Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint



Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of Mint EOs and Total Phenolic Compounds
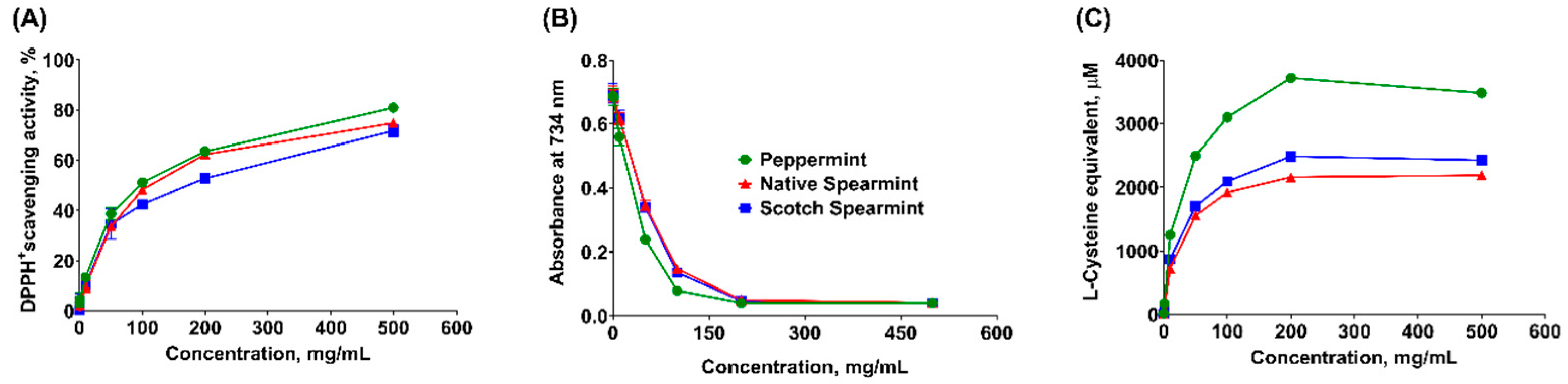
2.2. Chemical-Based Antioxidant Activity Assays
2.3. Lipid Peroxidation Assay
2.4. Cellular-Based Antioxidant Activity Assay
Intracellular Antioxidant Activity Assay
2.5. In Vivo Antioxidant Analysis with Nematode Model
3. Materials and Methods
3.1. Preparation of Mint Essential Oil
3.2. Analyses of Chemical Composition
3.3. Chemical-Based Antioxidant Assays
3.3.1. DPPH Radical Scavenging Capacity Assay
3.3.2. Trolox Equivalent Antioxidant Capacity Assay
3.3.3. Reducing Power Assay
3.4. Lipid Peroxidation Assay
3.5. Cellular-Based Antioxidant Activity Assay
3.5.1. Cell Culture
3.5.2. Cell Viability Assay
3.5.3. Cellular Antioxidant Activity (CAA) assay
3.5.4. Intracellular Antioxidant Activity (IAA) Assay
3.6. In Vivo Antioxidant Activity in Caenorhabditis Elegans Model
3.6.1. Cultures of Caenorhabditis Elegans (C. elegans)
3.6.2. Oxidative Stress Assays in C. elegans
3.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Valko, M.; Rhodes, C.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Shurson, G.C.; Kerr, B.J.; Hanson, A.R. Evaluating the quality of feed fats and oils and their effects on pig growth performance. J. Anim. Sci. Biotechnol. 2015, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Markers of oxidative status in plasma and erythrocytes of transition dairy cows during hot season. J. Dairy Sci. 2002, 85, 2173–2179. [Google Scholar] [CrossRef]
- Halliwell, B.; Zhao, K.; Whiteman, M. The gastrointestinal tract: A major site of antioxidant action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, A.; Parker, R.D.; Abdollahi, M. Oxidative stress and pathogenesis of inflammatory bowel disease: An epiphenomenon or the cause? Dig. Dis. Sci. 2007, 52, 2015–2021. [Google Scholar] [CrossRef]
- Fellenberg, M.A.; Speisky, H. Antioxidants: Their effects on broiler oxidative stress and its meat oxidative stability. World’s Poult. Sci. J. 2007, 62, 53–70. [Google Scholar] [CrossRef]
- Liang, F.; Jiang, S.; Mo, Y.; Zhou, G.; Yang, L. Consumption of oxidized soybean oil increased intestinal oxidative stress and affected intestinal immune variables in yellow-feathered broilers. Asian-Australas. J. Anim. Sci. 2015, 28, 1194–1201. [Google Scholar] [CrossRef]
- Hirose, M.; Takesada, Y.; Tanaka, H.; Tamano, S.; Kato, T.; Shirai, T. Carcinogenicity of antioxidants BHA, caffeic acid, sesamol, 4-methoxyphenol and catechol at low doses, either alone or in combination, and modulation of their effects in a rat medium-term multi-organ carcinogenesis model. Carcinogenesis 1998, 19, 207–212. [Google Scholar] [CrossRef]
- Safer, A.M.; Al-Nughamish, A.J. Hepatotoxicity induced by the anti-oxidant food additive, butylated hydroxytoluene (BHT), in rats: An electron microscopical study. Histol. Histopathol. 1999, 14, 391–406. [Google Scholar]
