
Oligonucleotide Binding to Non-B-DNA in MYC

 , and
, and
Abstract
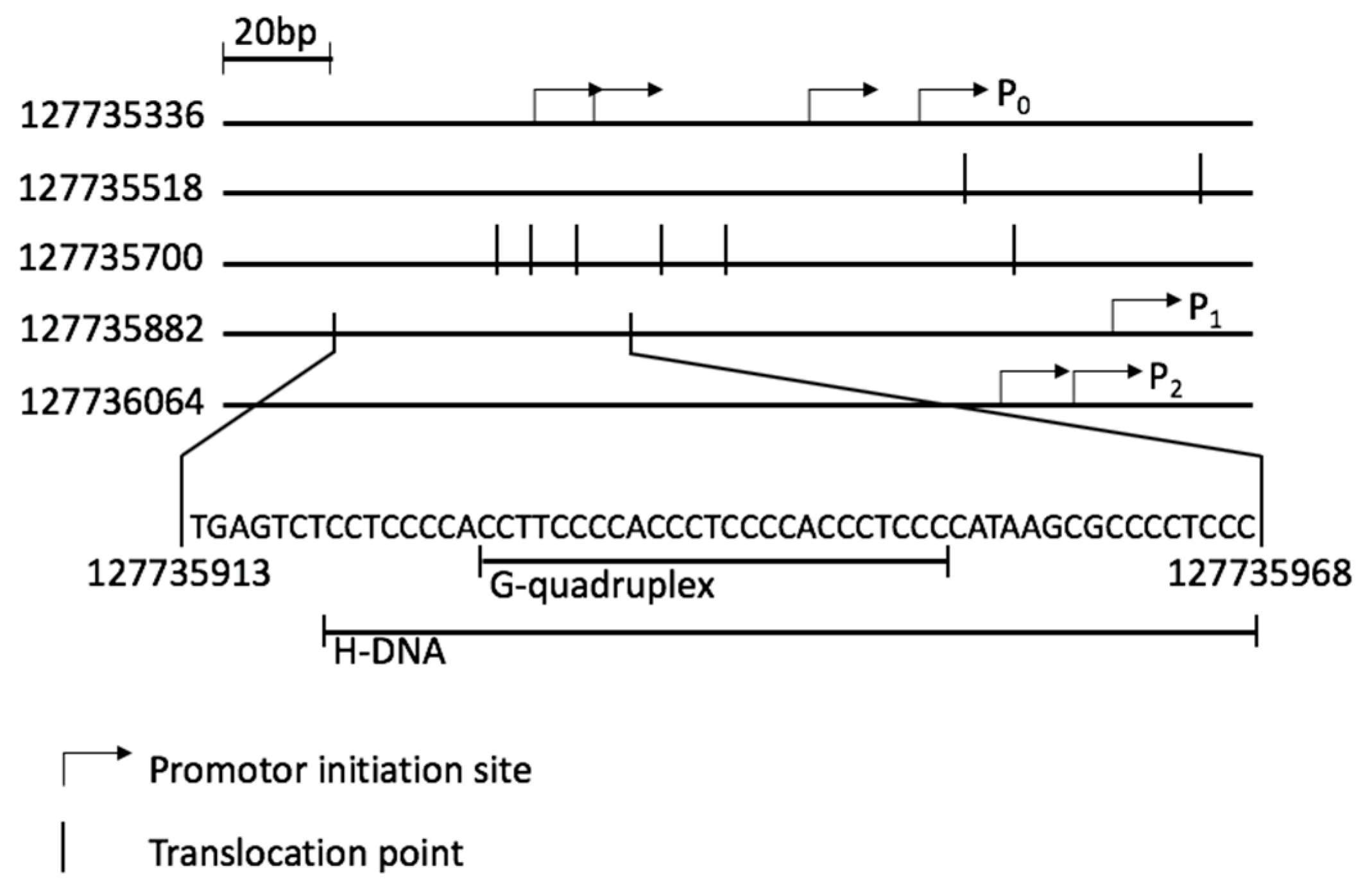
:1. Introduction
2. Results and Discussion
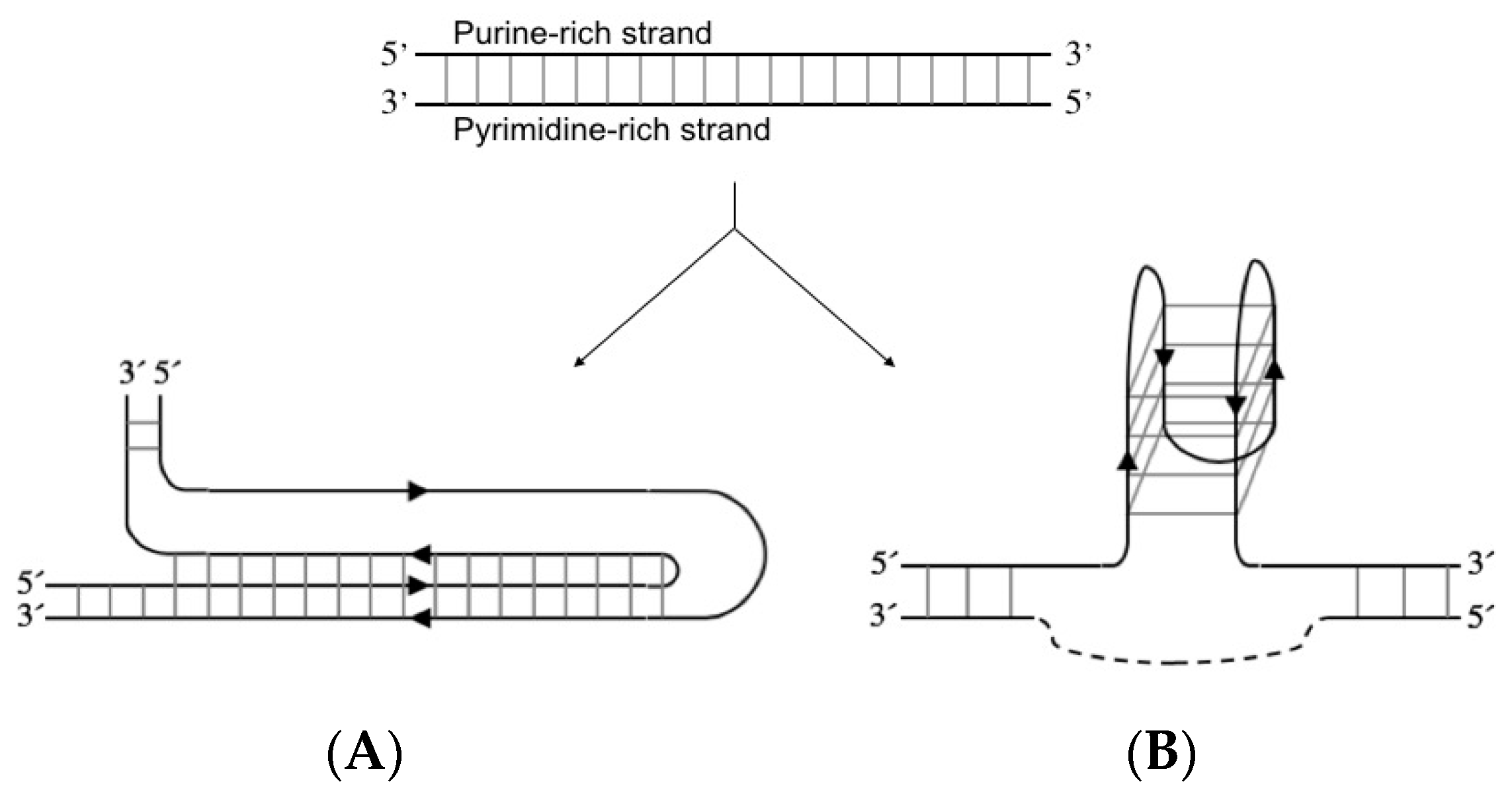
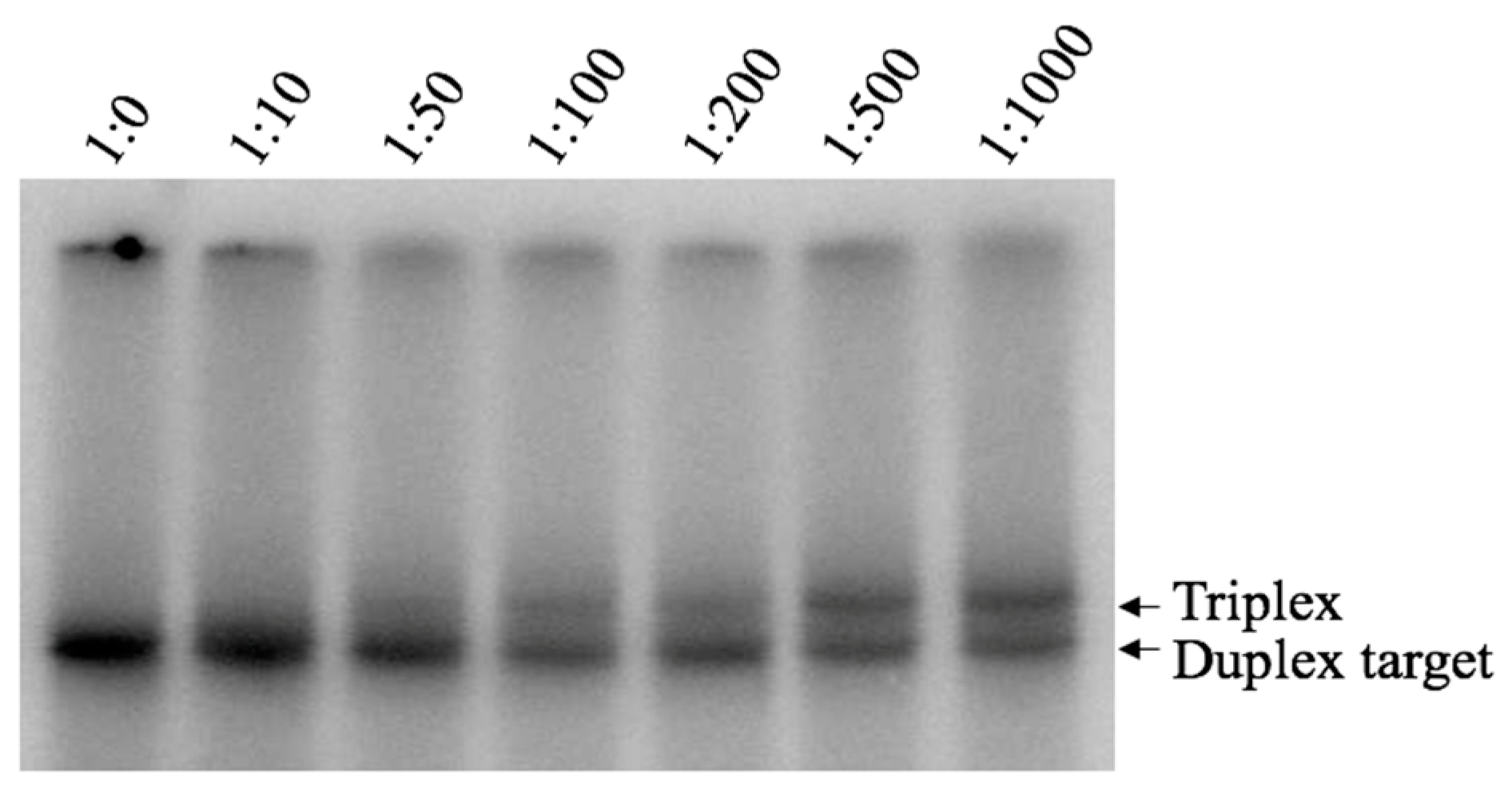
2.1. Intermolecular Triplex Is Formed with Pu TFO But Not with Py TFO
