
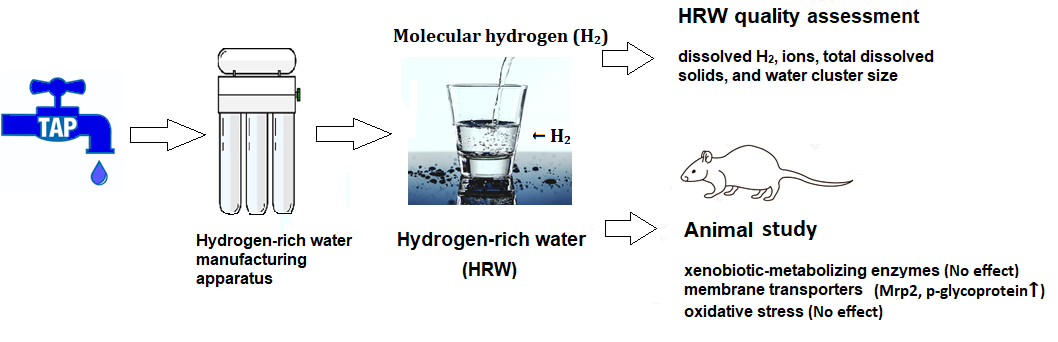
Intake of Molecular Hydrogen in Drinking Water Increases Membrane Transporters, p-Glycoprotein, and Multidrug Resistance-Associated Protein 2 without Affecting Xenobiotic-Metabolizing Enzymes in Rat Liver



Abstract

1. Introduction
2. Results
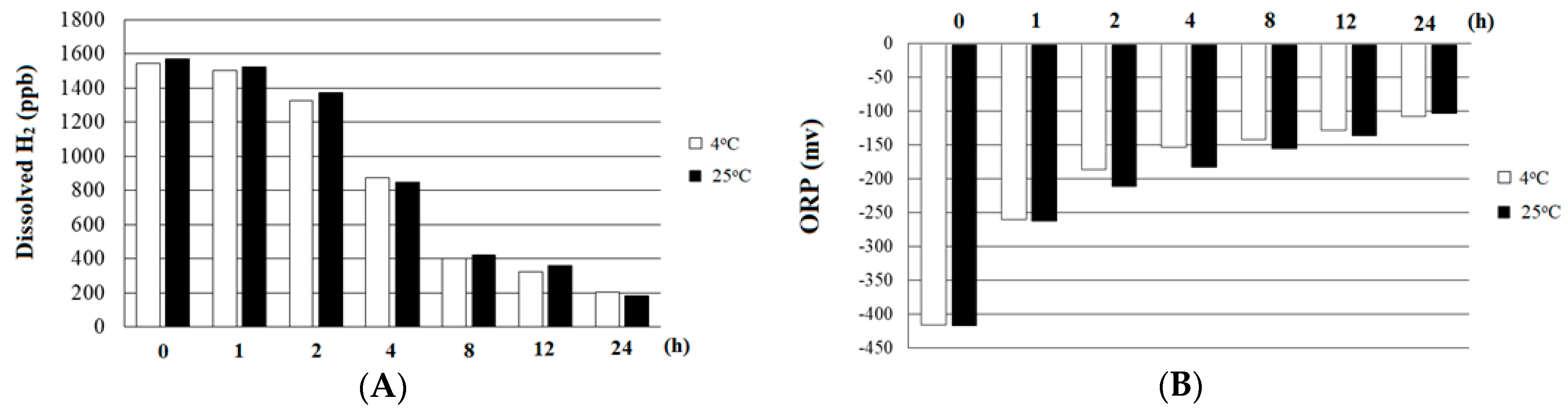
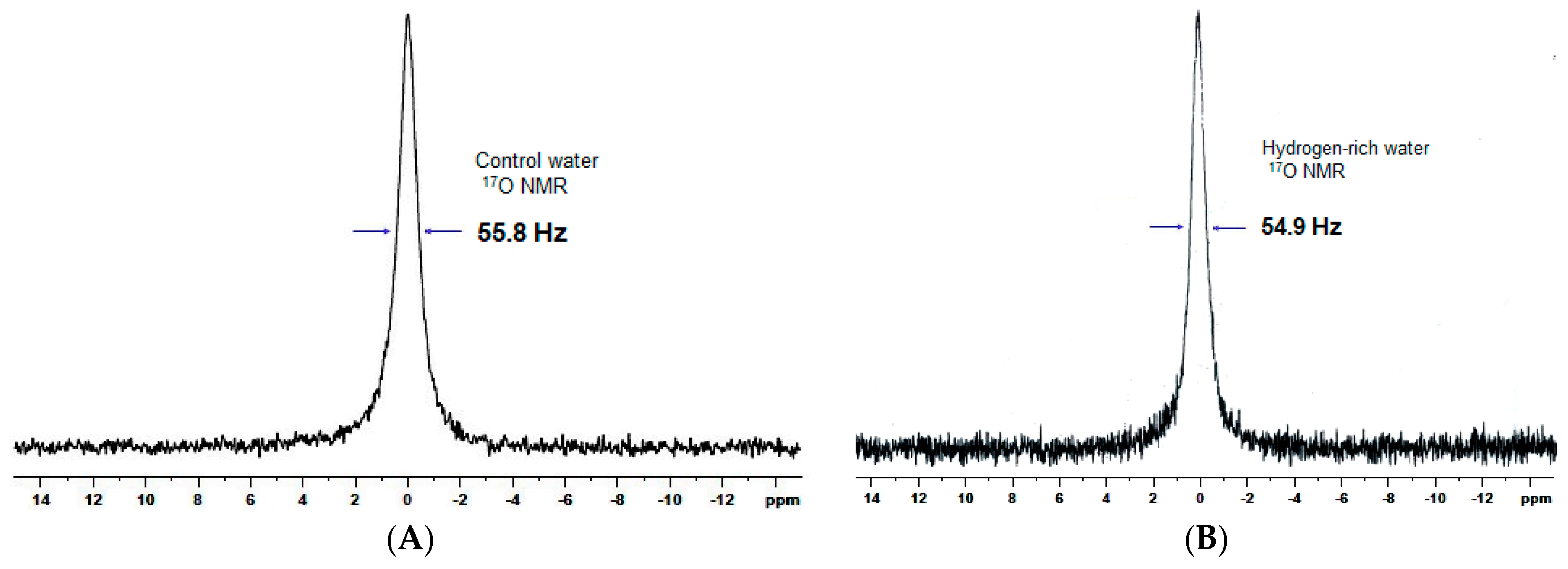
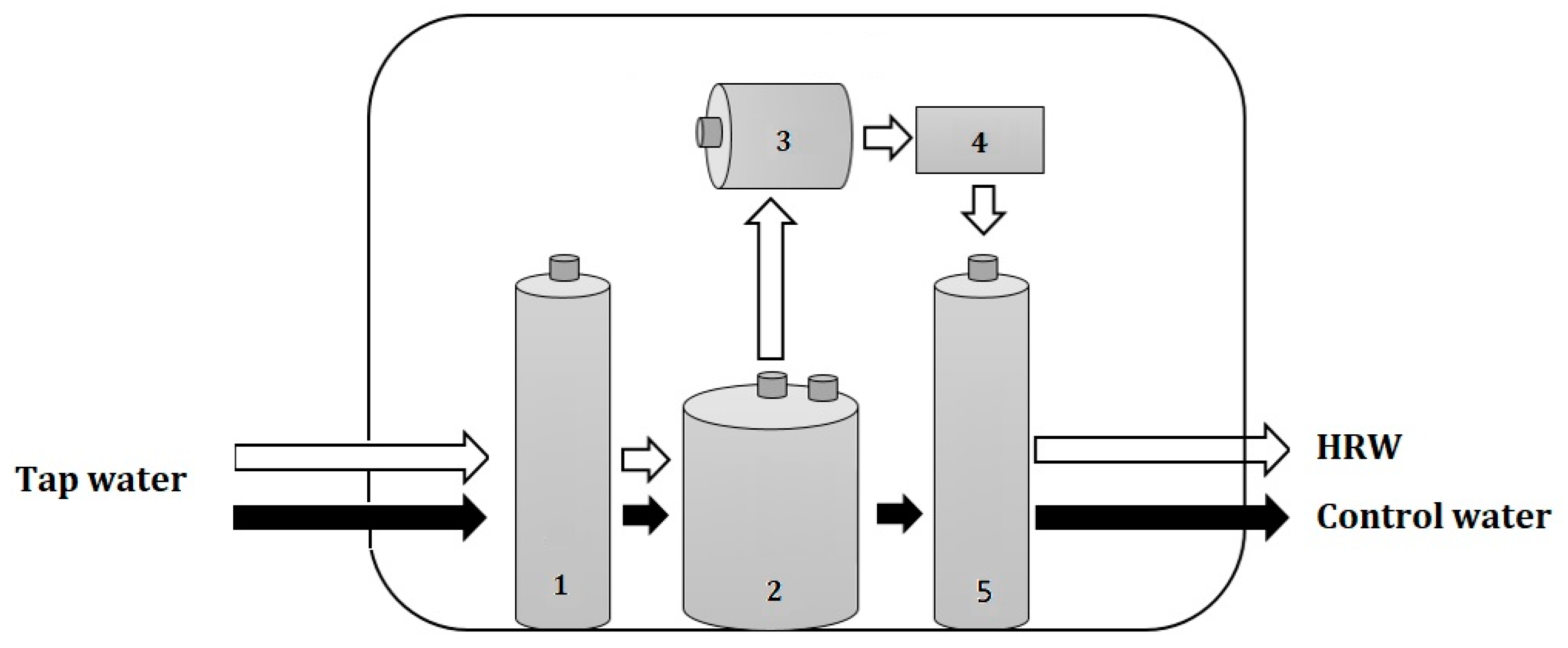
2.1. Quality of HRW
2.2. Body Weight, Tissue Weight, Water Drinking Volume, and Plasma Biochemical Parameters
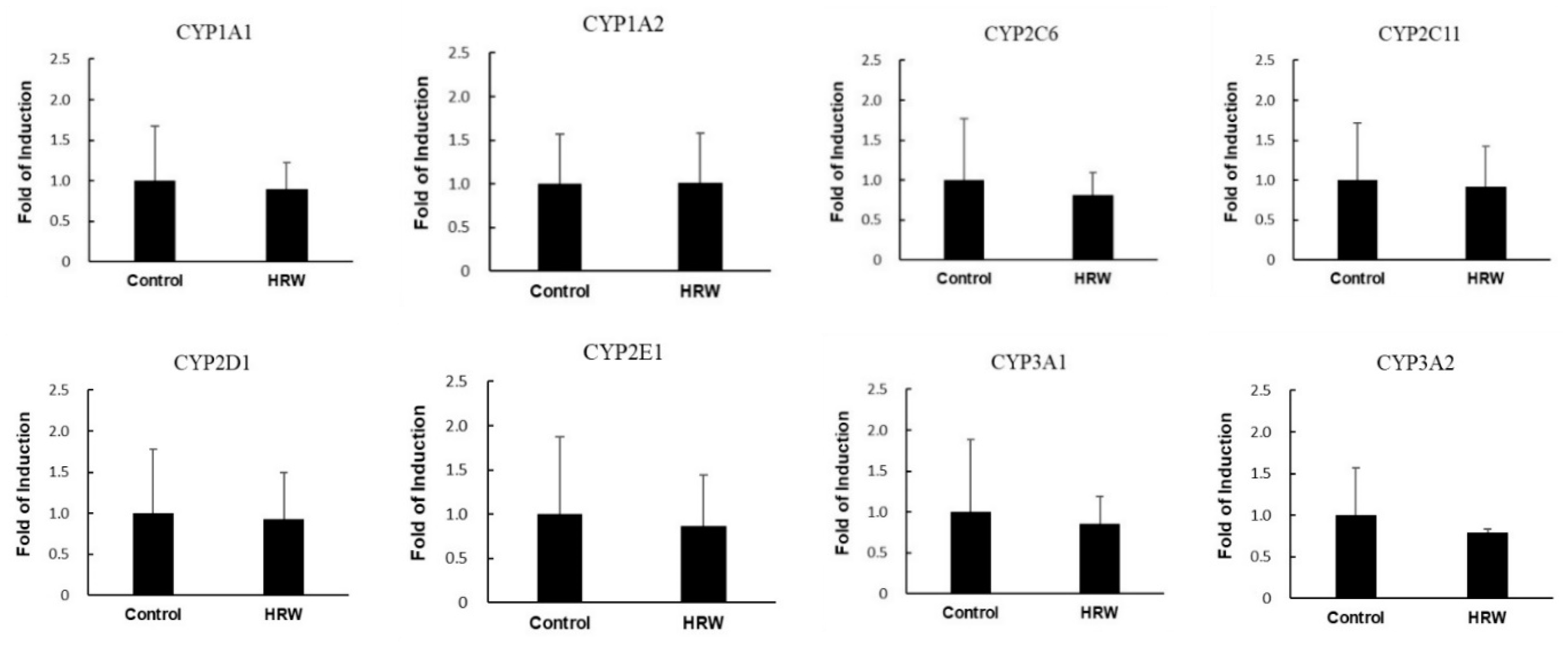
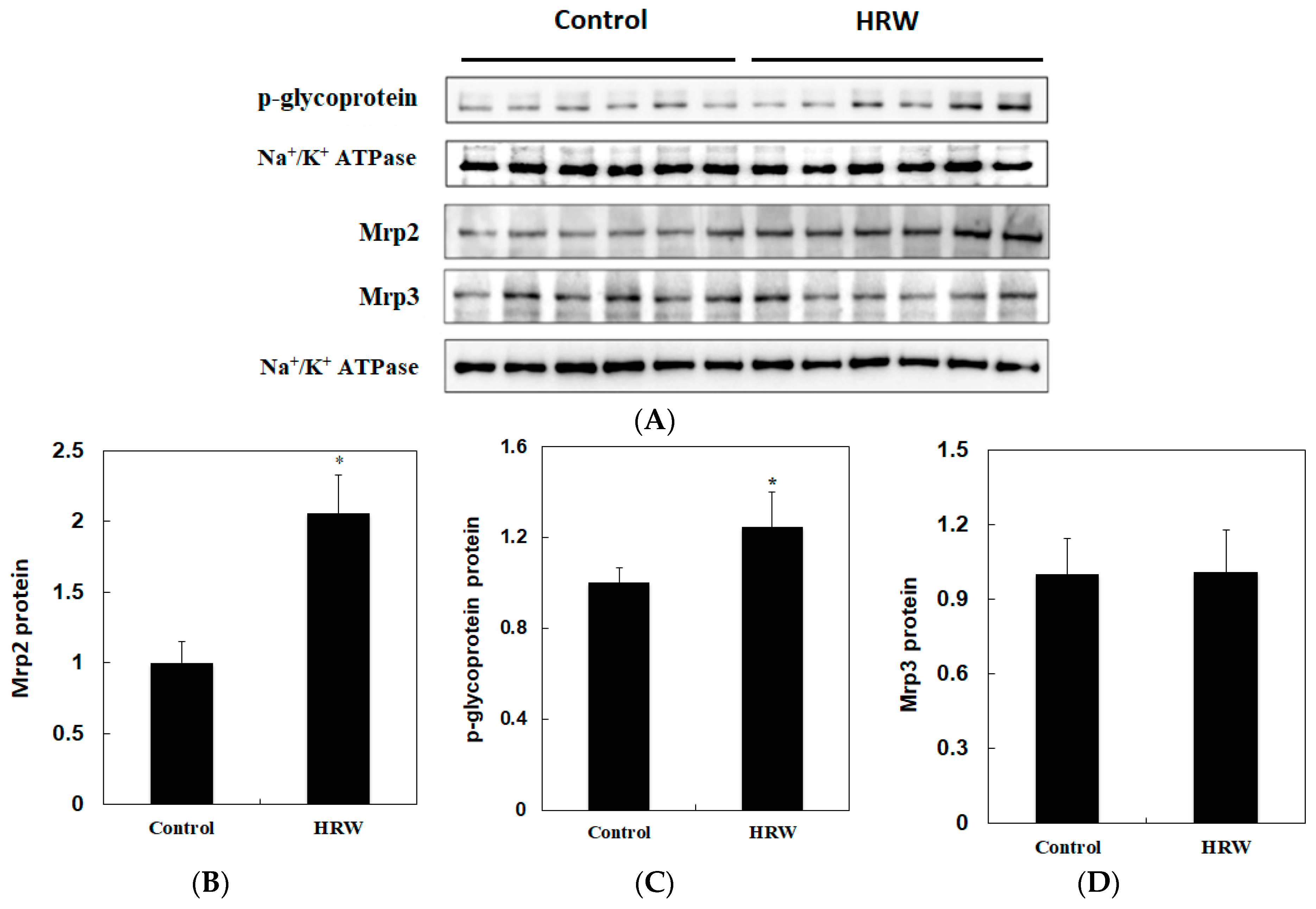
2.3. Activities of Xenobiotic-Metabolizing Enzymes and Membrane Transporters
2.4. Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. HRW Production
4.3. Determinations of the Quality of HRW
4.4. Animal Study
4.5. Preparation of Liver Microsomes
4.6. Xenobiotic-Metabolizing Enzyme Activity Assays
4.7. Determinations of GSH, GSH Peroxidase Activity, and Lipid Peroxidation
4.8. Determination of p-Glycoprotein and Mrp 2/3
4.9. Western Blot Analysis
4.10. Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rendić, S. Summary of information on human CYP enzymes: Human P450 metabolism data. Drug Metab. Rev. 2002, 34, 83–448. [Google Scholar] [CrossRef] [PubMed]
