
Comprehensive Analysis of Secondary Metabolites in Usnea longissima (Lichenized Ascomycetes, Parmeliaceae) Using UPLC-ESI-QTOF-MS/MS and Pro-Apoptotic Activity of Barbatic Acid


 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
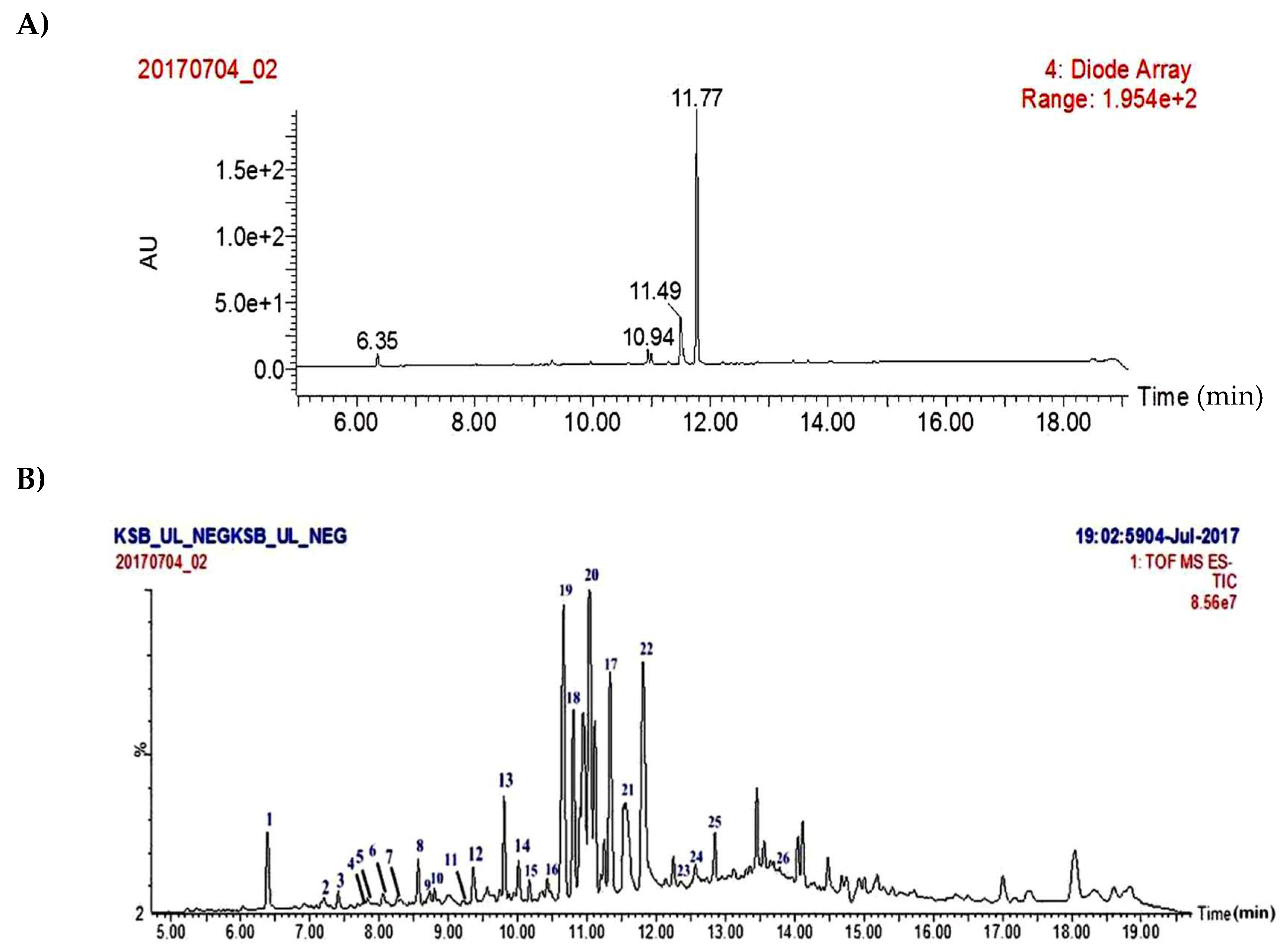
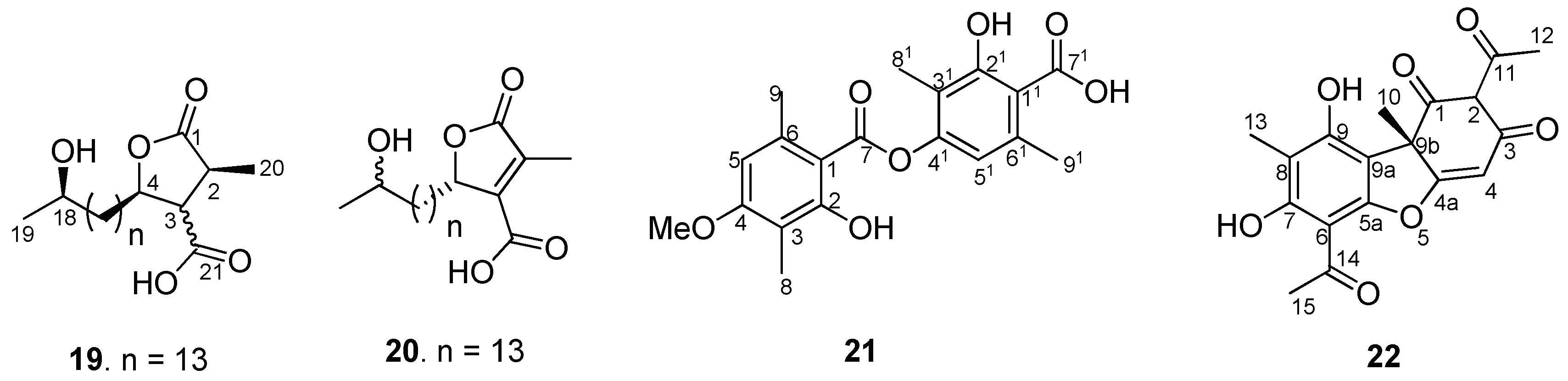
2.1. Isolation of Major Compounds from Acetone Extract of U. longissima and UPLC/PDA/ESI-QTOF-MS Analysis
2.2. Structural Fragmentation Analysis
2.3. Biological Activity
2.3.1. Cell Cycle Analysis
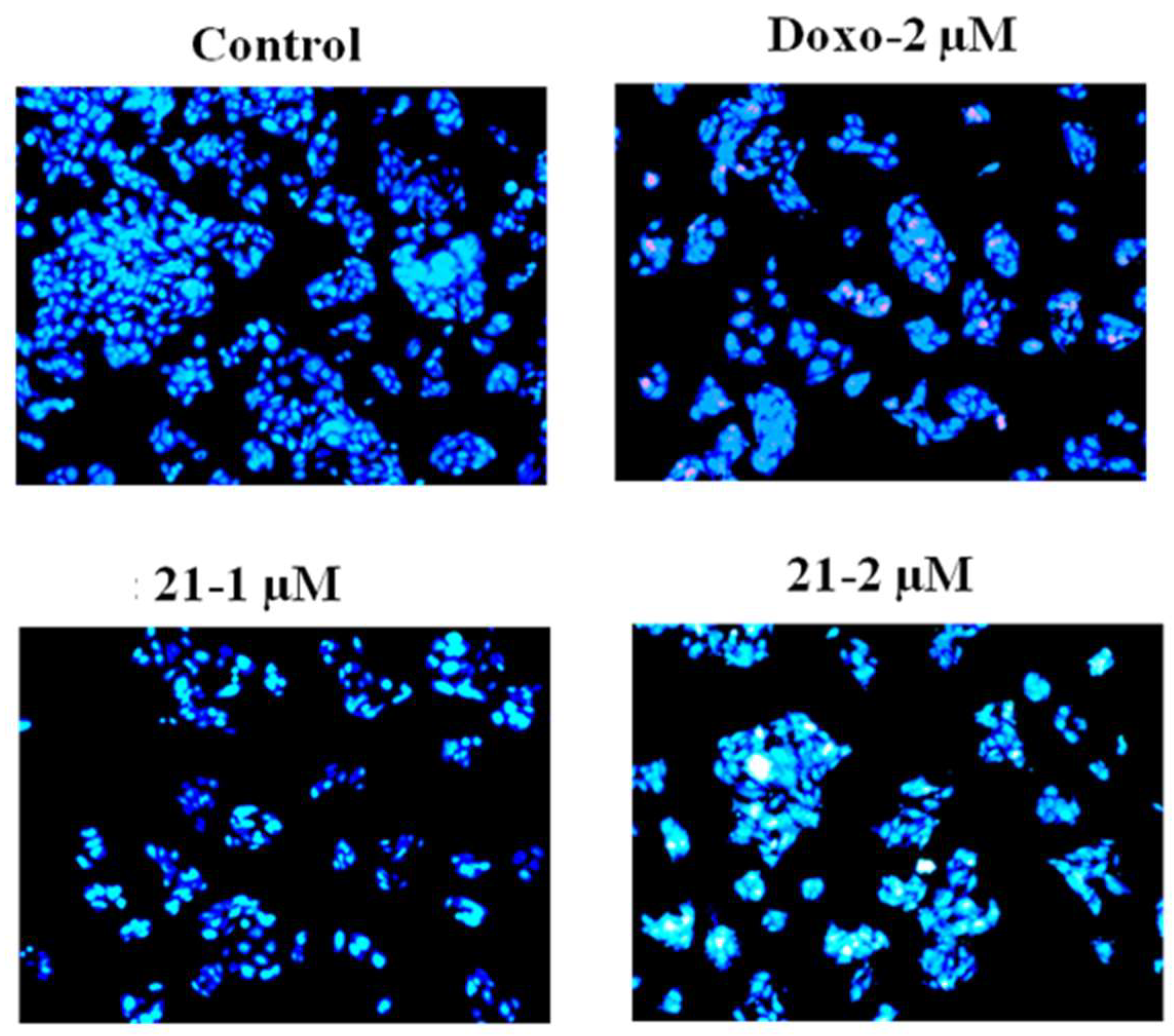
2.3.2. Hoechst Staining for Apoptosis
2.3.3. Annexin V Staining
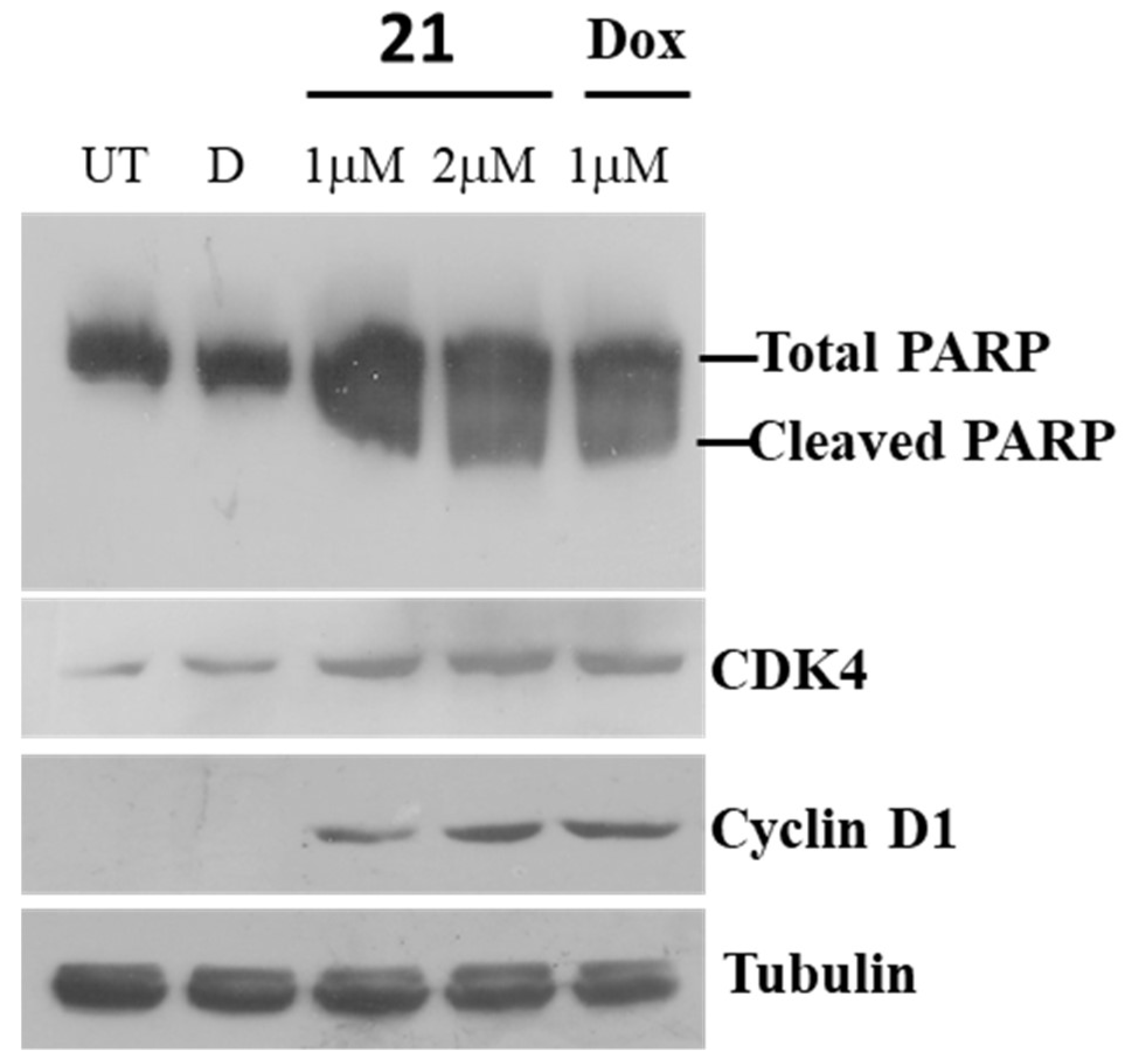
2.3.4. Immunoblot Assay for Poly (ADP-ribose) polymerase (PARP)
2.3.5. Caspase-3 Assay
3. Materials and Methods
3.1. Lichen Sample
3.2. Chemicals and Reagents
3.3. Extraction and Purification of Constituents from U. longissima
3.4. Spectral Data
3.5. Instrumental UPLC Conditions
3.6. Biological Activity
3.6.1. Cytotoxic Activity
3.6.2. Cell Cycle Analysis
3.6.3. Hoechst Staining
3.6.4. Annexin V Assay for Apoptosis
3.6.5. Western Blotting Assay
3.6.6. Caspase-3 Assay
3.6.7. Statistical Analysis
3.7. X-ray Crystal Data of 21
