Study on Hybrid Characteristics of Medicinally Used Cultivated Codonopsis Species Using Ribosomal Internal Transcribed Spacer (ITS) Sequencing

Abstract
:1. Introduction
2. Results and Discussion
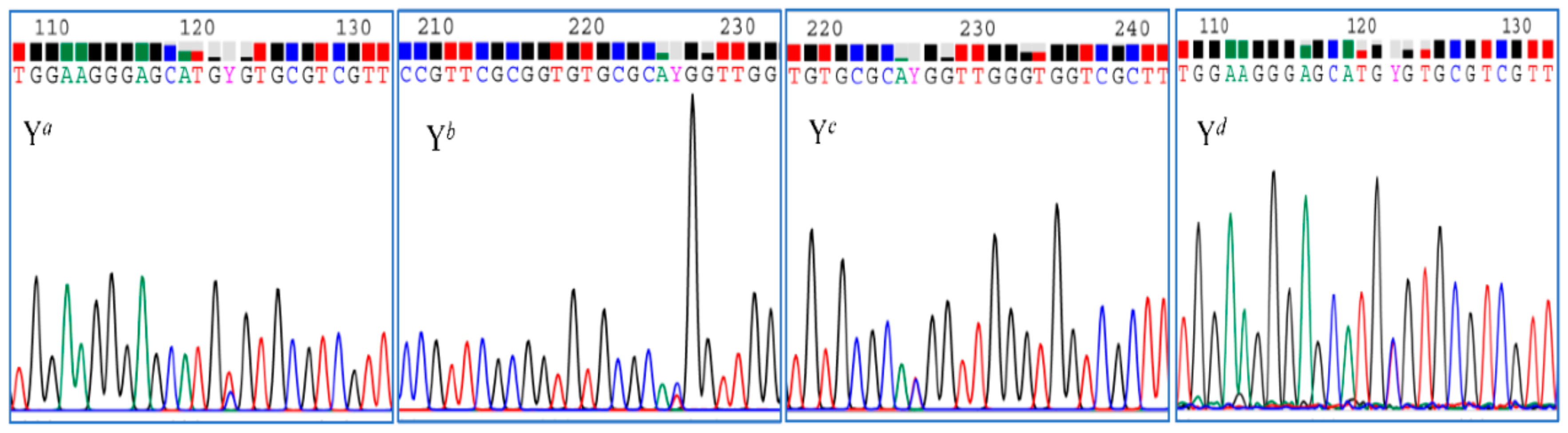
2.1. Verification of Additive Nucleotides
2.2. ITS Sequences of Codonopsis Plant Species
2.3. Molecular Phylogenetic Tree Based on the ITS Gene Sequences
2.4. Hybridization Analysis Based on the ITS Gene Sequences
3. Materials and Methods
3.1. Plant Materials
3.2. DNA Extraction
3.3. PCR Amplification
3.4. Verification on Variable Nucleotide
3.5. Processing on ITS Sequence
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Commission, C.P. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume I, pp. 281–282. ISBN 978-7-5067-7337-9. [Google Scholar]
- Wang, J.; Deng, C.-Q.; Shi, L.; Deng, M.-L. Progress of modern research on Codonopsis taxa. Guide China Med. 2011, 9, 279–281. [Google Scholar] [CrossRef]
- Liu, X.-X.; Chen, Y.-L.; Shi, Y.-T.; Luo, J.-J.; Hou, Y.-Q.; Zhang, Y.-P. Analysis on genetic diversity of germplasm resources of radix Codonopsis cultivated in gansu province based on ISSR marker. J. Chin. Med. Mater. 2016, 39, 1742–1747. [Google Scholar] [CrossRef]
- Yang, H.-Z.; Chen, Y.; Guo, F.-X.; Zhou, C.-M.; He, Y.-L. Resource investigation of wild Codonopsis pilosula inTanchang county of Gansu. China J. Chin. Mater. Med. 2016, 41, 186–191. [Google Scholar] [CrossRef]
- Zhao, S.; Xin, T.-Y.; Hou, D.-Y.; Pang, X.-H.; Chen, R.-Y.; Gao, J.-P. Identification of Codonopsis Radix and Its Adulterants Using the ITS/ITS2 Barcodes. World Sci. Technol./Mod. Tradit. Chin. Med. Mater. Med. 2013, 15, 421–428. [Google Scholar] [CrossRef]
- Liu, D.; Guo, H.-Y.; Tang, W.-Q.; Yang, J.-Q. Comparative Evolution of S7 Intron 1 and Ribosomal Internal Transcribed Spacer in Coilia nasus (Clupeiformes: Engraulidae). Int. J. Mol. Sci. 2012, 13, 3085–3100. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.-M.; Wang, Z.; Mu, X.-Q.; Liang, Z.-S.; Guo, H.-B. A Dominant Gene for Male Sterility in Salvia miltiorrhiza Bunge. PLoS ONE 2012, 7, e50903. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Nandagopal, K.; Jha, T.B. Molecular characterization of some Indian Aloe vera populations through RAPD and ITS markers. Plant Biosyst. 2017, 151, 695–703. [Google Scholar] [CrossRef]
- Han, E.-H.; Lee, S.-J.; Kim, M.-B.; Shin, Y.-W.; Kim, Y.-H.; Lee, S.-W. Molecular marker analysis of Cynanchum wilfordii and C-auriculatum using the simple ARMS-PCR method with mismatched primers. Plant Biotechnol. Rep. 2017, 11, 127–133. [Google Scholar] [CrossRef]
- Jia, Q.-J.; Wang, J.-M.; Zhu, J.-H.; Hua, W.; Shang, Y.; Yang, J.-M.; Liang, Z.-S. Toward Identification of Black Lemma and Pericarp Gene Blp1 in Barley Combining Bulked Segregant Analysis and Specific-Locus Amplified Fragment Sequencing. Front. Plant Sci. 2017, 8, 1414. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.-J.; Xia, P.-G.; He, Z.-G.; Liu, Y.; Liu, F.-H.; Zhao, H.-G.; Han, R.-L.; Liang, Z.