- Sun, B.; Fukuhara, M. Effects of co-administration of butylated hydroxytoluene, butylated hydroxyanisole and flavonoids on the activation of mutagens and drug-metabolizing enzymes in mice. Toxicology 1997, 122, 61–72. [Google Scholar] [CrossRef]
- Baydar, N.G.; Özkan, G.; Sağdiç, O. Total phenolic contents and antibacterial activities of grape (Vitis vinifera L.) extracts. Food Control 2004, 15, 335–339. [Google Scholar] [CrossRef]
- Sökmen, M.; Serkedjieva, J.; Daferera, D.; Gulluce, M.; Polissiou, M.; Tepe, B.; Akpulat, H.A.; Sahin, F.; Sokmen, A. In vitro antioxidant, antimicrobial, and antiviral activities of the essential oil and various extracts from herbal parts and callus cultures of Origanum acutidens. J. Agric. Food Chem. 2004, 52, 3309–3312. [Google Scholar] [CrossRef] [PubMed]
- Dundar, E.; Olgun, E.G.; Isiksoy, S.; Kurkcuoglu, M.; Baser, K.H.C.; Bal, C. The effects of intra-rectal and intra-peritoneal application of Origanum onites L. essential oil on 2,4,6-trinitrobenzenesulfonic acid-induced colitis in the rat. Exp. Toxicol. Pathol. 2008, 59, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, M.; Che, T.; Bravo, D.; Pettigrew, J. Anti-inflammatory effects of several plant extracts on porcine alveolar macrophages in vitro. J. Anim. Sci. 2012, 90, 2774–2783. [Google Scholar] [CrossRef]
- Amorati, R.; Foti, M.C.; Valgimigli, L. Antioxidant activity of essential oils. J. Agric. Food Chem. 2013, 61, 10835–10847. [Google Scholar] [CrossRef]
- Ruberto, G.; Baratta, M.T. Antioxidant activity of selected essential oil components in two lipid model systems. Food Chem. 2000, 69, 167–174. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef]
- Oh, J.; Jo, H.; Cho, A.R.; Kim, S.-J.; Han, J. Antioxidant and antimicrobial activities of various leafy herbal teas. Food Control 2013, 31, 403–409. [Google Scholar] [CrossRef]
- Arzani, A.; Zeinali, H.; Razmjo, K. Iron and magnesium concentrations of mint accessions (Mentha spp.). Plant Physiol. Biochem. 2007, 45, 323–329. [Google Scholar] [CrossRef]
- Bensabah, F.; Houbairi, S.; Essahli, M.; Lamiri, A.; Naja, J. Chemical composition and inhibitory effect of the essential oil from mentha spicata irrigated by wastewater on the corrosion of aluminum in 1 molar hydrochloric acid. Port. Electrochim. Acta 2013, 31, 195–206. [Google Scholar] [CrossRef]
- Riachi, L.G.; De Maria, C.A. Peppermint antioxidants revisited. Food Chem. 2015, 176, 72–81. [Google Scholar] [CrossRef]
- Naidu, J.R.; Ismail, R.; Yeng, C.; Sasidharan, S.; Kumar, P. Chemical composition and antioxidant activity of the crude methanolic extracts of Mentha spicata. J. Phytol. 2012, 4, 13–18. [Google Scholar]