- James, L.P. Acetaminophen-induced hepatotoxicity. Drug Metab. Dispos. 2003, 31, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.J.; Gelboin, H.V. Role of Human Cytochromes P450 in the Metabolic Activation of Chemical Carcinogens and Toxins. Drug Metab. Rev. 1994, 26, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Arlt, V.M.; Krais, A.M.; Godschalk, R.W.; Riffo-Vasquez, Y.; Mrizova, I.; Roufosse, C.A.; Corbin, C.; Shi, Q.; Frei, E.; Stiborova, M.; et al. Pulmonary Inflammation Impacts on CYP1A1-Mediated Respiratory Tract DNA Damage Induced by the Carcinogenic Air Pollutant Benzo[a]pyrene. Toxicol. Sci. 2015, 146, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Kondraganti, S.R.; Jiang, W.; Jaiswal, A.K.; Moorthy, B. Persistent induction of hepatic AND pulmonary phase II enzymes by 3-methylcholanthrene in rats. Toxicol. Sci. 2008, 102, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V. Induction of phase II enzymes as a strategy in the chemoprevention of cancer and other degenerative diseases. Postępy Hig. Med. Doświadczalnej 2007, 61, 627–638. [Google Scholar]
- Kong, L.L.; Shen, G.L.; Wang, Z.Y.; Zhuang, X.M.; Xiao, W.B.; Yuan, M.; Gong, Z.H.; Li, H. Inhibition of p-Glycoprotein and Multidrug Resistance-Associated Protein 2 Regulates the Hepatobiliary Excretion and Plasma Exposure of Thienorphine and Its Glucuronide Conjugate. Front. Pharmacol. 2016, 7, 242. [Google Scholar] [CrossRef]
- Liu, Y.T.; Chen, Y.H.; Uramaru, N.; Lin, A.H.; Yang, H.T.; Lii, C.K.; Yao, H.T. Soy isoflavones reduce acetaminophen-induced liver injury by inhibiting cytochrome P-450-mediated bioactivation and glutathione depletion and increasing urinary drug excretion in rats. J. Funct. Foods 2016, 26, 135–143. [Google Scholar] [CrossRef]
