloid biosynthesis is localized to sieve elements in opium poppy. Plant Cell 2003, 15, 2626–2635. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Khan, S.A.; Mathur, A.K.; Ghosh, S.; Shanker, K.; Kalra, A. Improved sanguinarine production via biotic and abiotic elicitations and precursor feeding in cell suspensions of latex-less variety of Papaver somniferum with their gene expression studies and upscaling in bioreactor. Protoplasma 2014, 251, 1359–1371. [Google Scholar] [CrossRef] [PubMed]
- Sandor, R.; Slanina, J.; Midlik, A.; Sebrlova, K.; Novotna, L.; Carnecka, M.; Slaninova, I.; Taborsky, P.; Taborska, E.; Pes, O. Sanguinarine is reduced by NADH through a covalent adduct. Phytochemistry 2018, 145, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Vogel, M.; Lawson, M.; Sippl, W.; Conrad, U.; Roos, W. Structure and mechanism of sanguinarine reductase, an enzyme of alkaloid detoxification. J. Biol. Chem. 2010, 285, 18397–18406. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Baumert, A.; Vogel, M.; Roos, W. Sanguinarine reductase, a key enzyme of benzophenanthridine detoxification. Plant Cell Environ. 2006, 29, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotti, H.; Rischer, H. The killer of Socrates: Coniine and related alkaloids in the plant kingdom. Molecules 2017, 22, E1962. [Google Scholar] [CrossRef] [PubMed]
- Arihan, O.; Boz, M.; Iskit, A.B.; Ilhan, M. Antinociceptive activity of coniine in mice. J. Ethnopharmacol. 2009, 125, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Castells, E.; Berenbaum, M.R. Resistance of the generalist moth Trichoplusia ni (Noctuidae) to a novel chemical defence in the invasive plant Conium maculatum. Chemoecology 2008, 18, 11–18. [Google Scholar] [CrossRef]
- Castells, E.; Mulder, P.P.; Pérez-Trujillo, M. Diversity of pyrrolizidine alkaloids in native and invasive Senecio pterophorus (Asteraceae): Implications for toxicity. Phytochemistry 2014, 108, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.; Stevens, K. Pyrrolizidine alkaloids. Nat. Prod. Rep. 2014, 31, 1721–1788. [Google Scholar] [CrossRef] [PubMed]
- Skoneczny, D.; Weston, P.A.; Zhu, X.; Gurr, G.M.; Callaway, R.M.; Weston, L.A. Metabolic profiling of pyrrolizidine alkaloids in foliage of two Echium spp. Invaders in Australia—A case of novel weapons? Int. J. Mol. Sci. 2015, 16, 26721–26737. [Google Scholar] [CrossRef] [PubMed]
- Schimming, O.; Challinor, V.L.; Tobias, N.J.; Adihou, H.; Gruen, P.; Poeschel, L.; Richter, C.; Schwalbe, H.; Bode, H.B. Structure, biosynthesis, and occurrence of bacterial pyrrolizidine alkaloids. Angew. Chem. Int. Ed. Engl. 2015, 54, 12702–12705. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T. Chemical ecology of pyrrolizidine alkaloids. Plants 1999, 207, 483–495. [Google Scholar] [CrossRef]
- Del Campo, M.L.; Smedley, S.R.; Eisner, T. Reproductive benefits derived from defensive plant alkaloid possession in an arctiid moth (Utetheisa ornatrix). Proc. Natl. Acad. Sci. USA 2005, 102, 13508–13512. [Google Scholar] [CrossRef] [PubMed]
- Eisner, T.; Eisner, M. Unpalatability of the pyrrolizidine alkaloid-containing moth Utetheisa ornatrix, and its larva, to wolf spiders. Psyche A J. Entomol. 1991, 98, 111–118. [Google Scholar] [CrossRef]
- Ehmke, A.; Witte, L.; Biller, A.; Hartmann, T. Sequestration, N-oxidation and transformation of plant pyrrolizidine alkaloids by the arctiid moth Tyria jacobaeae L. Z. Naturforsch. 1990, 45, 1185–1192. [Google Scholar]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Ann. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Roddick, J.G. Acetylcholinesterase-inhibitory activity of steroidal glycoalkaloids and their aglycones. Phytochemistry 1989, 28, 2631–2634. [Google Scholar] [CrossRef]
- Cardenas, P.D.; Sonawane, P.D.; Heinig, U.; Bocobza, S.E.; Burdman, S.; Aharoni, A. The bitter side of the nightshades: Genomics drives discovery in Solanaceae steroidal alkaloid metabolism. Phytochemistry 2015, 113, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Itkin, M.; Heinig, U.; Tzfadia, O.; Bhide, A.J.; Shinde, B.; Cardenas, P.D.; Bocobza, S.E.; Unger, T.; Malitsky, S.; Finkers, R.; et al. Biosynthesis of antinutritional alkaloids in solanaceous crops is mediated by clustered genes. Science 2013, 341, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.E.; McCance, D.J.; Drysdale, R.B. The detoxication of α-tomatine by Fusarium oxysporum f. sp. lycopersici. Phytochemistry 1977, 16, 545–546. [Google Scholar] [CrossRef]
- Oekmen, B.; Etalo, D.W.; Joosten, M.H.; Bouwmeester, H.J.; de Vos, R.C.; Collemare, J.; de Wit, P.J. Detoxification of α-tomatine by Cladosporium fulvum is required for full virulence on tomato. New Phytol. 2013, 198, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Blankemeyer, J.T.; White, J.B.; Stringer, B.K.; Friedman, M. Effect of α-tomatine and tomatidine on membrane potential of frog embryos and active transport of ions in frog skin. Food Chem. Toxicol. 1997, 35, 639–646. [Google Scholar] [CrossRef]
- Itkin, M.; Rogachev, I.; Alkan, N.; Rosenberg, T.; Malitsky, S.; Masini, L.; Meir, S.; Iijima, Y.; Aoki, K.; de Vos, R.; et al. GLYCOALKALOID METABOLISM1 is required for steroidal alkaloid glycosylation and prevention of phytotoxicity in tomato. Plant Cell 2011, 23, 4507–4525. [Google Scholar] [CrossRef] [PubMed]
- Wouters, F.C.; Blanchett, B.; Gershenzon, J.; Vassao, D.G. Plant defense and herbivore counter-defense: Benzoxazinoids and insect herbivores. Phytochem. Rev. 2016, 15, 1127–1151. [Google Scholar] [CrossRef] [PubMed]
- Pankoke, H.; Buschmann, T.; Mueler, C. Role of plant β-glucosidases in the dual defense system of iridoid glycosides and their hydrolyzing enzymes in Plantago lanceolata and Plantago major. Phytochemistry 2013, 94, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Pentzold, S.; Zagrobelny, M.; Rook, F.; Bak, S. How insects overcome two-component plant chemical defence: Plant β-glucosidases as the main target for herbivore adaptation. Biol. Rev. 2014, 89, 531–551. [Google Scholar] [CrossRef] [PubMed]
- Del Cueto, J.; Ionescu, I.A.; Picmanova, M.; Gericke, O.; Motawia, M.S.; Olsen, C.E.; Campoy, J.A.; Dicenta, F.; Møller, B.L.; Sánchez-Pérez, R. Cyanogenic glucosides and derivatives in almond and sweet cherry flower buds from dormancy to flowering. Front. Plant Sci. 2017, 8, 800. [Google Scholar] [CrossRef] [PubMed]
- Kautz, S.; Trisel, J.A.; Ballhorn, D.J. Jasmonic acid enhances plant cyanogenesis and resistance to herbivory in lima bean. J. Chem. Ecol. 2014, 40, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Huberty, M.; Tielboeger, K.; Harvey, J.A.; Mueller, C.; Macel, M. Chemical defenses (glucosinolates) of native and invasive populations of the range expanding invasive plant Rorippa austriaca. J. Chem. Ecol. 2014, 40, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Plant-derived cardiac glycosides: Role in heart ailments and cancer management. Biomed. Pharmacother. 2016, 84, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Opitz, S.E.W.; Mueller, C. Plant chemistry and insect sequestration. Chemoecology 2009, 19, 117–154. [Google Scholar] [CrossRef]
- Qiu, L.Y.; Swarts, H.G.; Tonk, E.C.; Willems, P.H.; Koenderink, J.B.; De Pont, J.J. Conversion of the low affinity ouabain-binding site of non-gastric H,K-ATPase into a high affinity binding site by substitution of only five amino acids. J. Biol. Chem. 2006, 281, 13533–13539. [Google Scholar] [CrossRef] [PubMed]
- Dalla, S.; Swarts, H.G.; Koenderink, J.B.; Dobler, S. Amino acid substitutions of Na,K-ATPase conferring decreased sensitivity to cardenolides in insects compared to mammals. Insect Biochem. Mol. Biol. 2013, 43, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Dalla, S.; Baum, M.; Dobler, S. Substitutions in the cardenolide binding site and interaction of subunits affect kinetics besides cardenolide sensitivity of insect Na, K-ATPase. Insect Biochem. Mol. Biol. 2017, 89, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.N.; Meinzer, F.; Dalla, S.; Romey-Gluesing, R.; Dobler, S. The function and evolutionary significance of a triplicated Na, K-ATPase gene in a toxin-specialized insect. BMC Evol. Biol. 2017, 17, 256. [Google Scholar] [CrossRef] [PubMed]
- Petschenka, G.; Pick, C.; Wagschal, V.; Dobler, S. Functional evidence for physiological mechanisms to circumvent neurotoxicity of cardenolides in an adapted and a non-adapted hawk-moth species. Proc. Biol. Sci. 2013, 280, 20123089. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, N.L. Pharmacology of nicotine: Addiction, smoking-induced disease, and therapeutics. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Rathi, P.; Schoettner, M.; Baldwin, I.T.; Pandit, S. Differences in nicotine metabolism of two Nicotiana attenuata herbivores render them differentially susceptible to a common native predator. PLoS ONE 2014, 9, e95982. [Google Scholar] [CrossRef] [PubMed]
- Fuerstenberg-Haegg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.D. Preface: Mechanism of plant-insect interaction. J. Exp. Biol. 2015, 66, 421–424. [Google Scholar]
- Kessler, A.; Halitschke, R.; Baldwin, I.T. Silencing the jasmonate cascade: Induced plant defenses and insect populations. Science 2004, 305, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Mithoefer, A.; Boland, W. Before gene expression: Early events in plant-insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R. Chemical ecology of insect-plant interactions: Ecological significance of plant secondary metabolites. Biosci. Biotechnol. Biochem. 2014, 78, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaquerel, E.; Steppuhn, A.; Baldwin, I.T. Nicotiana attenuata α-DIOXYGENASE1 through its production of 2-hydroxylinolenic acid is required for intact plant defense expression against attack from Manduca sexta larvae. New Phytol. 2012, 196, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M.; Lawrence, B.M. Menthol: A simple monoterpene with remarkable biological properties. Phytochemistry 2013, 96, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Tissier, A.; Morgan, J.A.; Dudareva, N. Plant volatiles: Going ‘in’ but not ‘out’ of trichome cavities. Trends Plant Sci. 2017, 22, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Frick, K.M.; Kamphuis, L.G.; Siddique, K.H.; Singh, K.B.; Foley, R.C. Quinolizidine alkaloid biosynthesis in lupins and prospects for grain quality improvement. Front. Plant Sci. 2017, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.P. Simple indolizidine and quinolizidine alkaloids. In The Alkaloids: Chemistry and Biology; Knoelker, H.-J., Ed.; Elsevier: New Yorrk, NY, USA, 2016; Volume 75, pp. 1–517. ISBN 978-0-12-803434-7. [Google Scholar]
- Wink, M.; Roemer, P. Acquired toxicity—The advantages of specializing on alkaloid-rich lupins to Macrosiphum albifrons (Aphidae). Naturwissenschaften 1986, 73, 210–212. [Google Scholar] [CrossRef]
- Dohnal, V.; Wu, Q.; Kuca, K. Metabolism of aflatoxins: Key enzymes and interindividual as well as interspecies differences. Arch. Toxicol. 2014, 88, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Neal, G.E. Genetic implications in the metabolism and toxicity of mycotoxins. Toxicol. Lett. 1995, 82, 861–867. [Google Scholar] [CrossRef]
- Cleveland, T.E.; Yu, J.; Fedorova, N.; Bhatnagar, D.; Payne, G.A.; Nierman, W.C.; Bennett, J.W. Potential of Aspergillus flavus genomics for applications in biotechnology. Trends Biotechnol. 2009, 27, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guan, X.; Xing, F.; Lv, C.; Dai, X.; Liu, Y. Effect of water activity and temperature on the growth of Aspergillus flavus, the expression of aflatoxin biosynthetic genes and aflatoxin production in shelled peanuts. Food Control 2017, 82, 325–332. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef]
- Tekaia, F.; Latge, J.P. Aspergillus fumigatus: Saprophyte or pathogen? Curr. Opin. Microbiol. 2005, 8, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Yu, J.H.; Kelkar, H.S.; Fernandes, M.; Nesbitt, T.C.; Keller, N.P.; Adams, T.H.; Leonard, T.J. Twenty-five coregulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 1996, 93, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Klejnstrup, M.L.; Frandsen, R.J.; Holm, D.K.; Nielsen, M.T.; Mortensen, U.H.; Larsen, T.O.; Nielsen, J.B. Genetics of polyketide metabolism in Aspergillus nidulans. Metabolites 2012, 2, 100–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a halogenase involved in the biosynthesis of ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microbiol. 2016, 82, 5631–5641. [Google Scholar] [CrossRef] [PubMed]
