Interplay between Residual Protease Activity in Commercial Lactases and the Subsequent Digestibility of ?-Casein in a Model System



 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
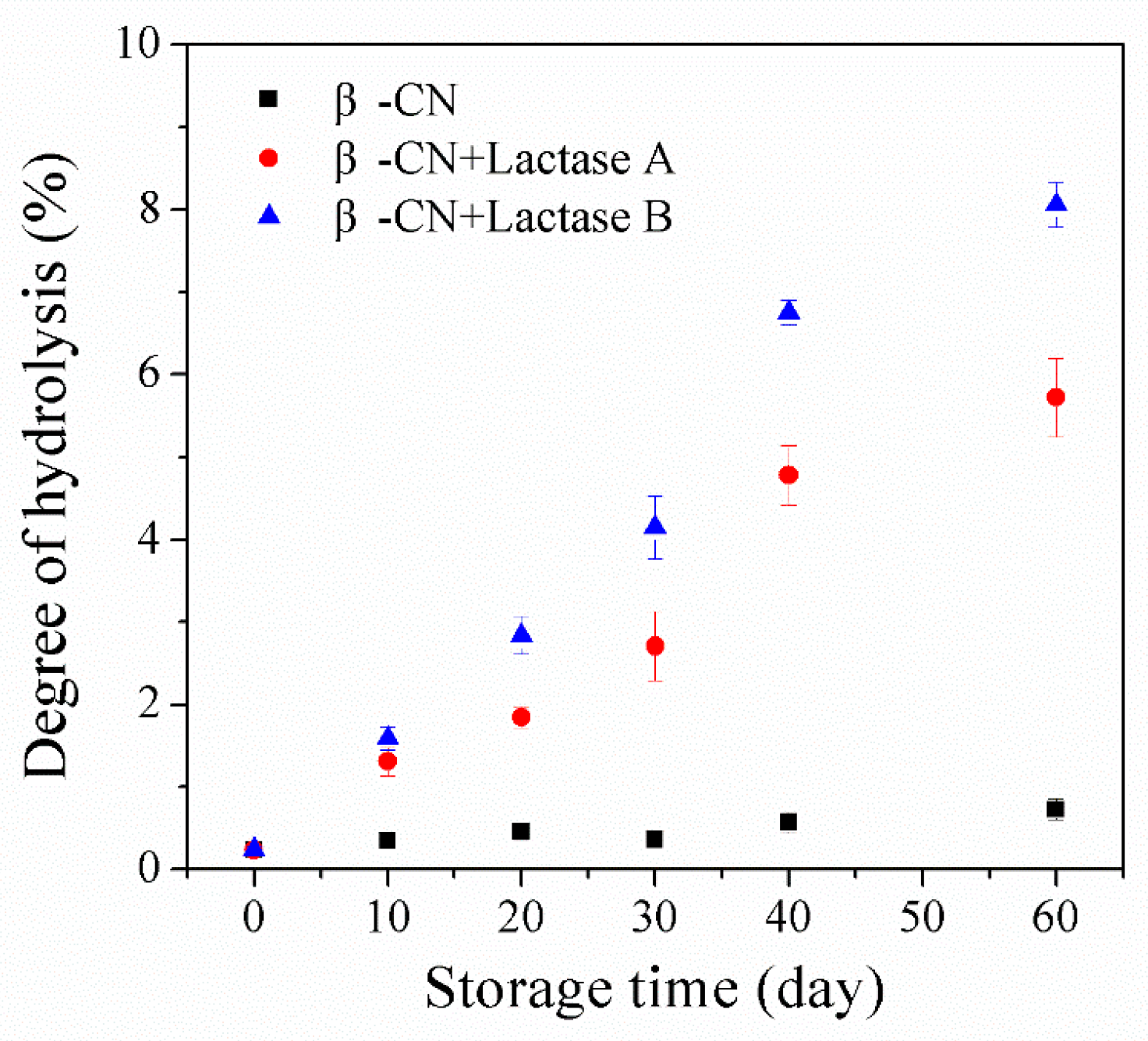
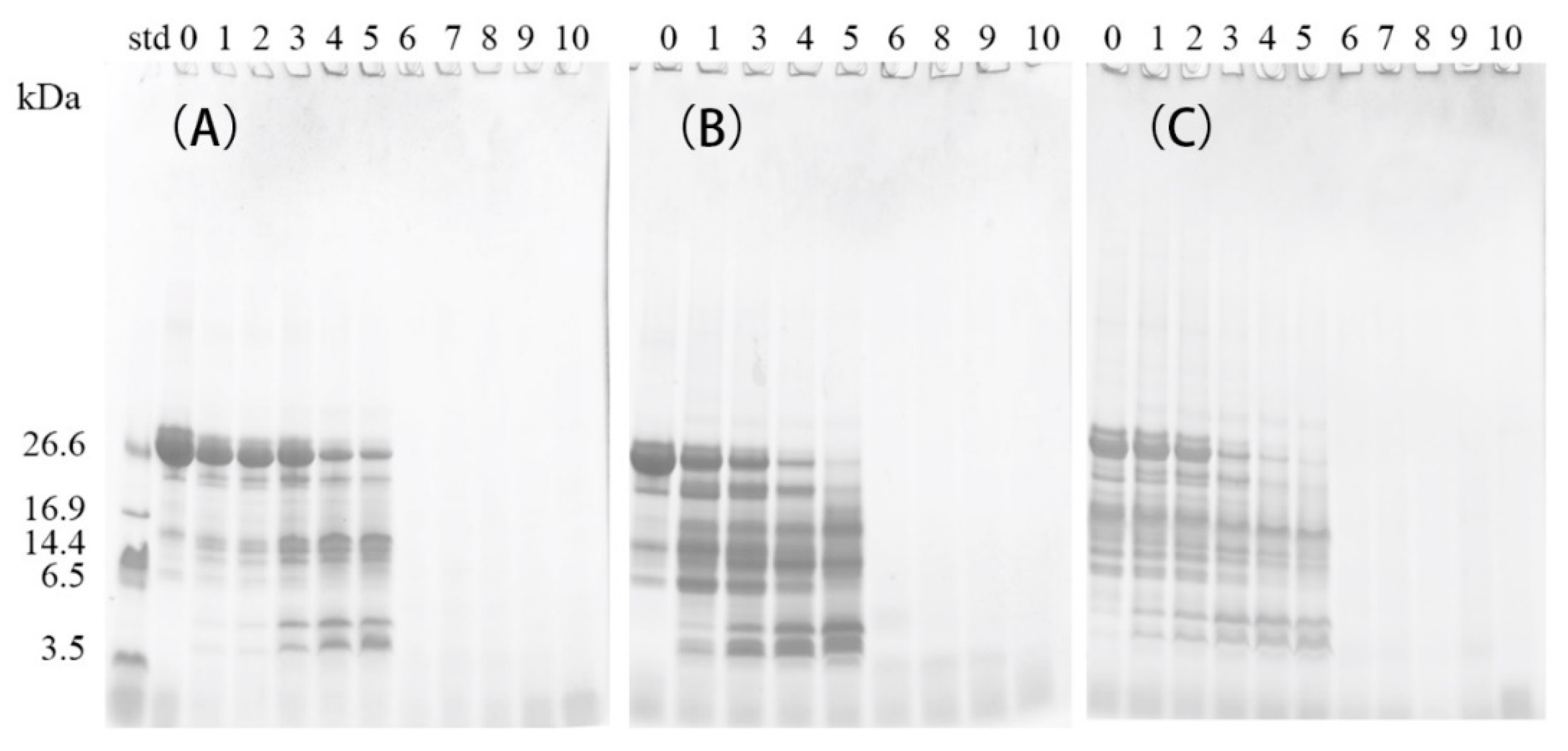
2.1. Proteolysis of β-casein (β-CN) during Storage
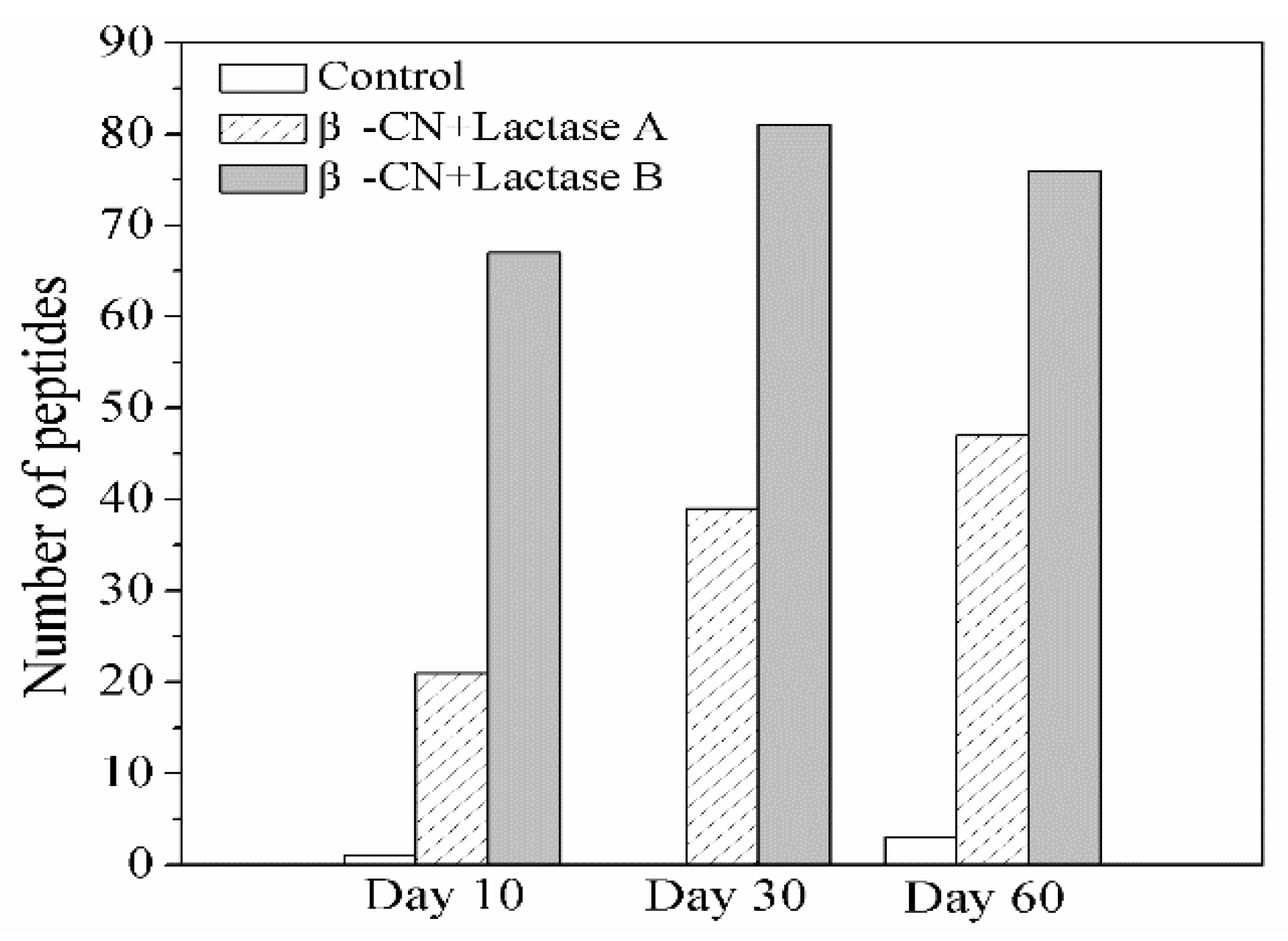
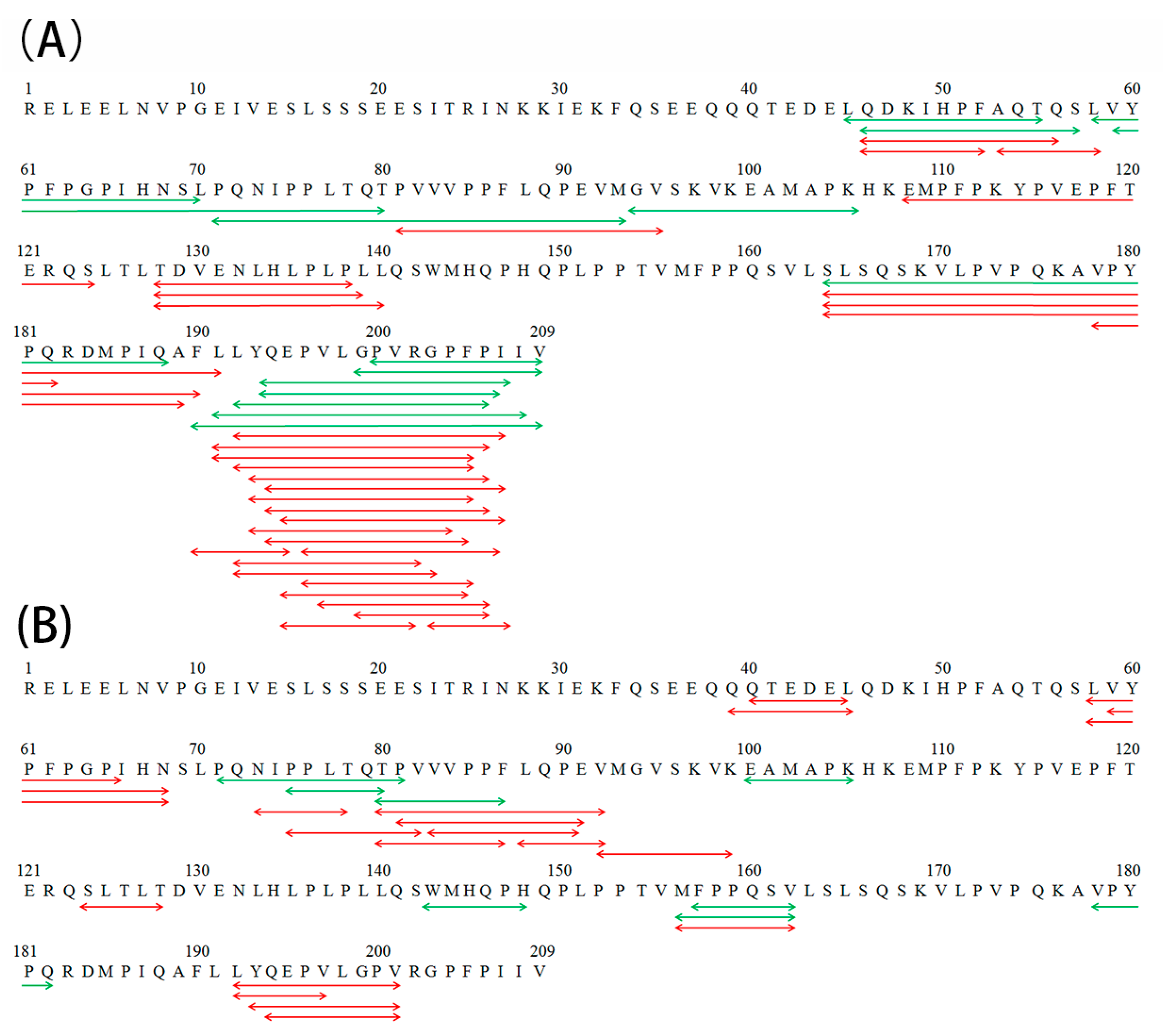
2.2. Formation of Bitter and N-Terminal Glycated Peptide
2.3. Change in Digestibility
3. Materials and Methods
3.1. Materials
3.2. Storage Experiment
3.3. In Vitro Digestion
3.4. SDS-PAGE
3.5. DH
3.6. LC-ESI-MS/MS Ion Trap
3.7. Statistics Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Itan, Y.; Jones, B.L.; Ingram, C.J.; Swallow, D.M.; Thomas, M.G. A worldwide correlation of lactase persistence phenotype and genotypes. BMC Evol. Biol. 2010, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.; Gerbault, P.; Thomas, M.G.; Burger, J. The evolution of lactase persistence in Europe. A synthesis of archaeological and genetic evidence. Int. Dairy J. 2012, 22, 88–97. [Google Scholar] [CrossRef]
- Frandsen, S.E.; Jensen, H.G.; Yu, W.; Walter-Jørgensen, A. Reform of EU sugar policy: Price cuts versus quota reductions. Eur. Rev. Agric. Econ. 2003, 30, 1–26. [Google Scholar] [CrossRef]
- Nielsen, S.D.; Jansson, T.; Le, T.T.; Jensen, S.; Eggers, N.; Rauh, V.; Sørensen, J.; Andersen, H.J.; Larsen, L.B. Correlation between sensory properties and peptides derived from hydrolysed-lactose UHT milk during storage. Int. Dairy J. 2017, 68, 23–31. [Google Scholar] [CrossRef]
