
The Versatile Role of Matrix Metalloproteinase for the Diverse Results of Fibrosis Treatment


1
Buddhist Tzu Chi Bioinnovation Center, Tzu Chi Foundation, Hualien 970, Taiwan
2
Department of Medical Research, Hualien Tzu Chi Hospital, Hualien 970, Taiwan
3
Department of Pathology, Hualien Tzu Chi Hospital & Tzu Chi University, Hualien 970, Taiwan
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(22), 4188; https://doi.org/10.3390/molecules24224188
Submission received: 2 October 2019
/
Revised: 14 November 2019
/
Accepted: 15 November 2019
/
Published: 19 November 2019
(This article belongs to the Special Issue Metalloproteins)
Abstract
:Fibrosis is a type of chronic organ failure, resulting in the excessive secretion of extracellular matrix (ECM). ECM protects wound tissue from infection and additional injury, and is gradually degraded during wound healing. For some unknown reasons, myofibroblasts (the cells that secrete ECM) do not undergo apoptosis; this is associated with the continuous secretion of ECM and reduced ECM degradation even during de novo tissue formation. Thus, matrix metalloproteinases (MMPs) are considered to be a potential target of fibrosis treatment because they are the main groups of ECM-degrading enzymes. However, MMPs participate not only in ECM degradation but also in the development of various biological processes that show the potential to treat diseases such as stroke, cardiovascular diseases, and arthritis. Therefore, treatment involving the targeting of MMPs might impede typical functions. Here, we evaluated the links between these MMP functions and possible detrimental effects of fibrosis treatment, and also considered possible approaches for further applications.
1. Introduction
Matrix metalloproteinases (MMPs) are endopeptidases with a Zn2+ ion catalytic domain [1]; they interact with multiple components of the extracellular matrix (ECM) and bioactive molecules such as receptors and cytosolic phosphatase [2,3]. Novel substrates of MMPs are still being identified, such as cytokines and growth factors [4]. The classification of MMPs is based on the substrate that they degrade, while the naming is not specific to the catalytic activity [5,6]. For example, MMP-1, also known as collagenase 1, can digest Col I, II, III, VII, VIII, X, and gelatin [7]. Subsequently, membrane-type MMPs (MT-MMPs) were discovered, which have a transmembrane domain from the extracellular to the cytosolic part of the cell [8]. There are other membrane-anchored metalloproteinases with a disintegrin domain, which belong to two new families, referred to as the ADAMs (A Disintegrin And Metalloproteinases) and ADAMTs (A Disintegrin And Metalloproteinases with Thrombospondin Motifs) [9]. The inhibitory pro-domain and the zinc-binding catalytic domain are the central features of MMPs, and domains corresponding to these are also present in ADAMs and ADAMTs, which have a cysteine-rich domain, epidermal growth factor (EGF)-like domain, and type-1 thrombospondin (TSP-1) domain [10]. These domains indicate that the key function of ADAMs is in the ectodomain shedding of membrane proteins, although some ADAMs can also degrade ECM substrates. The most intensively studied ADAM is ADAM17, which facilitates the release of the soluble form of tumor necrosis factor-α (TNF-α) from its membrane precursor. Unlike studies of the most critical biological functions of ADAMs on MMPs, there have been fewer studies on the use of ADAMs for ECM degradation. As such, this work focuses on the experimental evidence of using MMPs as targets in studies of organ fibrosis.
Given that their catalytic activity is specific to conserved collagen-like peptides, MMPs have often been linked to fibrosis and cancer metastasis [11]. The roles of MMPs in fibrogenesis are linked to an imbalance between ECM secretion and MMP degradation [12,13]; in tumor metastasis, MMPs degrade cell–cell junctions, which promotes invasiveness into adjacent tissues [14,15]. Therefore, the regulation of ECM-degrading enzymes may be a rational therapeutic target in both fibrosis and tumor metastasis [16,17]. Although most studies have shown that disruption of the activity or expression of MMPs reduced fibrosis, Giannandrea and Parks have listed the diverse treatment results for fibrosis in different types of MMPs [18]. Moreover, the contradictory roles of MMPs have been reported not only in the tumor microenvironment, but also in relation to the acquisition of properties for cancer growth and invasion [19]. Thus, cellular physiology or tissue homeostasis might change when targeting MMPs to treat organ fibrosis. For instance, MT1-MMP cleavage activates MMP2, thus maintaining its activity even in the presence of tissue inhibitors of metalloproteinases (TIMPs) and causes ECM remodeling [20]. Moreover, activated MMPs enhance EMT in epithelial cells, resulting in transformation of the cell type [21,22].
Interestingly, the expression of MMPs was elevated in the early stage of fibrosis, even before the accumulation of scar tissue, and they were reduced after the recovery stage [23]. It is believed that MMPs play an important role that could be inhibited to treat fibrosis. Notably, the results suggested a diverse therapeutic effect of MMP targeting. Here, we discuss the general and correlated functions of MMPs that might alter the treatment of fibrosis. Moreover, MMPs are also related to cancer, cardiovascular, and nervous system diseases. Based on the possible significance of MMPs for treating fibrosis, but also the uncertainty about their therapeutic potential, the possible mechanisms of action of MMPs are discussed in this review, and hypotheses are proposed about the roles of MMPs in fibrogenesis and its therapy.
2. General Functions and Regulation of MMPs
The endopeptidase activity of MMPs is derived from their catalytic domain, which is inhibited by the pro-domain (consisting of the conserved amino acid sequence PRCGXPD) [24]. Thus, the general MMP is secreted in a latent form and located depending on its domain-property; as such, the transmembrane domain-containing MT-MMPs act as membrane proteins. Some MMPs are not secreted and instead perform different functions in the cytoplasm. For example, as a regulator of cellular communication network factor 2 or connective tissue growth factor (CCN2/CTGF), MMP-3 plays a role in the nucleus, which is due to its attenuated signal peptide at the N-terminus [25]. Moreover, MMP-2 has been shown to be present in the cytosol of cardiomyocytes, due to its cleavage by troponin I, during ischemia–reperfusion injury [26,27]. However, the Human Genome Project revealed that the real gene numbers are far fewer than those that are predicted [28], which suggests that some protein-coding genes, including MMPs, may have more undiscovered functions.
2.1. The ECM Digestion Processes of MMPs
All MMPs catalyze the breaking of peptide bonds via their Zn2+-containing domain. Some of them, such as MMP-2 and MMP-9, contain a fibronectin-like region for more inseparable binding to their substrate [29,30]. The digestion process of MMP-3 was described by Pelmenschikov et al. [31,32]. Except for the catalytic domain, members of the MMP family have some motifs such as the N-terminal signal peptide that confer the optional secretory property of MMPs [33]. The hinge region contains a proline-rich region and cooperates with the hemopexin-like C-terminal domain that consists of a four-bladed β-propeller structure. This interacts with TIMP-1 and inhibits the activity of pro-MMP-9 and cell migration [34,35]. Furthermore, the hinge and hemopexin-like region form a proline zipper-like structure to unwind the triple helix of collagen [36,37] because the substrate-binding site is too narrow (about 5 Å) from the triple-helix-typed structure of collagen [38]. It is now clear that MMPs are very effective in catalyzing specific substrates like collagen and gelatin.
2.2. The Regulation of MMPs Corresponding to Physiological Processes
MMPs play major roles in cell development and migration because of their ECM-degrading activity, which is controlled by triggers such as growth factors or cytokines acting on cis-elements, including activator protein 1 (AP-1) and polyoma enhancer activator 3 (PEA3) at the promoter upstream [39]. For the wound-healing response, keratinocyte migration, angiogenesis, and contraction are correlated with MMP-13; thus, MMP-13 knockout mice exhibit impaired wound-healing response [40]. In skin diseases, epidermal growth factor (EGF)-induced MMP-1 expression in skin fibroblasts has been shown to be related to the deregulation of matrix metabolism [41]. Furthermore, studies have revealed that matrix stiffness regulates MMP-9 and TIMP-1 to perpetuate fibrosis in hepatic stellate cells, which play a pivotal role in fibrosis [42]. This suggests the intimate relationship between MMPs and fibrosis.
The enzyme activity of MMPs is regulated by a “cysteine switch” to restrict the contact between Zn2+ and H2O molecules [43]. Moreover, when the sulfide bond is broken or proteolytic cleavage occurs at the bait region, the catalytic domain is exposed and activated [44]. A well-known model of regulation is the binding of the catalytic domain by TIMPs [45]; the inhibitory process involves the 1:1 binding of MMP/TIMP on cell surfaces of cutaneous keratinocytes or fibroblasts [46]. Thus, regulation of the expression of either MMPs or TIMPs is essential for maintaining ECM balance.
