Relationships between Structures of Condensed Tannins from Texas Legumes and Methane Production During In Vitro Rumen Digestion

 , ,
, ,
Abstract
:1. Introduction
2. Results
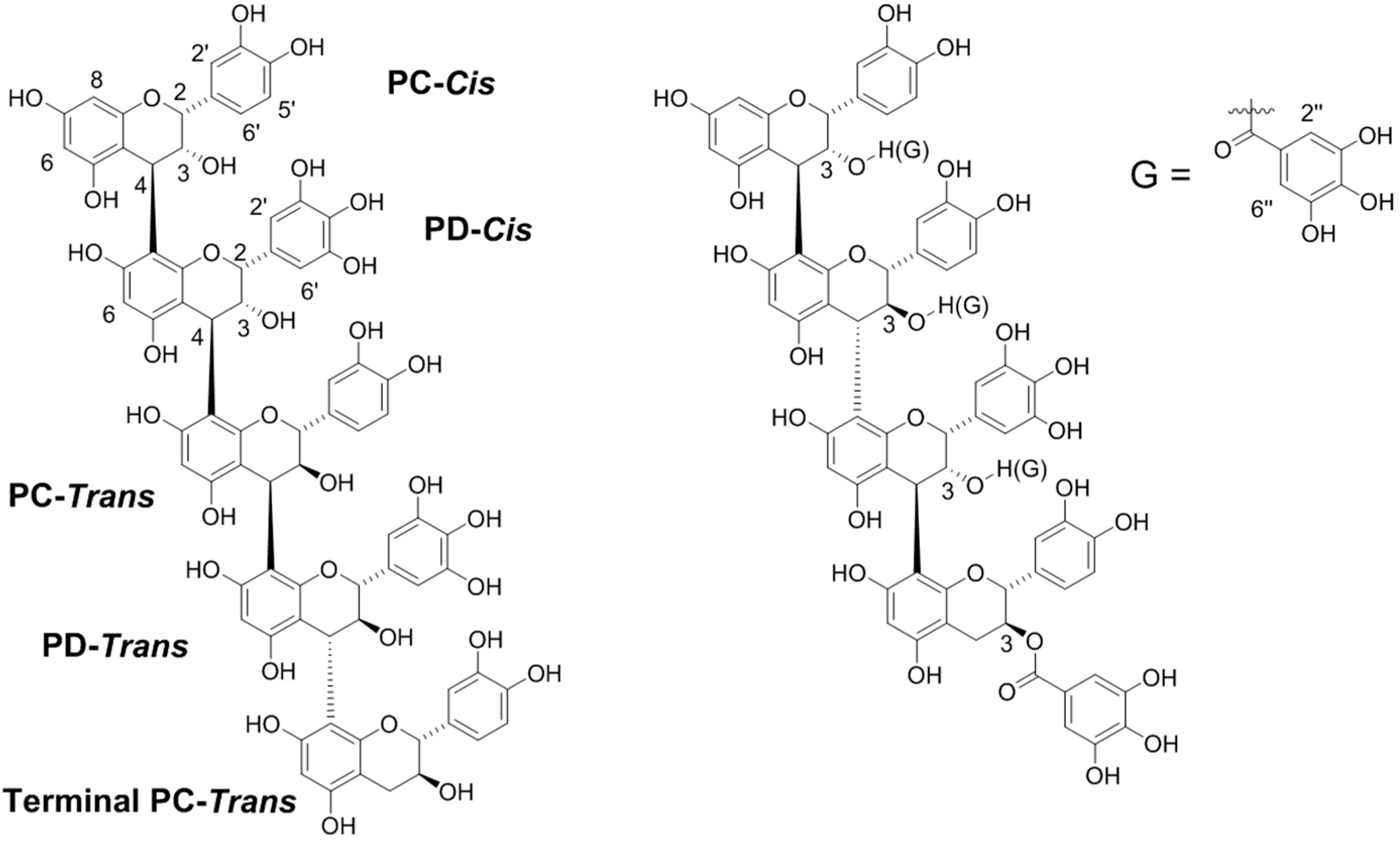
2.1. Degradative and NMR Analysis of Purified CTs
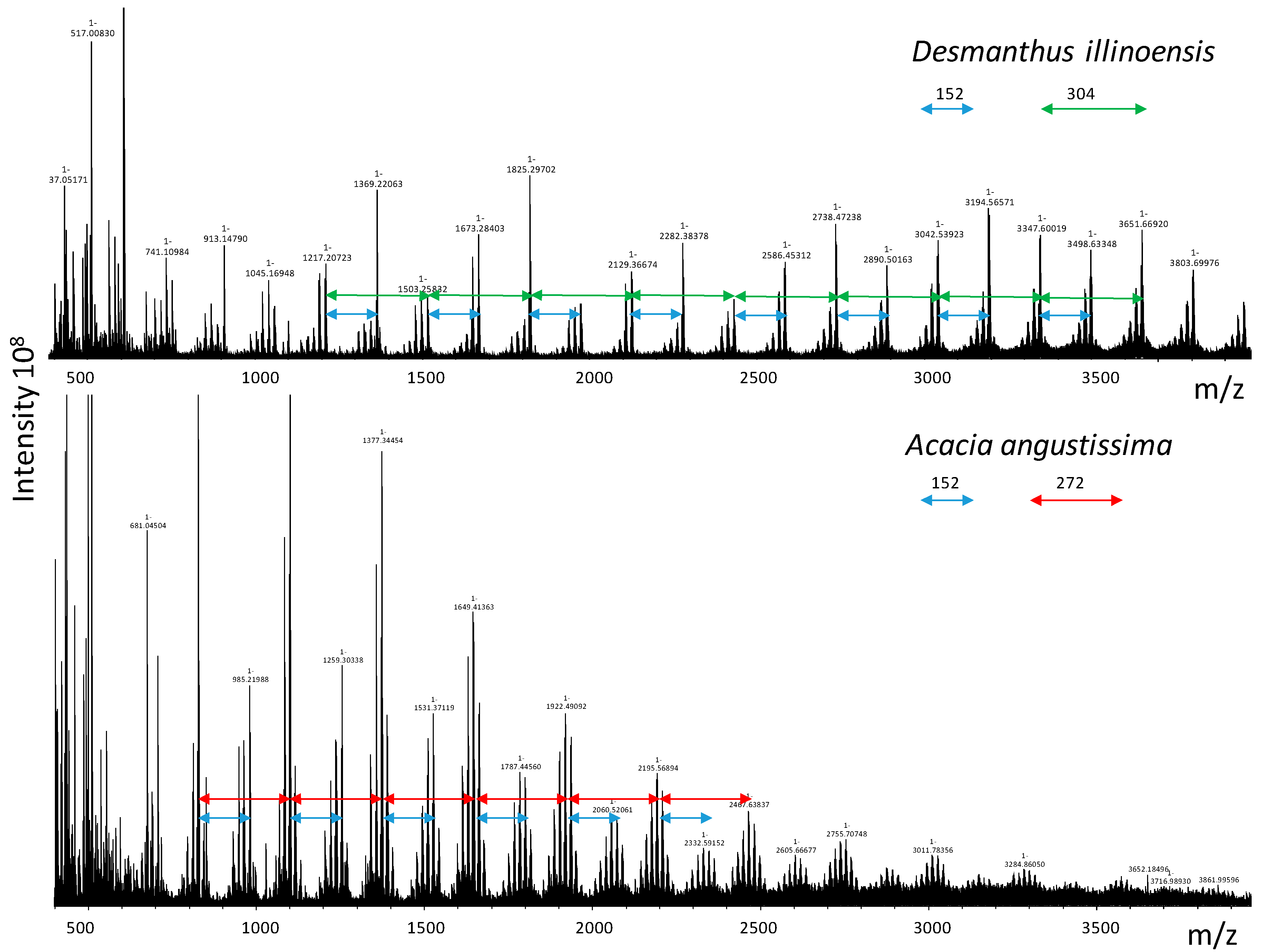
2.2. MALDI-TOF Mass Spectrometric Analysis
2.3. Antioxidant Activity
2.4. Protein Precipitation Activity
3. Discussion
3.1. Methods to Determine CT Structure
3.2. Chemotaxonomy
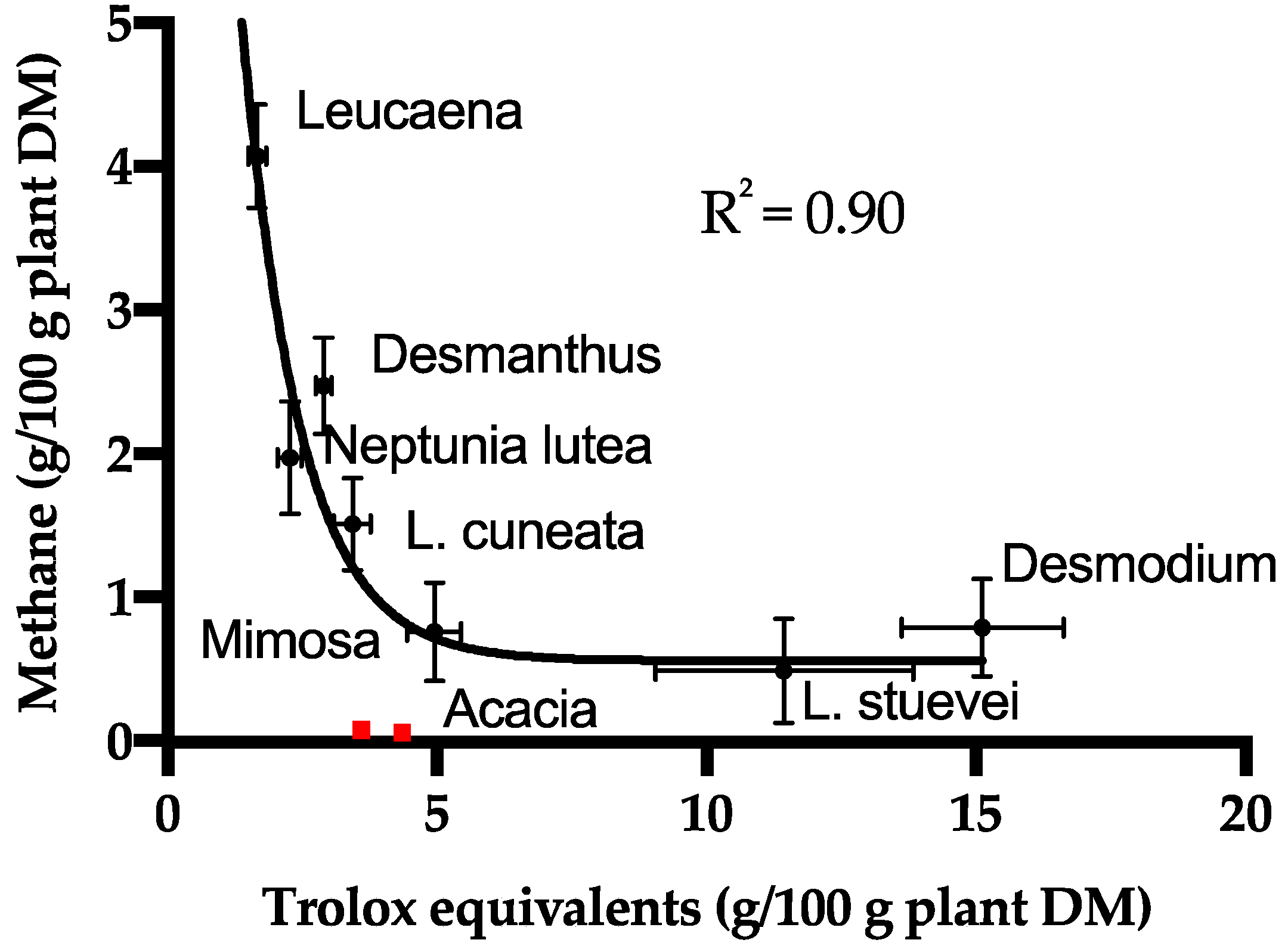
3.3. Methane Abatement
4. Materials and Methods
4.1. Chemicals
4.2. Tannin Purification
4.3. Anthocyanidins
4.4. Thiolysis
4.5. NMR Spectroscopy
4.6. MALDI-TOF Mass Spectrometry
4.7. Antioxidant Activity
4.8. Protein Precipitation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V. Phenolic compounds: From plants to foods. Phytochem. Rev. 2012, 11, 153–177. [Google Scholar] [CrossRef]
- Hanley, T.A.; Robbins, C.T.; Hagerman, A.E.; McArthur, C. Predicting digestible protein and digestible dry matter in tannin containing forages consumed by ruminants. Ecology 1992, 73, 537–541. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Doran-Browne, N.; Behrendt, R.; Kingwell, R.; Eckard, R. Modelling the potential of birdsfoot trefoil (Lotus corniculatus) to reduce methane emissions and increase production on wool and prime lamb farm enterprises. Anim. Prod. Sci. 2015, 55, 1097–1105. [Google Scholar] [CrossRef]
- Naumann, H.D.; Tedeschi, L.O.; Zeller, W.E.; Huntley, N.F. The role of condensed tannins in ruminant animal production: Advances, limitations and future directions. Rev. Bras. Zootec. 2017, 46, 929–949. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Jayanegara, A.; Leiber, F.; Kreuzer, M. Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments. J. Anim. Physiol. Anim. Nutr. 2012, 96, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Choudhury, P.K.; Dolores Carro, M.; Griffith, G.W.; Dagar, S.S.; Puniya, M.; Calabro, S.; Ravella, S.R.; Dhewa, T.; Upadhyay, R.C.; et al. New aspects and strategies for methane mitigation from ruminants. Appl. Microbiol. Biotechnol. 