- Farber, P.; Geisen, R. Analysis of differentially expressed ochratoxin A biosynthesis genes of Penicillium nordicum. Eur. J. Plant Pathol. 2004, 110, 661–669. [Google Scholar] [CrossRef]
- Geisen, R.; Schmidt-Heydt, M.; Karolewiez, A. A gene cluster of the ochratoxin a biosynthetic genes in Penicillium. Mycotoxin Res. 2006, 22, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Garcia-Diaz, M.; Gonzalez-Jaen, M.T.; Vazquez, C.; Patino, B. Description of an orthologous cluster of ochratoxin A biosynthetic genes in Aspergillus and Penicillium species. A comparative analysis. Int. J. Food Microbiol. 2018, 268, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Susca, A.; Proctor, R.H.; Morelli, M.; Haidukowski, M.; Gallo, A.; Logrieco, A.F.; Moretti, A. Variation in fumonisin and ochratoxin production associated with differences in biosynthetic gene content in Aspergillus niger and A. welwitschiae isolates from multiple crop and geographic origins. Front. Microbiol. 2016, 7, 1412. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Xu, Y.; Huang, Z.B. Isolation and characterization of the citrinin biosynthetic gene cluster from Monascus aurantiacus. Biotechnol. Lett. 2012, 34, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, C.A.; Gueldener, U.; Xu, J.R.; Trail, F.; Turgeon, B.G.; Di Pietro, A.; Walton, J.D.; Ma, L.J.; Baker, S.E.; Rep, M.; et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 2007, 317, 1400–1402. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M.; Manners, J.M. On the trail of a cereal killer: Recent advances in Fusarium graminearum pathogenomics and host resistance. Mol. Plant Pathol. 2012, 13, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Windram, O.; Denby, K.J. Modelling signaling networks underlying plant defence. Curr. Opin. Plant Biol. 2015, 27, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Choi, Y.E.; Zou, X.; Xu, J.R. The FvMK1 mitogen-activated protein kinase gene regulates conidiation, pathogenesis, and fumonisin production in Fusarium verticillioides. Fungal Genet. Biol. 2011, 48, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, S.A.; Boddu, J.; Berthiller, F.; Hametner, C.; Stupar, R.M.; Adam, G.; Muehlbauer, G.J. Transcriptome analysis of the barley-deoxynivalenol interaction: Evidence for a role of glutathione in deoxynivalenol detoxification. Mol. Plant Microbe Interact. 2010, 23, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Tokai, T.; O’Donnell, K.; Ward, T.J.; Fujimura, M.; Hamamoto, H.; Shibata, T.; Yamaguchi, I. The trichothecene biosynthesis gene cluster of Fusarium graminearum F15 contains a limited number of essential pathway genes and expressed non-essential genes. FEBS Lett. 2003, 539, 105–110. [Google Scholar] [CrossRef]
- Lee, T.; Lee, S.H.; Shin, J.Y.; Kim, H.K.; Yun, S.H.; Kim, H.Y.; Lee, S.; Ryu, J.G. Comparison of Trichothecene Biosynthetic Gene Expression between Fusarium graminearum and Fusarium asiaticum. Plant Pathol. J. 2014, 30, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Malz, S.; Grell, M.N.; Thrane, C.; Maier, F.J.; Rosager, P.; Felk, A.; Albertsen, K.S.; Salomon, S.; Bohn, L.; Schaefer, W.; et al. Identification of a gene cluster responsible for the biosynthesis of aurofusarin in the Fusarium graminearum species complex. Fungal Genet. Biol. 2005, 42, 420–433. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Gorczynska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Gaffoor, I.; Trail, F. Characterization of two polyketide synthase genes involved in zearalenone biosynthesis in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Lee, Y.R.; Jin, J.; Han, K.H.; Kim, H.; Kim, J.C.; Lee, T.; Yun, S.H.; Lee, Y.W. Two different polyketide synthase genes are required for synthesis of zearalenone in Gibberella zeae. Mol. Microbiol. 2005, 58, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Lysøe, E.; Klemsdal, S.S.; Bone, K.R.; Frandsen, R.J.; Johansen, T.; Thrane, U.; Giese, H. The PKS4 gene of Fusarium graminearum is essential for zearalenone production. Appl. Environ. Microbiol. 2006, 72, 3924–3932. [Google Scholar] [CrossRef] [PubMed]
- Kosawang, C.; Karlsson, M.; Velez, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D.F. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fungal Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Ando, N.; Kimura, M.; Kakeya, H.; Osada, H.; Yamaguchi, I. A novel lactonohydrolase responsible for the detoxification of zearalenone: Enzyme purification and gene cloning. Biochem. J. 2002, 365, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Brown, D.W.; Cheung, F.; Proctor, R.H.; Butchko, R.A.E.; Zheng, L.; Lee, Y.; Utterback, T.; Smith, S.; Feldblyum, T.; Glenn, A.E.; et al. Comparative analysis of 87,000 expressed sequence tags from the fumonisin-producing fungus Fusarium verticillioides. Fungal Genet. Biol. 2005, 42, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.E. Aspergillus niger genomics: Past, present and into the future. Med. Mycol. 2006, 44, S17–S21. [Google Scholar] [CrossRef] [PubMed]
- Susca, A.; Proctor, R.H.; Butchko, R.A.; Haidukowski, M.; Stea, G.; Logrieco, A.; Moretti, A. Variation in the fumonisin biosynthetic gene cluster in fumonisin-producing and nonproducing black aspergilli. Fungal Genet. Biol. 2014, 73, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tagami, K.; Minami, A.; Matsumoto, T.; Frisvad, J.C.; Suzuki, H.; Ishikawa, J.; Gomi, K.; Oikawa, H. Reconstitution of biosynthetic machinery for the synthesis of the highly elaborated indole diterpene penitrem. Angew. Chem. Int. Ed. Engl. 2015, 54, 5748–5752. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Nielsen, K.F.; Smedsgaard, J. Roquefortine/oxaline biosynthesis pathway metabolites in Penicillium ser. Corymbifera: In planta production and implications for competitive fitness. J. Chem. Ecol. 2005, 31, 2373–2390. [Google Scholar] [PubMed]
- Ali, H.; Ries, M.I.; Nijland, J.G.; Lankhorst, P.P.; Hankemeier, T.; Bovenberg, R.A.; Vreeken, R.J.; Driessen, A.J. A branched biosynthetic pathway is involved in production of roquefortine and related compounds in Penicillium chrysogenum. PLoS ONE 2013, 8, e65328. [Google Scholar] [CrossRef] [PubMed]
- Toyomasu, T.; Nakaminami, K.; Toshima, H.; Mie, T.; Watanabe, K.; Ito, H.; Matsui, H.; Mitsuhashi, W.; Sassa, T.; Oikawa, H. Cloning of a gene cluster responsible for the biosynthesis of diterpene aphidicolin, a specific inhibitor of DNA polymerase α. Biosci. Biotechnol. Biochem. 2004, 68, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Del Sorbo, G.; Ruocco, M.; Schoonbeek, H.J.; Scala, F.; Pane, C.; Vinale, F.; De Waard, M.A. Cloning and functional characterization of BcatrA, a gene encoding an ABC transporter of the plant pathogenic fungus Botryotinia fuckeliana (Botrytis cinerea). Mycol. Res. 2008, 112, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Cozijnsen, A.J.; Wilson, L.M.; Pedras, M.S.; Howlett, B.J. The sirodesmin biosynthetic gene cluster of the plant pathogenic fungus Leptosphaeria maculans. Mol. Microbiol. 2004, 53, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Jarvis, R.S.; Howlett, B.J. The ABC transporter gene in the sirodesmin biosynthetic gene cluster of Leptosphaeria maculans is not essential for sirodesmin production but facilitates self-protection. Fungal Genet. Biol. 2005, 42, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; O’Keeffe, G.; Jones, G.W.; Doyle, S. Resistance is not futile: Gliotoxin biosynthesis, functionality and utility. Trends Microbiol. 2015, 23, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Carberry, S.; Kavanagh, K.; Haas, H.; Jones, G.W.; O’Brien, J.; Nolan, A.; Stephens, J.; Fenelon, O.; Doyle, S. Self-protection against gliotoxin—A component of the gliotoxin biosynthetic cluster, GliT, completely protects Aspergillus fumigatus against exogenous gliotoxin. PLoS Pathog. 2010, 6, e1000952. [Google Scholar] [CrossRef] [PubMed]
- Guerre, P. Ergot alkaloids produced by endophytic fungi of the genus Epichloë. Toxins 2015, 7, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, T.; Machado, C.; Luebbe, Y.; Correia, T.; Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. The ergot alkaloid gene cluster in Claviceps purpurea: Extension of the cluster sequence and intra species evolution. Phytochemistry 2005, 66, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, N.; Haarmann, T.; Pazoutova, S.; Jung, M.; Tudzynski, P. The ergot alkaloid gene cluster: Functional analyses and evolutionary aspects. Phytochemistry 2009, 70, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Li, S.M. Ergot alkaloids: Structure diversity, biosynthetic gene clusters and functional proof of biosynthetic genes. Nat. Prod. Rep. 2011, 28, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Coyle, C.M.; Panaccione, D.G. An ergot alkaloid biosynthesis gene and clustered hypothetical genes from Aspergillus fumigatus. Appl. Environ. Microbiol. 2005, 71, 3112–3118. [Google Scholar] [CrossRef] [PubMed]
- Florea, S.; Panaccione, D.G.; Schardl, C.L. Ergot Alkaloids of the Family Clavicipitaceae. Phytopathology 2017, 107, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, N.; Wilson, E.V.; Machado, C.; Schardl, C.L.; Tudzynski, P. Comparison of ergot alkaloid biosynthesis gene clusters in Claviceps species indicates loss of late pathway steps in evolution of C. fusiformis. Appl. Environ. Microbiol. 2007, 73, 7185–7191. [Google Scholar] [CrossRef] [PubMed]
- Unsold, I.A.; Li, S.M. Overproduction, purification and characterization of FgaPT2, a dimethylallyltryptophan synthase from Aspergillus fumigatus. Microbiology 2005, 151, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Wallwey, C.; Matuschek, M.; Xie, X.L.; Li, S.M. Ergot alkaloid biosynthesis in Aspergillus fumigatus: Conversion of chanoclavine-I aldehyde to festuclavine by the festuclavine synthase FgaFS in the presence of the old yellow enzyme FgaOx3. Org. Biomol. Chem. 2010, 8, 3500–3508. [Google Scholar] [CrossRef] [PubMed]
- Goetz, K.E.; Coyle, C.M.; Cheng, J.Z.; O’Connor, S.E.; Panaccione, D.G. Ergot cluster-encoded catalase is required for synthesis of chanoclavine-I in Aspergillus fumigatus. Curr. Genet. 2011, 57, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Young, C.A.; Hesse, U.; Amyotte, S.G.; Andreeva, K.; Calie, P.J.; Fleetwood, D.J.; Haws, D.C.; Moore, N.; Oeser, B.; et al. Plant-symbiotic fungi as chemical engineers: Multi-genome analysis of the clavicipitaceae reveals dynamics of alkaloid loci. PLoS Genet. 2013, 9, e1003323. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Kutil, B.L.; Greenwald, C.; Liu, G.; Spiering, M.J.; Schardl, C.L.; Wilkinson, H.H. Comparison of loline alkaloid gene clusters across fungal endophytes: Predicting the co-regulatory sequence motifs and the evolutionary history. Fungal Genet. Biol. 2007, 44, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Bhardwaj, M.; Faulkner, J.R.; Nagabhyru, P.; Charlton, N.D.; Higashi, R.M.; Miller, A.F.; Young, C.A.; Grossman, R.B.; Schardl, C.L. Ether bridge formation in loline alkaloid biosynthesis. Phytochemistry 2014, 98, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiering, M.J.; Moon, C.D.; Wilkinson, H.H.; Schardl, C.L. Gene clusters for insecticidal loline alkaloids in the grass-endophytic fungus Neotyphodium uncinatum. Genetics 2005, 169, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Schardl, C.L.; Panaccione, D.G.; Florea, S.; Takach, J.E.; Charlton, N.D.; Moore, N.; Webb, J.S.; Jaromczyk, J. Genetics, genomics and evolution of ergot alkaloid diversity. Toxins 2015, 7, 1273–1302. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Tapper, B.A.; Popay, A.; Parker, E.J.; Scott, B. A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory. Mol. Microbiol. 2005, 57, 1036–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, S.; Takemoto, D.; Tapper, B.A.; Lane, G.A.; Fraser, K.; Scott, B. Functional analysis of an indole-diterpene gene cluster for lolitrem B biosynthesis in the grass endosymbiont Epichloë festucae. FEBS Lett. 2012, 586, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Bryant, M.K.; Christensen, M.J.; Tapper, B.A.; Bryan, G.T.; Scott, B. Molecular cloning and genetic analysis of a symbiosis-expressed gene cluster for lolitrem biosynthesis from a mutualistic endophyte of perennial ryegrass. Mol. Genet. Genom. 2005, 274, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Donzelli, B.G.G.; Creamer, R.; Baucom, D.L.; Gardner, D.R.; Pan, J.; Moore, N.; Krasnoff, S.B.; Jaromczyk, J.W.; Schardl, C.L. Swainsonine biosynthesis genes in diverse symbiotic and pathogenic fungi. G3 Genes Genomes Genet. 2017, 7, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Benveniste, R.; Davies, J. Aminoglycoside antibiotic-inactivating enzymes in actinomycetes similar to those present in clinical isolates of antibiotic-resistant bacteria. Proc. Natl. Acad. Sci. USA 1973, 70, 2276–2280. [Google Scholar] [CrossRef] [PubMed]