2.3. The Connection of Expression Profile and Organs
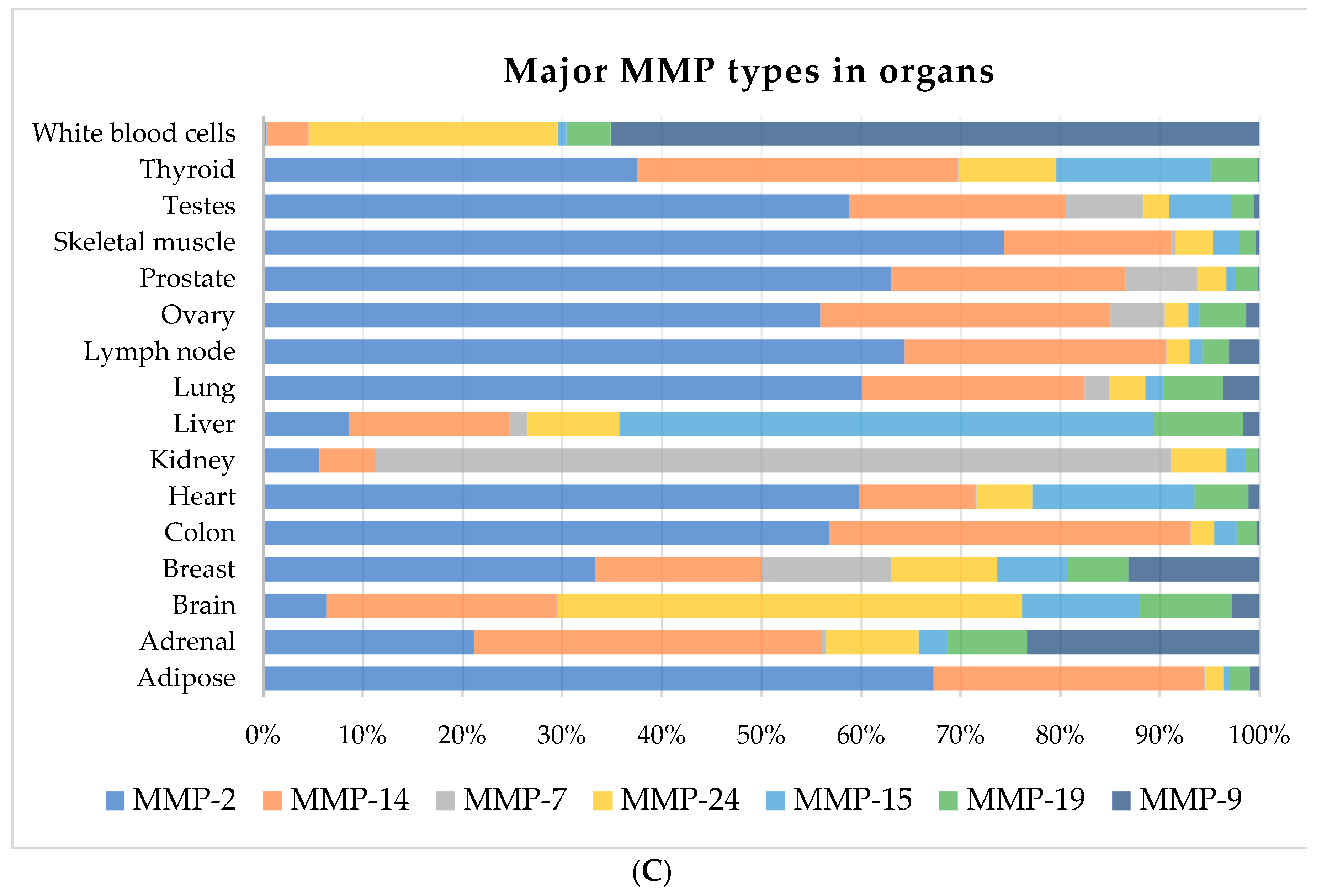
The functions of MMPs depend markedly on their localization. Since different physiological functions of various systems may need different types of MMPs, they exhibit diverse expression ratios in different organs. Thus, we compared the RPKM (reads per kilobase per million mapped reads) of 21 MMPs (MMP-1, 2, 3, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 19, 20, 21, 24, 25, 26, 27) in 16 human tissues from 19 biosamples of Expression Atlas by Illumina bodyMap2 in NCBI GENE (https://www.ncbi.nlm.nih.gov/gene). The RPKM represents the transcript reads in the same gene length arranged by their total expression rate (see Figure 1). Notably, four of 16 organs (heart, liver, lung, and kidney) are likely to suffer from fibrosis, in which 25.64% of all MMPs are expressed (Figure 1B). Furthermore, in fibrotic organs, MMP-2, MMP-14, MMP-7, MMP-24, MMP-15, MMP-19, and MMP-9 constitute more than 90% of total MMP expression, which are reported to be key regulators of tissue fibrosis [47,48,49,50,51]. As shown in Figure 1C, MMP-2 is a pivotal MMP enzyme, except for in the brain, liver, kidney, and white blood cell (WBC). Thus, targeting these major MMPs might be harmful in other organs away from the fibrosis tissue. In mouse MMP mutant strains, there are subtle differences in the phenotypes depending on the particular mutated MMP, including reduced body size [52], obesity [53], reduced hepatic fibrosis [48], delayed mammary tumorigenesis [54], and bone development defects [55]. Based on this, members of the MMP family show good compensation for deficits of other members, except that MMP-14 alone (MT1-MMP) mutant mice show lethality and die by 3–12 weeks of age [56]. Consistent with this, our data as presented in Figure 1 showed that MMP-14 was expressed in almost all listed organs, which might explain the lethality of its mutation.
3. The Role of ECM Degradation in Fibrosis Treatment
Fibrosis is a disease associated with an abnormal wound-healing response in tissues such as the skin, liver, lung, kidney, and heart. To protect the site of injury and prevent infection, fibroblasts or circulating fibrocytes migrate and proliferate to secrete ECM as well as form scar tissue. The fibroblasts are activated and transdifferentiate into myofibroblasts, resulting in excessive ECM secretion. In a normal state, these myofibroblasts undergo apoptosis once the injured tissue is repaired. However, if the regulation of myofibroblast apoptosis and ECM degradation is impaired, tissue fibrosis and damage can occur. However, the key mechanisms behind this, such as the inflammatory response, elevated cytokines, and changes in the microenvironment, are still unclear.
Given that MMPs are known to function in the degradation of ECM, it was considered that they may have potential for treating fibrosis. Specific components of the ECM, such as collagens (I, III, V, and VII), fibronectin, elastin, and proteoglycans, are present at increased levels during fibrogenesis [57]. Therefore, MMPs are upregulated in the early and late stages of fibrosis in response to ECM accumulation and correlate with the fibrotic process [58].
3.1. Rationale for MMPs in Digesting Fibers
For digesting excessive ECM and replacing fibrosing foci with normal tissue, numerous lines of evidence have revealed that MMPs such as MMP-2 and MMP-9 increase their expression to achieve wound healing in the recovery stage [59,60], at a stage generally called re-epithelialization [61]. Some researchers have considered that fibrosis might originate from the failure of re-epithelialization, with MMPs and their functions potentially playing important roles in the Drosophila basement membrane remodeling [62]. Particularly, expression of the MMP-1 gene is known to be remarkably increased in fibroblasts in hepatic fibrosis, but not in those with liver cirrhosis [63]. It has been noticed that MMP-7 activity is measured from the serum of children with cystic fibrosis [64]. Moreover, up to a 7–12-fold increase in MMP-2 gene expression was found in in CCl4-induced liver fibrosis rats, whereas the 65 kDa active form of MMP-2 was enhanced 13–28-fold in comparison to that in the control group, as revealed by a zonography assay [65]. These results proved that MMPs are directly and indirectly correlated to fibrosis, but the actual roles that they play in fibrogenesis remain unknown.
3.2. Therapeutic Potential of MMP Inhibition but Not Activation
To determine whether MMPs play key roles, the specific knockout of genes is beneficial. As a result of such a knockout, the relationships of MMPs with fibrotic diseases in the lung, skin, and kidney have started to be revealed [66,67,68]. For instance, unilateral ureteral obstruction (UUO)-induced kidney fibrosis in MMP-9 KO mice showed significantly lesser interstitial fibrosis than in wild type mice [69]. Moreover, in the 2000s, researchers started to identify the correlation of MMP/TIMP ratio and tissue remodeling [70,71], and also evaluated the therapeutic potential of each type of MMP [72]. However, almost all MMPs have highly overlapping substrates and shared functions, as described above, so the knockout of one specific MMP gene may not successfully result in loss of function. In this context, the use of tetracycline-like antibiotics, which inhibit MMPs, were previously approved for treating infection [73]. Compared with healthy human samples, in a patient with idiopathic pulmonary fibrosis (IPF), MMP-3, MMP-9, and TIMP-1 showed decreased levels in bronchoalveolar lavage fluid (BALF), but the forced vital capacity and six-minute walking distance showed no differences [74]. Although more studies have been performed in the liver and lung than in the kidney or other organs, it was concluded that the results are often diverse for each MMP type and also differ from the animal model in the systemic analysis by Giannandrea and Parks in 2014 [18]. For example, MMP-12 has both anti- and pro-fibrotic effects in pulmonary fibrosis [75,76], and also does not affect the genetic loss of MMP-9 in rodent models [77]. Rather than diminishing fibrotic scarring, MMPs were also surprisingly shown to enhance fibrogenesis more often than improve it [78]. Surprisingly, MMPs display pro-fibrotic activity in most cases, so the use of MMP inhibitors might have potential for treating fibrosis.