2014, 98, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, R.; Baruah, L.; Saravanan, M.; Suresh, K.P.; Sampath, K.T. Effect of medicinal and aromatic plants on rumen fermentation, protozoa population and methanogenesis in vitro. J. Anim. Physiol. Anim. Nutr. 2013, 97, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, T.T.; Lascano, C.E.; Wettstein, H.R.; Mayer, A.C.; Kreuzer, M.; Hess, H.D. Effect of the tropical tannin-rich shrub legumes Calliandra calothyrsus and Flemingia macrophylla on methane emission and nitrogen and energy balance in growing lambs. Animal 2008, 2, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Restrepo, C.A.; Barry, T.N. Alternative temperate forages containing secondary compounds for improving sustainable productivity in grazing ruminants. Anim. Feed Sci. Technol. 2005, 120, 179–201. [Google Scholar] [CrossRef]
- Bhatta, R.; Enishi, O.; Yabumoto, Y.; Nonaka, I.; Takusari, N.; Higuchi, K.; Tajima, K.; Takenaka, A.; Kurihara, M. Methane reduction and energy partitioning in goats fed two concentrations of tannin from Mimosa spp. J. Agric. Sci. 2013, 151, 119–128. [Google Scholar] [CrossRef]
- Moreira, G.D.; Lima, P.D.T.; Borges, B.O.; Primavesi, O.; Longo, C.; McManus, C.; Abdalla, A.; Louvandini, H. Tropical tanniniferous legumes used as an option to mitigate sheep enteric methane emission. Trop. Anim. Health Prod. 2013, 45, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Pellikaan, W.F.; Stringano, E.; Leenaars, J.; Bongers, D.J.G.M.; Van Laar-van Schuppen, S.; Plant, J.; Mueller-Harvey, I. Evaluating effects of tannins on extent and rate of in vitro gas and CH4 production using an automated pressure evaluation system (APES). Anim. Feed Sci. Technol. 2011, 166–167, 377–390. [Google Scholar] [CrossRef]
- Hassanat, F.; Benchaar, C. Assessment of the effect of condensed (acacia and quebracho) and hydrolysable (chestnut and valonea) tannins on rumen fermentation and methane production in vitro. J. Sci. Food Agric. 2013, 93, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Grainger, C.; Clarke, T.; Auldist, M.J.; Beauchemin, K.A.; McGinn, S.M.; Waghorn, G.C.; Eckard, R.J. Potential use of Acacia mearnsii condensed tannins to reduce methane emissions and nitrogen excretion from grazing dairy cows. Can. J. Anim. Sci. 2009, 89, 241–251. [Google Scholar] [CrossRef]