- Ogawara, H. Antibiotic resistance in pathogenic and producing bacteria, with special reference to β-lactam antibiotics. Microbiol. Rev. 1981, 45, 591–619. [Google Scholar] [PubMed]
- Walker, M.S.; Walker, J.B. Streptomycin biosynthesis and metabolism. Enzymatic phosphorylation of dihydrostreptobiosamine moieties of dihydrostreptomycin-(streptidino) phosphate and dihydrostreptomycin by Streptomyces extracts. J. Biol. Chem. 1970, 245, 6683–6689. [Google Scholar] [PubMed]
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Crofts, T.S.; Gasparrini, A.J.; Dantas, G. Next-generation approaches to understand and combat the antibiotic resistome. Nat. Rev. Microbiol. 2017, 15, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesme, J.; Simonet, P. The soil resistome: A critical review on antibiotic resistance origins, ecology and dissemination potential in telluric bacteria. Environ. Microbiol. 2015, 17, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Traxler, M.F.; Kolter, R. Natural products in soil microbe interactions and evolution. Nat. Prod. Rep. 2015, 32, 956–970. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Cooper, M.A. Aminoglycoside antibiotics in the 21st century. ACS Chem. Biol. 2013, 8, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ennifar, E.; Aslam, M.W.; Strasser, P.; Hoffmann, G.; Dumas, P.; van Delft, F.L. Structure-guided discovery of a novel aminoglycoside conjugate targeting HIV-1 RNA viral genome. ACS Chem. Biol. 2013, 8, 2509–2517. [Google Scholar] [CrossRef] [PubMed]
- Schatz, A.; Bugie, E.; Waksman, S.A. Streptomycin, a substance exhibiting antibiotic activity against Gram-positive and Gram-negative bacteria. Proc. Soc. Exp. Biol. Med. 1944, 55, 66–69. [Google Scholar] [CrossRef]
- Ohnuki, T.; Imanaka, T.; Aiba, S. Self-cloning in Streptomyces griseus of an str gene cluster for streptomycin biosynthesis and streptomycin resistance. J. Bacteriol. 1985, 164, 85–94. [Google Scholar] [PubMed]
- Distler, J.; Mansouri, K.; Mayer, G.; Stockmann, M.; Piepersberg, W. Streptomycin biosynthesis and its regulation in Streptomycetes. Gene 1992, 115, 105–111. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Ishikawa, J.; Hara, H.; Suzuki, H.; Ikenoya, M.; Ikeda, H.; Yamashita, A.; Hattori, M.; Horinouchi, S. Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J. Bacteriol. 2008, 190, 4050–4060. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H.; Ueda, M.; Maeda, K.; Yagishita, K.; Kondo, S.; Okami, Y.; Utahara, R.; Osato, Y.; Nitta, K.; Takeuchi, T. Production and isolation of a new antibiotic: Kanamycin. J. Antibiot. 1957, 10, 181–188. [Google Scholar] [PubMed]
- Kharel, M.K.; Subba, B.; Basnet, D.B.; Woo, J.S.; Lee, H.C.; Liou, K.; Sohng, J.K. A gene cluster for biosynthesis of kanamycin from Streptomyces kanamyceticus: Comparison with gentamicin biosynthetic gene cluster. Arch. Biochem. Biophys. 2004, 429, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.M.; Kikuchi, T.; Mashiko, H.; Ogawara, H. Analysis of the promoter region of the cloned kanamycin resistance gene (kmr) from Streptomyces kanamyceticus. J. Antibiot. 1989, 42, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Yanai, K.; Murakami, T. The kanamycin biosynthetic gene cluster from Streptomyces kanamyceticus. J. Antibiot. 2004, 57, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Yanai, K.; Murakami, T.; Bibb, M. Amplification of the entire kanamycin biosynthetic gene cluster during empirical strain improvement of Streptomyces kanamyceticus. Proc. Natl. Acad. Sci. USA 2006, 103, 9661–9666. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Park, S.R.; Nepal, K.K.; Han, A.R.; Ban, Y.H.; Yoo, Y.J.; Kim, E.J.; Kim, E.M.; Kim, D.; Sohng, J.K.; et al. Discovery of parallel pathways of kanamycin biosynthesis allows antibiotic manipulation. Nat. Chem. Biol. 2011, 7, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Tamegai, H.; Kudo, F.; Kojima, K.; Kakinuma, K.; Eguchi, T. Biosynthesis of 2-deoxystreptamine-containing antibiotics in Streptoalloteichus hindustanus JCM 3268: Characterization of 2-deoxy-scyllo-inosose synthase. J. Antibiot. 2006, 59, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Janssen, G.R.; Ward, J.M.; Bibb, M.J. Unusual transcriptional and translational features of the aminoglycoside phosphotransferase gene (aph) from Streptomyces fradiae. Genes Dev. 1989, 3, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Cabrera, M.; Perez-Gonzalez, J.A.; Heinzel, P.; Piepersberg, W.; Jimenez, A. Isolation and nucleotide sequencing of an aminocyclitol acetyltransferase gene from Streptomyces rimosus forma paromomycinus. J. Bacteriol. 1989, 171, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Pardo, J.M.; Malpartida, F.; Rico, M.; Jimenez, A. Biochemical basis of resistance to hygromycin B in Streptomyces hygroscopicus—The producing organism. Microbiology 1985, 131, 1289–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniappan, N.; Ayers, S.; Gupta, S.; Habib, E.-S.; Reynolds, K.A. Production of hygromycin A analogs in Streptomyces hygroscopicus NRRL 2388 through identification and manipulation of the biosynthetic gene cluster. Chem. Biol. 2006, 13, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Dhote, V.; Gupta, S.; Reynolds, K.A. An O-phosphotransferase catalyzes phosphorylation of hygromycin A in the antibiotic-producing organism Streptomyces hygroscopicus. Antimicrob. Agents Chemother. 2008, 52, 3580–3588. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Hotta, K.; Okami, Y.; Umezawa, H. Self-resistance of a Streptomyces which produces istamycins. J. Antibiot. 1981, 34, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Hasegawa, M. Analysis of the nucleotide sequence of fmrT encoding the self-defense gene of the istamycin producer, Streptomyces tenjimariensis ATCC 31602; comparison with the squences of kamB of Streptomyces tenebrarius NCIB 11028 and kamC of Saccharopolyspora hirsuta CL102. J. Antibiot. 1993, 46, 511–517. [Google Scholar] [PubMed]
- Hotta, K.; Ogata, T.; Ishikawa, J.; Okanishi, M.; Mizuno, S.; Morioka, M.; Naganawa, H.; Okami, Y. Mechanism of multiple aminoglycoside resistance of kasugamycin-producing Streptomyces kasugaensis MB273: Involvement of two types of acetyltransferases in resistance to astromicin group antibiotics. J. Antibiot. 1996, 49, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Ikeno, S.; Yamane, Y.; Ohishi, Y.; Kinoshita, N.; Hamada, M.; Tsuchiya, K.S.; Hori, M. ABC transporter genes, kasKLM, responsible for self-resistance of a kasugamycin producer strain. J. Antibiot. 2000, 53, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Hasegawa, M. Analysis of the self-defense gene (fmrO) of a fortimicin A (astromicin) producer, Micromonospora olivasterospora: Comparison with other aminoglycoside-resistance-encoding genes. Gene 1993, 127, 63–69. [Google Scholar] [PubMed]
- Kobayashi, T.; Uozumi, T.; Beppu, T. Cloning and characterization of the streptothricin-resistance gene which encodes streptothricin acetyltransferase from Streptomyces lavendulae. J. Antibiot. 1986, 39, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, C.; Toyoda, J.; Kato, Y.; Izumikawa, M.; Takagi, M.; Shin-ya, K.; Katano, H.; Utagawa, T.; Hamano, Y. A stand-alone adenylation domain forms amide bonds in streptothricin biosynthesis. Nat. Chem. Biol. 2012, 8, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Tang, B.; Dai, R.; Zhang, B.; Chen, L.; Yang, H.; Zhao, G.; Ding, X. Identification of the streptothricin and tunicamycin biosynthetic gene clusters by genome mining in Streptomyces sp. strain fd1-xmd. Appl. Microbiol. Biotechnol. 2018, 102, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Arsic, B.; Barber, J.; Ckos, A.; Mladenovic, M.; Stankovic, N.; Novak, P. 16-membered macrolide antibiotics: A review. Int. J. Antimicrob. Agents 2018, 51, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Katz, L.; Mankin, A.S. Macrolides. In Encyclopedia of Microbiology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 529–558. [Google Scholar]
- Chen, D.; Feng, J.; Huang, L.; Zhang, Q.; Wu, J.; Zhu, X.; Duan, Y.; Xu, Z. Identification and characterization of a new erythromycin biosynthetic gene cluster in Actinopolyspora erythraea YIM90600, a novel erythronolide-producing halophilic actinomycete isolated from salt field. PLoS ONE 2014, 9, e108129. [Google Scholar] [CrossRef] [PubMed]
- Gaisser, S.; Bohm, G.A.; Cortes, J.; Leadlay, P.F. Analysis of seven genes from the eryAI-eryK region of the erythromycin biosynthetic gene cluster in Saccharopolyspora erythraea. Mol. Gen. Genet. 1997, 256, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Oliynyk, M.; Samborskyy, M.; Lester, J.B.; Mironenko, T.; Scott, N.; Dickens, S.; Haydock, S.F.; Leadlay, P.F. Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea NRRL23338. Nat. Biotechnol. 2007, 25, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Olano, C.; Rodriguez, A.M.; Michel, J.M.; Mendez, C.; Raynal, M.C.; Salas, J.A. Analysis of a Streptomyces antibioticus chromosomal region involved in oleandomycin biosynthesis, which encodes two glycosyltransferases responsible for glycosylation of the macrolactone ring. Mol. Gen. Genet. 1998, 259, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.M.; Olano, C.; Vilches, C.; Mendez, C.; Salas, J.A. Streptomyces antibioticus contains at least three oleandomycin-resistance determinants, one of which shows similarity with proteins of the ABC-transporter superfamily. Mol. Microbiol. 1993, 8, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Stratigopoulos, G.; Cundliffe, E. Expression analysis of the tylosin-biosynthetic gene cluster: Pivotal regulatory role of the tylQ product. Chem. Biol. 2002, 9, 71–78. [Google Scholar] [CrossRef]
- Anzai, Y.; Saito, N.; Tanaka, M.; Kinoshita, K.; Koyama, Y.; Kato, F. Organization of the biosynthetic gene cluster for the polyketide macrolide mycinamicin in Micromonospora griseorubida. FEMS Microbiol. Lett. 2003, 218, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Inouye, M.; Horinouchi, S. Establishment of a host-vector system for Micromonospora griseorubida and characterization of mycinamicin biosynthetic genes. Actinomycetologica 1995, 9, 1–12. [Google Scholar] [CrossRef]
- Zhao, L.; Beyer, N.J.; Borisova, S.A.; Liu, H.W. β-Glucosylation as a part of self-resistance mechanism in methymycin/pikromycin producing strain Streptomyces venezuelae. Biochemistry 2003, 42, 14794–14804. [Google Scholar] [CrossRef] [PubMed]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.L.; Levy, S.B. Tetracyclines and tetracycline derivatives. In Comprehensive Biotechnology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 3, pp. 269–283. [Google Scholar]
- Binnie, C.; Warren, M.; Butler, M.J. Cloning and heterologous expression in Streptomyces lividans of Streptomyces rimosus genes involved in oxytetracycline biosynthesis. J. Bacteriol. 1989, 171, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ames, B.D.; Tsai, S.C.; Tang, Y. Engineered biosynthesis of a novel amidated polyketide, using the malonamyl-specific initiation module from the oxytetracycline polyketide synthase. Appl. Environ. Microbiol. 2006, 72, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Donhofer, A.; Franckenberg, S.; Wickles, S.; Berninghausen, O.; Beckmann, R.; Wilson, D.N. Structural basis for TetM-mediated tetracycline resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 16900–16905. [Google Scholar] [CrossRef] [PubMed]