In the literature, findings from studies on activation or knockout animals have indicated the importance of MMPs in the development of fibrosis [78]. However, the anti-fibrotic effects of MMPs in the lungs vary from those in organs including the liver, heart, and kidney [18]. In Table 1, we present the MMP inhibitors that have been shown to be effective in fibrosis animal models, including those with lung, liver, and myocardial fibrosis. In general, broad-spectrum MMP inhibitors are effective in all three organs. According to fewer reports on selective inhibitors, their development by pharmaceutical companies might only have been reported in patents. Moreover, some studies are not shown in Table 1 since they only involved using cell models for mechanistic analyses or for treatments having potential only at the acute injury phase [79,80]. Nevertheless, the drugs shown to have potential by animal studies still need to be tested in preclinical studies before being applied to patients.
4. More Functions of MMPs
Although substantial studies have revealed the functions and physiological roles of almost all MMPs, it appears that many proteins that interact with MMPs have yet to be discovered. A major clinical use of MMPs is to prevent cancer metastasis. However, studies have also utilized MMPs as targets in different body systems, including the immune, cardiovascular, and central nervous systems. Therefore, the unexpected results for fibrosis treatment may be because these additional targets also interact in fibrogenesis, such as in immunity and cell growth, and transduce messages [91,92].
4.1. Promotion of Cancer Invasiveness
The process of tumor cell spread from the primary site into other normal tissue, referred to as metastasis, occurs in invasive cancer via movement through the bloodstream. The surrounding tissue border and the vessel walls have to split, and typically MMPs secreted on the cell surface break the basal membrane, allowing escape into the blood flow [93,94]. Further studies showed that when the space and nutrient support is no longer sufficient for tumor growth, MMPs indirectly promote invasiveness [95].
4.2. Macrophage Degradation of the Basal Membrane
Macrophages play important roles in the immune system via their ability to perform phagocytosis and amoeboid movement toward wound tissue that has become infected. The activation of macrophages leads them to secrete MMPs, which is required for degradation of the ECM of the basement membrane, followed by migration into injured tissues [96,97] and the engulfment of pathogens. Macrophages from MMP-12 deficient mice exhibit abrogated migration ability [98]. In experimental studies, it was shown that the MMP-2 and MMP-9 inhibitors ARP100 and AG-L-66085 significantly reduced migration via different mechanisms: ARP100 inhibited MMP-2 and subsequent transforming growth factor β1 (TGF-β1) secretion, whereas AG-L-66085 diminished the angiogenesis response by reducing vascular epithelial growth factor (VEGF) in a retinoblastoma model [99]. A clinical trial for recurrent glioblastoma is going to test the combination of monoclonal antibody of MMP-9 combined with bevacizumab (NCT03631836); however, the concept of blocking MMP-9 is related to tumor vascularization but not to its ECM-degrading role [100].
4.3. MMPs Treat Stroke or Cardiovascular Diseases
MMPs also have the potential to treat diseases such as stroke, cardiovascular diseases, and arthritis [25,101,102,103]. In leukocytes, MMP-2 expression is positively correlated with the formation of sclerotic plaques, but the mechanism behind this are still unknown [104]. The above diseases are not related to degradation of the ECM, but functional components could be involved, such as NF-κB and hypoxia-inducible factor 1α (HIF-1α) [105,106]. An increasing number of additional catalytic targets of MMPs in various tissue types have also been found recently [107].
4.4. Central Nervous System (CNS) and the Microenvironment
Obstruction of the breakdown of extracellular constituents can cause CNS disease such as multiple sclerosis (MS); several types of MMPs are reported to be involved in MS [108]. In a Theiler’s murine encephalomyelitis (TME)-induced MS disease model, MMP-12 was shown to play a pivotal role in the development of astrogliosis and demyelination [109]. These processes are similar to fibrosis, including a collagenous region and focal proliferation in brain tissue. MMP-12 was also shown to reduce activated microglia and reactive macrophages, influencing the M1/M2 balance in virus-infected mice [110]. The therapeutic mechanisms include the macrophage-mediated proteolysis and matrix invasion and the basement membrane penetration potential of macrophages [111].
Recently, it is reported that MMPs regulate the microenvironment via the shedding of the exosome [112]. The exosome is a small releasing vesicle (30–100 nm in diameter), the trafficking of which enables communication with other cells and the movement of cargo such as proteins, cytokines, and miRNAs [113]. It is closely related to the physiological and regulatory mechanisms of the exosome; thus, MMP shedding for appropriate release plays a role in mediating their functions. As such, adipocyte-derived exosomes of the liver were shown to induce TGF-β signaling in hepatocytes, leading to the initiation of fibrosis [114]. Moreover, exosomes containing MT1-MMP activated pro-MMP-2 and caused the subsequent degradation of type 1 collagen and gelatin in the fibroblast-like cell line COS-1 [115]. Furthermore, the enzymatic activity of MMP-2 is involved in exosome trafficking from fibroblasts to endothelial cells and facilitates the breakdown of ECM in an MMP-14-dependent manner [116].
5. Possible Participating Role of MMPs in Fibrosis
In terms of the possible mechanisms of MMP involvement in fibrosis, the first candidate is the inflammatory response after injury [117,118], which was utilized as one of the clinical drugs for treating lung and liver fibrosis [119,120]. Anti-inflammatory therapy has been proven to have potential in animal models; however, in a clinical trial, treatment with steroids such as prednisolone in patients with pulmonary fibrosis resulted in unfavorable outcomes and increased hospitalization events [121]. Some researchers have been shifting their focus away from immunology and fibrosis, but the pathological relationship remains [122]. Notably, the correlation between fibrosis and MMPs in macrophages and neutrophils [123], such as MMP-1, MMP3, MMP-7, MMP-9, MMP-13, and MMP-19, was shown to be increased in BALF. These infiltrated macrophages and neutrophils refer to the inflammatory response and are essential in pulmonary fibrosis [50,71,124,125,126,127].
5.1. Immunomodulation or Inflammatory Regulation
Additionally, MMP-10 is known to be related to the transition from the M1 to M2 type of alveolar macrophages and regulates the immune tolerance to TLR-7 induced inflammation [128]. Besides the inflammatory effect, the following wound healing response usually includes peripheral epithelium activation through the EMT in renal tubular epithelial, hepatic stellate, and alveolar epithelial cells [129,130,131,132]; these cells transdifferentiate into fibroblast-like or myofibroblast cells and secrete collagen fibers, leading to fibrosis [133,134]. Accordingly, MMPs contribute to EMT-related cancer metastasis in breast and gastric cancers [21,135], although we mentioned their anti-cancer potential in the previous section. In the final stage of wound healing, the recruited myofibroblasts undergo apoptosis and allow tissue regeneration, whereas in fibrosis, the myofibroblasts are resistant to apoptosis and proliferate to form fibrotic foci [136].
The pro-apoptotic role of MMP-7 is due to the cleavage of CD95 in apoptosis-resistant tumor cells [137]. Moreover, MMP-10 promotes tumor progression by stimulating HIF-1α and MMP-2 in cervical tumors [138]. Notably, MMP is needed and would be increased in tissues in cases of fibrosis; however, the ECM does not decrease in these circumstances. Therefore, MMPs are strongly suspected to interact with pro-fibrotic factors, such as molecules involved in inflammation, EMT, and apoptosis resistance (e.g., TGF-β1, IL-1β, and TNF-α) [139,140].
5.2. ECM and Vasculature in Angiogenesis
Vessel walls have an ECM-containing, three-layered structure, including the tunica intima, tunica media, and tunica adventitia, and provide mechanical strength and elasticity. Neoplastic proliferation requires angiogenesis in malignant or benign tumors; the existence of MMPs and VEGF facilitates tumor growth [141]. Moreover, neutrophilic MMP-9 acts as a pro-angiogenic proteinase, as revealed using a developing chicken embryo model [142]. Studies in the adventitia layer revealed that MMP-2 activation and fibroblast proliferation both induce a phenotypic switch in a hypoxic state, converting fibroblasts into myofibroblasts, which often form a fibrotic focus [143].
6. Conclusions
Since the complete mechanisms of fibrosis remain a mystery, the origin and each stage are a complex and mutual effect. To decrease the excessive ECM in fibrotic tissue, the activated MMPs interact with the molecules involved inflammation, EMT, and apoptosis [144]. Unfortunately, the complete functions and substrates of MMPs have not yet been revealed [145,146], although some of them are secondary messengers, such as the cleavage of the AMP-activated protein kinase-α (AMPK-α) by MMP-9 in Toll-like receptor 4 (TLR4) signaling [105]. Although studies have suggested that simple inhibition of the expression of an MMP is insufficient to treat fibrosis, we aim to clarify the roles of MMPs in fibrosis in more detail to increase the potential for using them in a clinical context in the future.
Author Contributions
Writing—original Draft Preparation, H.-M.C.; Writing—review, Y.-S.C.; Conceptualization, H.-J.H.