- White, R.G.; Lawler, J.P. Can methane suppression during digestion of woody and leafy browse compensate for energy costs of detoxification of plant secondary compounds? A test with muskoxen fed willows and birch. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 849–859. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.; Ramirez-Restrepo, C.A.; Muir, J.P. Developing a conceptual model of possible benefits of condensed tannins for ruminant production. Animal 2014, 8, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumann, H.D.; Tedeschi, L.O.; Muir, J.P.; Lambert, B.D.; Kothmann, M.M. Effect of molecular weight of condensed tannins from warm-season perennial legumes on ruminal methane production in vitro. Biochem. Syst. Ecol. 2013, 50, 154–162. [Google Scholar] [CrossRef]
- Hatew, B.; Carbonero, C.H.; Stringano, E.; Sales, L.F.; Smith, L.M.J.; Mueller-Harvey, I.; Hendriks, W.H.; Pellikaan, W.F. Diversity of condensed tannin structures affects rumen in vitro methane production in sainfoin (Onobrychis viciifolia) accessions. Grass Forage Sci. 2015, 70, 474–490. [Google Scholar] [CrossRef]
- Hatew, B.; Stringano, E.; Mueller-Harvey, I.; Hendriks, W.H.; Carbonero, C.H.; Smith, L.M.J.; Pellikaan, W.F. Impact of variation in structure of condensed tannins from sainfoin (Onobrychis viciifolia) on in vitro ruminal methane production and fermentation characteristics. J. Anim. Physiol. Anim. Nutr. 2016, 100, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Huyen, N.T.; Fryganas, C.; Uittenbogaard, G.; Mueller-Harvey, I.; Verstegen, M.W.A.; Hendriks, W.H.; Pellikaan, W.F. Structural features of condensed tannins affect in vitro ruminal methane production and fermentation characteristics. J. Agric. Sci. 2016, 154, 1474–1487. [Google Scholar] [CrossRef]
- Saminathan, M.; Sieo, C.C.; Abdullah, N.; Wong, C.; Ho, Y.W. Effects of condensed tannin fractions of different molecular weights from a Leucaena leucocephala hybrid on in vitro methane production and rumen fermentation. J. Sci. Food Agric. 2015, 95, 2742–2749. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E. Fifty years of polyphenol-protein complexes. Rec. Adv. Polyphen. Res. 2012, 3, 71–97. [Google Scholar]
- Hagerman, A.E.; Riedl, K.M.; Rice, R.E. Tannins as biological antioxidants. In Plant Polyphenols 2: Chemistry, Biology, Pharmacology, Ecology; Gross, G.G., Hemingway, R.W., Yoshida, T., Eds.; Kluwer Academic/Plenum: New York, NY, USA, 1999; pp. 495–505. [Google Scholar]