- Dairi, T.; Aisaka, K.; Katsumata, R.; Hasegawa, M. A self-defense gene homologous to tetracycline effluxing gene essential for antibiotic production in Streptomyces aureofaciens. Biosci. Biotechnol. Biochem. 1995, 59, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Connell, S.R.; Tracz, D.M.; Nierhaus, K.H.; Taylor, D.E. Ribosomal protection proteins and their mechanism of tetracycline resistance. Antimicrob. Agents Chemother. 2003, 47, 3675–3681. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Yan, X.; Wang, L.; Chu, J.; Zhuang, Y.; Zhang, S.; Guo, M. Molecular cloning and functional characterization of an ATP-binding cassette transporter OtrC from Streptomyces rimosus. BMC Biotechnol. 2012, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Magarvey, N.; Piraee, M.; Vining, L.C. The gene cluster for chloramphenicol biosynthesis in Streptomyces venezuelae ISP5230 includes novel shikimate pathway homologues and a monomodular non-ribosomal peptide synthetase gene. Microbiology 2001, 147, 2817–2829. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, F.; Lewis, E.A.; Piraee, M.; van Pee, K.H.; Vining, L.C.; White, R.L. Isolation of 3'-O-acetylchloramphenicol: A possible intermediate in chloramphenicol biosynthesis. Bioorg. Med. Chem. Lett. 2002, 12, 283–286. [Google Scholar] [CrossRef]
- Izard, T. Structural basis for chloramphenicol tolerance in Streptomyces venezuelae by chloramphenicol phosphotransferase activity. Protein Sci. 2001, 10, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Mosher, R.H.; Camp, D.J.; Yang, K.; Brown, M.P.; Shaw, W.V.; Vining, L.C. Inactivation of chloramphenicol by O-phosphorylation. A novel resistance mechanism in Streptomyces venezuelae ISP5230, a chloramphenicol producer. J. Biol. Chem. 1995, 270, 27000–27006. [Google Scholar] [CrossRef] [PubMed]
- Mosher, R.H.; Ranade, N.P.; Schrempf, H.; Vining, L.C. Chloramphenicol resistance in Streptomyces: Cloning and characterization of a chloramphenicol hydrolase gene from Streptomyces venezuelae. J. Gen. Microbiol. 1990, 136, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Nakano, H.; Matsuhashi, Y.; Takeuchi, T.; Umezawa, H. Distribution of chloramphenicol acetyltransferase and chloramphenicol-3-acetate esterase among Streptomyces and Corynebacterium. J. Antibiot. 1977, 30, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Shaw, W.V.; Hopwood, D.A. Chloramphenicol acetylation in Streptomyces. J. Gen. Microbiol. 1976, 94, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Schaberle, T.F.; Vollmer, W.; Frasch, H.J.; Huttel, S.; Kulik, A.; Rottgen, M.; von Thaler, A.K.; Wohlleben, W.; Stegmann, E. Self-resistance and cell wall composition in the glycopeptide producer Amycolatopsis balhimycina. Antimicrob. Agents Chemother. 2011, 55, 4283–4289. [Google Scholar] [CrossRef] [PubMed]
- Arthur, M.; Reynolds, P.; Courvalin, P. Glycopeptide resistance in enterococci. Trends Microbiol. 1996, 4, 401–407. [Google Scholar] [CrossRef]
- Marshall, C.G.; Lessard, I.A.; Park, I.; Wright, G.D. Glycopeptide antibiotic resistance genes in glycopeptide-producing organisms. Antimicrob. Agents Chemother. 1998, 42, 2215–2220. [Google Scholar] [PubMed]
- Walsh, C.T.; Fisher, S.L.; Park, I.S.; Prahalad, M.; Wu, Z. Bacterial resistance to vancomycin: Five genes and one missing hydrogen bond tell the story. Chem. Biol. 1996, 3, 21–28. [Google Scholar] [CrossRef]
- Sosio, M.; Kloosterman, H.; Bianchi, A.; de Vreugd, P.; Dijkhuizen, L.; Donadio, S. Organization of the teicoplanin gene cluster in Actinoplanes teichomyceticus. Microbiology 2004, 150, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Pootoolal, J.; Thomas, M.G.; Marshall, C.G.; Neu, J.M.; Hubbard, B.K.; Walsh, C.T.; Wright, G.D. Assembling the glycopeptide antibiotic scaffold: The biosynthesis of A47934 from Streptomyces toyocaensis NRRL15009. Proc. Natl. Acad. Sci. USA 2002, 99, 8962–8967. [Google Scholar] [CrossRef] [PubMed]
- Li, T.L.; Huang, F.; Haydock, S.F.; Mironenko, T.; Leadlay, P.F.; Spencer, J.B. Biosynthetic gene cluster of the glycopeptide antibiotic teicoplanin: Characterization of two glycosyltransferases and the key acyltransferase. Chem. Biol. 2004, 11, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Pelzer, S.; Sussmuth, R.; Heckmann, D.; Recktenwald, J.; Huber, P.; Jung, G.; Wohlleben, W. Identification and analysis of the balhimycin biosynthetic gene cluster and its use for manipulating glycopeptide biosynthesis in Amycolatopsis mediterranei DSM5908. Antimicrob. Agents Chemother. 1999, 43, 1565–1573. [Google Scholar] [PubMed]
- Recktenwald, J.; Shawky, R.; Puk, O.; Pfennig, F.; Keller, U.; Wohlleben, W.; Pelzer, S. Nonribosomal biosynthesis of vancomycin-type antibiotics: A heptapeptide backbone and eight peptide synthetase modules. Microbiology 2002, 148, 1105–1118. [Google Scholar] [CrossRef] [PubMed]
- Beltrametti, F.; Consolandi, A.; Carrano, L.; Bagatin, F.; Rossi, R.; Leoni, L.; Zennaro, E.; Selva, E.; Marinelli, F. Resistance to glycopeptide antibiotics in the teicoplanin producer is mediated by van gene homologue expression directing the synthesis of a modified cell wall peptidoglycan. Antimicrob. Agents Chemother. 2007, 51, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Van Wageningen, A.M.; Kirkpatrick, P.N.; Williams, D.H.; Harris, B.R.; Kershaw, J.K.; Lennard, N.J.; Jones, M.; Jones, S.J.; Solenberg, P.J. Sequencing and analysis of genes involved in the biosynthesis of a vancomycin group antibiotic. Chem. Biol. 1998, 5, 155–162. [Google Scholar] [CrossRef]
- Novotna, G.; Hill, C.; Vincent, K.; Liu, C.; Hong, H.J. A novel membrane protein, VanJ, conferring resistance to teicoplanin. Antimicrob. Agents Chemother. 2012, 56, 1784–1796. [Google Scholar] [CrossRef] [PubMed]
- Sosio, M.; Stinchi, S.; Beltrametti, F.; Lazzarini, A.; Donadio, S. The gene cluster for the biosynthesis of the glycopeptide antibiotic A40926 by Nonomuraea species. Chem. Biol. 2003, 10, 541–549. [Google Scholar] [CrossRef]
- Marcone, G.L.; Beltrametti, F.; Binda, E.; Carrano, L.; Foulston, L.; Hesketh, A.; Bibb, M.; Marinelli, F. Novel mechanism of glycopeptide resistance in the A40926 producer Nonomuraea sp. ATCC 39727. Antimicrob. Agents Chemother. 2010, 54, 2465–2472. [Google Scholar] [CrossRef] [PubMed]
- Binda, E.; Marcone, G.L.; Pollegioni, L.; Marinelli, F. Characterization of VanYn, a novel D,D-peptidase/D,D-carboxypeptidase involved in glycopeptide antibiotic resistance in Nonomuraea sp. ATCC 39727. FEBS J. 2012, 279, 3203–3213. [Google Scholar] [CrossRef] [PubMed]
- Ostash, B.; Doud, E.H.; Lin, C.; Ostash, I.; Perlstein, D.L.; Fuse, S.; Wolpert, M.; Kahne, D.; Walker, S. Complete characterization of the seventeen step moenomycin biosynthetic pathway. Biochemistry 2009, 48, 8830–8841. [Google Scholar] [CrossRef] [PubMed]
- Horbal, L.; Ostash, B.; Luzhetskyy, A.; Walker, S.; Kalinowski, J.; Fedorenko, V. A gene cluster for the biosynthesis of moenomycin family antibiotics in the genome of teicoplanin producer Actinoplanes teichomyceticus. Appl. Microbiol. Biotechnol. 2016, 100, 7629–7638. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.; Nolden, S.; Gebhardt, P.; Heinzelmann, E.; Lange, C.; Puk, O.; Welzel, K.; Wohlleben, W.; Schwartz, D. Sequencing and analysis of the biosynthetic gene cluster of the lipopeptide antibiotic friulimicin in Actinoplanes friuliensis. Antimicrob. Agents Chemother. 2007, 51, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M.; Thompson, C.J.; Kumagai, T.; Suzuki, K.; Deblaere, R.; Villarroel, R.; Davies, J. Characterisation by molecular cloning of two genes from Streptomyces verticillus encoding resistance to bleomycin. Gene 1994, 151, 11–16. [Google Scholar] [PubMed]
- Galm, U.; Wendt-Pienkowski, E.; Wang, L.; George, N.P.; Oh, T.J.; Yi, F.; Tao, M.; Coughlin, J.M.; Shen, B. The biosynthetic gene cluster of zorbamycin, a member of the bleomycin family of antitumor antibiotics, from Streptomyces flavoviridis ATCC 21892. Mol. Biosyst. 2009, 5, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A. On the antibacterial action of cultures of penicillium, with special reference to their use in the isolation of B. influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Martin, J.F.; Ullan, R.V.; Garcia-Estrada, C. Regulation and compartmentalization of β-lactam biosynthesis. Microb. Biotechnol. 2010, 3, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, C.A. Convergent biosynthetic pathways to β-lactam antibiotics. Curr. Opin. Chem. Biol. 2016, 35, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, J.M.; Hodgson, J.E. The biosynthetic genes for clavulanic acid and cephamycin production occur as a ‘super-cluster’ in three Streptomyces. FEMS Microbiol. Lett. 1993, 110, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Trefzer, A.; Kovalchuk, A.; van den Berg, M.; Mueller, U.; Heijne, W.; Wu, L.; Alam, M.T.; Ronning, C.M.; Nierman, W.C.; et al. The sequence of a 1.8-mb bacterial linear plasmid reveals a rich evolutionary reservoir of secondary metabolic pathways. Genome Biol. Evol. 2010, 2, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Barbe, V.; Bouzon, M.; Mangenot, S.; Badet, B.; Poulain, J.; Segurens, B.; Vallenet, D.; Marliere, P.; Weissenbach, J. Complete genome sequence of Streptomyces cattleya NRRL 8057, a producer of antibiotics and fluorometabolites. J. Bacteriol. 2011, 193, 5055–5056. [Google Scholar] [CrossRef] [PubMed]
- Coque, J.J.; Liras, P.; Martin, J.F. Genes for a β-lactamase, a penicillin-binding protein and a transmembrane protein are clustered with the cephamycin biosynthetic genes in Nocardia lactamdurans. EMBO J. 1993, 12, 631–639. [Google Scholar] [PubMed]
- Liras, P. Biosynthesis and molecular genetics of cephamycins. Cephamycins produced by actinomycetes. Antonie Van Leeuwenhoek 1999, 75, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Miyashita, H.; Sumino, Y. Organization and expression in Pseudomonas putida of the gene cluster involved in cephalosporin biosynthesis from Lysobacter lactamgenus YK90. Appl. Microbiol. Biotechnol. 1996, 45, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Barredo, J.L.; Cantoral, J.M.; Alvarez, E.; Diez, B.; Martin, J.F. Cloning, sequence analysis and transcriptional study of the isopenicillin N synthase of Penicillium chrysogenum AS-P-78. Mol. Gen. Genet. 1989, 216, 91–98. [Google Scholar] [CrossRef] [PubMed]
- MacCabe, A.P.; van Liempt, H.; Palissa, H.; Unkles, S.E.; Riach, M.B.; Pfeifer, E.; von Dohren, H.; Kinghorn, J.R. δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine synthetase from Aspergillus nidulans. Molecular characterization of the acvA gene encoding the first enzyme of the penicillin biosynthetic pathway. J. Biol. Chem. 1991, 266, 12646–12654. [Google Scholar] [PubMed]
- Paradkar, A.S.; Aidoo, K.A.; Wong, A.; Jensen, S.E. Molecular analysis of a β-lactam resistance gene encoded within the cephamycin gene cluster of Streptomyces clavuligerus. J. Bacteriol. 1996, 178, 6266–6274. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Hung, T.V.; Liou, K.; Lee, H.C.; Shin, C.H.; Sohng, J.K. Characterization of pbpA and pbp2 encoding penicillin-binding proteins located on the downstream of clavulanic acid gene cluster in Streptomyces clavuligerus. Biotechnol. Lett. 2006, 28, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Ogawara, H. Self-resistance in Streptomyces, with special reference to β-lactam antibiotics. Molecules 2016, 21, E605. [Google Scholar] [CrossRef] [PubMed]
- Reading, C.; Cole, M. Clavulanic acid: A beta-lactamase-inhiting beta-lactam from Streptomyces clavuligerus. Antimicrob. Agents Chemother. 1977, 11, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.E.; Paradkar, A.S.; Mosher, R.H.; Anders, C.; Beatty, P.H.; Brumlik, M.J.; Griffin, A.; Barton, B. Five additional genes are involved in clavulanic acid biosynthesis in Streptomyces clavuligerus. Antimicrob. Agents Chemother. 2004, 48, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Tahlan, K.; Park, H.U.; Jensen, S.E. Three unlinked gene clusters are involved in clavam metabolite biosynthesis in Streptomyces clavuligerus. Can. J. Microbiol. 2004, 50, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, A. Clavulanic acid production by Streptomyces clavuligerus: Biogenesis, regulation and strain improvement. J. Antibiot. 2013, 66, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Kahan, J.S.; Kahan, F.M.; Goegelman, R.; Currie, S.A.; Jackson, M.; Stapley, E.O.; Miller, T.W.; Miller, A.K.; Hendlin, D.; Mochales, S.; et al. Thienamycin, a new β-lactam antibiotic. I. Discovery, taxonomy, isolation and physical properties. J. Antibiot. 1979, 32, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nunez, L.E.; Mendez, C.; Brana, A.F.; Blanco, G.; Salas, J.A. The biosynthetic gene cluster for the β-lactam carbapenem thienamycin in Streptomyces cattleya. Chem. Biol. 2003, 10, 301–311. [Google Scholar] [CrossRef]