Funding
This study was funded by the Buddhist Tzu Chi Bioinnovation Center, Tzu Chi Foundation, Hualien, Taiwan (project title: Development of a small molecule regulates transcription activity of SOX2 in type I collagen synthesis in fibroblasts for treating pulmonary fibrosis, MF00A130SS01) and Ministry of Science and Technology, Taiwan (MOST 106-2320-B-303-001-MY3 and MOST 106-2320-B-303-002-MY3).
Acknowledgments
We are grateful to Tina Emily Shih and Enago (www.enago.tw) for English proofreading.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
| Abbreviation | Definition |
| ECM | extracellular matrix |
| MMPs | matrix metalloproteinases |
| MT-MMPs | membrane-type MMPs |
| ADAMs | a disintegrin and metalloproteinases |
| ADAMTs | a disintegrin and metalloproteinases with thrombospondin motifs |
| EGF | epidermal growth factor |
| TSP-1 | type-1 thrombospondin |
| TNF-α | tumor necrosis factor-α |
| TIMPs | tissue inhibitors of metalloproteinases |
| TIMPs | unilateral ureteral obstruction |
| VEGF | vascular epithelial growth factor |
| TLR4 | Toll-like receptor 4 |
| AMPK-α | AMP-activated protein kinase α |
| CCN2/CTGF | cellular communication network factor 2 or connective tissue growth factor |
| AP-1 | activator protein 1 |
| PEA3 | polyoma enhancer activator 3 |
| EGF | epidermal growth factor |
| WBC | white blood cell |
| TGF -β1 | transforming growth factor β1 |
| HIF-1α | hypoxia-inducible factor 1α |
References
- Huntley, G.W. Synaptic circuit remodelling by matrix metalloproteinases in health and disease. Nat. Rev. Neurosci. 2012, 13, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed]
- Lopez-Otin, C.; Hunter, T. The regulatory crosstalk between kinases and proteases in cancer. Nat. Rev. Cancer 2010, 10, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Urbach, C.; Gordon, N.C.; Strickland, I.; Lowne, D.; Joberty-Candotti, C.; May, R.; Herath, A.; Hijnen, D.; Thijs, J.L.; Bruijnzeel-Koomen, C.A.; et al. Combinatorial Screening Identifies Novel Promiscuous Matrix Metalloproteinase Activities that Lead to Inhibition of the Therapeutic Target IL-13. Chem. Biol. 2015, 22, 1442–1452. [Google Scholar] [CrossRef]
- Gross, J.; Lapiere, C.M. Collagenolytic activity in amphibian tissues: A tissue culture assay. Proc. Natl. Acad. Sci. USA 1962, 48, 1014–1022. [Google Scholar] [CrossRef]
- Jablonska-Trypuc, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef]
- Pardo, A.; Selman, M. MMP-1: The elder of the family. Int. J. Biochem. Cell Biol. 2005, 37, 283–288. [Google Scholar] [CrossRef]
- Ra, H.J.; Parks, W.C. Control of matrix metalloproteinase catalytic activity. Matrix Biol. 2007, 26, 587–596. [Google Scholar] [CrossRef]
- Klein, T.; Bischoff, R. Active metalloproteases of the A Disintegrin and Metalloprotease (ADAM) family: Biological function and structure. J. Proteome Res. 2011, 10, 17–33. [Google Scholar] [CrossRef]
- Kelwick, R.; Desanlis, I.; Wheeler, G.N.; Edwards, D.R. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombospondin motifs) family. Genome Biol. 2015, 16, 113. [Google Scholar] [CrossRef]
- Fingleton, B. MMPs as therapeutic targets—Still a viable option? Semin. Cell Dev. Biol. 2008, 19, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Manoury, B.; Nenan, S.; Guenon, I.; Lagente, V.; Boichot, E. Influence of early neutrophil depletion on MMPs/TIMP-1 balance in bleomycin-induced lung fibrosis. Int. Immunopharmacol. 2007, 7, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.C.; Markle, L.S.; Vickers, J.L.; Petitt, M.S.; Raimer, S.S.; McNeese, C. The imbalanced expression of matrix metalloproteinases in nephrogenic systemic fibrosis. J. Am. Acad. Dermatol. 2010, 63, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Bramhall, S.R.; Neoptolemos, J.P.; Stamp, G.W.; Lemoine, N.R. Imbalance of expression of matrix metalloproteinases (MMPs) and tissue inhibitors of the matrix metalloproteinases (TIMPs) in human pancreatic carcinoma. J. Pathol. 1997, 182, 347–355. [Google Scholar] [CrossRef]
- Shin, D.Y.; Kim, G.Y.; Kim, J.I.; Yoon, M.K.; Kwon, T.K.; Lee, S.J.; Choi, Y.W.; Kang, H.S.; Yoo, Y.H.; Choi, Y.H. Anti-invasive activity of diallyl disulfide through tightening of tight junctions and inhibition of matrix metalloproteinase activities in LNCaP prostate cancer cells. Toxicol. In Vitro 2010, 24, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.P.; Shah, S.V.; Shukla, S.N.; Shah, P.M.; Patel, P.S. Clinical significance of MMP-2 and MMP-9 in patients with oral cancer. Head Neck 2007, 29, 564–572. [Google Scholar] [CrossRef]
- Sutnar, A.; Pesta, M.; Liska, V.; Treska, V.; Skalicky, T.; Kormunda, S.; Topolcan, O.; Cerny, R.; Holubec, L., Jr. Clinical relevance of the expression of mRNA of MMP-7, MMP-9, TIMP-1, TIMP-2 and CEA tissue samples from colorectal liver metastases. Tumour Biol. 2007, 28, 247–252. [Google Scholar] [CrossRef]
- Giannandrea, M.; Parks, W.C. Diverse functions of matrix metalloproteinases during fibrosis. Dis. Model. Mech. 2014, 7, 193–203. [Google Scholar] [CrossRef]
- Noel, A.; Gutierrez-Fernandez, A.; Sounni, N.E.; Behrendt, N.; Maquoi, E.; Lund, I.K.; Cal, S.; Hoyer-Hansen, G.; Lopez-Otin, C. New and paradoxical roles of matrix metalloproteinases in the tumor microenvironment. Front. Pharmacol. 2012, 3, 140. [Google Scholar] [CrossRef]
- Nishida, Y.; Miyamori, H.; Thompson, E.W.; Takino, T.; Endo, Y.; Sato, H. Activation of matrix metalloproteinase-2 (MMP-2) by membrane type 1 matrix metalloproteinase through an artificial receptor for proMMP-2 generates active MMP-2. Cancer Res. 2008, 68, 9096–9104. [Google Scholar] [CrossRef]
- Radisky, E.S.; Radisky, D.C. Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J. Mammary Gland. Biol. Neoplasia 2010, 15, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.C.; Pastar, I.; Ojeh, N.; Chen, V.; Liu, S.; Garzon, K.I.; Tomic-Canic, M. Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. 2016, 365, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Hemmann, S.; Graf, J.; Roderfeld, M.; Roeb, E. Expression of MMPs and TIMPs in liver fibrosis—A systematic review with special emphasis on anti-fibrotic strategies. J. Hepatol. 2007, 46, 955–975. [Google Scholar] [CrossRef] [PubMed]
- Park, A.J.; Matrisian, L.M.; Kells, A.F.; Pearson, R.; Yuan, Z.Y.; Navre, M. Mutational analysis of the transin (rat stromelysin) autoinhibitor region demonstrates a role for residues surrounding the “cysteine switch”. J. Biol. Chem. 1991, 266, 1584–1590. [Google Scholar]
- Eguchi, T.; Kubota, S.; Kawata, K.; Mukudai, Y.; Uehara, J.; Ohgawara, T.; Ibaragi, S.; Sasaki, A.; Kuboki, T.; Takigawa, M. Novel transcription-factor-like function of human matrix metalloproteinase 3 regulating the CTGF/CCN2 gene. Mol. Cell. Biol. 2008, 28, 2391–2413. [Google Scholar] [CrossRef]
- Ali, M.A.; Chow, A.K.; Kandasamy, A.D.; Fan, X.; West, L.J.; Crawford, B.D.; Simmen, T.; Schulz, R. Mechanisms of cytosolic targeting of matrix metalloproteinase-2. J. Cell. Physiol. 2012, 227, 3397–3404. [Google Scholar] [CrossRef]
- Cheng, Z.; Liu, L.; Wang, Z.; Cai, Y.; Xu, Q.; Chen, P. Hypoxia Activates Src and Promotes Endocytosis Which Decreases MMP-2 Activity and Aggravates Renal Interstitial Fibrosis. Int. J. Mol. Sci. 2018, 19, 581. [Google Scholar] [CrossRef]
- Venter, J.C.; Adams, M.D.; Myers, E.W.; Li, P.W.; Mural, R.J.; Sutton, G.G.; Smith, H.O.; Yandell, M.; Evans, C.A.; Holt, R.A.; et al. The sequence of the human genome. Science 2001, 291, 1304–1351. [Google Scholar] [CrossRef]
- Fischer, T.; Riedl, R. Inhibitory Antibodies Designed for Matrix Metalloproteinase Modulation. Molecules 2019, 24, 2265. [Google Scholar] [CrossRef]
- Peralta, F.A.; Huidobro-Toro, J.P. Zinc as Allosteric Ion Channel Modulator: Ionotropic Receptors as Metalloproteins. Int. J. Mol. Sci. 2016, 17, 1059. [Google Scholar] [CrossRef]