- Huang, X.D.; Liang, J.B.; Tan, H.Y.; Yahya, R.; Khamseekhiew, B.; Ho, Y.W. Molecular weight and protein binding affinity of Leucaena condensed tannins and their effects on in vitro fermentation parameters. Anim. Feed Sci. Technol. 2010, 159, 81–87. [Google Scholar] [CrossRef]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Ishimoto, H.; Tai, A.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Hatano, T.; Ito, H. Antioxidative properties of functional polyphenols and their metabolites assessed by an ORAC assay. Biosci. Biotech. Biochem. 2012, 76, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Joslyn, M.A.; Goldstein, J.L. Conversion of leucoanthocyanins into corresponding anthocyanidins. Science 1964, 143, 954–955. [Google Scholar] [CrossRef] [PubMed]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutounet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Zeller, W.E.; Ramsay, A.; Ropiak, H.M.; Fryganas, C.; Mueller-Harvey, I.; Brown, R.H.; Drake, C.; Grabber, J.H. H-1-C-13 HSQC NMR spectroscopy for estimating procyanidin/prodelphinidin and cis/trans-flavan-3-ol ratios of condensed tannin samples: Correlation with thiolysis. J. Agric. Food Chem. 2015, 63, 1967–1973. [Google Scholar] [CrossRef] [PubMed]
- Roux, D.G.; Evelyn, S.R. Condensed tannins: A study of complex leuco-anthocyanins present in condensed tannins. Biochem. J. 1958, 69, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Steynberg, P.J.; Bezuidenhoudt, B.C.B.; Ferreira, D.; Steynberg, J.P. Cleavage of the interflavanyl bond in 5-deoxy (A-ring) proanthocyanidins. J. Chem. Soc. Chem. Comm. 1994, 31–32. [Google Scholar] [CrossRef]
- Steynberg, P.J.; Steynberg, J.P.; Bezuidenhoudt, B.C.B.; Ferreira, D. Oligomeric flavonoids. Part 19. Reductive cleavage of the interflavanyl bond in proanthocyanidins. J. Chem. Soc. Perk. Trans. 1995, 1, 3005–3012. [Google Scholar] [CrossRef]
- Li, C.; Leverence, R.; Trombley, J.D.; Xu, S.; Yang, J.; Tian, Y.; Reed, J.D.; Hagerman, A.E. High molecular weight persimmon (Diospyros kaki L.) proanthocyanidin: A highly galloylated, A-linked tannin with an unusual flavonol terminal unit, myricetin. J. Agric. Food Chem. 2010, 58, 9033–9042. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Quintanilla-Lopez, J.E.; Gomez-Cordoves, C.; Bartolome, B.; Lebron-Aguilar, R. MALDI-TOF MS analysis of plant proanthocyanidins. J. Pharm. Biomed. Anal. 2010, 51, 358–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, C.G.; Vestling, M.M.; Reed, J.D. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of heteropolyflavan-3-ols and glucosylated heteropolyflavans in sorghum [Sorghum bicolor (L.) Moench]. J. Agric. Food Chem. 2003, 51, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Stringano, E.; Cramer, R.; Hayes, W.; Smith, C.; Gibson, T.; Mueller-Harvey, I. Deciphering the complexity of sainfoin (Onobrychis viciifolia) proanthocyanidins by MALDI-TOF mass spectrometry with a judicious choke of isotope patterns and matrixes. Anal. Chem. 2011, 83, 4147–4153. [Google Scholar] [CrossRef] [PubMed]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Food. 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef] [PubMed]
- Naumann, H.D.; Hagerman, A.E.; Lambert, B.D.; Muir, J.P.; Tedeschi, L.O.; Kothmann, M.M. Molecular weight and protein-precipitating ability of condensed tannins from warm-season perennial legumes. J. Plant. Interact. 2014, 9, 212–219. [Google Scholar] [CrossRef]
- Zeller, W.E.; Sullivan, M.L.; Mueller-Harvey, I.; Grabber, J.H.; Ramsay, A.; Drake, C.; Brown, R.H. Protein precipitation behavior of condensed tannins from Lotus pedunculatus and Trifolium repens with different mean degrees of polymerization. J. Agric. Food Chem. 2015, 63, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Scioneaux, A.N.; Schmidt, M.A.; Moore, M.A.; Lindroth, R.L.; Wooley, S.C.; Hagerman, A.E. Qualitative variation in proanthocyanidin composition of Populus species and hybrids: Genetics is the key. J. Chem. Ecol. 2011, 37, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Kelm, M.A.; Hammerstone, J.F.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Screening of foods containing proanthocyanidins and their structural characterization using LC-MS/MS and thiolytic degradation. J. Agric. Food Chem. 2003, 51, 7513–7521. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Cunningham, D.G.; Liu, H.Y.; Khoo, C.; Gu, L.W. Development of a thiolysis HPLC method for the analysis of procyanidins in cranberry products. J. Agric. Food Chem. 2018, 66, 2159–2167. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Butler, L.G. Condensed tannin purification and characterization of tannin-associated proteins. J. Agric. Food Chem. 1980, 28, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Seigler, D.S. Phytochemistry of Acacia—Sensu lato. Biochem. Syst. Ecol. 2003, 31, 845–873. [Google Scholar] [CrossRef]