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Calcott, M.J.; Ackerley, D.F.; Knight, A.; Keyzers, R.A.; Owen, J.G. Secondary metabolism in the lichen symbiosis. Chem. Soc. Rev. 2018, 47, 1730–1760. [Google Scholar] [CrossRef] [PubMed]
- Culberson, C.F. Chemical and Botanical Guide to Lichen Products; University of North Carolina press: Chapel Hill, NC, USA, 1969. [Google Scholar]
- Elix, J.A. A Catalogue of Standardized Chromatographic Data and Biosynthetic Relationships for Lichens Substances, 3rd ed.; Research School of Chemistry, Australian National University: Canberra, Australia, 2014. [Google Scholar]
- Boustie, J.; Grube, M. Lichens-a promising source of bioactive secondary metabolites. Plant Genet. Resour. 2005, 3, 273–287. [Google Scholar] [CrossRef]
- Kayes, L.J.; Wormer, K.V.; Kofranek, D.; Sale, K.; McCune, B. Chemical variation of Usnea longissima Ach. in the central Oregon Coast Range. N. Am. Fungi 2008, 3, 1–10. [Google Scholar] [CrossRef]
- Prateeksha; Paliya, B.S.; Bajpai, R.; Jadaun, V.; Kumar, S.; Upreti, D.K.; Singh, B.R.; Nayaka, S.; Joshi, Y.; Singh, B.N. The genus Usnea: A potent phytomedicine with multifactorious ethnobotany, phytochemistry and pharmacology. RSC Adv. 2016, 6, 21672–21696. [Google Scholar] [CrossRef]