- Pelmenschikov, V.; Siegbahn, P.E. Catalytic mechanism of matrix metalloproteinases: Two-layered ONIOM study. Inorg. Chem. 2002, 41, 5659–5666. [Google Scholar] [CrossRef] [PubMed]
- Manzetti, S.; McCulloch, D.R.; Herington, A.C.; van der Spoel, D. Modeling of enzyme-substrate complexes for the metalloproteases MMP-3, ADAM-9 and ADAM-10. J. Comput. Aided Mol. Des. 2003, 17, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Massova, I.; Kotra, L.P.; Fridman, R.; Mobashery, S. Matrix metalloproteinases: Structures, evolution, and diversification. FASEB J. 1998, 12, 1075–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, S.J.; Pereira, H.M.; Jacchieri, S.; Brentani, R.R. Collagen/collagenase interaction: Does the enzyme mimic the conformation of its own substrate? FASEB J. 1996, 10, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Dufour, A.; Sampson, N.S.; Zucker, S.; Cao, J. Role of the hemopexin domain of matrix metalloproteinases in cell migration. J. Cell. Physiol. 2008, 217, 643–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, W. A helping hand for collagenases: The haemopexin-like domain. Structure 1995, 3, 527–530. [Google Scholar] [CrossRef]
- Nagase, H. Substrate Specificity of MMPs. In Matrix Metalloproteinase Inhibitors in Cancer Therapy; Clendeninn, N.J., Appelt, K., Eds.; Humana Press: Totowa, NJ, USA, 2001; pp. 39–66. [Google Scholar]
- Gomis-Ruth, F.X.; Gohlke, U.; Betz, M.; Knauper, V.; Murphy, G.; Lopez-Otin, C.; Bode, W. The helping hand of collagenase-3 (MMP-13): 2.7 A crystal structure of its C-terminal haemopexin-like domain. J. Mol. Biol. 1996, 264, 556–566. [Google Scholar] [CrossRef]
- Yan, C.; Boyd, D.D. Regulation of matrix metalloproteinase gene expression. J. Cell. Physiol. 2007, 211, 19–26. [Google Scholar] [CrossRef]
- Hattori, N.; Mochizuki, S.; Kishi, K.; Nakajima, T.; Takaishi, H.; D’Armiento, J.; Okada, Y. MMP-13 plays a role in keratinocyte migration, angiogenesis, and contraction in mouse skin wound healing. Am. J. Pathol. 2009, 175, 533–546. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Chung, J.H. Epidermal growth factor-induced matrix metalloproteinase-1 expression is negatively regulated by p38 MAPK in human skin fibroblasts. J. Dermatol. Sci. 2011, 64, 134–141. [Google Scholar] [CrossRef]
- Lachowski, D.; Cortes, E.; Rice, A.; Pinato, D.; Rombouts, K.; Hernandez, A.D.R. Matrix stiffness modulates the activity of MMP-9 and TIMP-1 in hepatic stellate cells to perpetuate fibrosis. Sci. Rep. 2019, 9, 7299. [Google Scholar] [CrossRef] [PubMed]
- Van Wart, H.E.; Birkedal-Hansen, H. The cysteine switch: A principle of regulation of metalloproteinase activity with potential applicability to the entire matrix metalloproteinase gene family. Proc. Natl. Acad. Sci. USA 1990, 87, 5578–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, D.E.; Alonso, D.F.; Yoshiji, H.; Thorgeirsson, U.P. Tissue inhibitors of metalloproteinases: Structure, regulation and biological functions. Eur. J. Cell Biol. 1997, 74, 111–122. [Google Scholar] [PubMed]
- Tandara, A.A.; Mustoe, T.A. MMP- and TIMP-secretion by human cutaneous keratinocytes and fibroblasts—Impact of coculture and hydration. J. Plast. Reconstr. Aesthet. Surg. 2011, 64, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Nkyimbeng, T.; Ruppert, C.; Shiomi, T.; Dahal, B.; Lang, G.; Seeger, W.; Okada, Y.; D’Armiento, J.; Gunther, A. Pivotal role of matrix metalloproteinase 13 in extracellular matrix turnover in idiopathic pulmonary fibrosis. PLoS ONE 2013, 8, e73279. [Google Scholar] [CrossRef]
- Uchinami, H.; Seki, E.; Brenner, D.A.; D’Armiento, J. Loss of MMP 13 attenuates murine hepatic injury and fibrosis during cholestasis. Hepatology 2006, 44, 420–429. [Google Scholar] [CrossRef]
- Pardo, A.; Cabrera, S.; Maldonado, M.; Selman, M. Role of matrix metalloproteinases in the pathogenesis of idiopathic pulmonary fibrosis. Respir. Res. 2016, 17, 23. [Google Scholar] [CrossRef] [Green Version]
- Rosas, I.O.; Richards, T.J.; Konishi, K.; Zhang, Y.; Gibson, K.; Lokshin, A.E.; Lindell, K.O.; Cisneros, J.; Macdonald, S.D.; Pardo, A.; et al. MMP1 and MMP7 as potential peripheral blood biomarkers in idiopathic pulmonary fibrosis. PLoS Med. 2008, 5, e93. [Google Scholar] [CrossRef] [Green Version]
- Sokai, A.; Handa, T.; Tanizawa, K.; Oga, T.; Uno, K.; Tsuruyama, T.; Kubo, T.; Ikezoe, K.; Nakatsuka, Y.; Tanimura, K.; et al. Matrix metalloproteinase-10: A novel biomarker for idiopathic pulmonary fibrosis. Respir. Res. 2015, 16, 120. [Google Scholar] [CrossRef] [Green Version]
- Itoh, T.; Ikeda, T.; Gomi, H.; Nakao, S.; Suzuki, T.; Itohara, S. Unaltered secretion of beta-amyloid precursor protein in gelatinase A (matrix metalloproteinase 2)-deficient mice. J. Biol. Chem. 1997, 272, 22389–22392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendas, A.M.; Folgueras, A.R.; Llano, E.; Caterina, J.; Frerard, F.; Rodriguez, F.; Astudillo, A.; Noel, A.; Birkedal-Hansen, H.; Lopez-Otin, C. Diet-induced obesity and reduced skin cancer susceptibility in matrix metalloproteinase 19-deficient mice. Mol. Cell. Biol. 2004, 24, 5304–5313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, R.; Lefebvre, O.; Noel, A.; Fahime, M.E.; Chenard, M.P.; Wendling, C.; Kebers, F.; LeMeur, M.; Dierich, A.; Foidart, J.M.; et al. In vivo evidence that the stromelysin-3 metalloproteinase contributes in a paracrine manner to epithelial cell malignancy. J. Cell Biol. 1998, 140, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.H.; Shipley, J.M.; Bergers, G.; Berger, J.E.; Helms, J.A.; Hanahan, D.; Shapiro, S.D.; Senior, R.M.; Werb, Z. MMP-9/gelatinase B is a key regulator of growth plate angiogenesis and apoptosis of hypertrophic chondrocytes. Cell 1998, 93, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Pardo, A.; Selman, M. Matrix metalloproteases in aberrant fibrotic tissue remodeling. Proc. Am. Thorac. Soc. 2006, 3, 383–388. [Google Scholar] [CrossRef]
- Kim, J.Y.; Choeng, H.C.; Ahn, C.; Cho, S.H. Early and late changes of MMP-2 and MMP-9 in bleomycin-induced pulmonary fibrosis. Yonsei Med. J. 2009, 50, 68–77. [Google Scholar] [CrossRef]
- Corbel, M.; Boichot, E.; Lagente, V. Role of gelatinases MMP-2 and MMP-9 in tissue remodeling following acute lung injury. Braz. J. Med. Biol. Res. 2000, 33, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Pardo, A.; Barrios, R.; Maldonado, V.; Melendez, J.; Perez, J.; Ruiz, V.; Segura-Valdez, L.; Sznajder, J.I.; Selman, M. Gelatinases A and B are up-regulated in rat lungs by subacute hyperoxia: Pathogenetic implications. Am. J. Pathol. 1998, 153, 833–844. [Google Scholar] [CrossRef]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef]
- Stevens, L.J.; Page-McCaw, A. A secreted MMP is required for reepithelialization during wound healing. Mol. Biol. Cell 2012, 23, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, I.; Arai, M.; Wada, N.; Maruyama, K. Gene expression of MMPs and TIMPs in the process of hepatic fibrosis. Nihon Rinsho 1993, 51, 428–434. [Google Scholar] [PubMed]
- Laszlo, A.; Sohar, I.; Gyurkovits, K. Activity of the lysosomal cysteine proteinases (cathepsin B,H,L) and a metalloproteinase (MMP-7-ase) in the serum of cystic fibrosis homozygous children. Acta Paediatr. Hung. 1987, 28, 175–178. [Google Scholar] [PubMed]
- Takahara, T.; Furui, K.; Funaki, J.; Nakayama, Y.; Itoh, H.; Miyabayashi, C.; Sato, H.; Seiki, M.; Ooshima, A.; Watanabe, A. Increased expression of matrix metalloproteinase-II in experimental liver fibrosis in rats. Hepatology 1995, 21, 787–795. [Google Scholar] [CrossRef]
- Gonzalez-Avila, G.; Iturria, C.; Vadillo-Ortega, F.; Ovalle, C.; Montano, M. Changes in matrix metalloproteinases during the evolution of interstitial renal fibrosis in a rat experimental model. Pathobiology 1998, 66, 196–204. [Google Scholar] [CrossRef]