-S. Identification of seeds based on molecular markers and secondary metabolites in Senna obtusifolia and Senna occidentalis. Bot. Stud. 2017, 58, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yan, S.; Li, J.-J.; Xiong, C.; Shi, Y.-H.; Wu, L.; Xiang, L.; Deng, B.; Ma, W.; Chen, S. Study of Commercially Available Lobelia chinensis Products Using Bar-HRM Technology. Front. Plant Sci. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-Y.; Chen, X.-C.; Yang, P.; Wang, L.-L.; Han, J.-P. Barcoding the Dendrobium (Orchidaceae) Species and Analysis of the Intragenomic Variation Based on the Internal Transcribed Spacer 2. Biomed. Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Jia, X.-R.; Liu, P.; Zhang, Q.-M.; Liu, F.; Ma, C.-D.; Xi, P.-Z.; Liang, Z.-S. Species identification of polygonati rhizoma in China by both morphological and molecular marker methods. Comptes Rendus Biol. 2018, 341, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.-J.; Xia, P.-G.; Liu, J.-L.; Li, X.; Han, R.-L.; Liu, F.-H.; Zhao, H.-G.; Liang, Z.-S. Genetic diversity and population structure assessment of Chinese Senna obtusifolia L. by molecular markers and morphological traits of seed. Acta Physiol. Plant. 2018, 40, 12. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hsieh, C.C.; Agrawal, D.C.; Kuo, C.-L.; Chue, F.-S.; Tsay, H.S. ITS sequence based phylogenetic relationship of Dangshen radix. J. Food Drug Anal. 2007, 15, 428–432. [Google Scholar] [CrossRef]
- Kitani, Y.; Zhu, S.; Batkhuu, J.; Sanchir, C.; Komatsu, K. Genetic diversity of Ephedra plants in mongolia inferred from internal transcribed spacer sequence of nuclear ribosomal DNA. Biol. Pharm. Bull. 2011, 34, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.-S.; Yuan, Q.-J.; Li, Q.-Q.; Huang, L.-Q. Molecular Systematics of Genus Atractylodes (Compositae, Cardueae): Evidence from Internal Transcribed Spacer (ITS) and trnL-F Sequences. Int. J. Mol. Sci. 2012, 13, 14623–14633. [Google Scholar] [CrossRef] [PubMed]
- He, J.-Y.; Zhu, S.; Komatsu, K.; Goda, Y.; Cai, S.-Q. Genetic polymorphism of medicinally-used Codonopsis species in an internal transcribed spacer sequence of nuclear ribosomal DNA and its application to authenticate Codonopsis Radix. J. Nat. Med. 2014, 68, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-C.; Zhao, M.-M.; Sun, W.; Teng, H.-L.; Huang, B.-S.; Zhao, X.-P. Differentiation of the Chinese minority medicinal plant genus Berchemia spp. by evaluating three candidate barcodes. Springerplus 2016, 5, 658. [Google Scholar] [CrossRef] [PubMed]
- Han, E.-H.; Cho, K.; Goo, Y.; Kim, M.; Shin, Y.-W.; Kim, Y.-H.; Lee, S.-W. Development of molecular markers, based on chloroplast and ribosomal DNA regions, to discriminate three popular medicinal plant species, Cynanchum wilfordii, Cynanchum auriculatum, and Polygonum multiflorum. Mol. Biol. Rep. 2016, 43, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.-G.; Guo, H.-B.; Zhang, Y.; Deyholos, M.K.; Peng, L.; Jia, Y.-Y.; Yan, X.-J.; Liu, Y.; Liang, Z.-S. Wild Panax vietnamensis and Panax stipuleanatus markedly increase the genetic diversity of Panax notoginseng (Araliaceae) revealed by start codon targeted (SCoT) markers and ITS DNA barcode. Biochem. Syst. Ecol. 2016, 66, 37–42. [Google Scholar] [CrossRef]