- Dorman, H.J.; Kosar, M.; Kahlos, K.; Holm, Y.; Hiltunen, R. Antioxidant properties and composition of aqueous extracts from Mentha species, hybrids, varieties, and cultivars. J. Agric. Food Chem. 2003, 51, 4563–4569. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Cantrell, C.L.; Astatkie, T.; Hristov, A. Yield, content, and composition of peppermint and spearmints as a function of harvesting time and drying. J. Agric. Food Chem. 2010, 58, 11400–11407. [Google Scholar] [CrossRef]
- Khanna, R.; MacDonald, J.K.; Levesque, B.G. Peppermint oil for the treatment of irritable bowel syndrome: A systematic review and meta-analysis. J. Clin. Gastroenterol. 2014, 48, 505–512. [Google Scholar] [CrossRef]
- Oh, J.Y.; Park, M.A.; Kim, Y.C. Peppermint oil promotes hair growth without toxic signs. Toxicol. Res. 2014, 30, 297–304. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity and potential health benefits of peppermint tea (Mentha piperita L.). Phytother. Res. 2006, 20, 619–633. [Google Scholar] [CrossRef]
- Salim, R. Carvone content and chemical composition in spearmint (mentha spicata var. viridis l.) as affected by herb storage under ambient temperature. J. Food Nutr. Popul. Health 2016, 1, 1–5. [Google Scholar]
- Buleandra, M.; Oprea, E.; Popa, D.E.; David, I.G.; Moldovan, Z.; Mihai, I.; Badea, I.A. Comparative chemical analysis of mentha piperita and m. spicata and a fast assessment of commercial peppermint teas. Nat. Prod. Commun. 2016, 11, 551–555. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Chauhan, R.S.; Kaul, M.K.; Shahi, A.K.; Kumar, A.; Ram, G.; Tawa, A. Chemical composition of essential oils in Mentha spicata L. accession [IIIM(J)26] from North-West Himalayan region, India. Ind. Crop. Prod. 2009, 29, 654–656. [Google Scholar] [CrossRef]
- Clark, R.J.; Menary, R.C. The effect of two harvests per year on the yield and composition of Tasmanian peppermint oil (Mentha piperita L.). J. Sci. Food Agric. 1984, 35, 1191–1195. [Google Scholar] [CrossRef]
- Loganayaki, N.; Siddhuraju, P.; Manian, S. Antioxidant activity and free radical scavenging capacity of phenolic extracts from Helicteres isora L. and Ceiba pentandra L. J. Food Sci. Technol. 2013, 50, 687–695. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Neng, N.R.; Nogueira, J.M.; Saraiva, J.A.; Nunes, M.L. Chemical composition and antibacterial and antioxidant properties of commercial essential oils. Ind. Crop. Prod. 2013, 43, 587–595. [Google Scholar] [CrossRef]
- Ciftci, O.; Ozdemir, I.; Tanyildizi, S.; Yildiz, S.; Oguzturk, H. Antioxidative effects of curcumin, beta-myrcene and 1,8-cineole against 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced oxidative stress in rats liver. Toxicol. Ind. Health 2011, 27, 447–453. [Google Scholar] [CrossRef]
- Ladan Moghadam, A.R. Antioxidant activity and chemical composition of rosmarinus officinalis l. essential oil from iran. J. Essent. Oil Bear. Plants 2015, 18, 1490–1494. [Google Scholar] [CrossRef]