- Slitt, A.L.; Cherrington, N.J.; Maher, J.M.; Klaassen, C.D. Induction of multidrug resistance protein 3 in rat liver is associated with altered vectorial excretion of acetaminophen metabolites. Drug Metab. Dispos. 2003, 31, 1176–1186. [Google Scholar] [CrossRef]
- Setsukinai, K.; Urano, Y.; Kakinuma, K.; Majima, H.J.; Nagano, T. Development of novel fluorescence probes that can reliably detect reactive oxygen species and distinguish specific species. J. Biol. Chem. 2003, 278, 3170–3175. [Google Scholar] [CrossRef]
- Ohta, S. Molecular hydrogen as a preventive and therapeutic medical gas: Initiation, development and potential of hydrogen medicine. Pharmacol. Ther. 2014, 144, 1–11. [Google Scholar] [CrossRef]
- Cejka, C.; Kubinova, S.; Cejkova, J. The preventive and therapeutic effects of molecular hydrogen in ocular diseases and injuries where oxidative stress is involved. Free Radic. Res. 2019, 53, 237–247. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the 1st Addendum; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Yan, Y.; Ou, X.X.; Zhang, H.P.; Shao, Y. Effects of nano-materials on 17O NMR line-width of water clusters. J. Mol. Struct. 2013, 1051, 211–214. [Google Scholar] [CrossRef]