- Editorial Committee of the Administration Bureau of Traditional Chinese Medicine. Chinese Materia Medica (Zhong Hua Ben Cao), Vol. of Mongolian Medicine; Shanghai Science & Technology Press: Shanghai, China, 2004; pp. 253–254. [Google Scholar]
- Yu, X.; Guo, Q.; Su, G.; Yang, A.; Hu, Z.; Qu, C.; Wan, Z.; Li, R.; Tu, P.; Chai, X. Usnic acid derivatives with cytotoxic and antifungal activities from the lichen Usnea longissima. J. Nat. Prod. 2016, 79, 1373–1380. [Google Scholar] [CrossRef]
- Sun, C.; Liu, F.; Sun, J.; Li, J.; Wang, X. Optimisation and establishment of separation conditions of organic acids from Usnea longissima Ach. by pH-zone-refining counter-current chromatography: Discussion of the eluotropic sequence. J. Chromatogr. A 2016, 1427, 96–101. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Azizuddin; Jalil, S.; Rahman, A.U. Bioactive phenolic compounds from a medicinal lichen, Usnea longissima. Phytochemistry 2005, 66, 2346–2350. [Google Scholar] [CrossRef]
- Shukla, P.; Upreti, D.K.; Tewari, L.M. Lichen genus Usnea (Parmeliaceae, Ascomycota) in Uttarakhand, India. Curr. Res. Environ. Appl. Myco. 2014, 4, 188–201. [Google Scholar] [CrossRef]