- Hadacek, F. Secondary metabolites as plant traits: Current assessment and future perspectives. Crit. Rev. Plant Sci. 2002, 21, 273–322. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Iwasaki, M.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Harborne, J.B. Flavonoids and the evolution of the angiosperms. Biochem. Syst. Ecol. 1977, 5, 7–72. [Google Scholar] [CrossRef]
- Hoong, Y.B.; Pizzi, A.; Tahir, P.M.; Pasch, H. Characterization of Acacia mangium polyflavonoid tannins by MALDI-TOF mass spectrometry and CP-MAS 13C NMR. Eur. Polym. J. 2010, 46, 1268–1277. [Google Scholar] [CrossRef]
- Venter, P.B.; Senekal, N.D.; Kemp, G.; Amra-Jordaan, M.; Khan, P.; Bonnet, S.L.; Van der Westhuizen, J.H. Analysis of commercial proanthocyanidins. Part 3: The chemical composition of wattle (Acacia mearnsii) bark extract. Phytochemistry 2012, 83, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Hattas, D.; Julkunen-Tiitto, R. The quantification of condensed tannins in African savanna tree species. Phytochem. Lett. 2012, 5, 329–334. [Google Scholar] [CrossRef]
- Crestini, C.; Lange, H.; Bianchetti, G. Detailed Chemical Composition of Condensed Tannins via Quantitative P-31 NMR and HSQC Analyses: Acacia catechu, Schinopsis balansae, and Acacia mearnsii. J. Nat. Prod. 2016, 79, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Durmic, Z.; Moate, P.J.; Eckard, R.; Revell, D.K.; Williams, R.; Vercoe, P.E. In vitro screening of selected feed additives, plant essential oils and plant extracts for rumen methane mitigation. J. Sci. Food Agric. 2014, 94, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Carulla, J.E.; Kreuzer, M.; Machmuller, A.; Hess, H.D. Supplementation of Acacia mearnsii tannins decreases methanogenesis and urinary nitrogen in forage-fed sheep. Aust. J. Agric. Res. 2005, 56, 961–970. [Google Scholar] [CrossRef]
- Tavendale, M.H.; Meagher, L.P.; Pacheco, D.; Walker, N.; Attwood, G.T.; Sivakumaran, S. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Technol. 2005, 123, 403–419. [Google Scholar] [CrossRef]