- Schmidt, E.; Bail, S.; Buchbauer, G.; Stoilova, I.; Atanasova, T.; Stoyanova, A.; Krastanov, A.; Jirovetz, L. Chemical composition, olfactory evaluation and antioxidant effects of essential oil from Mentha x piperita. Nat. Prod. Commun. 2009, 4, 1107–1112. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Cillard, J.; Cillard, P.; Cormier, M. Effect of experimental factors on the prooxidant behavior of α-tocopherol. J. Am. Oil Chem. Soc. 1980, 57, 255–261. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, H.; Wang, J.; Zhou, L.; Yang, P. Chemical composition and anti-inflammatory, cytotoxic and antioxidant activities of essential oil from leaves of mentha piperita grown in china. PLoS ONE 2014, 9, e114767. [Google Scholar] [CrossRef]
- Da Silva Ramos, R.; Rodrigues, A.B.; Farias, A.L.; Simoes, R.C.; Pinheiro, M.T.; Ferreira, R.M.; Costa Barbosa, L.M.; Picanco Souto, R.N.; Fernandes, J.B.; Santos, L.D.; et al. Chemical composition and in vitro antioxidant, cytotoxic, antimicrobial, and larvicidal activities of the essential oil of Mentha piperita L. (Lamiaceae). Sci. World J. 2017, 2017, 4927214. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Ou, Y.; Chen, H.; Xiao, S.; Liu, G.; Cao, Y.; Huang, Q. Cellular antioxidant activities of polyphenols isolated from Eucalyptus leaves (Eucalyptus grandis× Eucalyptus urophylla GL9). J. Funct. Foods 2014, 7, 737–745. [Google Scholar] [CrossRef]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Aherne, S.A.; Kerry, J.P.; O’brien, N.M. Effects of plant extracts on antioxidant status and oxidant-induced stress in Caco-2 cells. Br. J. Nutr. 2007, 97, 321–328. [Google Scholar] [CrossRef] [Green Version]
- O’sullivan, A.; O’Callaghan, Y.; O’Grady, M.; Queguineur, B.; Hanniffy, D.; Troy, D.; Kerry, J.; O’Brien, N. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H. Oregano essential oil induces SOD1 and GSH expression through nrf2 activation and alleviates hydrogen peroxide-induced oxidative damage in IPEC-J2 cells. Oxidative Med. Cell. Longev. 2016, 2016, 13. [Google Scholar] [CrossRef]
- Mimica-Dukic, N.; Bozin, B. Mentha L. species (Lamiaceae) as promising sources of bioactive secondary metabolites. Curr. Pharm. Des. 2008, 14, 3141–3150. [Google Scholar] [CrossRef]
- Khalil, A.F.; Elkatry, H.O.; El Mehairy, H.F. Protective effect of peppermint and parsley leaves oils against hepatotoxicity on experimental rats. Ann. Agric. Sci. 2015, 60, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.A.; Shukitt-Hale, B.; Kalt, W.; Ingram, D.K.; Joseph, J.A.; Wolkow, C.A. Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditis elegans. Aging Cell 2006, 5, 59–68. [Google Scholar] [CrossRef]
- Shukla, V.; Yadav, D.; Phulara, S.C.; Gupta, M.M.; Saikia, S.K.; Pandey, R. Longevity-promoting effects of 4-hydroxy-E-globularinin in Caenorhabditis elegans. Free Radic. Biol. Med. 2012, 53, 1848–1856. [Google Scholar] [CrossRef]
- Kampkotter, A.; Timpel, C.; Zurawski, R.F.; Ruhl, S.; Chovolou, Y.; Proksch, P.; Watjen, W. Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 314–323. [Google Scholar] [CrossRef]