- Mallavadhani, U.V.; Sudhakar, A.V.S.; Mahapatra, A.; Narasimhan, K.; Thirunavokkarasu, M.; Elix, J.A. Phenolic and steroidal constituents of the lichen Usnea longissima. Biochem. Syst. Ecol. 2004, 32, 95–97. [Google Scholar] [CrossRef]
- Ohmura, Y.; Skirina, I.; Skirin, F. Contribution to the knowledge of the genus Usnea (Parmeliaceae, Ascomycota) in Southern Far East Russia. Bull. Natl. Mus. Nat. Sci. Ser. B. 2017, 43, 1–10. [Google Scholar]
- Wang, H.; Yang, T.; Cheng, X.; Kwong, S.; Liu, C.; An, R.; Li, G.; Wang, X.; Wang, C. Simultaneous determination of usnic, diffractaic, evernic and barbatic acids in rat plasma by ultra-high-performance liquid chromatography–quadrupole exactive Orbitrap mass spectrometry and its application to pharmacokinetic studies. Biomed. Chromatogr. 2018, 32, 4123. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Siva, B.; Sarma, V.U.M.; Mohabe, S.; Reddy, A.M.; Boustie, J.; Tiwari, A.K.; Rao, N.R.; Babu, K.S. UPLC-MS/MS quantitative analysis and structural fragmentation study of five Parmotrema lichens from the Eastern Ghats. J. Pharm. Biomed. Anal. 2018, 156, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Laxinamu, J.; Tang, Y.X.; Bao, H.Y.; Bau, T. Chemical constituents from Usnea longissima, a traditional mongolian medicine. Zhongguo Zhong Yao Za Zhi 2013, 38, 2125–2128. [Google Scholar] [PubMed]