- Szcześ, A.; Chibowski, E.; Hołysz, L.; Rafalski, P. Effects of static magnetic field on water at kinetic condition. Chem. Eng. Process. Process. Intensif. 2011, 50, 124–127. [Google Scholar] [CrossRef]
- Ayrton, A.; Morgan, P. Role of transport proteins in drug absorption, distribution and excretion. Xenobiotica 2001, 31, 469–497. [Google Scholar] [CrossRef]
- Ghanem, C.I.; Gómez, P.C.; Arana, M.C.; Perassolo, M.; Ruiz, M.L.; Villanueva, S.S.; Ochoa, E.J.; Catania, V.A.; Bengochea, L.A.; Mottino, A.D. Effect of acetaminophen on expression and activity of rat liver multidrug resistance-associated protein 2 and p-glycoprotein. Biochem. Pharmacol. 2004, 68, 791–798. [Google Scholar] [CrossRef]
- Yang, C.Y. Calcium and Magnesium in Drinking Water and Risk of Death from Cerebrovascular Disease. Stroke 1998, 29, 411–414. [Google Scholar] [CrossRef]
- Catling, L.A.; Abubakar, I.; Lake, I.R.; Swift, L.; Hunter, P.R. A systematic review of analytical observational studies investigating the association between cardiovascular disease and drinking water hardness. J. Water Heal. 2008, 6, 433–442. [Google Scholar] [CrossRef]
- Leurs, L.J.; Schouten, L.J.; Mons, M.N.; Goldbohm, R.A.; van den Brandt, P.A. Relationship between tap water hardness, magnesium, and calcium concentration and mortality due to ischemic heart disease or stroke in The Netherlands. Environ. Health Perspect 2010, 118, 414–420. [Google Scholar] [CrossRef]