- Zhao, D.; Le, T.T.; Nielsen, S.D.; Larsen, L.B. Effect of storage on lactase-treated β-casein and β-lactoglobulin with respect to bitter peptide formation and subsequent in vitro digestibility. J. Agric. Food Chem. 2017, 65, 8409–8417. [Google Scholar] [CrossRef] [PubMed]
- Harju, M.; Kallioinen, H.; Tossavainen, O. Lactose hydrolysis and other conversions in dairy products: Technological aspects. Int. Dairy J. 2012, 22, 104–109. [Google Scholar] [CrossRef]
- Nielsen, S.D.; Zhao, D.; Le, T.T.; Rauh, V.; Sørensen, J.; Andersen, H.J.; Larsen, L.B. Proteolytic side-activity of lactase preparations. Int. Dairy J. 2018, 78, 159–168. [Google Scholar] [CrossRef]
- Jansson, T.; Jensen, H.B.; Sundekilde, U.K.; Clausen, M.R.; Eggers, N.; Larsen, L.B.; Ray, C.; Andersen, H.; Bertram, H.C. Chemical and proteolysis-derived changes during long-term storage of lactose-hydrolyzed ultrahigh-temperature (UHT) milk. J. Agric. Food Chem. 2014, 62, 11270–11278. [Google Scholar] [CrossRef] [PubMed]
- Jansson, T.; Jensen, S.; Eggers, N.; Clausen, M.R.; Larsen, L.B.; Ray, C.; Sundgren, A.; Andersen, H.; Bertram, H.C. Volatile component profiles of conventional and lactose-hydrolyzed UHT milk—a dynamic headspace gas chromatography-mass spectrometry study. Dairy Sci. Technol. 2014, 94, 311–325. [Google Scholar] [CrossRef]
- Jansson, T.; Clausen, M.R.; Sundekilde, U.K.; Eggers, N.; Nyegaard, S.; Larsen, L.B.; Ray, C.; Sundgren, A.; Andersen, H.J.; Bertram, H.C. Lactose-hydrolyzed milk is more prone to chemical changes during storage than conventional ultra-high-temperature (UHT) milk. J. Agric. Food Chem. 2014, 62, 7886–7896. [Google Scholar] [CrossRef]