- Ghogomu, R.T.; Bodo, B. Structural elucidation of 13-acetoxylichesterinic and 13-acetoxyprotolichesterinic acids, two aliphatic lichen metabolites from Neuropogon trachycarpus. Phytochemistry 1982, 21, 2355–2358. [Google Scholar] [CrossRef]
- Nishitoba, Y.; Nishimura, I.; Nishiyama, T.; Mizutani, J. Lichen acids, plant growth inhibitors from Usnea longissima. Phytochemistry 1987, 26, 3181–3185. [Google Scholar] [CrossRef]
- Rashid, M.A.; Majid, M.A.; Quader, M.A. Complete NMR assignments of (+)-usnic acid. Fitoterapia 1999, 70, 113–115. [Google Scholar] [CrossRef]
- Salgado, F.; Albornoz, L.; Cortez, C.; Stashenko, E.; Urrea Vallejo, K.; Nagles, E.; Galicia Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary metabolite profiling of species of the genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2018, 23, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Siva, B.; Rama Rao, N.; Babu, K.S. Rapid identification of limonoids from Cipadessa baccifera and Xylocarpus granatum using ESI-Q-TOF-MS/MS and their structure-fragmentation study. J. Pharm. Biomed. Anal. 2018, 152, 224–233. [Google Scholar] [CrossRef]