- Rodriguez, M.; Nunez, L.E.; Brana, A.F.; Mendez, C.; Salas, J.A.; Blanco, G. Identification of transcriptional activators for thienamycin and cephamycin C biosynthetic genes within the thienamycin gene cluster from Streptomyces cattleya. Mol. Microbiol. 2008, 69, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Sakai, H.; Kohsaka, M.; Konomi, T.; Hosoda, J. Nocardicin A, a new monocyclic β-lactam antibiotic. I. Discovery, isolation and characterization. J. Antibiot. 1976, 29, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Gunsior, M.; Breazeale, S.D.; Lind, A.J.; Ravel, J.; Janc, J.W.; Townsend, C.A. The biosynthetic gene cluster for a monocyclic β-lactam antibiotic, nocardicin A. Chem. Biol. 2004, 11, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Asai, M.; Haibara, K.; Muroi, M.; Kintaka, K.; Kishi, T. Sulfazecin, a novel β-lactam antibiotic of bacterial origin. Isolation and chemical characterization. J. Antibiot. 1981, 34, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Oliver, R.A.; Townsend, C.A. Identification and Characterization of the Sulfazecin Monobactam Biosynthetic Gene Cluster. Cell. Chem. Biol. 2017, 24, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.G.; Chan, Y.A.; Ozanick, S.G. Deciphering tuberactinomycin biosynthesis: Isolation, sequencing, and annotation of the viomycin biosynthetic gene cluster. Antimicrob. Agents Chemother. 2003, 47, 2823–2830. [Google Scholar] [CrossRef] [PubMed]
- Bamas-Jacques, N.; Lorenzon, S.; Lacroix, P.; De Swetschin, C.; Crouzet, J. Cluster organization of the genes of Streptomyces pristinaespiralis involved in pristinamycin biosynthesis and resistance elucidated by pulsed-field gel electrophoresis. J. Appl. Microbiol. 1999, 87, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Mast, Y.; Weber, T.; Goelz, M.; Ort-Winklbauer, R.; Gondran, A.; Wohlleben, W.; Schinko, E. Characterization of the ‘pristinamycin supercluster’ of Streptomyces pristinaespiralis. Microb. Biotechnol. 2011, 4, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.J.; Weisblum, B.; Fahnestock, S.R.; Nomura, M. Alteration of 23 S ribosomal RNA and erythromycin-induced resistance to lincomycin and spiramycin in Staphylococcus aureus. J. Mol. Biol. 1973, 74, 67–72. [Google Scholar] [CrossRef]
- Calcutt, M.J.; Cundliffe, E. Cloning of a lincosamide resistance determinant from Streptomyces caelestis, the producer of celesticetin, and characterization of the resistance mechanism. J. Bacteriol. 1990, 172, 4710–4714. [Google Scholar] [CrossRef] [PubMed]
- Guilfoile, P.G.; Hutchinson, C.R. A bacterial analog of the mdr gene of mammalian tumor cells is present in Streptomyces peucetius, the producer of daunorubicin and doxorubicin. Proc. Natl. Acad. Sci. USA 1991, 88, 8553–8557. [Google Scholar] [CrossRef] [PubMed]
- Furuya, K.; Hutchinson, C.R. The DrrC protein of Streptomyces peucetius, a UvrA-like protein, is a DNA-binding protein whose gene is induced by daunorubicin. FEMS Microbiol. Lett. 1998, 168, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, N.; Doi-Katayama, Y.; Filippini, S.; Nastro, C.; Fonstein, L.; Gallo, M.; Colombo, A.L.; Hutchinson, C.R. The Streptomyces peucetius dpsY and dnrX genes govern early and late steps of daunorubicin and doxorubicin biosynthesis. J. Bacteriol. 1998, 180, 2379–2386. [Google Scholar] [PubMed]
- Menendez, N.; Nur-e-Alam, M.; Brana, A.F.; Rohr, J.; Salas, J.A.; Mendez, C. Biosynthesis of the antitumor chromomycin A3 in Streptomyces griseus: Analysis of the gene cluster and rational design of novel chromomycin analogs. Chem. Biol. 2004, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Lombo, F.; Menendez, N.; Salas, J.A.; Mendez, C. The aureolic acid family of antitumor compounds: Structure, mode of action, biosynthesis, and novel derivatives. Appl. Microbiol. Biotechnol. 2006, 73, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Florez, A.B.; Alvarez, S.; Zabala, D.; Brana, A.F.; Salas, J.A.; Mendez, C. Transcriptional regulation of mithramycin biosynthesis in Streptomyces argillaceus: Dual role as activator and repressor of the PadR-like regulator MtrY. Microbiology 2015, 161, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Steffensky, M.; Muehlenweg, A.; Wang, Z.X.; Li, S.M.; Heide, L. Identification of the novobiocin biosynthetic gene cluster of Streptomyces spheroides NCIB 11891. Antimicrob. Agents Chemother. 2000, 44, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.X.; Li, S.M.; Heide, L. Identification of the coumermycin A1 biosynthetic gene cluster of Streptomyces rishiriensis DSM 40489. Antimicrob. Agents Chemother. 2000, 44, 3040–3048. [Google Scholar] [CrossRef] [PubMed]
- Pojer, F.; Li, S.M.; Heide, L. Molecular cloning and sequence analysis of the clorobiocin biosynthetic gene cluster: New insights into the biosynthesis of aminocoumarin antibiotics. Microbiology 2002, 148, 3901–3911. [Google Scholar] [CrossRef] [PubMed]
- August, P.R.; Flickinger, M.C.; Sherman, D.H. Cloning and analysis of a locus (mcr) involved in mitomycin C resistance in Streptomyces lavendulae. J. Bacteriol. 1994, 176, 4448–4454. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, P.J.; Johnson, D.A.; August, P.R.; Liu, H.W.; Sherman, D.H. Characterization of a mitomycin-binding drug resistance mechanism from the producing organism, Streptomyces lavendulae. J. Bacteriol. 1997, 179, 1796–1804. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Xu, H.; Li, Y.; Zhang, F.; Chen, X.Y.; He, Q.L.; Igarashi, Y.; Tang, G.L. Characterization of yatakemycin gene cluster revealing a radical S-adenosylmethionine dependent methyltransferase and highlighting spirocyclopropane biosynthesis. J. Am. Chem. Soc. 2012, 134, 8831–8840. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Huang, W.; He, Q.L.; Zhao, Z.X.; Zhang, F.; Wang, R.; Kang, J.; Tang, G.L. Self-resistance to an antitumor antibiotic: A DNA glycosylase triggers the base-excision repair system in yatakemycin biosynthesis. Angew. Chem. Int. Ed. Engl. 2012, 51, 10532–10536. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Shan, Y.M.; Li, H.; Dou, W.W.; Jiang, X.H.; Mao, X.M.; Liu, S.P.; Guan, W.J.; Li, Y.Q. Multiple transporters are involved in natamycin efflux in Streptomyces chattanoogensis L10. Mol. Microbiol. 2017, 103, 713–728. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Goswami, A.; Yang, Z.; Liu, X.; Green, K.D.; Barnard-Britson, S. The biosynthesis of capuramycin-type antibiotics: Identification of the A-102395 biosynthetic gene cluster, mechanism of self-resistance, and formation of uridine-5’-carboxamide. J. Biol. Chem. 2015, 290, 13710–13724. [Google Scholar] [CrossRef] [PubMed]
- Funabashi, M.; Nonaka, K.; Yada, C.; Hosobuchi, M.; Masuda, N.; Shibata, T.; Van Lanen, S.G. Identification of the biosynthetic gene cluster of A-500359s in Streptomyces griseus SANK60196. J. Antibiot. 2009, 62, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Naganawa, H.; Hamada, M.; Maeda, K.; Okami, Y.; Takeushi, T.; Umezawa, H. Laspartomycin, a new anti-staphylococcal peptide. J. Antibiot. 1968, 21, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Shen, Q.; Yin, X. Molecular cloning and identification of the laspartomycin biosynthetic gene cluster from Streptomyces viridochromogenes. Gene 2011, 483, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.M.; Huang, T.; Rudolf, J.D.; Smanski, M.J.; Shen, B. Mechanisms of self-resistance in the platensimycin- and platencin-producing Streptomyces platensis MA7327 and MA7339 strains. Chem. Biol. 2014, 21, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Koyama, Y.; Oda, K.; Noda, M.; Matoba, Y.; Sugiyama, M. Molecular cloning and heterologous expression of a biosynthetic gene cluster for the antitubercular agent D-cycloserine produced by Streptomyces lavendulae. Antimicrob. Agents Chemother. 2010, 54, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Kuzuyama, T.; Seto, H. Characterization of the fomA and fomB gene products from Streptomyces wedmorensis, which confer fosfomycin resistance on Escherichia coli. Antimicrob. Agents Chemother. 2000, 44, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Roza, J.; Blanco, M.G.; Hardisson, C.; Salas, J.A. Self-resistance in actinomycetes producing inhibitors of RNA polymerase. J. Antibiot. 1986, 39, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Spanogiannopoulos, P.; Thaker, M.; Koteva, K.; Waglechner, N.; Wright, G.D. Characterization of a rifampin-inactivating glycosyltransferase from a screen of environmental actinomycetes. Antimicrob. Agents Chemother. 2012, 56, 5061–5069. [Google Scholar] [CrossRef] [PubMed]
- Baumann, S.; Schoof, S.; Bolten, M.; Haering, C.; Takagi, M.; Shin-ya, K.; Arndt, H.D. Molecular determinants of microbial resistance to thiopeptide antibiotics. J. Am. Chem. Soc. 2010, 132, 6973–6981. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, V.; Noda-Garcia, L.; Tawfik, D.S.; Fujimori, D.G. Antibiotic resistance evolved via inactivation of a ribosomal RNA methylating enzyme. Nucleic Acids Res. 2016, 44, 8897–8907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, H.W.; Huang, C.C.; Chin, D.H. Thiols screened by the neocarzinostatin protein for preserving or detoxifying its bound enediyne antibiotic. Chemistry 2012, 18, 6238–6249. [Google Scholar] [CrossRef] [PubMed]
- Ahlert, J.; Shepard, E.; Lomovskaya, N.; Zazopoulos, E.; Staffa, A.; Bachmann, B.O.; Huang, K.; Fonstein, L.; Czisny, A.; Whitwam, R.E.; et al. The calicheamicin gene cluster and its iterative type I enediyne PKS. Science 2002, 297, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Whitwam, R.E.; Ahlert, J.; Holman, T.R.; Ruppen, M.; Thorson, J.S. The gene calC encodes for a non-heme iron metalloprotein responsible for calicheamicin self-resistance in Micromonospora. J. Am. Chem. Soc. 2000, 122, 1556–1557. [Google Scholar] [CrossRef]
- Shen, B.; Yan, X.; Huang, T.; Ge, H.; Yang, D.; Teng, Q.; Rudolf, J.D.; Lohman, J.R. Enediynes: Exploration of microbial genomics to discover new anticancer drug leads. Bioorg. Med. Chem. Lett. 2015, 25, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lohman, J.R.; Huang, S.X.; Horsman, G.P.; Dilfer, P.E.; Huang, T.; Chen, Y.; Wendt-Pienkowski, E.; Shen, B. Cloning and sequencing of the kedarcidin biosynthetic gene cluster from Streptoalloteichus sp. ATCC 53650 revealing new insights into biosynthesis of the enediyne family of antitumor antibiotics. Mol. Biosyst. 2013, 9, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.L.; Nam, S.J.; Fukuda, T.; Yamanaka, K.; Kauffman, C.A.; Jensen, P.R.; Fenical, W.; Moore, B.S. Structures and comparative characterization of biosynthetic gene clusters for cyanosporasides, enediyne-derived natural products from marine actinomycetes. J. Am. Chem. Soc. 2013, 135, 4171–4174. [Google Scholar] [CrossRef] [PubMed]
- Kale, A.J.; McGlinchey, R.P.; Lechner, A.; Moore, B.S. Bacterial self-resistance to the natural proteasome inhibitor salinosporamide A. ACS Chem. Biol. 2011, 6, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Daduang, R.; Kitani, S.; Hashimoto, J.; Thamchaipenet, A.; Igarashi, Y.; Shin-ya, K.; Ikeda, H.; Nihira, T. Characterization of the biosynthetic gene cluster for maklamicin, a spirotetronate-class antibiotic of the endophytic Micromonospora sp. NBRC 110955. Microbiol. Res. 2015, 180, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Foulston, L.C.; Bibb, M.J. Microbisporicin gene cluster reveals unusual features of lantibiotic biosynthesis in actinomycetes. Proc. Natl. Acad. Sci. USA 2010, 107, 13461–13466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, K.; Ohata, Y.; Shoda, M. Gene yerP, involved in surfactin self-resistance in Bacillus subtilis. Antimicrob. Agents Chemother. 2001, 45, 3566–3573. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.; Wu, Q.; Xu, Y. Genome and transcriptome analysis of surfactin biosynthesis in Bacillus amyloliquefaciens MT45. Sci. Rep. 2017, 7, 40976. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Entian, K.D. Genes involved in self-protection against the lantibiotic subtilin produced by Bacillus subtilis ATCC 6633. Appl. Environ. Microbiol. 1994, 60, 2793–2801. [Google Scholar] [PubMed]
- Wang, H.; van der Donk, W.A. Substrate selectivity of the sublancin S-glycosyltransferase. J. Am. Chem. Soc. 2011, 133, 16394–16397. [Google Scholar] [CrossRef] [PubMed]
- Emmert, E.A.; Klimowicz, A.K.; Thomas, M.G.; Handelsman, J. Genetics of zwittermicin a production by Bacillus cereus. Appl. Environ. Microbiol. 2004, 70, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ruan, L.F.; Zhao, C.M.; Wang, C.X.; Peng, D.H.; Sun, M. Validation of the intact zwittermicin A biosynthetic gene cluster and discovery of a complementary resistance mechanism in Bacillus thuringiensis. Antimicrob. Agents Chemother. 2011, 55, 4161–4169. [Google Scholar] [CrossRef] [PubMed]