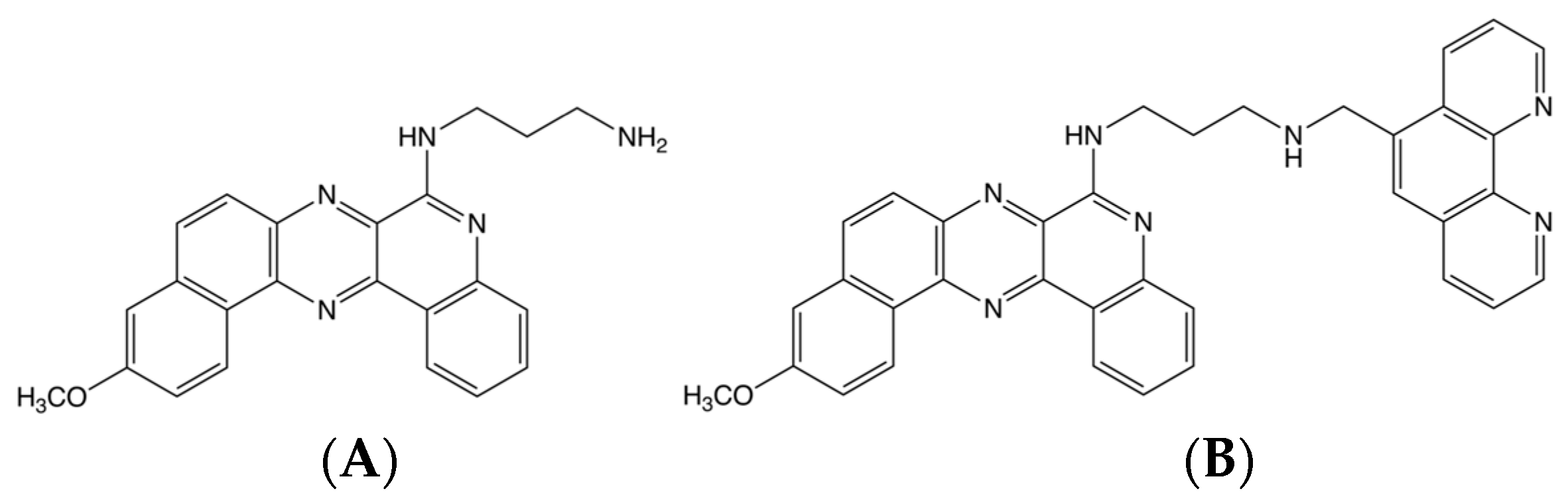
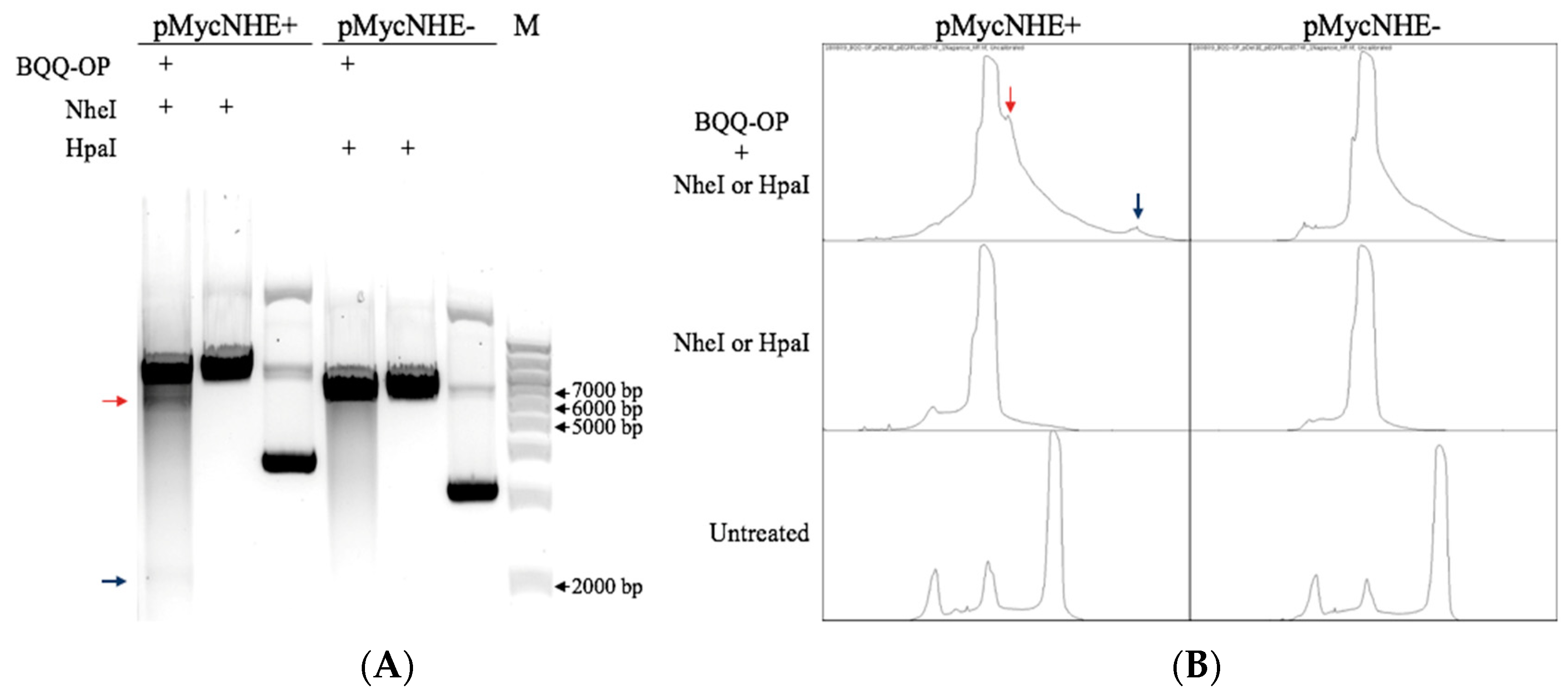
2.2. BQQ-OP Cleavage of pMycNHE+ Detects the Presence of H-DNA
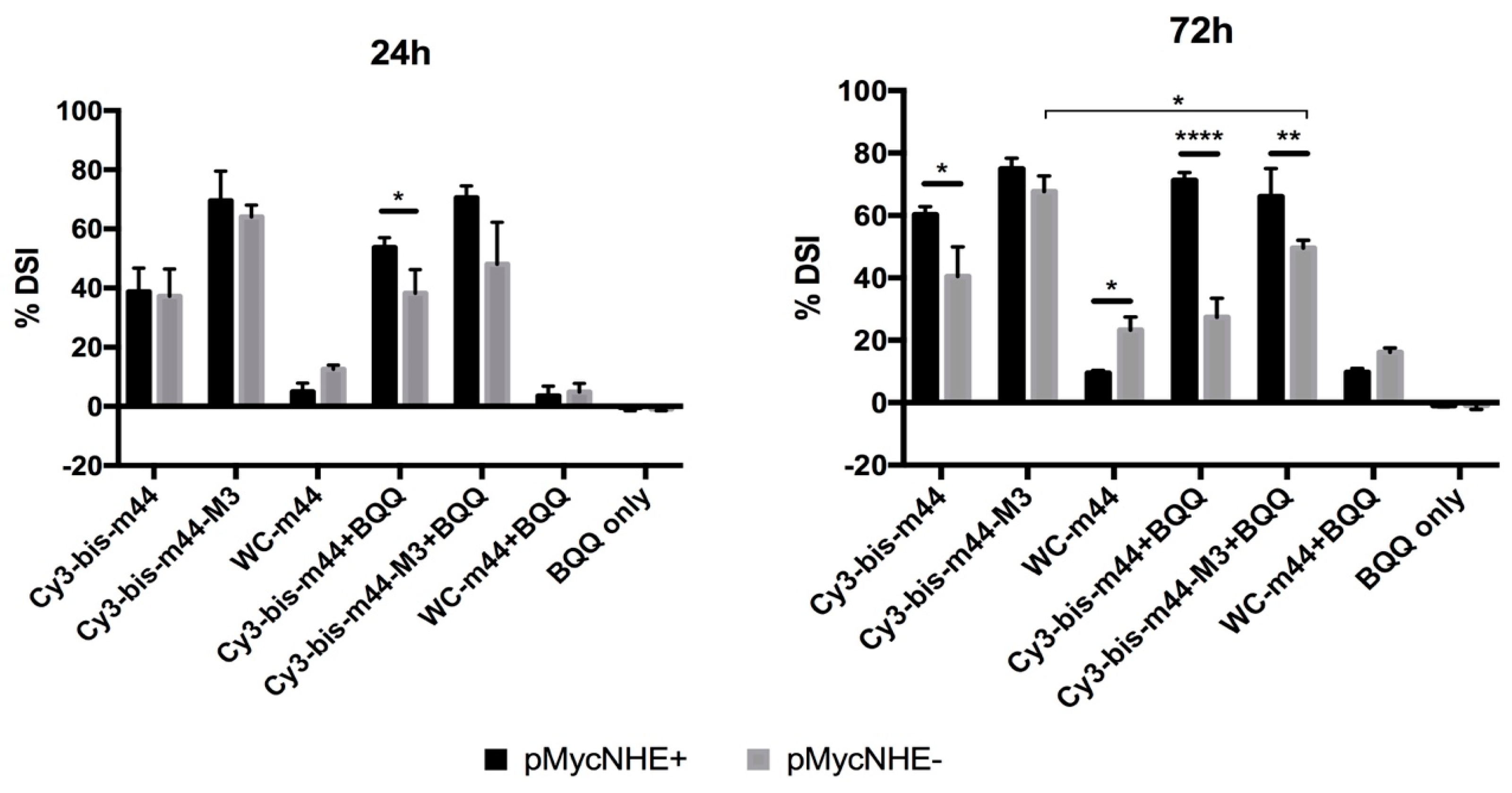
2.3. Influence of H-DNA Formation on bisLNA Mediated dsDNA Strand-Invasion
3. Materials and Methods