- Vanfleteren, J.R. Oxidative stress and ageing in Caenorhabditis elegans. Biochem. J. 1993, 292 Pt 2, 605–608. [Google Scholar] [CrossRef]
- Larsen, P.L. Aging and resistance to oxidative damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 1993, 90, 8905–8909. [Google Scholar] [CrossRef]
- Pandey, S.; Tiwari, S.; Kumar, A.; Niranjan, A.; Chand, J.; Lehri, A.; Chauhan, P.S. Antioxidant and anti-aging potential of Juniper berry (Juniperus communis L.) essential oil in Caenorhabditis elegans model system. Ind. Crop. Prod. 2018, 120, 113–122. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Zhou, H.-C.; Lin, Y.-M.; Wei, S.-D.; Tam, N.F.-y. Structural diversity and antioxidant activity of condensed tannins fractionated from mangosteen pericarp. Food Chem. 2011, 129, 1710–1720. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activity of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A. Characterization of the antioxidant properties of phenolic extracts from some citrus peels. J. Food Sci. Technol. 2012, 49, 729–736. [Google Scholar] [CrossRef]
- Oboh, G.; Henle, T. Antioxidant and inhibitory effects of aqueous extracts of Salvia officinalis leaves on pro-oxidant-induced lipid peroxidation in brain and liver in vitro. J. Med. Food 2009, 12, 77–84. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Dai, T.; Liu, C.; Li, T.; McClements, D.J.; Chen, J.; Liu, J. Proanthocyanidins, isolated from Choerospondias axillaris fruit peels, exhibit potent antioxidant activities in vitro and a novel anti-angiogenic property in vitro and in vivo. J. Agric. Food Chem. 2016, 64, 3546–3556. [Google Scholar] [CrossRef]
- Martorell, P.; Forment, J.V.; de Llanos, R.; Monton, F.; Llopis, S.; Gonzalez, N.; Genoves, S.; Cienfuegos, E.; Monzo, H.; Ramon, D. Use of Saccharomyces cerevisiae and Caenorhabditis elegans as model organisms to study the effect of cocoa polyphenols in the resistance to oxidative stress. J. Agric. Food Chem. 2011, 59, 2077–2085. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds 1 | Retention Time, min | Peppermint EO, % | Native Spearmint EO, % | Scotch Spearmint EO, % |
|---|---|---|---|---|
| Tricyclene | 13.23 | - 2 | 0.1 | 0.05 |
| alpha-Pinene | 14.03 | 0.56 | - | - |
| 1-Octen-3-ol | 15.8 | 0.11 | - | - |
| Sabinene | 15.91 | 0.43 | 0.48 | 0.46 |
| beta-Pinene | 16.16 | 0.81 | 0.62 | 0.59 |
| 3-Octanol | 16.82 | 0.26 | 1 | 2.68 |
| beta-Myrcene | 16.87 | 0.2 | 2.41 | 0.56 |
| alpha-Terpinene | 18.41 | 0.35 | 0.16 | - |
| para-Cymene | 18.6 | 0.32 | 0.22 | - |
| 1,8-Cineole | 19.07 | 5.62 | 2.24 | 1.32 |
| Limonene | 19.31 | 1.58 | 6.6 | 12.99 |
| cis-beta-Ocimene | 19.53 | 0.34 | 0.2 | - |
| gamma-Terpinene | 20.91 | 0.56 | 0.36 | 0.05 |
| trans-Sabinene Hydrate | 21.15 | 0.86 | 1.1 | 0.14 |
| alpha-Terpinolene | 22.81 | 0.18 | 0.12 | - |
| cis-Sabinene Hydrate | 23.04 | 0.11 | 0.16 | 0.09 |
| Linalool | 23.21 | 0.37 | 0.09 | 0.07 |