- Jensen, S.; Jansson, T.; Eggers, N.; Clausen, M.R.; Larsen, L.B.; Jensen, H.B.; Ray, C.; Sundgren, A.; Andersen, H.J.; Bertram, H.C. Storage-induced changes in the sensory characteristics and volatiles of conventional and lactose-hydrolyzed UHT processed milk. Eur. Food Res. Technol. 2015, 240, 1247–1257. [Google Scholar] [CrossRef]
- Carulli, S.; Calvano, C.D.; Palmisano, F.; Pischetsrieder, M. MALDI-TOF MS characterization of glycation products of whey proteins in a glucose/galactose model system and lactose-free milk. J. Agric. Food Chem. 2011, 59, 1793–1803. [Google Scholar] [CrossRef] [PubMed]
- Rauh, V.M.; Johansen, L.B.; Ipsen, R.; Paulsson, M.; Larsen, L.B.; Hammershøj, M. Plasmin activity in UHT milk: Relationship between proteolysis, age gelation, and bitterness. J. Agric. Food Chem. 2014, 62, 6852–6860. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Deeth, H.C.; Larsen, L.B. Proteomics of major bovine milk proteins: Novel insights. Int. Dairy J. 2017, 67, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Troise, A.D.; Bandini, E.; De Donno, R.; Meijer, G.; Trezzi, M.; Fogliano, V. The quality of low lactose milk is affected by the side proteolytic activity of the lactase used in the production process. Food Res. Int. 2016, 89, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledesma-Osuna, A.I.; Ramos-Clamont, G.; Vázquez-Moreno, L. Characterization of bovine serum albumin glycated with glucose, galactose and lactose. Acta Biochim. Pol. 2008, 55, 491–497. [Google Scholar] [PubMed]