- Hayashi, T.; Stetler-Stevenson, W.G.; Fleming, M.V.; Fishback, N.; Koss, M.N.; Liotta, L.A.; Ferrans, V.J.; Travis, W.D. Immunohistochemical study of metalloproteinases and their tissue inhibitors in the lungs of patients with diffuse alveolar damage and idiopathic pulmonary fibrosis. Am. J. Pathol. 1996, 149, 1241–1256. [Google Scholar]
- Lafuma, C.; El Nabout, R.A.; Crechet, F.; Hovnanian, A.; Martin, M. Expression of 72-kDa gelatinase (MMP-2), collagenase (MMP-1), and tissue metalloproteinase inhibitor (TIMP) in primary pig skin fibroblast cultures derived from radiation-induced skin fibrosis. J. Investig. Dermatol. 1994, 102, 945–950. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Qiao, X.; Tan, T.K.; Zhao, H.; Zhang, Y.; Liu, L.; Zhang, J.; Wang, L.; Cao, Q.; Wang, Y.; et al. Matrix metalloproteinase 9-dependent Notch signaling contributes to kidney fibrosis through peritubular endothelial-mesenchymal transition. Nephrol. Dial. Transplant. 2017, 32, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Arthur, M.J. Fibrogenesis II. Metalloproteinases and their inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G245–G249. [Google Scholar] [CrossRef]
- Henry, M.T.; McMahon, K.; Mackarel, A.J.; Prikk, K.; Sorsa, T.; Maisi, P.; Sepper, R.; Fitzgerald, M.X.; O’Connor, C.M. Matrix metalloproteinases and tissue inhibitor of metalloproteinase-1 in sarcoidosis and IPF. Eur. Respir. J. 2002, 20, 1220–1227. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Hovell, C.J.; Pawley, S.; Hutchings, M.I.; Arthur, M.J.; Iredale, J.P.; Benyon, R.C. Expression of matrix metalloproteinase-2 and -14 persists during early resolution of experimental liver fibrosis and might contribute to fibrolysis. Liver Int. 2004, 24, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.; Nag, S.; Bardhan, S.; Acharya, D.; Paul, R.; Dey, R.; Ghosh, M.; Saha, I. The role of long-term doxycycline in patients of idiopathic pulmonaryfibrosis: The results of an open prospective trial. Lung India 2009, 26, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Bhattacharya, P.; Paul, S.; Paul, R.; Swarnakar, S. An alternative therapy for idiopathic pulmonary fibrosis by doxycycline through matrix metalloproteinase inhibition. Lung India 2011, 28, 174–179. [Google Scholar] [PubMed]
- England, K.A.; Price, A.P.; Tram, K.V.; Shapiro, S.D.; Blazar, B.R.; Panoskaltsis-Mortari, A. Evidence for early fibrosis and increased airway resistance in bone marrow transplant recipient mice deficient in MMP12. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L519–L526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.R.; Cho, S.J.; Lee, C.G.; Homer, R.J.; Elias, J.A. Transforming growth factor (TGF)-beta1 stimulates pulmonary fibrosis and inflammation via a Bax-dependent, bid-activated pathway that involves matrix metalloproteinase-12. J. Biol. Chem. 2007, 282, 7723–7732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanone, S.; Zheng, T.; Zhu, Z.; Liu, W.; Lee, C.G.; Ma, B.; Chen, Q.; Homer, R.J.; Wang, J.; Rabach, L.A.; et al. Overlapping and enzyme-specific contributions of matrix metalloproteinases-9 and -12 in IL-13-induced inflammation and remodeling. J. Clin. Investig. 2002, 110, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Craig, V.J.; Zhang, L.; Hagood, J.S.; Owen, C.A. Matrix metalloproteinases as therapeutic targets for idiopathic pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 53, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Margulis, A.; Nocka, K.H.; Wood, N.L.; Wolf, S.F.; Goldman, S.J.; Kasaian, M.T. MMP dependence of fibroblast contraction and collagen production induced by human mast cell activation in a three-dimensional collagen lattice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L236–L247. [Google Scholar] [CrossRef]
- Malek, V.; Gaikwad, A.B. Telmisartan and thiorphan combination treatment attenuates fibrosis and apoptosis in preventing diabetic cardiomyopathy. Cardiovasc. Res. 2019, 115, 373–384. [Google Scholar] [CrossRef]
- Corbel, M.; Caulet-Maugendre, S.; Germain, N.; Molet, S.; Lagente, V.; Boichot, E. Inhibition of bleomycin-induced pulmonary fibrosis in mice by the matrix metalloproteinase inhibitor batimastat. J. Pathol. 2001, 193, 538–545. [Google Scholar] [CrossRef]
- George, J.; Tsutsumi, M.; Tsuchishima, M. MMP-13 deletion decreases profibrogenic molecules and attenuates N-nitrosodimethylamine-induced liver injury and fibrosis in mice. J. Cell. Mol. Med. 2017, 21, 3821–3835. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.T. Matrix metalloproteinase inhibition and the prevention of heart failure. Trends Cardiovasc. Med. 2001, 11, 202–205. [Google Scholar] [CrossRef]
- Fujita, M.; Ye, Q.; Ouchi, H.; Harada, E.; Inoshima, I.; Kuwano, K.; Nakanishi, Y. Doxycycline attenuated pulmonary fibrosis induced by bleomycin in mice. Antimicrob. Agents Chemother. 2006, 50, 739–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, Y.; Kunihiro, S.; Sato, S.; Yoshioka, K.; Hara, Y.; Kanai, K.; Hoshi, F.; Itoh, N.; Higuchi, S. Doxycycline attenuates isoproterenol-induced myocardial fibrosis and matrix metalloproteinase activity in rats. Biol. Pharm. Bull. 2009, 32, 1678–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, R.J.; Fattman, C.L.; Niehouse, L.M.; Tobolewski, J.M.; Hanford, L.E.; Li, Q.; Monzon, F.A.; Parks, W.C.; Oury, T.D. Matrix metalloproteinases promote inflammation and fibrosis in asbestos-induced lung injury in mice. Am. J. Respir. Cell Mol. Biol. 2006, 35, 289–297. [Google Scholar] [CrossRef]
- de Meijer, V.E.; Sverdlov, D.Y.; Popov, Y.; Le, H.D.; Meisel, J.A.; Nose, V.; Schuppan, D.; Puder, M. Broad-spectrum matrix metalloproteinase inhibition curbs inflammation and liver injury but aggravates experimental liver fibrosis in mice. PLoS ONE 2010, 5, e11256. [Google Scholar] [CrossRef] [Green Version]
- Pulli, B.; Ali, M.; Iwamoto, Y.; Zeller, M.W.; Schob, S.; Linnoila, J.J.; Chen, J.W. Myeloperoxidase-Hepatocyte-Stellate Cell Cross Talk Promotes Hepatocyte Injury and Fibrosis in Experimental Nonalcoholic Steatohepatitis. Antioxid. Redox Signal. 2015, 23, 1255–1269. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, J.; Telljohann, R.; Jiang, L.; Wu, J.; Monticone, R.E.; Kapoor, K.; Talan, M.; Lakatta, E.G. Chronic matrix metalloproteinase inhibition retards age-associated arterial proinflammation and increase in blood pressure. Hypertension 2012, 60, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, P.; Peterson, J.T.; Subramanian, V.; Singh, M.; Singh, K. Inhibition of matrix metalloproteinases improves left ventricular function in mice lacking osteopontin after myocardial infarction. Mol. Cell. Biochem. 2009, 322, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Parks, W.C.; Wilson, C.L.; Lopez-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Jia, M.X.; Wang, J.H.; Lu, J.L.; Deng, J.; Tang, J.X.; Liu, C. Association of MMP9-1562C/T and MMP13-77A/G Polymorphisms with Non-Small Cell Lung Cancer in Southern Chinese Population. Biomolecules 2019, 9, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamenkovic, I. Matrix metalloproteinases in tumor invasion and metastasis. Semin. Cancer Biol. 2000, 10, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Hart, E.; Shchurin, A.; Hoover-Plow, J. Inflammatory macrophage migration requires MMP-9 activation by plasminogen in mice. J. Clin. Investig. 2008, 118, 3012–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.Y.; Birkland, T.P.; Howe, J.D.; Rowan, A.D.; Fidock, M.; Parks, W.C.; Gavrilovic, J. Macrophage migration and invasion is regulated by MMP10 expression. PLoS ONE 2013, 8, e63555. [Google Scholar] [CrossRef] [Green Version]