- Cai, T.; Guo, Z.Q.; Xu, X.Y.; Wu, Z.J. Recent (2000–2015) developments in the analysis of minor unknown natural products based on characteristic fragment information using LC-MS. Mass Spectrom Rev. 2018, 37, 202–216. [Google Scholar] [CrossRef]
- Benitez, A.T.; Montalvo, M.R.; Sepulveda, B.; Castro, O.N.; Nagles, E.; Simirgiotis, M.J.; Beltran, O.G.; Areche, C. Metabolomic analysis of two Parmotrema lichens: P. robustum (Degel.) Hale and P. andinum (Mull. Arg.) Hale using UHPLC-ESI-OT-MS-MS. Molecules 2017, 22, 1861–1874. [Google Scholar] [CrossRef]
- Musharraf, S.G.; Kanwal, N.; Thadhani, V.M.; Choudhary, M.I. Rapid identification of lichen compounds based on the structure–fragmentation relationship using ESI-MS/MS analysis. Anal. Methods 2015, 7, 6066–6076. [Google Scholar] [CrossRef]
- Le Pogam, P.; Pillot, A.; Lohezic-Le Devehat, F.; Le Lamer, A.C.; Legouin, B.; Gadea, A.; Sauvager, A.; Ertz, D.; Boustie, J. Mass spectrometry as a versatile ancillary technique for the rapid in situ identification of lichen metabolites directly from TLC plates. The Lichenologist 2017, 49, 507–520. [Google Scholar] [CrossRef]
- McCune, B. Usnea in the Pacific Northwest. 2015. Available online: http://oregonstate.edu/~mccuneb/Usnea.PDF (accessed on 18 June 2019).