- Barooti, A.; Kamran, M.; Kharazmi, F.; Eftakhar, E.; Malekzadeh, K.; Talebi, A.; Soltani, N. Effect of oral magnesium sulfate administration on blood glucose hemostasis via inhibition of gluconeogenesis and FOXO1 gene expression in liver and muscle in diabetic rats. Biomed. Pharmacother. 2019, 109, 1819–1825. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kim, H.K. Anti-diabetic effects of electrolyzed reduced water in streptozotocin-induced and genetic diabetic mice. Life Sci. 2006, 79, 2288–2292. [Google Scholar] [CrossRef]
- Kajiyama, S.; Hasegawa, G.; Asano, M.; Hosoda, H.; Fukui, M.; Nakamura, N.; Kitawaki, J.; Imai, S.; Nakano, K.; Ohta, M.; et al. Supplementation of hydrogen-rich water improves lipid and glucose metabolism in patients with type 2 diabetes or impaired glucose tolerance. Nutr. Res. 2008, 28, 137–143. [Google Scholar] [CrossRef]
- Yao, H.T.; Hsu, Y.R.; Lii, C.K.; Lin, A.H.; Chang, K.H.; Yang, H.T. Effect of commercially available green and black tea beverages on drug-metabolizing enzymes and oxidative stress in Wistar rats. Food Chem. Toxicol. 2014, 70, 120–127. [Google Scholar] [CrossRef]
- Hanioka, N.; Jinno, H.; Tanaka-Kagawa, T.; Nishimura, T.; Ando, M. Determination of UDP-glucuronosyltransferase UGT1A6 activity in human and rat liver microsomes by HPLC with UV detection. J. Pharm. Biomed. Anal. 2001, 25, 65–75. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-Transferases. Methods Enzymol. 1981, 77, 398–405. [Google Scholar]