- Baldwin, B.G. Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: An example from the compositae. Mol. Phylogenetics Evol. 1992, 1, 3–16. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F. The ITS Region of Nuclear Ribosomal DNA: A Valuable Source of Evidence on Angiosperm Phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Denk, T.; Grimm, G.W. Phylogeny and biogeography of Zelkova (Ulmaceae sensu stricto) as inferred from leaf morphology, ITS sequence data and the fossil record. Bot. J. Linn. Soc. 2005, 147, 129–157. [Google Scholar] [CrossRef]
- Calonje, M.; Martín-Bravo, S.; Dobeš, C.; Koch, M.A. Non-coding nuclear DNA markers in phylogenetic reconstruction. Plant Syst. Evol. 2009, 282, 257–280. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Documentation of reticulate evolution in peonies (Paeonia) using internal transcribed spacer sequences of nuclear ribosomal DNA: Implications for biogeography and concerted evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 6813–6817. [Google Scholar] [CrossRef] [PubMed]
- Emshwiller, E.; Doyle, J. Origins of domestication and polyploidy in oca (Oxalis tuberosa: Oxalidaceae): NrDNA ITS data. Am. J. Bot. 1998, 85, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Franzke, A.; Mummenhoff, K. Recent hybrid speciation in Cardamine(Brassicaceae)+conversion of nuclear ribosomal ITS sequences in statu nascendi. Theor. Appl. Genet. 1999, 98, 831–834. [Google Scholar] [CrossRef]
- Widmer, A.; Baltisberger, M. Molecular evidence for allopolyploid speciation and a single origin of the narrow endemic Draba ladina (Brassicaceae). Am. J. Bot. 1999, 86, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Whittall, J.; Liston, A.; Gisler, S.; Meinke, R. Detecting nucelotide additivity from direct sequences is a SNAP: An example from Sidalcea (Malvaceae). Plant Biol. 2000, 2, 211–217. [Google Scholar] [CrossRef]
- Jason, T.; Rauscher, J.; Doyle, J.J.; Brown, A.H.D. Internal transcribed spacer repeat-specific primers and the analysis of hybridization in the Glycine tomentella (Leguminosae) polyploid complex. Mol. Ecol. 2002, 11, 2691–2702. [Google Scholar] [CrossRef]
- Feng, T.; Zhou, Z.; Tang, J.; Cheng, M.; Zhou, S. ITS sequence variation supports the hybrid origin of Malus toringoides hughes. Can. J. Bot 2007, 85, 659–666. [Google Scholar] [CrossRef]
- Liao, L.; Xu, L.-L.; Zhang, D.-M.; Fang, L.; Deng, H.-S.; Shi, J.-W.; Li, T.-J. Multiple hybridization origin of Ranunculus cantoniensis (4×): Evidence from trnL-F and ITS sequences and fluorescent in situ hybridization (FISH). Plant Syst. Evol. 2008, 276, 31–37. [Google Scholar] [CrossRef]
- Soltis, D.E.; Mavrodiev, E.V.; Doyle, J.J.; Rauscher, J.; Soltis, P.S. ITS and ETS sequence data and phylogeny reconstruction in allopolyploids and hybrids. Syst. Bot. 2008, 33, 7–20. [Google Scholar] [CrossRef]