- Gagarina, L.V.; Poryadina, L.N.; Chesnokov, S.V.; Konoreva, L.A. The lichen genus Usnea Dill. exAdans. in the Sakha Republic (Yakutia). Bot. Pac. 2017, 6, 31–36. [Google Scholar] [CrossRef]
- Chan, K.T.; Meng, F.Y.; Li, Q.; Ho, C.Y.; Lam, T.S.; To, Y.; Lee, W.H.; Li, M.; Chu, K.H.; Toh, M. Cucurbitacin B induces apoptosis and S phase cell cycle arrest in BEL-7402 human hepatocellular carcinoma cells and is effective via oral administration. Cancer Lett. 2010, 294, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, J.P.; Moura, D.F.; Rocha, T.A.; Souza, T.G.S.; Falcao, E.P.S.; Silva, N.H. Potential cytotoxic of barbatic acid on MCF-7 cell line by MTT assay. SociedadeBrasileira de Bioquimica e Biologia Molecular SBBq. 2014, 01. [Google Scholar]
- Manojlovic, N.T.; Vasiljevic, P.; Juskovic, M.; Jankovic, S.N.S.; Andjelkovic, A.M. HPLC analysis and cytotoxic potential of extracts from the lichen, Thamnolia vermicularis var. Subuliformis. J. Med. Plant Res. 2010, 4, 817–823. [Google Scholar]
- Sukhramani, P.S.; Sukhramani, P.S.; Desai S., A.; Suthar, M.P. In-vitro cytotoxicity evaluation of novel N-substituted bis-benzimidazole derivatives for anti-lung and anti-breast cancer activity. Ann. of Biol. Res. 2011, 2, 51–59. [Google Scholar]
- Prasad, C.V.; Nayak, V.L.; Ramakrishna, S.; Mallavadhani, U.V. Novel menadione hybrids: Synthesis, anticancer activity, and cell-based studies. Chem. Biol. Drug Des. 2018, 91, 220–233. [Google Scholar] [CrossRef]
- Martin, S.J.; Finucane, D.M.; Amarante-Mendes, G.P.; O’Brien, G.A.; Green, D.R. Phosphatidylserine externalization during CD95-induced apoptosis of cells and cytoplasts requires ICE/CED-3 protease activity. J. Biol. Chem. 1996, 271, 28753–28756. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, S.; Song, J. Caspase-dependent apoptosis and independent poly (ADP-ribose) polymerase cleavage induced by transforming growth factor beta1. Int. J. Biochem. Cell Biol. 2004, 36, 223–234. [Google Scholar] [CrossRef]
- Brodo, I.M.; Sharnoff, S.D.; Sharnoff, S. Lichens of North America; Canadian Museum of Nature, Yale University Press: New Haven, CT, USA, 2001. [Google Scholar]
- Clerc, P. Usnea. In Nordic Lichen Flora: Parmeliaceae; Thell, A., Moberg, R., Forening, N.L., Eds.; Museum of Evolution, Uppsala University: Uppsala, Sweden, 2011; Volume 4, pp. 107–127. [Google Scholar]
- Subba Rao, A.V.; Swapna, K.; Shaik, S.P.; Nayak V., L.; Reddy T., S.; Sunkari, S.; Shaik, T.B.; Bagul, C.; Kamal, A. Synthesis and biological evaluation of cis-restricted triazole/tetrazole mimics of combretastatin-benzothiazole hybrids as tubulin polymerization inhibitors and apoptosis inducers. Bioorg. Med. Chem. 2017, 25, 977–999. [Google Scholar] [CrossRef] [PubMed]
- Amujuri, D.; Siva, B.; Poornima, B.; Sirisha, K.; Sarma, A.V.S.; Nayak V., L.; Tiwari, A.K.; Purushotham, U.; Suresh Babu, K. Synthesis and biological evaluation of Schizandrin derivatives as potential anti-cancer agents. Eur. J. Med. Chem. 2018, 149, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.F.; Nayak, V.L.; Budaganaboyina, P.; Mullagiri, K.; Sunkari, S.; Gour, J.; Kamal, A. Synthesis and biological evaluation of imidazo [2,1-b]thiazole-benzimidazole conjugates as microtubule-targeting agents. Bioorg. Chem. 2018, 77, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Vangala, J.R.; Dudem, S.; Jain, N.; Kalivendi, S.V. Regulation of PSMB5 protein and β subunits of mammalian proteasome by constitutively activated signal transducer and activator of transcription 3 (STAT3): Potential role in bortezomib-mediated anticancer therapy. J. Biol. Chem. 2014, 289, 12612–12622. [Google Scholar] [CrossRef] [PubMed]