- Izzo, H.V.; Ho, C.T. Peptide-specific Maillard reaction products: A new pathway for flavor chemistry. Trends Food Sci. Tech. 1992, 3, 253–257. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; He, C.; Song, H.; Chen, F. Effect of thermal treatment on the flavor generation from Maillard reaction of xylose and chicken peptide. Lwt-Food Sci. Technol. 2015, 64, 316–325. [Google Scholar] [CrossRef]
- Zhao, D.; Li, L.; Le, T.T.; Larsen, L.B.; Su, G.; Liang, Y.; Li, B. Digestibility of glyoxal-glycated β-casein and β-lactoglobulin and distribution of peptide-bound advanced glycation end products in gastrointestinal digests. J. Agric. Food Chem. 2017, 65, 5778–5788. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Le, T.T.; Larsen, L.B.; Li, L.; Qin, D.; Su, G.; Li, B. Effect of glycation derived from α-dicarbonyl compounds on the in vitro digestibility of β-casein and β-lactoglobulin: A model study with glyoxal, methylglyoxal and butanedione. Food Res. Inter. 2017, 102, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Geissler, S.; Peto, A.; Knütter, I.; Brandsch, M.; Henle, T. Transport of free and peptide-bound pyrraline at intestinal and renal epithelial cells. J. Agric. Food Chem. 2009, 57, 6474–6480. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, M.; Geissler, S.; Matthes, R.; Peto, A.; Silow, C.; Brandsch, M.; Henle, T. Transport of free and peptide-bound glycated amino acids: Synthesis, transepithelial flux at Caco-2 cell monolayers, and interaction with apical membrane transport proteins. ChemBioChem 2011, 12, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.S.; Leonil, J.; Henry, G.; Cauty, C.; Carvalho, A.F.; Bouhallab, S. Heating and glycation of beta-lactoglobulin and beta-casein: Aggregation and in vitro digestion. Food Res. Int. 2014, 55, 70–76. [Google Scholar] [CrossRef]
- Wada, Y.; Lonnerdal, B. Effects of different industrial heating processes of milk on site-specific protein modifications and their relationship to in vitro and in vivo digestibility. J. Agric. Food Chem. 2014, 62, 4175–4185. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Petrat-Melin, B.; Andersen, P.; Rasmussen, J.T.; Poulsen, N.A.; Larsen, L.B.; Young, J.F. In vitro digestion of purified β-casein variants A1, A2, B, and I: Effects on antioxidant and angiotensin-converting enzyme inhibitory capacity. J. Dairy Sci. 2015, 98, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Nielsen, S.D.; Villumsen, N.S.; Kristiansen, G.H.; Nielsen, L.R.; Nielsen, S.B.; Hammershøj, M.; Larsen, L.B. Using proteomics to characterise storage-induced aggregates in acidic whey protein isolate drinks. Int. Dairy J. 2016, 60, 39–46. [Google Scholar] [CrossRef]
- Zhao, D.; Li, L.; Le, T.T.; Larsen, L.B.; Xu, D.; Jiao, W.; Sheng, B.; Li, B.; Zhang, X. Digestibility of glycated milk proteins and the peptidomics of their in vitro digests. J. Sci. Food Agri. 2019, 99, 3069–3077. [Google Scholar] [CrossRef]