- Consaul, L.L.; Gillespie, L.J.; Waterway, M.J. Evolution and polyploid origins in north American arctic puccinellia (poaceae) based on nuclear ribosomal spacer and chloroplast DNA sequences. Am. J. Bot. 2010, 97, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.A.; Karl, R.; German, D.A.; Al-Shehbaz, I.A. Systematics, taxonomy and biogeography of three new Asian genera of Brassicaceae tribe Arabideae: An ancient distribution circle around the Asian high mountains. Taxon 2012, 61, 955–969. [Google Scholar]
- Punina, E.O.; Machs, E.M.; Krapivskaya, E.E.; Kim, E.-S.; Mordak, E.V.; Myakoshina, Y.A.; Rodionov, A.V. Interspecific hybridization in the genus Paeonia (Paeoniaceae): Polymorphic sites in transcribed spacers of the 45S rRNA genes as indicators of natural and artificial peony hybrids. Russ. J. Genet. 2012, 48, 684–697. [Google Scholar] [CrossRef]
- Mandakova, T.; Kovarik, A.; Zozomova-Lihova, J.; Shimizu-Inatsugi, R.; Shimizu, K.K.; Mummenhoff, K.; Marhold, K.; Lysak, M.A. The More the Merrier: Recent Hybridization and Polyploidy in Cardamine. Plant Cell 2013, 25, 3280–3295. [Google Scholar] [CrossRef] [PubMed]
- Hodac, L.; Scheben, A.P.; Hojsgaard, D.; Paun, O.; Horandl, E. ITS Polymorphisms Shed Light on Hybrid Evolution in Apomictic Plants: A Case Study on the Ranunculus auricomus Complex. PLoS ONE 2014, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zozomova Lihova, J.; Krak, K.; Mandakova, T.; Shimizu, K.K.; Spaniel, S.; Vit, P.; Lysak, M.A. Multiple hybridization events in Cardamine (Brassicaceae) during the last 150 years: Revisiting a textbook example of neoallopolyploidy. Ann. Bot. 2014, 113, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.-B.; Zhang, J.; Hu, Z.-H.; Yuan, D.-P. Molecular identification of wild and cultivated banqiao Codonopsis species. Lishizhen Med. Mater. Res. 2010, 21, 3272–3273. [Google Scholar] [CrossRef]
- Li, D.-Z.; Gao, L.-M.; Li, H.-T.; Wang, H.; Ge, X.-J.; Liu, J.-Q.; Chen, Z.-D.; Zhou, S.-L.; Chen, S.-L.; Yang, J.-B. Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporated into the core barcode for seed plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19641–19646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkins, K.M. Chapter 15—Analysis of Deoxyribonucleic Acid (DNA) Sequence Data Using BioEdit. In Forensic DNA Biology; Academic Press: San Diego, CA, USA, 2013; pp. 129–132. [Google Scholar]
- Lu, G.-Q.; Moriyama, E.N. Vector NTI, a balanced all-in-one sequence analysis suite. Brief. Bioinform. 2004, 5, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the Codonopsis taxa are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| pCT46 (%) a | Main Signal | S-Value (%) | N-Value (%) | Significance b |
|---|---|---|---|---|
| 10 | C | 11.54 ± 1.46 | 10.82 ± 0.98 | |
| 15 | C | 14.82 ± 1.16 | 10.64 ± 1.13 | ** |
| 20 | C | 21.08 ± 2.23 | 10.66 ± 1.07 | ** |
| 30 | C | 29.14 ± 1.65 | 8.92 ± 1.18 | ** |
| 40 | C | 39.60 ± 1.87 | 9.02 ± 1.19 | ** |
| 50 | C | 47.72 ± 1.37 | 8.84 ± 1.16 | ** |
| Species | ITS Type | G + C Content (%) | Nucleotide Position | Sequence Type of Supposed Parental Lineages (Nucleotides at 122nd and 226th) | Number of Specimens | GenBank Accession No. | |||