| Amyl Isovalerate | 23.83 | 0.15 | - | - |
| 3-Octanol Acetate | 24.84 | 0.05 | 0.32 | 0.13 |
| trans-Pinocarveol | 25.66 | - | 0.09 | 0.13 |
| trans-Verbenol | 26.02 | - | - | 0.11 |
| Menthone | 26.33 | 21.8 | - | 1.21 |
| Isomenthone | 26.91 | 3.75 | - | 0.16 |
| Menthofuran | 27.41 | 2.08 | - | - |
| neo-Menthol | 27.56 | 4.19 | - | - |
| 4-Terpineol | 28.25 | - | 1.09 | 0.19 |
| Menthol | 28.59 | 38.45 | 0.12 | - |
| iso-Menthol | 28.78 | 0.71 | - | - |
| cis-Dihydrocarvone | 28.88 | - | 0.99 | 0.94 |
| alpha-Terpineol | 29 | - | 0.29 | 0.2 |
| neoiso-Menthol/alpha-Terpineol | 29.21 | 0.41 | - | - |
| trans-Dihydrocarvone/Dihydrocarveol | 29.29 | - | 0.44 | 0.11 |
| neo-Dihydrocarveol | 29.52 | - | 0.26 | - |
| trans-Carveol | 31.4 | - | 0.68 | 0.53 |
| Pulegone | 31.83 | 0.91 | - | - |
| Carvone | 32.59 | - | 70.36 | 70.91 |
| Piperitone | 32.86 | 0.65 | 0.26 | 0.31 |
| Carvone Oxide (1st Isomer) | 33.25 | - | 0.07 | 0.14 |
| Carvone Oxide (2nd Isomer) | 33.97 | - | 0.16 | 0.25 |
| neo-Menthyl Acetate | 34.96 | 0.22 | - | - |
| Thymol | 35.66 | 0.1 | - | - |
| Menthyl Acetate | 36.05 | 3.9 | - | - |
| iso-Menthyl Acetate | 37.03 | 0.19 | - | - |
| Dihydrocarvyl Acetate | 38.12 | - | 0.17 | - |
| Eugenol | 39.33 | 0.05 | 0.13 | 0.11 |
| cis-Carvyl Acetate | 40.24 | - | 0.7 | 0.12 |
| cis-Jasmone | 41.71 | 0.03 | 0.44 | 0.37 |
| beta-Bourbonene | 42.73 | 0.43 | 1.69 | 1.36 |
| beta-Elemene | 43.05 | 0.23 | 0.15 | 0.08 |
| beta-Caryophyllene | 44.8 | 2.87 | 1.1 | 0.88 |
| beta-Copaene | 45.37 | 0.07 | 0.25 | 0.18 |
| trans-beta-Farnesene | 46.88 | 0.49 | 0.97 | 0.44 |
| Phenethyl Isovalerate | 47.62 | - | 0.16 | 0.23 |
| Germacrene D | 48.47 | 3.24 | 1.01 | 0.5 |
| Bicyclogermacrene | 49.33 | 0.53 | - | - |
| alpha-Muurolene | 49.52 | - | 0.48 | 0.49 |
| delta-Cadinene | 50.78 | 0.16 | 0.05 | 0.06 |
| Viridiflorol | 54.51 | 0.8 | 0.45 | - |
| Assay 1 | EC50, mg/mL | ||
|---|---|---|---|
| Peppermint EO | Native Spearmint EO | Scotch Spearmint EO | |
| DPPH scavenging capacity | 70.29 ± 4.59 c | 86.51 ± 5.45 b | 109.8 ± 6.70 a |
| TEAC assay | 29.51 ± 1.30 b | 45.74 ± 1.71 a | 44.38 ± 1.67 a |
| Reducing power assay | 22.7 ± 1.66 | 23.39 ± 2.66 | 22.91 ± 2.35 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Tan, B.; Liu, Y.; Dunn, J.; Martorell Guerola, P.; Tortajada, M.; Cao, Z.; Ji, P. Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint. Molecules 2019, 24, 2825. https://doi.org/10.3390/molecules24152825
Wu Z, Tan B, Liu Y, Dunn J, Martorell Guerola P, Tortajada M, Cao Z, Ji P. Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint. Molecules. 2019; 24(15):2825. https://doi.org/10.3390/molecules24152825
Chicago/Turabian StyleWu, Zhaohai, Bie Tan, Yanhong Liu, James Dunn, Patricia Martorell Guerola, Marta Tortajada, Zhijun Cao, and Peng Ji. 2019. "Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint" Molecules 24, no. 15: 2825. https://doi.org/10.3390/molecules24152825