- Ney, K.H. Bitterness of peptides: Amino acid composition and chain length. Food Tast. Chem. 1979, 115, 149–173. [Google Scholar]
Sample Availability: Samples of the compounds β-CN and glucose are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Representative Cleavage Sites |
|---|---|
| β-CN + Lactase A (day 10) | F (190), I (207), I (208) |
| β-CN + Lactase A (day 30) | A (177), P (206), K (176), I (207), F (190), I (208), L (191), F (205), Y (193) |
| β-CN + Lactase A (day 60) | A (177), F (205), P (206), K (176), I (207), F (190), I (207), I (208), L (191), E (195), |
| β-CN + Lactase B (day 10) | F (190), V (170), P (206), F (205), Y (193), A (189), Q (194), L (191), L (192) |
| β-CN + Lactase B (day 30) | F (190), V (170), P (206), F (205), Y (193), A (189), Q (194), L (192), A (177), P (204) |
| β-CN + Lactase B (day 10) | I (207), F (190), P (206), I (208), Y (193), A (189), Q (194), L (192), A (177), S (164), I (207) |
| No. | Peptides | Identified Mass (Da) | Q Value | Origin |
|---|---|---|---|---|
| 1 | G(203)-V(209) | 642.37 | 2305 | β-CN + Lactase B |
| 2 | G(199)-V(209) | 1150.69 | 1869 | β-CN + Lactase B |
| 3 | E(195)-I(208) | 1489.87 | 1868 | β-CN + Lactase B |
| 4 | E(195)-I(209) | 1588.93 | 1856 | β-CN + Lactase B |
| 5 | L(171)-I(187) | 1948.06 | 1840 | β-CN + Lactase A |
| 6 | Y(193)-I(208) | 1780.99 | 1808 | β-CN + Lactase B |
| 7 | Y(193)-V(209) | 1880.06 | 1801 | β-CN + Lactase A or B |
| 8 | F(190)-P(206) | 1928.06 | 1792 | β-CN + Lactase A |
| 9 | V(178)-I(187) | 1214.61 | 1786 | β-CN + Lactase A |