- Getachew, G.; Makkar, H.P.S.; Becker, K. Tannins in tropical browses: Effects on in vitro microbial fermentation and microbial protein synthesis in media containing different amounts of nitrogen. J. Agric. Food Chem. 2000, 48, 3581–3588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, R.; Gung, B.W.; Tindall, S.; Gonzalez, J.M.; Halvorson, J.J.; Hagerman, A.E. Polyphenol-aluminum complex formation: Implications for aluminum tolerance in plants. J. Agric. Food Chem. 2016, 64, 3025–3033. [Google Scholar] [CrossRef] [PubMed]
- Andersen, O.M.; Jordheim, M. Chemistry of flavonoid-based colors in plants. In Comprehensive Natural Products II: Chemistry and Biology; Liu, H., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 3, pp. 547–614. [Google Scholar]
Sample Availability: Samples of purified CT from Desmodium paniculatum, Lespedeza stuevei, Lespedeza cuneata, Mimosa strigillosa, Desmanthus illinoensis, Leucaena retusa, and Acacia angustissima are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Plant | Subfamily | Subtribe | λmax in HCl-Methanol (nm) | Methane Production 1 (g/kg DM) |
|---|---|---|---|---|
| Desmodium paniculatum | Papilionoideae | - | 543 | 7.9 |
| Lespedeza stuevei | Papilionoideae | - | 543 | 4.9 |
| Lespedeza cuneata | Papilionoideae | - | 547 | 15.1 |
| Mimosa strigillosa | Mimosoideae | Mimoseae | 547 | 7.6 |
| Desmanthus illinoensis | Mimosoideae | Mimoseae | 547 | 24.9 |
| Neptunia lutea | Mimosoideae | Mimoseae | 547 | 19.7 |
| Leucaena retusa | Mimosoideae | Mimoseae | 538 | 40.7 |
| Acacia angustissima STP5 | Mimosoideae | Acacieae | 508 | 0.6 |
| Acacia angustissima STX | Mimosoideae | Acacieae | 505 | 0.8 |
| Plant Sample | PC/PD Ratio | cis/trans | % Galloyl | mDP | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Thiol | NMR | Thiol | NMR H/C-4 | NMR H/C-2 | Thiol | NMR H/C-4 | NMR H/C-2′,5′ | Thiol | NMR | |
| Desmodium paniculatum | 55/45 | 52.5/47.5 | 90/10 | 87.8/12.2 | 84.2/15.8 | None | None | None | 18.6 | ND 1 |
| Lespedeza stuevei | 42/58 | 41.4/58.6 | 44/56 | 34.8/65.2 | 33.9/66.1 | 3 | 2.4 | 1.0 | 9.3 | 6.7 |
| Lespedeza cuneata | 8/92 | 4.3/95.7 | 80/20 | 82.1/17.9 | 75.4/24.6 | None | ND 2 | 5.3 | 10.6 | 9.1 |
| Mimosa strigillosa | 15/85 | 15.3/84.7 | 89/11 | ND 3 | ND 4 | 40 | 50.8 | ND 5 | 7.6 | 6.1 |
| Desmanthus illinoensis | 3/97 | 1.8/98.2 | 98/2 | ND 3 | 96.2/3.8 | 75 | 76.2 | 87.5 | 6.0 | 5.97 |
| Neptunia lutea | 12/88 | 8.2/91.8 | 93/7 | ND 3 | 91.5/8.5 | 32 | 34.4 | 25.6 | 11.5 | 8.1 |
| Leucaena retusa | 99/1 | 98.6/1.4 | 98/2 | ND 3 | ND 4 | 29 | 21.3 | 34.3 | 39.2 | 6.3 |