- Neumuller, A.M.; Konz, D.; Marahiel, M.A. The two-component regulatory system BacRS is associated with bacitracin ‘self-resistance’ of Bacillus licheniformis ATCC 10716. Eur. J. Biochem. 2001, 268, 3180–3189. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Park, S.Y.; Kim, R.; Kim, S.B.; Lee, C.H.; Kim, J.F.; Park, S.H. Identification of a polymyxin synthetase gene cluster of Paenibacillus polymyxa and heterologous expression of the gene in Bacillus subtilis. J. Bacteriol. 2009, 191, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Westman, E.L.; Yan, M.; Waglechner, N.; Koteva, K.; Wright, G.D. Self resistance to the atypical cationic antimicrobial peptide edeine of Brevibacillus brevis Vm4 by the N-acetyltransferase EdeQ. Chem. Biol. 2013, 20, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Kamigiri, K.; Suzuki, Y.; Shibazaki, M.; Morioka, M.; Suzuki, K.; Tokunaga, T.; Setiawan, B.; Rantiatmodjo, R.M. Kalimantacins A, B and C, novel antibiotics from Alcaligenes sp. YL-02632S. I. Taxonomy, fermentation, isolation and biological properties. J. Antibiot. 1996, 49, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Mattheus, W.; Gao, L.J.; Herdewijn, P.; Landuyt, B.; Verhaegen, J.; Masschelein, J.; Volckaert, G.; Lavigne, R. Isolation and purification of a new kalimantacin/batumin-related polyketide antibiotic and elucidation of its biosynthesis gene cluster. Chem. Biol. 2010, 17, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Mattheus, W.; Masschelein, J.; Gao, L.J.; Herdewijn, P.; Landuyt, B.; Volckaert, G.; Lavigne, R. The kalimantacin/batumin biosynthesis operon encodes a self-resistance isoform of the FabI bacterial target. Chem. Biol. 2010, 17, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, T.; Kawakami, M. How does Pseudomonas fluorescens avoid suicide from its antibiotic pseudomonic acid? Evidence for two evolutionarily distinct isoleucyl-tRNA synthetases conferring self-defense. J. Biol. Chem. 2003, 278, 25887–25894. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Baker, A.T.; Raab, A.; Huang, S.; Wang, T.; Yu, Y.; Jaspars, M.; Secombes, C.J.; Deng, H. The fish pathogen Yersinia ruckeri produces holomycin and uses an RNA methyltransferase for self-resistance. J. Biol. Chem. 2013, 288, 14688–14697. [Google Scholar] [CrossRef] [PubMed]
- Kenig, M.; Reading, C. Holomycin and an antibiotic (MM 19290) related to tunicamycin, metabolites of Streptomyces clavuligerus. J. Antibiot. 1979, 32, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Forseth, R.R.; Bowers, A.A.; Schroeder, F.C.; Walsh, C.T. A backup plan for self-protection: S-methylation of holomycin biosynthetic intermediates in Streptomyces clavuligerus. ChemBioChem 2012, 13, 2521–2526. [Google Scholar] [CrossRef] [PubMed]
- Chopra, S.; Palencia, A.; Virus, C.; Schulwitz, S.; Temple, B.R.; Cusack, S.; Reader, J. Structural characterization of antibiotic self-immunity tRNA synthetase in plant tumour biocontrol agent. Nat. Commun. 2016, 7, 12928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.G.; Genee, H.J.; Kaas, C.S.; Nielsen, J.B.; Regueira, T.B.; Mortensen, U.H.; Frisvad, J.C.; Patil, K.R. A new class of IMP dehydrogenase with a role in self-resistance of mycophenolic acid producing fungi. BMC Microbiol. 2011, 11, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regueira, T.B.; Kildegaard, K.R.; Hansen, B.G.; Mortensen, U.H.; Hertweck, C.; Nielsen, J. Molecular basis for mycophenolic acid biosynthesis in Penicillium brevicompactum. Appl. Environ. Microbiol. 2011, 77, 3035–3043. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L. How do antibiotic-producing microorganisms avoid suicide? Ann. N. Y. Acad. Sci. 1974, 235, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Vining, L.C. Antibiotic tolerance in producer organisms. Adv. Appl. Microbiol. 1979, 25, 147–168. [Google Scholar] [PubMed]
- Cundliffe, E. How antibiotic-producing organisms avoid suicide. Annu. Rev. Microbiol. 1989, 43, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Cundliffe, E.; Demain, A.L. Avoidance of suicide in antibiotic-producing microbes. J. Ind. Microbiol. Biotechnol. 2010, 37, 643–672. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.; Wright, G.D. Intrinsic antibiotic resistance: Mechanisms, origins, challenges and solutions. Int. J. Med. Microbiol. 2013, 303, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Olivares, J.; Bernardini, A.; Garcia-Leon, G.; Corona, F.; Sanchez, M.B.; Martinez, J.L. The intrinsic resistome of bacterial pathogens. Front. Microbiol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Culyba, M.J.; Mo, C.Y.; Kohli, R.M. Targets for combating the evolution of acquired antibiotic resistance. Biochemistry 2015, 54, 3573–3582. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Gao, G.F.; Zhu, B. The antibiotic resistome: Gene flow in environments, animals and human beings. Front. Med. 2017, 11, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Ying, G.G.; Singer, A.C.; Zhu, Y.G. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Wencewicz, T.A. New antibiotics from nature’s chemical inventory. Bioorg. Med. Chem. 2016, 24, 6227–6252. [Google Scholar] [CrossRef] [PubMed]
- Matano, L.M.; Morris, H.G.; Wood, B.M.; Meredith, T.C.; Walker, S. Accelerating the discovery of antibacterial compounds using pathway-directed whole cell screening. Bioorg. Med. Chem. 2016, 24, 6307–6314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltz, R.H. Molecular beacons to identify gifted microbes for genome mining. J. Antibiot. 2017, 70, 639–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.K.; Koeris, M.S. The next generation of bacteriophage therapy. Curr. Opin. Microbiol. 2011, 14, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Donovan, D.M. Antimicrobial bacteriophage-derived proteins and therapeutic applications. Bacteriophage 2015, 5, e1062590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiGiandomenico, A.; Sellman, B.R. Antibacterial monoclonal antibodies: The next generation? Curr. Opin. Microbiol. 2015, 27, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Oleksiewicz, M.B.; Nagy, G.; Nagy, E. Anti-bacterial monoclonal antibodies: Back to the future? Arch. Biochem. Biophys. 2012, 526, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Quintanilla, M.; Pulido, M.R.; Carretero-Ledesma, M.; McConnell, M.J. Vaccines for antibiotic-resistant bacteria: Possibility or pipe dream? Trends Pharmacol. Sci. 2016, 37, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M.; Siber, G.R. How can vaccines contribute to solving the antimicrobial resistance problem? MBio 2016, 7, e00428-16. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018. [Google Scholar] [CrossRef] [PubMed]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Fortman, J.L.; Mukhopadhyay, A. The future of antibiotics: Emerging technologies and stewardship. Trends Microbiol. 2016, 24, 515–517. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaeberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Iscla, I.; Wray, R.; Blount, P.; Larkins-Ford, J.; Conery, A.L.; Ausubel, F.M.; Ramu, S.; Kavanagh, A.; Huang, J.X.; Blaskovich, M.A.; et al. A new antibiotic with potent activity targets MscL. J. Antibiot. 2015, 68, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, P.; Martens, E. Antibiotics in late clinical development. Biochem. Pharmacol. 2017, 133, 152–163. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Construction of human voltage-gated sodium channel 1.4. (B) Comparison of amino acid residues of human Nav1.4, human Nav1.5, Fugu Nav1.4a, Tetraodon Nav1.4a, and Mya arenaria Nav. The amino acid numbering is according to that of human Nav1.4. The GenBank accession numbers are human Nav1.4: P35499; human Nav1.5: Q14524; fugu Nav1.4a: ABB29441; Tetraodon Nav1.4a: ABB29443; and Mya arenaria Nav: AAX14719. The amino acids in alanine (A) in DIV (DEKA) are marked with red, those in two aspartate residues in DIII and DIV (EEDD) are marked with light blue, and the amino acid residues at 407 are marked with dark blue.
Figure 1.
(A) Construction of human voltage-gated sodium channel 1.4. (B) Comparison of amino acid residues of human Nav1.4, human Nav1.5, Fugu Nav1.4a, Tetraodon Nav1.4a, and Mya arenaria Nav. The amino acid numbering is according to that of human Nav1.4. The GenBank accession numbers are human Nav1.4: P35499; human Nav1.5: Q14524; fugu Nav1.4a: ABB29441; Tetraodon Nav1.4a: ABB29443; and Mya arenaria Nav: AAX14719. The amino acids in alanine (A) in DIV (DEKA) are marked with red, those in two aspartate residues in DIII and DIV (EEDD) are marked with light blue, and the amino acid residues at 407 are marked with dark blue.

Figure 2.
(A) Amino acid sequence of α-bungarotoxin (GenBank accession No. P01378). Amino acids involved in the binding to thr acetylcholine receptor are marked with bold red. Red underlines indicate loop II and loop III, respectively. (B) Comparison of amino acid sequences of human α2-acetylcholine receptor (GenBank accession No. CAD89000) and Torpedo marmorata α-acetylcholine receptor (GenBank accession No. AAA96704). Amino acid residues involved in the ligand-binding are marked with bold red. Wavy red underlines show the trans-membrane regions. (C). Comparison of critical amino acid residues of acetylcholine receptors from species that are sensitive or resistant to α-bungarotoxin. Dots indicate the same amino acid residues as that of a mouse.
Figure 2.
(A) Amino acid sequence of α-bungarotoxin (GenBank accession No. P01378). Amino acids involved in the binding to thr acetylcholine receptor are marked with bold red. Red underlines indicate loop II and loop III, respectively. (B) Comparison of amino acid sequences of human α2-acetylcholine receptor (GenBank accession No. CAD89000) and Torpedo marmorata α-acetylcholine receptor (GenBank accession No. AAA96704). Amino acid residues involved in the ligand-binding are marked with bold red. Wavy red underlines show the trans-membrane regions. (C). Comparison of critical amino acid residues of acetylcholine receptors from species that are sensitive or resistant to α-bungarotoxin. Dots indicate the same amino acid residues as that of a mouse.

Figure 3.
Comparison of amino acid sequences in site 3 of the human sodium channels and that of Haplopelma schmidti (OhNav1). The critical residues are marked with blue and red. The amino acid numbering is according to OhNav1.
Figure 3.
Comparison of amino acid sequences in site 3 of the human sodium channels and that of Haplopelma schmidti (OhNav1). The critical residues are marked with blue and red. The amino acid numbering is according to OhNav1.

Figure 4.
Comparison of the amino acid sequences of CPT is of the CPT-producing and CPT-nonproducing plants and humans. Ophiorrhiza liukiuensis, Camptotheca acuminate, and Ophiorrhiza pumila are CPT-producing plants, while Ophiorrhiza japonica and Homo sapiens are nonproducing. GenBank accession numbers are BAG31374, BAG31376, BAG31373, BAG31375, and AAA61206, respectively. The critical amino acid residues for the resistance (aspragine and serine) were marked with red.
Figure 4.
Comparison of the amino acid sequences of CPT is of the CPT-producing and CPT-nonproducing plants and humans. Ophiorrhiza liukiuensis, Camptotheca acuminate, and Ophiorrhiza pumila are CPT-producing plants, while Ophiorrhiza japonica and Homo sapiens are nonproducing. GenBank accession numbers are BAG31374, BAG31376, BAG31373, BAG31375, and AAA61206, respectively. The critical amino acid residues for the resistance (aspragine and serine) were marked with red.

Figure 5.
Comparison of amino acid residues of three isoforms of Oncopeltus fasciatus Na+/K+-ATPases, and those of Drosophila melanogaster and Homo sapiens. The critical residues are marked with red. GenBank accession numbers are AFU25689, AFU25688, AFU25687, AAC05260, and NP_000692, respectively. The critical amino acid residues for the binding of ouabain were marked with red.
Figure 5.
Comparison of amino acid residues of three isoforms of Oncopeltus fasciatus Na+/K+-ATPases, and those of Drosophila melanogaster and Homo sapiens. The critical residues are marked with red. GenBank accession numbers are AFU25689, AFU25688, AFU25687, AAC05260, and NP_000692, respectively. The critical amino acid residues for the binding of ouabain were marked with red.

Figure 6.
The relationship between predators and prey, and herbivores and plant.