- Shipley, J.M.; Wesselschmidt, R.L.; Kobayashi, D.K.; Ley, T.J.; Shapiro, S.D. Metalloelastase is required for macrophage-mediated proteolysis and matrix invasion in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 3942–3946. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.H.; Gao, B.T.; Goldsmith, Z.K.; Irvine, A.S.; Saleh, N.; Lee, R.P.; Lendermon, J.B.; Bheemreddy, R.; Zhang, Q.; Brennan, R.C.; et al. Inhibition of MMP-2 and MMP-9 decreases cellular migration, and angiogenesis in in vitro models of retinoblastoma. BMC Cancer 2017, 17, 434. [Google Scholar] [CrossRef]
- Masson, V.; de la Ballina, L.R.; Munaut, C.; Wielockx, B.; Jost, M.; Maillard, C.; Blacher, S.; Bajou, K.; Itoh, T.; Itohara, S.; et al. Contribution of host MMP-2 and MMP-9 to promote tumor vascularization and invasion of malignant keratinocytes. FASEB J. 2005, 19, 234–236. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Rosenberg, G.A. Matrix metalloproteinases as therapeutic targets for stroke. Brain Res. 2015, 1623, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix metalloproteinase-9: Many shades of function in cardiovascular disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, T.Y.; Tsao, S.M.; Yeh, C.B.; Yang, S.F. Matrix metalloproteinases in pneumonia. Clin. Chim. Acta 2014, 433, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh, G.; Quchani, S.H.; Sahraian, M.A.; Abolhassani, F.; Gilani, M.A.S.; Tarzjani, M.D.; Atoof, F. Leukocyte Gene Expression and Plasma Concentration in Multiple Sclerosis: Alteration of Transforming Growth Factor-betas, Claudin-11, and Matrix Metalloproteinase-2. Cell. Mol. Neurobiol. 2016, 36, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Amorosa, L.F.; Coyle, S.M.; Macor, M.A.; Lubitz, S.E.; Carson, J.L.; Birnbaum, M.J.; Lee, L.Y.; Haimovich, B. Proteolytic Cleavage of AMPKalpha and Intracellular MMP9 Expression Are Both Required for TLR4-Mediated mTORC1 Activation and HIF-1alpha Expression in Leukocytes. J. Immunol. 2015, 195, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.S.; Park, J.S.; Choi, I.Y.; Kim, W.K.; Kim, H.S. Inhibition of MMP-3 or -9 suppresses lipopolysaccharide-induced expression of proinflammatory cytokines and iNOS in microglia. J. Neurochem. 2008, 106, 770–780. [Google Scholar] [CrossRef]
- Berg, G.; Barchuk, M.; Miksztowicz, V. Behavior of Metalloproteinases in Adipose Tissue, Liver and Arterial Wall: An Update of Extracellular Matrix Remodeling. Cells 2019, 8, 158. [Google Scholar] [CrossRef] [Green Version]
- Javaid, M.A.; Abdallah, M.N.; Ahmed, A.S.; Sheikh, Z. Matrix metalloproteinases and their pathological upregulation in multiple sclerosis: An overview. Acta Neurol. Belg. 2013, 113, 381–390. [Google Scholar] [CrossRef]
- Hansmann, F.; Zhang, N.; Herder, V.; Leitzen, E.; Baumgartner, W. Delayed Astrogliosis Associated with Reduced M1 Microglia Activation in Matrix Metalloproteinase 12 Knockout Mice during Theiler’s Murine Encephalomyelitis. Int. J. Mol. Sci. 2019, 20, 1702. [Google Scholar] [CrossRef] [Green Version]
- Vedam-Mai, V.; Yachnis, A.; Ullman, M.; Javedan, S.P.; Okun, M.S. Postmortem observation of collagenous lead tip region fibrosis as a rare complication of DBS. Mov. Disord. 2012, 27, 565–569. [Google Scholar] [CrossRef]
- Yong, V.W.; Power, C.; Forsyth, P.; Edwards, D.R. Metalloproteinases in biology and pathology of the nervous system. Nat. Rev. Neurosci. 2001, 2, 502–511. [Google Scholar] [CrossRef]
- Shimoda, M. Extracellular vesicle-associated MMPs: A modulator of the tissue microenvironment. Adv. Clin. Chem. 2019, 88, 35–66. [Google Scholar] [PubMed]
- Ferguson, S.W.; Wang, J.; Lee, C.J.; Liu, M.; Neelamegham, S.; Canty, J.M.; Nguyen, J. The microRNA regulatory landscape of MSC-derived exosomes: A systems view. Sci. Rep. 2018, 8, 1419. [Google Scholar] [CrossRef] [PubMed]
- Koeck, E.S.; Iordanskaia, T.; Sevilla, S.; Ferrante, S.C.; Hubal, M.J.; Freishtat, R.J.; Nadler, E.P. Adipocyte exosomes induce transforming growth factor beta pathway dysregulation in hepatocytes: A novel paradigm for obesity-related liver disease. J. Surg. Res. 2014, 192, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Hakulinen, J.; Sankkila, L.; Sugiyama, N.; Lehti, K.; Keski-Oja, J. Secretion of active membrane type 1 matrix metalloproteinase (MMP-14) into extracellular space in microvesicular exosomes. J. Cell. Biochem. 2008, 105, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Han, K.Y.; Dugas-Ford, J.; Seiki, M.; Chang, J.H.; Azar, D.T. Evidence for the Involvement of MMP14 in MMP2 Processing and Recruitment in Exosomes of Corneal Fibroblasts. Invest. Ophthalmol. Vis. Sci. 2015, 56, 5323–5329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalfe, P.D.; Wang, J.; Jiao, H.; Huang, Y.; Hori, K.; Moore, R.B.; Tredget, E.E. Bladder outlet obstruction: Progression from inflammation to fibrosis. BJU Int. 2010, 106, 1686–1694. [Google Scholar] [CrossRef]
- Prabhu, S.D.; Frangogiannis, N.G. The Biological Basis for Cardiac Repair After Myocardial Infarction: From Inflammation to Fibrosis. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef]
- Daniels, C.E.; Lasky, J.A.; Limper, A.H.; Mieras, K.; Gabor, E.; Schroeder, D.R. Imatinib treatment for idiopathic pulmonary fibrosis: Randomized placebo-controlled trial results. Am. J. Respir. Crit. Care Med. 2010, 181, 604–610. [Google Scholar] [CrossRef]
- Boettcher, E.; Csako, G.; Pucino, F.; Wesley, R.; Loomba, R. Meta-analysis: Pioglitazone improves liver histology and fibrosis in patients with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2012, 35, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Raghu, G.; Anstrom, K.J.; King, T.E., Jr.; Lasky, J.A.; Martinez, F.J. Prednisone, azathioprine, and N-acetylcysteine for pulmonary fibrosis. N. Engl. J. Med. 2012, 366, 1968–1977. [Google Scholar] [CrossRef]
- Rafii, R.; Juarez, M.M.; Albertson, T.E.; Chan, A.L. A review of current and novel therapies for idiopathic pulmonary fibrosis. J. Thorac. Dis. 2013, 5, 48–73. [Google Scholar] [PubMed]
- Ahluwalia, N.; Shea, B.S.; Tager, A.M. New therapeutic targets in idiopathic pulmonary fibrosis. Aiming to rein in runaway wound-healing responses. Am. J. Respir. Crit. Care Med. 2014, 190, 867–878. [Google Scholar] [CrossRef] [PubMed]
- McKeown, S.; Richter, A.G.; O’Kane, C.; McAuley, D.F.; Thickett, D.R. MMP expression and abnormal lung permeability are important determinants of outcome in IPF. Eur. Respir. J. 2009, 33, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, C.M.; Dolgonos, L.; Zemans, R.L.; Young, S.K.; Robertson, J.; Briones, N.; Suzuki, T.; Campbell, M.N.; Gauldie, J.; Radisky, D.C.; et al. Matrix metalloproteinase 3 is a mediator of pulmonary fibrosis. Am. J. Pathol. 2011, 179, 1733–1745. [Google Scholar] [CrossRef]
- Willems, S.; Verleden, S.E.; Vanaudenaerde, B.M.; Wynants, M.; Dooms, C.; Yserbyt, J.; Somers, J.; Verbeken, E.K.; Verleden, G.M.; Wuyts, W.A. Multiplex protein profiling of bronchoalveolar lavage in idiopathic pulmonary fibrosis and hypersensitivity pneumonitis. Ann. Thorac. Med. 2013, 8, 38–45. [Google Scholar]
- Yu, G.; Kovkarova-Naumovski, E.; Jara, P.; Parwani, A.; Kass, D.; Ruiz, V.; Lopez-Otin, C.; Rosas, I.O.; Gibson, K.F.; Cabrera, S.; et al. Matrix metalloproteinase-19 is a key regulator of lung fibrosis in mice and humans. Am. J. Respir. Crit. Care Med. 2012, 186, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Rohani, M.G.; Dimitrova, E.; Beppu, A.; Wang, Y.; Jefferies, C.A.; Parks, W.C. Macrophage MMP10 Regulates TLR7-Mediated Tolerance. Front. Immunol. 2018, 9, 2817. [Google Scholar] [CrossRef]