| Flavan-3-ol | - | (Epi)afzelechin | (Epi)catechin | Epi(gallocatechin) | |
|---|---|---|---|---|---|
| 5-Deoxy Flavan-3-ol | Guibourtinidol | Fisetinidol | Robinetinidol | - | Gallate Ester |
| Plant Sample | 256.07 | 272.07 | 288.06 | 304.06 | 152.01 |
| Desmodium paniculatum | - | - | ++ | ++ | - |
| Lespedeza stuevei | - | - | ++ | ++ | + |
| Lespedeza cuneata | - | - | + | ++ | - |
| Desmanthus illinoensis | - | - | + | ++ | ++ |
| Neptunia lutea | - | + | + | ++ | + |
| Leucaena retusa | - | - | ++ | - | + |
| Acacia angustissima 2 | + | ++ | + | - | + |
| Plant Sample | Observed Mass | Formula | Exact Mass | Error (ppm) | Interpretation |
|---|---|---|---|---|---|
| Desmodium paniculatum | 2065.43585 | C105H85O45 | 2065.43630 | 0.22 | cat4-gallocat3 |
| Lespedeza stuevei | 1473.31433 | C75H61O32 | 1473.31461 | 0.19 | cat3-gallocat2 |
| Lespedeza cuneata | 1825.36676 | C90H73O42 | 1825.35766 | 4.99 | gallocat6 |
| Desmanthus illinoensis | 1369.22063 | C66H49O33 | 1369.21562 | 3.66 | gallocat3-gallate3 |
| Neptunia lutea | 3025.55520 | C135H125O80 | 3025.55958 | 1.45 | cat-gallocat8-gallate2 |
| Leucaena retusa | 1745.34814 | C89H69O38 | 1745.34670 | 0.83 | cat5-gallate2 |
| Acacia angustissima 2 | 1241.29289 | C67H53O24 | 1241.29269 | 0.17 | cat-gui-fis2-gallate |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naumann, H.; Sepela, R.; Rezaire, A.; Masih, S.E.; Zeller, W.E.; Reinhardt, L.A.; Robe, J.T.; Sullivan, M.L.; Hagerman, A.E. Relationships between Structures of Condensed Tannins from Texas Legumes and Methane Production During In Vitro Rumen Digestion. Molecules 2018, 23, 2123. https://doi.org/10.3390/molecules23092123
Naumann H, Sepela R, Rezaire A, Masih SE, Zeller WE, Reinhardt LA, Robe JT, Sullivan ML, Hagerman AE. Relationships between Structures of Condensed Tannins from Texas Legumes and Methane Production During In Vitro Rumen Digestion. Molecules. 2018; 23(9):2123. https://doi.org/10.3390/molecules23092123
Chicago/Turabian StyleNaumann, Harley, Rebecka Sepela, Aira Rezaire, Sonia E. Masih, Wayne E. Zeller, Laurie A. Reinhardt, Jamison T. Robe, Michael L. Sullivan, and Ann E. Hagerman. 2018. "Relationships between Structures of Condensed Tannins from Texas Legumes and Methane Production During In Vitro Rumen Digestion" Molecules 23, no. 9: 2123. https://doi.org/10.3390/molecules23092123