{kind=link}
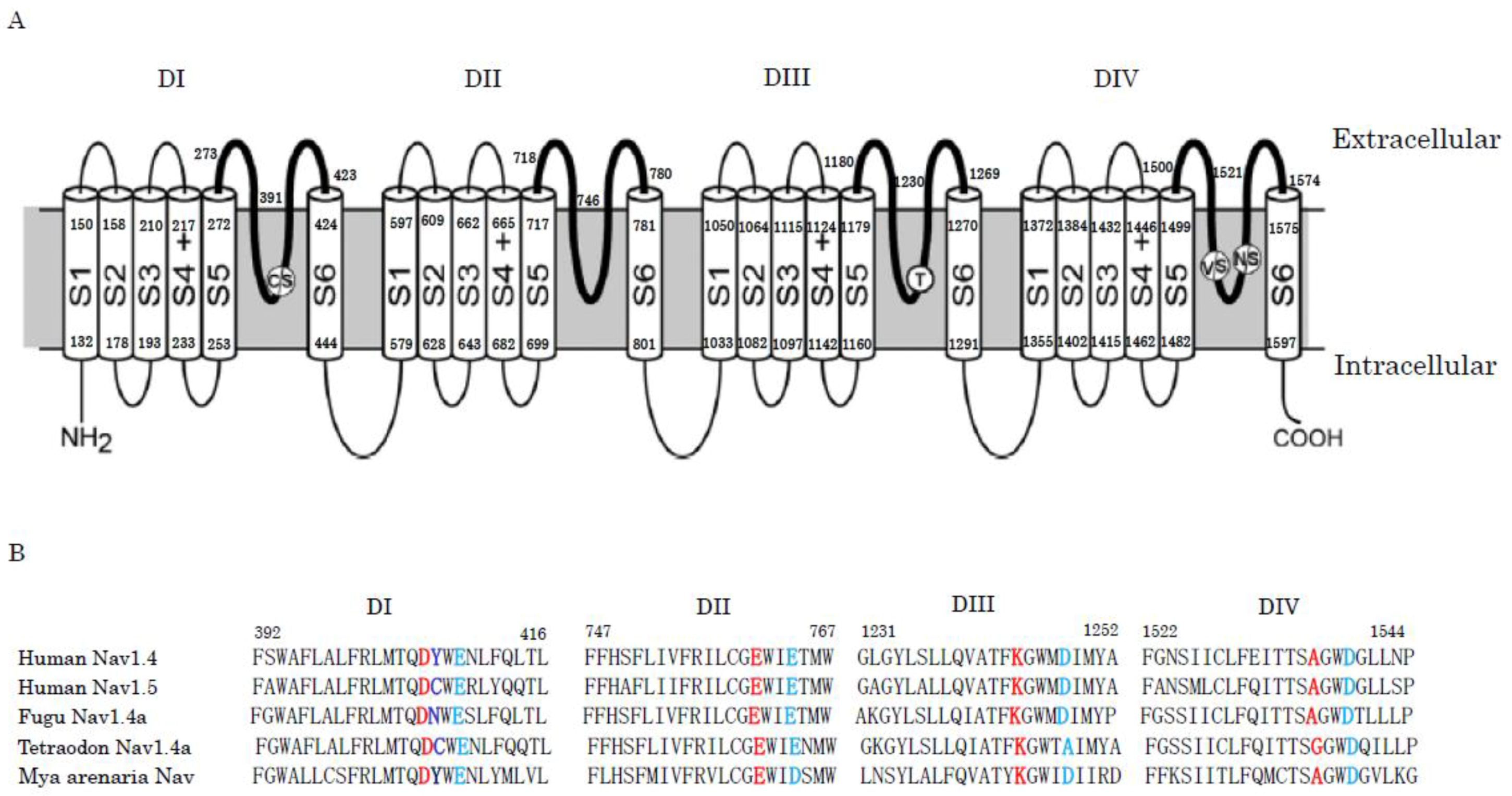{kind=link}
{kind=link}
{kind=link}
{kind=link}
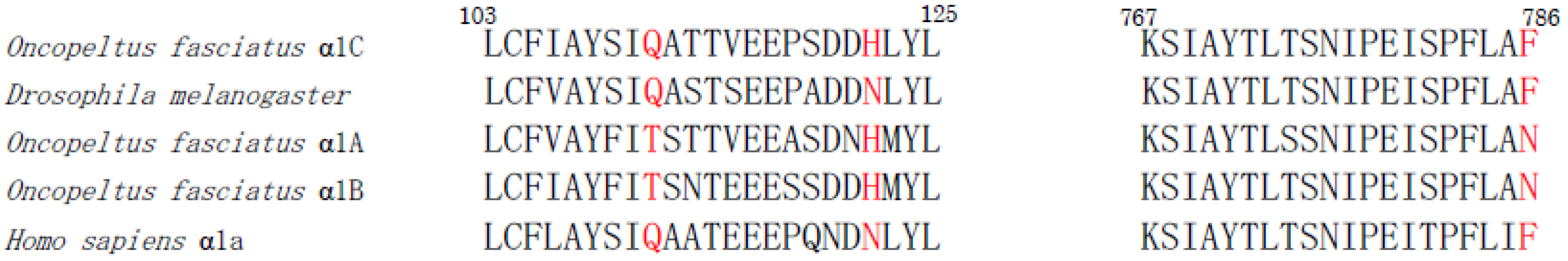{kind=link}
{kind=link}
Table 1.
Resistance-related genes in biosynthetic gene clusters of phycotoxins and related compounds.
Table 1.
Resistance-related genes in biosynthetic gene clusters of phycotoxins and related compounds.
| Toxin | Producing Species | GenBank Accession No. *1 | Resistance-Related Gene/Protein | GenBank Accession No. *2 | Reference |
|---|---|---|---|---|---|
| Cylindrospermopsin | Cylindrospermopsis raciborskii AWT205 | EU140798 | Multi-drug exporter: CyrK | ABX60156 (465aa) | [48] |
| Aphanizomenon sp. strain 10E6 | GQ385961 | Multi-drug exporter: CyrK | ADF88272 (479aa) | [49] | |
| Oscillatoria sp. strain PCC 6506 | FJ418586 | Multi-drug exporter: CyrK | ADI48264 (479aa) | [47] | |
| Jamaicamide A | Lyngbya majuscula | AY522504 | No resistance-related gene | [50] | |
| Hectochlorin | Lyngbya majuscula | AY974560 | No resistance-related gene | [53] | |
| Curacin A | Lyngbya majuscula strain 19L | AY652953 | No resistance-related gene | [54] | |
| Moorea producens 3L | HQ696500 | No resistance-related gene | [55] | ||
| Apratoxin A | Lyngbya (Moorea) bouillonii PNG5-198 | MKZS01000001 | No resistance-related gene | [57,58] | |
| Lyngbyatoxin | Lyngbya majuscula | AY588942 | No resistance-related gene | [59,60,61] | |
| Teleocidin B | Streptomyces blastmyceticus NBRC 12747 | AB937114 | ABC transporters: Orf1, Orf2, Orf3 | BAP27936-BAP27938 | [62] |
| Microcystins | Microcystis aeruginosa PCC 7806 | AF183408 | ABC transporter: McyH | AAF00956 (538aa) | [65,66] |
| Mutation of target: phosphatase? | [68] | ||||
| Nodularin | Nodularia spumigena | AY210783 | ABC transporter: NdaI | AAO64410 (601aa) | [64] |
| Saxitoxins | Cylindrospermopsis raciborskii T3 | DQ787200 | MATE: SxtF MATE: SxtM | ABI75096(471aa) ABI75103 (482aa) | [74,75] |
| Anabaena circinalis AWQC131C | DQ787201 | MATE: SxtM | ABI75138 (485aa) | [75] | |
| Aphanizomenon sp. NH5 | EU603710 | MATE: SxtM | ACG63815 (485aa) | [75] | |
| Lyngbya wollei | EU603711 | MATE: SxtM1 MATE: SxtM2 MATE: SxtM3 | ACG63829 (479aa) ACG63832 (485aa) ACZ26231 (503aa) | [76] | |
| Raphidiopsis brookii D9 | ACYB00000000 | MATE: SxtF (CRD_02147) MATE: SxtM (CRD_02155) | WP_009343300 (471aa) WP_040553734 (475aa) | [77] | |
| (Mya arenaria) | Mutation of target: sodium channel | AAX14719 (1435aa, partial) | [72] | ||
| Anatoxin A | Oscillatoria sp. PCC6506 | FJ477836 | AnaI: MATE-like transporter | AMO66168 (466aa) | [88,89] |
| Anabaena flos-aquae 37 | JF803645 | No resistance-related gene | [90] | ||
| Hapalindole | Fischerella sp. ATCC 43239 | KJ742064 | No resistance-related gene | [96] | |
| Fischerella sp. PCC 9339 | ABC transporter: Orf2 (277aa) ABC transporter: Orf3 (298aa) ABC transporter: Orf4 (331aa) | IMG Gene ID: 2517064626 IMG Gene ID: 2517064627 IMG Gene ID: 2517064628 | [96] | ||
| Ambiguine | Fischerella ambigua UTEX 1903 | KJ742065 KF664586 | Efflux pump: AmbE1 Efflux pump: AmbE2 Efflux pump: AmbE3 | AIJ28573 (388aa) AHB62754 (388aa) AIJ28574 (397aa) AHB62753 (397aa) AIJ28575 (151aa) AHB62752 (151aa) | [95,96] |
| Welwitindolinone | Hapalosiphon welwitschii UH IC-52-3 | KJ767017 | Multidrug resistance protein: WelE4 | AIH14769 (105aa) | [96] |
| Hapalosiphon welwitschii UTEX B1830 | KF811479 | No resistance-related gene | [94] | ||
| Westiella intricate UH HT-29-1 | KJ767018 | Multidrug resistance protein: WelE4 | AIH14815 (105aa) | [96] | |
| Monensin | Streptomyces cinnnamonensis | AF440781 | Efflux protein: MonT | ANZ52456 | [103,104] |
| Palytoxin | Palythora, Ostreopsis, Trichodesmium | Sequestration? | [108] | ||
| Okadaic acid | Protocentrum | sulfated diesters? | [109] | ||
| Domoic acid | Pseudo-nitzschia australis | SLC6 transporter? | [116,117] | ||
| Patellamides | Prochloron diene | AY986476 | No resistance-related gene | [123] | |
| ABC transporter? | [124] | ||||
| Microcin B | Pseudomonas antarctica PAMC 27494 | CP015600 | ABC transporter: McbE ABC transporter: McbF | ANF87042 (237aa) ANF87073 (250aa) | [131] |
| Microcin C7 | Escherichia coli | X57583 | Efflux pump: MccC Acetyltransferase: MccE Self-immunity protein: MccF | CAA40810 (404aa) CAA40813 (521aa) CAA40814 (344aa) | [132,133,134] |
| Goadsporin | Streptomyces sp. TP-A0584 | AB205012 | ABC transporter: GodB ABC transporter: GodC Acetyltransferase: GodH | BAE46917 (550aa) BAE46948 (557aa) BAE46923 (222aa) | [129,135] |
*1: GenBank accession number for the biosynthetic gene cluster of the toxin. *2: GenBank accession number for the resistance-related gene.
Table 2.
Toxin resistance-related strategies in marine and terrestrial animals.
| Toxin | Strategies | Animals | Reference |
|---|---|---|---|
| Tetrodotoxin | Mutation of target (Nav) | Fugu | [147,149] |
| Newt | [144,145,148] | ||
| Snake | [144,145,148] | ||
| Saxitoxin | Mutation of target (Nav) | Softshell clam | [72] |
| Conotoxin/Conopeptide | Mutation of target (Nav) | Conus | [154] |
| Sequestration | Conus | [157,164] | |
| Post-traslational modification | Conus | [160,161,162,163] | |
| Actinoporin | Modification of target (sphingomyelin) | Sea anemone | [168] |
| Hydralysin | Lack of receptor | Hydra | [169] |
| α-Bungarotoxin | Mutation and modification of target | Snake, mongoose, hedgehog, human | [180,181,182,183] |
| Atrolysins | Inhibition by serum | Opossum | [187] |
| Rattlesnake | [188,189] | ||
| Botrocetin | Mutation of target (von Willebrand factor) | Opossum | [190] |
| Micrurotoxins | Mutation of target (GABAA receptor) | Coral snake | [191] |
| LqTx (Scorpion α-toxin) | Mutation of target (Nav) | Scorpion | [199,200] |
| CssIV (Scorpion β-toxin) | Mutation of target (Nav) | Scorpion | [203,204] |
| Scorpion toxins | Mutation of target (Nav)? | Bat | [205,206] |
| Huwentoxin-IV (Tarantula toxin) | Mutation of target (Nav) | Tarantula | [215,216] |
| Jingzhaotoxin-I (Tarantula toxin) | Mutation of target (Nav) | Human | [218,219] |
| Lipophilic alkaloids | Sequestration | Frog | [223] |
| Batrachotoxin | Mutation of target (Nav) | Frog | [225,226] |
Table 3.