| 10 | G(199)-I(207) | 938.53 | 1767 | β-CN + Lactase B |
| 11 | V(170)-Q(182) | 1434.82 | 1759 | β-CN + Lactase B |
| 12 | F(190)-F(205) | 1831.00 | 1741 | β-CN + Lactase B |
| 13 | L(191)-P(206) | 1780.99 | 1739 | β-CN + Lactase A or B |
| 14 | V(178)-R(183) | 758.41 | 1738 | β-CN + Lactase B |
| 15 | A(177)-V(209) | 3720.03 | 1737 | β-CN + Lactase B |
| 16 | Y(193)-I(207) | 1667.90 | 1734 | β-CN + Lactase B |
| 17 | Q(194)-V(209) | 1716.99 | 1730 | β-CN + Lactase B |
| 18 | D(184)-I(207) | 2696.44 | 1729 | β-CN + Lactase B |
| 19 | V(170)-I(187) | 2047.13 | 1724 | β-CN + Lactase A |
| 20 | L(171)-P(186) | 1834.98 | 1704 | β-CN + Lactase A |
| No. | N-Terminally Glycated Peptides | Identified Mass (Da) | Theoretical Mass (Da) | Origin |
|---|---|---|---|---|
| 1 | Q(194)EPVLGPVRGPFPI | 1667.90 | 1668.15 | β-CN + Lactase B |
| 2 | Q(188)AFLLYQEPVLGPVRGP | 2046.09 | 2046.37 | β-CN + Lactase A |
| 3 | Q(194)EPVLGPVRG | 1212.59 | 1212.64 | β-CN + Lactase B |
| 4 | Q(194)EPVLGPVRGP | 1309.83 | 1309.69 | β-CN + Lactase B |
| 5 | F(190)LLYQEPVLGP | 1536.60 | 1535.81 | β-CN + Lactase A or B |
| 6 | Y(193)QEPVLGPVRGPFP | 1716.35 | 1716.87 | β-CN + Lactase B |
| Sample (Day 30) | DH (%) | ||||
|---|---|---|---|---|---|
| G0 | G15 | G120 | GI15 | GI120 | |
| β-CN (control) | 0.36 ± 0.06a | 4.37 ± 0.37a | 9.97 ± 0.87a | 44.28 ± 3.63a | 58.91 ± 2.81a |
| β-CN+Lactase A | 2.71 ± 0.42b | 10.72 ± 1.83b | 13.74 ± 2.10b | 60.82 ± 4.17b | 69.19 ± 4.37b |
| β-CN+Lactase B | 4.15±0.04c | 16.57 ± 2.19c | 22.41 ± 3.04c | 67.53 ± 5.82b | 74.85 ± 5.80c |