- Guan, X.; Hoffman, B.; Dwivedi, C.; Matthees, D.P. A simultaneous liquid chromatography/mass spectrometric assay of glutathione, cysteine, homocysteine and their disulfides in biological samples. J. Pharm. Biomed. Anal. 2003, 31, 251–261. [Google Scholar] [CrossRef]
- Mohandas, J.; Marshall, J.J.; Duggin, G.G.; Horvath, J.S.; Tiller, D.J. Low activities of glutathione-related enzymes as factors in the genesis of urinary bladder cancer. Cancer Res. 1984, 44, 5086–5091. [Google Scholar]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Aleksunes, L.M.; Scheffer, G.L.; Jakowski, A.B.; Pruimboom-Brees, I.M.; Manautou, J.E. Coordinated expression of multidrug resistance-associated proteins (Mrps) in mouse liver during toxicant-induced injury. Toxicol. Sci. 2006, 89, 370–379. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | HRW |
|---|---|---|
| H2 (ppb) | 0 | 1550 |
| ORP (mv) | 292.6 | −452.7 |
| TDS (g/L) | 197.3 | 196.7 |
| Salt (kU.m) | 149.3 | 149.3 |
| EC (ds/m) | 197 | 197 |
| DO (mg/L) | 6.1 | 5.8 |
| pH | 7.6 | 9.3 |
| Ions (ppm) | ||
| Ca2+ | 28.0 | 9.1 |
| Mg2+ | 10.2 | 22.8 |
| Na+ | 4.8 | 5.4 |
| K+ | 1 | 1 |
| Cl− | 2.9 | 3.2 |
| SO42− | 38.5 | 59.8 |
| Water cluster (Hz) | 55.8 | 54.9 |
| Parameters | Control | HRW |
|---|---|---|
| Total cholesterol (mg/dL) | 72.0 ± 8.2 | 78.9 ± 9.1 |
| Triglyceride (mg/dL) | 53.9 ± 20.5 | 58.2 ± 13.3 |
| Glucose (mg/dL) | 155.9 ± 10.3 | 143.9 ± 8.1 * |
| BUN (mg/dL) | 16.8 ± 2.4 | 18.8 ± 2.6 |