- Kriz, W.; Kaissling, B.; Le Hir, M. Epithelial-mesenchymal transition (EMT) in kidney fibrosis: Fact or fantasy? J. Clin. Investig. 2011, 121, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.L.; Zhu, R.T.; Sun, Y.L. Epithelial-mesenchymal transition in liver fibrosis. Biomed. Rep. 2016, 4, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Chuang, H.M.; Su, H.L.; Li, C.; Lin, S.Z.; Yen, S.Y.; Huang, M.H.; Ho, L.I.; Chiou, T.W.; Harn, H.J. The Role of Butylidenephthalide in Targeting the Microenvironment Which Contributes to Liver Fibrosis Amelioration. Front. Pharmacol. 2016, 7, 112. [Google Scholar] [CrossRef] [Green Version]
- Marmai, C.; Sutherland, R.E.; Kim, K.K.; Dolganov, G.M.; Fang, X.; Kim, S.S.; Jiang, S.; Golden, J.A.; Hoopes, C.W.; Matthay, M.A.; et al. Alveolar epithelial cells express mesenchymal proteins in patients with idiopathic pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L71–L78. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.Y.; Huang, T.P.; Yang, W.C.; Chen, Z.P.; Yang, A.H.; Mu, W.; Nikolic-Paterson, D.J.; Atkins, R.C.; Lan, H.Y. Tubular epithelial-myofibroblast transdifferentiation in progressive tubulointerstitial fibrosis in 5/6 nephrectomized rats. Kidney Int. 1998, 54, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, S.H. Genesis of the myofibroblast in lung injury and fibrosis. Proc. Am. Thorac. Soc. 2012, 9, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Tian, L.; He, L.S.; Soni, B.; Shang, H.T. Myofibroblasts and their resistance to apoptosis: A possible mechanism of osteoradionecrosis. Clin. Cosmet. Investig. Dent. 2012, 4, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Strand, S.; Vollmer, P.; van den Abeelen, L.; Gottfried, D.; Alla, V.; Heid, H.; Kuball, J.; Theobald, M.; Galle, P.R.; Strand, D. Cleavage of CD95 by matrix metalloproteinase-7 induces apoptosis resistance in tumour cells. Oncogene 2004, 23, 3732–3736. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Miyake, M.; Lawton, A.; Goodison, S.; Rosser, C.J. Matrix metalloproteinase-10 promotes tumor progression through regulation of angiogenic and apoptotic pathways in cervical tumors. BMC Cancer 2014, 14, 310. [Google Scholar] [CrossRef] [Green Version]
- Seeland, U.; Haeuseler, C.; Hinrichs, R.; Rosenkranz, S.; Pfitzner, T.; Scharffetter-Kochanek, K.; Bohm, M. Myocardial fibrosis in transforming growth factor-beta(1) (TGF-beta(1)) transgenic mice is associated with inhibition of interstitial collagenase. Eur. J. Clin. Investig. 2002, 32, 295–303. [Google Scholar] [CrossRef]
- Robert, S.; Gicquel, T.; Bodin, A.; Lagente, V.; Boichot, E. Characterization of the MMP/TIMP Imbalance and Collagen Production Induced by IL-1beta or TNF-alpha Release from Human Hepatic Stellate Cells. PLoS ONE 2016, 11, e0153118. [Google Scholar] [CrossRef]
- Zheng, H.; Takahashi, H.; Murai, Y.; Cui, Z.; Nomoto, K.; Niwa, H.; Tsuneyama, K.; Takano, Y. Expressions of MMP-2, MMP-9 and VEGF are closely linked to growth, invasion, metastasis and angiogenesis of gastric carcinoma. Anticancer. Res. 2006, 26, 3579–3583. [Google Scholar]
- Ardi, V.C.; Kupriyanova, T.A.; Deryugina, E.I.; Quigley, J.P. Human neutrophils uniquely release TIMP-free MMP-9 to provide a potent catalytic stimulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 20262–20267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Fu, A.A.; Misra, K.D.; Shergill, U.M.; Leof, E.B.; Mukhopadhyay, D. Hypoxia-induced phenotypic switch of fibroblasts to myofibroblasts through a matrix metalloproteinase 2/tissue inhibitor of metalloproteinase-mediated pathway: Implications for venous neointimal hyperplasia in hemodialysis access. J. Vasc. Interv. Radiol. 2010, 21, 896–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Novoa, J.M.; Nieto, M.A. Inflammation and EMT: An alliance towards organ fibrosis and cancer progression. EMBO Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhard, U.; Huesgen, P.F.; Schilling, O.; Bellac, C.L.; Butler, G.S.; Cox, J.H.; Dufour, A.; Goebeler, V.; Kappelhoff, R.; Keller, U.A.D.; et al. Active site specificity profiling of the matrix metalloproteinase family: Proteomic identification of 4300 cleavage sites by nine MMPs explored with structural and synthetic peptide cleavage analyses. Matrix Biol. 2016, 49, 37–60. [Google Scholar] [CrossRef]
- Huang, H. Matrix Metalloproteinase-9 (MMP-9) as a Cancer Biomarker and MMP-9 Biosensors: Recent Advances. Sensors 2018, 18, 3249. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Expression of 20 types of matrix metalloproteinases (MMPs) (MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-16, MMP-17, MMP-19, MMP-20, MMP-21, MMP-24, MMP-25, and MMP-27) in 16 human organs (adipose, adrenal, brain, breast, colon, heart, kidney, liver, lung, lymph node, ovary, prostate, skeletal muscle, testes, thyroid, and white blood cells). The reads per kilobase per million mapped reads (RPKM) value compares the gene expression with the sample sequencing depth and gene length. (A) The expression ratio of different MMP types in human organs. (B) The distribution ratio of (B) all MMPs and (C) seven major MMPs in 16 organs.
Figure 1.
Expression of 20 types of matrix metalloproteinases (MMPs) (MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-16, MMP-17, MMP-19, MMP-20, MMP-21, MMP-24, MMP-25, and MMP-27) in 16 human organs (adipose, adrenal, brain, breast, colon, heart, kidney, liver, lung, lymph node, ovary, prostate, skeletal muscle, testes, thyroid, and white blood cells). The reads per kilobase per million mapped reads (RPKM) value compares the gene expression with the sample sequencing depth and gene length. (A) The expression ratio of different MMP types in human organs. (B) The distribution ratio of (B) all MMPs and (C) seven major MMPs in 16 organs.


{kind=link}
{kind=link}
Table 1.
Experimental evidence of effective MMP inhibitors against fibrosis in animal models including of lung, liver, and myocardial fibrosis.
Table 1.
Experimental evidence of effective MMP inhibitors against fibrosis in animal models including of lung, liver, and myocardial fibrosis.
| Compound Name | Description | Effects | CAS Number | Refs |
|---|---|---|---|---|
| Batimastat | A broad-spectrum MMP inhibitor | Inhibit pulmonary fibrosis | 130370-60-4 | [81] |
| CL 82198 hydrochloride | A selective inhibitor of MMP-13 | Blocks liver fibrosis | 307002-71-7 | [82] |
| CP 471474 | An MMP inhibitor | Inhibit collagen in myocardial fibrosis | 210755-45-6 | [83] |
| Doxycycline Hyclate | An antimicrobial tetracycline that acts as an inhibitor of MMP-1, MMP-8 and MMP-9 | Attenuated pulmonary/myocardial fibrosis | 24390-14-5 | [84,85] [74] |
| Reduced parameters in IPF patients | ||||
| GM 6001 | A cell permeable MMP and fibroblast collagenase inhibitor | Reduced pulmonary inflammation and fibrosis | 142880-36-2 | [86] |
| Marimastat | A broad-spectrum MMP inhibitor and selective TACE inhibitor | Aggravates liver fibrosis | 154039-60-8 | [87,88] |
| PD166793 | A potent MMP-2, MMP-3, and MMP-13 inhibitor | Retardation of age-associated arterial fibrosis | 199850-67-4 | [89] [90] |
| Reduced myocardial fibrosis | ||||
| Thiorphan (DL) | An enkephalinase and metalloproteinase inhibitor | Reduced myocardial fibrosis | 76721-89-6 | [80] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chuang, H.-M.; Chen, Y.-S.; Harn, H.-J. The Versatile Role of Matrix Metalloproteinase for the Diverse Results of Fibrosis Treatment. Molecules 2019, 24, 4188. https://doi.org/10.3390/molecules24224188
AMA Style
Chuang H-M, Chen Y-S, Harn H-J. The Versatile Role of Matrix Metalloproteinase for the Diverse Results of Fibrosis Treatment. Molecules. 2019; 24(22):4188. https://doi.org/10.3390/molecules24224188
Chicago/Turabian StyleChuang, Hong-Meng, Yu-Shuan Chen, and Horng-Jyh Harn. 2019. "The Versatile Role of Matrix Metalloproteinase for the Diverse Results of Fibrosis Treatment" Molecules 24, no. 22: 4188. https://doi.org/10.3390/molecules24224188