- Kalivendi, S.V.; Konorev, E.A.; Cunningham, S.; Vanamala, S.K.; Kaji, E.H.; Joseph, J.; Kalyanaraman, B. Doxorubicin activates nuclear factor of activated T-lymphocytes and Fas ligand transcription: Role of mitochondrial reactive oxygen species and calcium. Biochem. J. 2005, 389, 527–539. [Google Scholar] [CrossRef] [PubMed]
- SMART & SAINT. Software Reference Manuals; Versions 6.28a & 5.625; Bruker Analytical X-ray Systems Inc.: Madison, WI, USA, 2001. [Google Scholar]
- Sheldrick, G.M. SHELXS97 and SHELXL97, Programs for Crystal Structure Solution and Refinement; University of Gottingen: Göttingen, Germany, 1997. [Google Scholar]
- Barroso Martins, M.C.; Rocha, T.A.; Santos Silva, T.D.; Cavalcanti Neto, M.P.; Da Silva Santos, N.P.; Da Silva, T.G.; Aguiar-Junoir, F.C.A.; Da Silva Falcao, E.P.; Pereira, E.C.; Da Silva, N.H. In vitro and In vivo antineoplastic activity of barbatic acid. Int. Arch. Med. 2016, 9, 1–9. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds 19, 20, 21 and 22 are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HeLa a | A549 b | MCF-7 c | DU-145 d | HEK-293 e | |
|---|---|---|---|---|---|
| Crude extract | 25.33 ± 1.23 | 9.11 ± 2.76 | 14.67 ± 2.85 | 20.88 ± 3.74 | 78.33 ± 2.11 |
| 19 | 22.67 ± 0.97 | 20.20 ± 1.99 | 20.01 ± 0.74 | 26.97 ± 2.92 | 55.87 ± 1.55 |
| 20 | 23.63 ± 0.74 | 19.13 ± 3.86 | 22.30 ± 4.05 | 26.10 ± 1.09 | 112.77 ± 3.82 |
| 21 | 3.02 ± 0.11 | 1.78 ± 0.62 | 3.16 ± 0.88 | 9.03 ± 2.11 | 124.90 ± 2.65 |
| 22 | 18.88 ± 2.06 | 13.80 ± 1.11 | 19.18 ± 2.05 | 24.21 ± 0.99 | 125.20 ± 1.98 |
| Doxorubicin | 1.55 ± 1.32 | 1.95 ± 0.97 | 1.62 ± 0.36 | 1.51 ± 0.84 | NT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, S.D.; Siva, B.; Kumar, K.; Babu, V.S.P.; Sravanthi, V.; Boustie, J.; Nayak, V.L.; Tiwari, A.K.; Rao, C.V.; Sridhar, B.; et al. Comprehensive Analysis of Secondary Metabolites in Usnea longissima (Lichenized Ascomycetes, Parmeliaceae) Using UPLC-ESI-QTOF-MS/MS and Pro-Apoptotic Activity of Barbatic Acid. Molecules 2019, 24, 2270. https://doi.org/10.3390/molecules24122270
Reddy SD, Siva B, Kumar K, Babu VSP, Sravanthi V, Boustie J, Nayak VL, Tiwari AK, Rao CV, Sridhar B, et al. Comprehensive Analysis of Secondary Metabolites in Usnea longissima (Lichenized Ascomycetes, Parmeliaceae) Using UPLC-ESI-QTOF-MS/MS and Pro-Apoptotic Activity of Barbatic Acid. Molecules. 2019; 24(12):2270. https://doi.org/10.3390/molecules24122270
Chicago/Turabian StyleReddy, S. Divya, Bandi Siva, Katragunta Kumar, V. S. Phani Babu, Vemireddy Sravanthi, Joel Boustie, V. Lakshma Nayak, Ashok K Tiwari, CH. V. Rao, B. Sridhar, and et al. 2019. "Comprehensive Analysis of Secondary Metabolites in Usnea longissima (Lichenized Ascomycetes, Parmeliaceae) Using UPLC-ESI-QTOF-MS/MS and Pro-Apoptotic Activity of Barbatic Acid" Molecules 24, no. 12: 2270. https://doi.org/10.3390/molecules24122270