| Creatinine (mg/dL) | 0.31 ± 0.02 | 0.31 ± 0.03 |
| Uric acid (mg/dL) | 2.2 ± 0.6 | 2.0 ± 0.3 |
| ALT (U/L) | 44.6 ± 6.1 | 44.3 ± 5.1 |
| TBARS (nmol/mL) | 0.6 ± 0.1 | 0.6 ± 0.1 |
| GSH (nmol/mL) | 6.4 ± 2.6 | 6.5 ± 2.4 |
| Na+ (mg/dL) | 140.2 ± 1.6 | 144.0 ± 5.9 |
| K+ (mg/dL) | 6.1 ± 0.3 | 6.1 ± 0.3 |
| Cl− (mg/dL) | 1.3 ± 0.1 | 1.4 ± 0.2 |
| Ca2+ (mg/dL) | 91.5 ± 1.1 | 93.3 ± 3.9 |
| Mg2+ (mg/dL) | 2.9 ± 0.2 | 3.0 ± 0.2 |
| Control | HRW | |
|---|---|---|
| Phase I enzymes (pmol/min/mg protein) | ||
| Testosterone 6β-hydroxylase (CYP3A) | 682.3 ± 75.6 | 640.9 ± 168.1 |
| Diclofenac 4-hydroxylase (CYP2C) | 155.9 ± 10.3 | 147.9 ± 8.1 |
| Dextromethorphan O-demethylase (CYP2D) | 51.9 ± 6.1 | 45.2 ± 4.6 |
| Nitrophenol 6-hydroxylase (CYP2E1) | 121.1 ± 12.3 | 109.8 ± 15.6 |
| Ethoxyresorufin O-deethylase (CYP1A1) | 167.8 ± 17.2 | 171.5 ± 10.1 |
| Methoxyresorufin O-demethylase (CYP1A2) | 133.2 ± 18.2 | 140.6 ± 17.4 |
| Pentoxyresorufin O-depentylase (CYP2B) | 61.9 ± 10.4 | 62.3 ± 11.3 |
| Lauric acid 12-hydroxylauric acid (CYP4A) | 1524 ± 125 | 1488 ± 132 |
| Phase II enzymes (nmol/min/mg protein) | ||
| Glutathione S-transferase (GST) | 1072 ± 175 | 1132 ± 88 |
| UDP-glucurosyltransferase (UGT) | 23.4 ± 2.3 | 25.5 ± 3.0 |
| Control | HRW | |
|---|---|---|
| GSH (nmol/mg protein) | 4.1 ± 0.7 | 5.0 ± 1.4 |
| GSSG (nmol/mg protein) | 0.14 ± 0.03 | 0.16 ± 0.02 |
| GSH/GSSG | 29.4 ± 4.2 | 31.3 ± 10.2 |