Toxin resistance-related strategies in plants.
| Toxin | Strategies | Plant/Animal | Reference |
|---|---|---|---|
| Camptothecin | Mutation of target (Topo I) | Ophiorrhiza japonica | [233,234] |
| Sclareol | ABC transporter (NpPDR1) | Nicotiana plumbaginifolia | [235,236,237] |
| Berberine (Benzylisoquinoline alkaloid) | Excretion, ABC transporter | Thalictrum minus | [243] |
| Sequestration, ABC transporter (CjABCB1) | Coptis japonica | [242,244] | |
| Catharanthine (Indole alkaloid) | ABC transporter (CrTPT2) | Catharanthus roseus | [248] |
| Flavonoids | Sequestration, ABC transporter | Arabidopsis | [241,250] |
| Sanguinarine (Benzophenanthridine alkaloid) | Sequestration | Papaver somniferum | [252,253] |
| Chemical modification | Eschscholzia californica | [254,255,256] | |
| Coniine (Piperidine alkaloid) | Detoxication by cytochrome P450 | Trichoplusia ni | [259] |
| Pyrrolizidine alkaloids | Sequestration as N-oxides | Senecio | [264] |
| Sequestration as N-oxides | Utetheisa ornatrix | [265,266] | |
| Steroidal glycoalkaloids | Deglycosylation to aglycones | Solanaceae | [269,270,271,275] |
| Deglycosylation to aglycones | Fusarium oxysporum, Cladosporium fulvum | [272,273] | |
| Benzoxazinoids | Glycosylation | Secale cereale | [276] |
| Iridoid glucosides | Glycosylation | Plantago lanceolata | [277,278] |
| Cyanogenic glucosides | Glycosylation | Prunus | [279,280] |
| Glucosinolates | Glycosylation, Myrosinase | Brassicaceae | [278,281] |
| Cardenolides | High K+ concentration | Lepidopteran | [282] |
| Sequestration | Lepidopteran | [283] | |
| Mutation of target (Na/K+-ATPase) | Danaus plexippus, Oncopeltus fasciatus | [284,285,286,287] | |
| Sequestration/Exclusion, ABC transporter | Daphnis nerii | [288] | |
| Nicotine | Exclusion/Cytochrome P-450 | Manduca sexta | [290] |
| Sequestration | Nicotiana | [291] | |
| α-DIOXYGENASE1 | Nicotiana attenuata/Manduca sexta | [296] | |
| Menthol (Monoterpene) | Sequestration | Mentha canadensis | [298] |
| Quinolizidine alkaloids | Sequestration | Lupinus | [299,301] |
| Sequestration | Macrosiphum albifrons | [301] |
Table 4.
Toxin resistance-related strategies in fungi.
| Toxin | Strategies | Fungi/Plant | Reference |
|---|---|---|---|
| Aflatoxin | Transporter, Hydroxylation | Aspergillus flavus, Aspergillus parasiticus | [304,305,306,307] |
| Sterigmatocystin | Not defined | Aspergillus nidulans | [309,310] |
| Ochratoxins | Transporter | Aspergillus, Penicillium | [312,313] |
| Citrinin | Transporter | Monascus aurantiacus, Monascus purpureus | [317,318] |
| Deoxynivalenol/Trichothecene | Glutathione-S-transferase, ABC transporter | Barley (plant) | [324] |
| Trichothecenes | Transporter | Fusarium graminearum | [325] |
| Zearalenone/Trichothecene | Monocarboxylate transporter | Fusarium graminearum/Gibberella zeae | [329,330] |
| Lactonohydrolase | Clonostachys rosea | [331,332] | |
| Fumonisins | Transporter | Fusarium verticillioides | [333,334] |
| Transporter | Aspergillus niger, A. welwitschiae | [335,336] | |
| Penitrem A | Transporter | Penicillium crustosum, P. simplicissimum | [337] |
| Roquefortine C | Transporter | Penicillium chrysogenum | [339] |
| Aphidicolin | Transporter | Cephalosporium aphidicola | [340] |
| Sirodesmin PL | Transporter | Leptosphaeris maculans | [342,343] |
| Gliotoxin | Transporter, Reductase | Aspergillus fumigatus | [342,344,345] |
| Ergot alkaloids | Not defined | Family Clavicipitaceae | [347,348,349,350,351,354] |
| Loline | Not defined | Neotyphodium/Epichloae/Endophyte | [356,358,359,360] |
| Lolitrem B | Not defined | Neotyphodium/Epichloae/Endophyte | [363,364] |
| Peramine | Transporter | Epichloae festucae | [362] |
| Swainsonine | Transporter | Orders Hypocreales, Chaetothyriales and others | [365] |
Table 5.
Antibiotic resistance-related strategies in bacteria and fungi.
| Antibiotic | Strategies | Bacteria/Fungi | Reference |
|---|---|---|---|
| Streptomycin | Phosphorylation, Transporter | Streptomyces griseus | [377,378,379] |
| Kanamycin | Acetylation, rRNA methylation, Transporter | Streptomyces kanamyceticus | [381,382,383,384] |
| Gentamicin | rRNA methylation, Transporter | Micromonospora echinospora | [381] |
| Tobramycin | Phosphorylation, Acetylation, Transporter | Streptoalloteichus hindustanus | [381,386] |
| Neomycin | Phosphorylation, Acetylation, Transporter | Streptomyces fradiae | [387] |
| Paromomycin | Phosphorylation, Acetylation, Transporter | Streptomyces rimosus | [388] |
| Lividomycin | Transporter | Streptomyces lividus | AJ748832 *1 |
| Hygromycin B | Phosphorylation | Streptomyces hygroscopicus | [389] |
| Hygromycin A | Phosphorylation, Transporter | Streptomyces hygroscopicus | [390,391] |
| Istamycin | rRNA methylation | Streptomyces tenjimariensis | [393] |
| Kasugamycin | Transporter, Acetylation | Streptomyces kasugaensis | [395] |
| Fortimicin/astromicin | rRNA methylation | Micromonospora olivasterospora | [396] |
| Streptothricin | Acetylation, Transporter | Streptomyces lavendulae | [397,398,399] |
| Erythromycin | rRNA methylation, Phosphorylation, Acetylation, Transporter | Saccharopolyspora erythraea | [402,403,404] |
| Oleandomycin | Glycosylation, Transporter | Streptomyces antibioticus | [405,406] |
| Tylosin | rRNA methylation, Transporter | Streptomyces fradiae | [407] |
| Mycinamicin | rRNA methylation | Micromonospora griseorubida | [408] |
| Methymycin | rRNA methylation, Glycosylation | Streptomyces venezuelae | [410] |
| Oxytetracycline | Ribosome protection, Transporter | Streptomyces rimosus | [413,414,415,417] |
| Chlortetracycline | Ribosome protection, Transporter | Kitasatospora aureofaciens | [416,417] |
| Chloramphenicol | Phosphorylation, Transporter, Acetylation, Hydrolase | Streptomyces venezuelae | [420,421,422,423,424] |
| Vancomycin | Transporter, Redirection of peptidoglycan biosynthesis | Amycolatopsis orientalis | [428,430], HE589771 *1 |
| Balhimycin | Transporter, Redirection of peptidoglycan biosynthesis | Amycolatopsis mediterranei | [428,435,436] |
| Chloroeremomycin | Transporter | Amycolatopsis orientalis | [438] |
| Teicoplanin | Transporter, Redirection of peptidoglycan biosynthesis | Actinoplanes teichomyceticus | [432,434,437,439] |
| A40926 | d,d-carboxypeptidase | Nonomuraea species | [440,441,442] |
| Moenomycin | Transporters | Streptomyces ghanaensis | [443,444] |
| Friulimicin | Transporters | Actinoplanes friuliensis | [445] |
| Bleomycin | Acetylation, Bleomycin-binding protein | Streptomyces verticillus | [446] |
| Zorbamycin | Zorbamycin-binding protein, Transporter | Streptomyces flavoviridis | [447] |
| Penicillin N/Cephamycin C | β-Lactamase, Penicillin-binding protein, Transporter | Streptomyces clavuligerus | [451,452] |
| β-Lactamase, Penicillin-binding protein, Transporter | Streptomyces cattleya | [453] | |
| β-Lactamase, Penicillin-binding protein, Transporter | Nocardia lactamdurans | [454,455] | |
| β-Lactamase, Transporter | Lysobacter lactamgenus | [456], X56660 *1 | |
| Penicillin G | Transporter? | Penicillium chrysogenum | [457], AM920436 *1 |
| Transporter? | Aspergillus nidulans | [458], X54853 *1 | |
| Cephalosporin C | Unknown | Acremonium chrysogenum | AJ404737 *1 |
| Clavulanic acid | β-Lactamase?, Transporter | Streptomyces clavuligerus | [451,459,460,463,464,465] |
| Thienamycin | β-Lactamase, Transporter, Acetylation | Streptomyces cattleya | [467,468] |
| Nocardicin A | Transporter, Acetylation | Nocardia uniformis | [470], AY541063 *1 |
| Sulfazecin | β-Lactamase, Transporter | Pseudomonas acidophila | [472], KX757706 *1 |
| Viomycin | Phosphorylation | Streptomyces species | [473], AY263398 *1 |
| Pristinamycin | Transporter, Efflux pump | Streptomyces pristinaespiralis | [474,475] |
| Lincomycin | rRNA methylation | Streptomyces caelestis | [476,477] |
| Daunorubicin/Doxorubicin | Transporter, DNA-binding protein, Oxidoreductase? | Streptomyces peucetius | [478,479,480] |
| Chromomycin A3 | Transporter, DNA-binding protein | Streptomyces griseus | [481] |
| Mithramycin | Transporter | Streptomyces species | [483], X89899 *1 |
| Novobiocin | Transporter, Modification of target (gyrase) | Streptomyces spheroides | [484], AF170880 *1 |
| Coumermycin A1 | Transporter, Modification of target (gyrase, topoisomerase IV) | Streptomyces rishiriensis | [485] |
| Clorobiocin | Transporter, Modification of target (gyrase, topoisomerase IV) | Streptomyces species | [486] |
| Mitomycin | Oxidoreductases, Mitomycin-binding protein | Streptomyces lavendulae | [487,488] |
| Yatakemycin | Transporter, DNA glycosylase (DNA repair enzyme) | Streptomyces species | [489,490], JF429418 *1 |
| Natamycin | Transporter | Streptomyces chattanoogensis | [491] |
| Capuramycin | Phosphorylation | Amycolatopsis species | [492], KP995196 *1 |
| A-500359s | Phosphorylation | Streptomyces griseus | [493] |
| Laspartomycin | Transporters | Streptomyces viridochromogenes | [495] |
| Platensimycin/Platencin | Transporter, Target replacement/modification | Streptomyces platensis | [496] |
| d-Cycloserine | d-alanyl-d-alanine ligase, Membrane protein DcsJ | Streptomyces lavendulae | [497] |
| Fosfomycin | Phosphorylation | Streptomyces wedmorensis | [498] |
| Rifamycin | Transporter, Glycosylation, Phosphorylation, Target modification | Nocardia species | [499,500] |
| Thiopeptide antibiotics | Target modification (rRNA protein), rRNA methylation? | Streptomyces azureus | [501,502] |
| Neocarzinostatin | Modification of apo-protein, Sequestration, Mycothiol-dependent detoxication | Streptomyces carzinostaticus | [503] |
| Calicheamicin | Non-hem iron metalloprotein | Micromonospora echinospora | [504,505], AF497482 *1 |
| Kedarcidin | Apoprotein, Transporters | Streptoalloteichus species | [506,507] |
| Cyanosporaside | Transporters | Salinispora pacifica, Streptomyces species | [508] |
| Salinosporamide A | Mutation of target (proteasome) | Salinispora tropica | [509], EF397502 *1 |
| Maklamicin | Transporters | Micromonospora species | [510], LC021382 *1 |
| Microbisporicin | Transporters | Microbispora corallina | [511], HM536998 *1 |
| Surfactin | Transporters | Bacillus subtilis | [512,513] |
| Subtilin | Transporter | Bacillus subtilis | [514] |
| Sublancin | S-glycosylation | Bacillus subtilis | [515] |
| Zwittermicin | Transporters | Bacillus cereus, Bacillus thuringiensis | [516,517], HQ846969 *1 |
| Bacitracin | Transporters | Bacillus lichenifomis | [518] |
| Polymyxin | Transporters | Paenibacillus polymyxa | [519], EU371992 *1 |
| Edeine | Transporter, Acetylation | Brevibacillus brevis | [520], KC771276 *1 |
| Kalimantacin | ACP reductase, Transporter | Pseudomonas fluorescens, Alcaligenes sp. | [522,523], GU479979 *1 |
| Pseudomonic acid | tRNA synthetase | Pseudomonas fluorescens | [524] |
| Holomycin | rRNA methylation | Yersinia ruckeri | [525] |
| S-methylation, Transporter | Streptomyces clavuligerus | [527], DS570652 *1 | |
| Agrocin 84 | Mutation of target (tRNA synthetase) | Agrobacterium radiobacter | [528] |
| Griseofulvin | Transporter | Penicillium species | [529] |
| Mycophenolic acid | IMP dehydrogenase | Penicillium species | [530,531], HQ731031 *1 |
*1: GenBank accession number for the resistance-ralated gene.
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ogawara, H. Comparison of Strategies to Overcome Drug Resistance: Learning from Various Kingdoms. Molecules 2018, 23, 1476. https://doi.org/10.3390/molecules23061476
AMA Style
Ogawara H. Comparison of Strategies to Overcome Drug Resistance: Learning from Various Kingdoms. Molecules. 2018; 23(6):1476. https://doi.org/10.3390/molecules23061476
Chicago/Turabian StyleOgawara, Hiroshi. 2018. "Comparison of Strategies to Overcome Drug Resistance: Learning from Various Kingdoms" Molecules 23, no. 6: 1476. https://doi.org/10.3390/molecules23061476