| Samples | Peptides after G Digestion | Peptides after G+I Digestion | ||||
|---|---|---|---|---|---|---|
| Total | Missed | New | Total | Missed | New | |
| β-CN (control) | 63 | _ | _ | 53 | _ | _ |
| β-CN + Lactase A | 61 | 16 | 14 | 55 | 6 | 8 |
| β-CN + Lactase B | 80 | 15 | 32 | 62 | 10 | 19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, D.; Le, T.T.; Larsen, L.B.; Nian, Y.; Wang, C.; Li, C.; Zhou, G. Interplay between Residual Protease Activity in Commercial Lactases and the Subsequent Digestibility of ?-Casein in a Model System. Molecules 2019, 24, 2876. https://doi.org/10.3390/molecules24162876
Zhao D, Le TT, Larsen LB, Nian Y, Wang C, Li C, Zhou G. Interplay between Residual Protease Activity in Commercial Lactases and the Subsequent Digestibility of ?-Casein in a Model System. Molecules. 2019; 24(16):2876. https://doi.org/10.3390/molecules24162876
Chicago/Turabian StyleZhao, Di, Thao T. Le, Lotte Bach Larsen, Yingqun Nian, Cong Wang, Chunbao Li, and Guanghong Zhou. 2019. "Interplay between Residual Protease Activity in Commercial Lactases and the Subsequent Digestibility of ?-Casein in a Model System" Molecules 24, no. 16: 2876. https://doi.org/10.3390/molecules24162876
APA StyleZhao, D., Le, T. T., Larsen, L. B., Nian, Y., Wang, C., Li, C., & Zhou, G. (2019). Interplay between Residual Protease Activity in Commercial Lactases and the Subsequent Digestibility of ?-Casein in a Model System. Molecules, 24(16), 2876. https://doi.org/10.3390/molecules24162876