|---|---|---|---|---|---|---|---|---|---|
| 122 | 135 | 226 | 500 | ||||||
| C. pilosula | P0 | 60.61 | C | G | C | G | EF190460 | ||
| P1 | 60.31 | Y a | * | Y b | * | P0 (C C) × S0 (T T) | 33 | ||
| P2 | 60.46 | * | * | Y b | * | 1 | |||
| P3 | 60.31 | Y a | * | Y c | * | P0 (C C) × S0 (T T) | 1 | ||
| P4 | 60.31 | Y d | * | Y b | * | P0 (C C) × S0 (T T) | 2 | ||
| P5 | 60.46 | Y d | * | * | * | P0 (C C) × PM0 (T C) | 2 | ||
| C. pilosula var. modesta | PM0 | 60.46 | T | * | * | * | EF190461 | ||
| PM1 | 60.31 | Y a | * | Y b | * | P0 (C C) × S0 (T T) | 4 | ||
| PM2 | 60.31 | T | * | Y b | * | S0 (T T) × PM0 (T C) | 1 | ||
| PM3 | 60.46 | Y a | * | * | * | P0 (C C) × PM0 (T C) | 1 | ||
| C. tangshen | T0 | 60.31 | T | * | * | A | EF190462 | ||
| T1 | 60.15 | T | R | * | R | 1 | |||
| C.SP. | S0 | 60.31 | T | * | T | * | AB769272 | ||
| Voucher No. | Species | Sequence Type (ITS) | Hybrid Ratio (%, 122nd) | Hybrid Ratio (%, 226th) |
|---|---|---|---|---|
| CP01 | C. pilosula | P1 | 33.46 ± 1.29 | 46.89 ± 1.35 |
| CP02 | C. pilosula | P2 | 33.49 ± 1.55 | |
| CP03 | C. pilosula | P1 | 37.47 ± 1.47 | 37.07 ± 0.50 |
| CP04 | C. pilosula | P1 | 36.14 ± 1.73 | 39.39 ± 1.17 |
| CP05 | C. pilosula | P1 | 48.33 ± 1.56 | 25.14 ± 2.52 |
| CP06 | C. pilosula | P1 | 35.23 ± 1.31 | 39.18 ± 1.52 |
| CP07 | C. pilosula | P1 | 33.25 ± 0.70 | 41.59 ± 0.45 |
| CP08 | C. pilosula | P1 | 44.77 ± 1.09 | 39.70 ± 1.62 |
| CP09 | C. pilosula | P1 | 31.9 ± 1.17 | 33.57 ± 2.13 |
| CP10 | C. pilosula | P3 | 32.72 ± 1.48 | 47.79 ± 1.77 |
| CP11 | C. pilosula | P1 | 40.86 ± 1.71 | 29.05 ± 2.04 |
| CP12 | C. pilosula | P1 | 38.34 ± 1.26 | 39.92 ± 2.39 |
| CP13 | C. pilosula | P1 | 35.94 ± 0.98 | 40.33 ± 3.32 |
| CP14 | C. pilosula | P1 | 44.52 ± 0.72 | 38.14 ± 1.68 |
| CP15 | C. pilosula | P1 | 35.05 ± 1.40 | 43.61 ± 2.30 |
| CP16 | C. pilosula | P1 | 30.65 ± 0.99 | 32.98 ± 2.03 |
| CP17 | C. pilosula | P1 | 32.57 ± 0.80 | 28.64 ± 0.82 |
| CP18 | C. pilosula | P1 | 45.63 ± 0.93 | 32.35 ± 2.46 |
| CP19 | C. pilosula | P1 | 39.53 ± 3.48 | 40.20 ± 2.11 |
| CP20 | C. pilosula | P1 | 32.24 ± 0.63 | 37.79 ± 1.06 |
| CP21 | C. pilosula | P1 | 40.25 ± 0.21 | 40.62 ± 1.27 |
| CP22 | C. pilosula | P1 | 35.93 ± 0.35 | 37.99 ± 1.30 |
| CP23 | C. pilosula | P1 | 39.33 ± 0.94 | 33.61 ± 1.03 |
| CP24 | C. pilosula | P1 | 36.48 ± 0.57 | 34.09 ± 1.12 |
| CP25 | C. pilosula | P1 | 39.65 ± 0.17 | 34.66 ± 0.83 |
| CP26 | C. pilosula | P4 | 47.64 ± 1.72 | 32.05 ± 2.11 |
| CP27 | C. pilosula | P1 | 34.86 ± 1.51 | 22.40 ± 1.51 |
| CP28 | C. pilosula | P1 | 43.02 ± 0.83 | 31.94 ± 0.82 |
| CP29 | C. pilosula | P1 | 40.71 ± 1.63 | 25.37 ± 1.88 |
| CP30 | C. pilosula | P1 | 48.02 ± 0.41 | 22.07 ± 1.46 |
| CP31 | C. pilosula | P1 | 43.49 ± 0.46 | 20.11 ± 1.74 |
| CP32 | C. pilosula | P1 | 47.36 ± 0.41 | 17.41 ± 1.73 |
| CP33 | C. pilosula | P1 | 28.92 ± 0.49 | 29.00 ± 5.90 |
| CP34 | C. pilosula | P5 | 34.88 ± 5.96 | |
| CP35 | C. pilosula | P1 | 46.22 ± 6.07 | 28.87 ± 1.41 |
| CP36 | C. pilosula | P1 | 43.10 ± 4.92 | 29.25 ± 2.70 |