| GSH peroxidase (nmol/min/mg protein) | 314.3 ± 25.6 | 302.0 ± 41.3 |
| TBARS (nmol/g protein) | 18.8 ± 3.6 | 18.9 ± 2.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, H.-T.; Yang, Y.-H.; Li, M.-L. Intake of Molecular Hydrogen in Drinking Water Increases Membrane Transporters, p-Glycoprotein, and Multidrug Resistance-Associated Protein 2 without Affecting Xenobiotic-Metabolizing Enzymes in Rat Liver. Molecules 2019, 24, 2627. https://doi.org/10.3390/molecules24142627
Yao H-T, Yang Y-H, Li M-L. Intake of Molecular Hydrogen in Drinking Water Increases Membrane Transporters, p-Glycoprotein, and Multidrug Resistance-Associated Protein 2 without Affecting Xenobiotic-Metabolizing Enzymes in Rat Liver. Molecules. 2019; 24(14):2627. https://doi.org/10.3390/molecules24142627
Chicago/Turabian StyleYao, Hsien-Tsung, Yu-Hsuan Yang, and Mei-Ling Li. 2019. "Intake of Molecular Hydrogen in Drinking Water Increases Membrane Transporters, p-Glycoprotein, and Multidrug Resistance-Associated Protein 2 without Affecting Xenobiotic-Metabolizing Enzymes in Rat Liver" Molecules 24, no. 14: 2627. https://doi.org/10.3390/molecules24142627
APA StyleYao, H.-T., Yang, Y.-H., & Li, M.-L. (2019). Intake of Molecular Hydrogen in Drinking Water Increases Membrane Transporters, p-Glycoprotein, and Multidrug Resistance-Associated Protein 2 without Affecting Xenobiotic-Metabolizing Enzymes in Rat Liver. Molecules, 24(14), 2627. https://doi.org/10.3390/molecules24142627