| CP37 | C. pilosula | P5 | 39.67 ± 6.60 | |
| CP38 | C. pilosula | P4 | 24.14 ± 0.85 | 16.47 ± 0.66 |
| CP39 | C. pilosula | P1 | 34.67 ± 1.01 | 42.17 ± 0.71 |
| CPM40 | C. pilosula var. modesta | PM1 | 36.99 ± 4.01 | 38.67 ± 0.54 |
| CPM41 | C. pilosula var. modesta | PM1 | 39.29 ± 0.25 | 33.92 ± 1.33 |
| CPM42 | C. pilosula var. modesta | PM1 | 36.88 ± 0.49 | 21.23 ± 0.66 |
| CPM43 | C. pilosula var. modesta | PM1 | 22.94 ± 0.80 | 19.67 ± 0.95 |
| CPM44 | C. pilosula var. modesta | PM2 | 25.56 ± 1.40 | |
| CPM45 | C. pilosula var. modesta | PM3 | 27.35 ± 1.01 |
| Voucher No. | Species | Locality | Locality No. a | Altitude (m) | Date of Collection | Sequence Type (ITS) b |
|---|---|---|---|---|---|---|
| CP01 | C. pilosula | Liancai, Longde, Guyuan, Ningxia, China | 1 | 1760 | 20 July 2016 | P1 |
| CP02 | C. pilosula | Kezhai, Longxi, Dingxi, Gansu, China | 2 | 2220 | 25 October 2016 | P2 |
| CP03 | C. pilosula | Xiaozhai, Minxian, Dingxi, Gansu, China | 3 | 2550 | 25 October 2016 | P1 |
| CP04 | C. pilosula | Weixin, Minxian, Dingxi, Gansu, China | 4 | 2225 | 25 October 2016 | P1 |
| CP05 | C. pilosula | Xijiang, Minxian, Dingxi, Gansu, China | 5 | 2254 | 25 October 2016 | P1 |
| CP06 | C. pilosula | Meichuan, Minxian, Dingxi, Gansu, China | 6 | 2328 | 25 October 2016 | P1 |
| CP07 | C. pilosula | Minyang, Minxian, Dingxi, Gansu, China | 7 | 2305 | 25 October 2016 | P1 |
| CP27 | C. pilosula | Minyang, Minxian, Dingxi, Gansu, China | 2305 | 14 October 2017 | P1 | |
| CP08 | C. pilosula | Chabu, Minxian, Dingxi, Gansu, China | 8 | 2313 | 25 October 2016 | P1 |
| CP28 | C. pilosula | Mazichuan, Minxian, Dingxi, Gansu, China | 9 | 2510 | 14 October 2017 | P1 |
| CP09 | C. pilosula | Zhongzhai, Minxian, Dingxi, Gansu, China | 10 | 2381 | 25 October 2016 | P1 |
| CP10 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 11 | 2281 | 28 October 2016 | P3 |
| CP11 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2250 | 28 October 2016 | P1 | |
| CP12 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2238 | 28 October 2016 | P1 | |
| CP13 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2233 | 28 October 2016 | P1 | |
| CP14 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2271 | 28 October 2016 | P1 | |
| CP34 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2445 | 14 October 2017 | P4 | |
| CP35 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2435 | 14 October 2017 | P1 | |
| CP36 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2188 | 14 October 2017 | P1 | |
| CP37 | C. pilosula | Hadapu, Tanchang, Longnan, Gansu, China | 2242 | 14 October 2017 | P3 | |
| CP15 | C. pilosula | Awu, Tanchang, Longnan, Gansu, China | 12 | 2421 | 28 October 2016 | P1 |
| CP32 | C. pilosula | Awu, Tanchang, Longnan, Gansu, China | 2351 | 14 October 2017 | P1 | |
| CP33 | C. pilosula | Awu, Tanchang, Longnan, Gansu, China | 2329 | 14 October 2017 | P1 | |
| CP16 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 13 | 2503 | 28 October 2016 | P1 |
| CP17 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2368 | 28 October 2016 | P1 | |
| CP18 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2390 | 28 October 2016 | P1 | |
| CP30 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2431 | 14 October 2017 | P1 | |
| CP31 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2456 | 14 October 2017 | P1 | |
| CP38 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2320 | 14 October 2017 | P5 | |
| CP39 | C. pilosula | Pangjia, Tanchang, Longnan, Gansu, China | 2306 | 14 October 2017 | P1 | |
| CP19 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 14 | 2255 | 28 October 2016 | P1 |
| CP29 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2255 | 14 October 2017 | P1 | |
| CP20 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2286 | 28 October 2016 | P1 | |
| CP21 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2388 | 28 October 2016 | P1 | |
| CP22 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2475 | 28 October 2016 | P1 | |
| CP23 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2314 | 28 October 2016 | P1 | |
| CP24 | C. pilosula | Lichuan, Tanchang, Longnan, Gansu, China | 2320 | 28 October 2016 | P1 | |
| CP25 | C. pilosula | Guanzhuang, Yaozhou, Tongchuan, Shaanxi, China | 15 | 880 | 15 November 2016 | P1 |
| CP26 | C. pilosula | Hongtiguan, Pingshun, Changzhi, Shanxi, China | 16 | 1245 | 12 March 2017 | P4 |
| CPM40 | C. pilosula var. modesta | Danbao, Wenxian, Longnan, Gansu, China | 17 | 895 | 15 October 2016 | PM1 |
| CPM41 | C. pilosula var. modesta | Koutouba, Wenxian, Longnan, Gansu, China | 18 | 1266 | 15 October 2016 | PM1 |
| CPM42 | C. pilosula var. modesta | Shifang, Wenxian, Longnan, Gansu, China | 19 | 995 | 28 October 2016 | PM2 |
| CPM43 | C. pilosula var. modesta | Baoziba, Wenxian, Longnan, Gansu, China | 20 | 1634 | 28 October 2016 | PM2 |
| CPM44 | C. pilosula var. modesta | Baoziba, Wenxian, Longnan, Gansu, China | 1480 | 28 October 2016 | PM3 | |
| CPM45 | C. pilosula var. modesta | Zhongzhai, Wenxian, Longnan, Gansu, China | 21 | 1361 | 28 October 2016 | PM2 |
| CT46 | C. tangshen | Banqiao, Enshi, Enshi, Hubei, China | 22 | 1775 | 18 August 2016 | T1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, L.-J.; Wang, E.-H.; Yang, Y.-C.; Xing, B.-C.; Ji, W.; Liu, F.; Liang, Z.-S. Study on Hybrid Characteristics of Medicinally Used Cultivated Codonopsis Species Using Ribosomal Internal Transcribed Spacer (ITS) Sequencing. Molecules 2018, 23, 1565. https://doi.org/10.3390/molecules23071565
Liang L-J, Wang E-H, Yang Y-C, Xing B-C, Ji W, Liu F, Liang Z-S. Study on Hybrid Characteristics of Medicinally Used Cultivated Codonopsis Species Using Ribosomal Internal Transcribed Spacer (ITS) Sequencing. Molecules. 2018; 23(7):1565. https://doi.org/10.3390/molecules23071565
Chicago/Turabian StyleLiang, Li-Jun, Er-Huan Wang, Yi-Chen Yang, Bing-Cong Xing, Wei Ji, Feng Liu, and Zong-Suo Liang. 2018. "Study on Hybrid Characteristics of Medicinally Used Cultivated Codonopsis Species Using Ribosomal Internal Transcribed Spacer (ITS) Sequencing" Molecules 23, no. 7: 1565. https://doi.org/10.3390/molecules23071565