3.1. TFO Design and Electrophoretic Mobility Shift Assay (EMSA)
3.2. BQQ-OP-Cleavage
3.3. bisLNA Strand-Invasion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, J.C. Helical repeat of DNA in solution. Proc. Natl. Acad. Sci. USA 1979, 76, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Guliang, W.; Vasquez, K.M. Naturally occurring H-DNA-forming sequences are mutagenic in mammalian cells. Proc. Natl. Acad. Sci. USA 2004, 101, 13448–13453. [Google Scholar]
- Gabay, M.; Li, Y.; Felsher, D.W. MYC activation is a hallmark of cancer initiation and maintenance. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.; Ceribelli, M.; Pittaluga, S.; Wright, G.; Staudt, L.M. Oncogenic mechanisms in Burkitt lymphoma. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.A. Burkitt’s Lymphoma: Clinicopathologic Features and Differential Diagnosis. Oncologist 2006, 11, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.L.; Aster, J.C. Molecular biology of Burkitt’s lymphoma. J. Clin. Oncol. 2000, 18, 3707–3721. [Google Scholar] [CrossRef] [PubMed]
- Casulo, C.; Friedberg, J.W. Burkitt lymphoma—A rare but challenging lymphoma. Best Pract. Res. Clin. Haematol. 2018, 31, 279–284. [Google Scholar] [CrossRef]
- Carroll, P.; Freie, B.; Mathsyaraja, H.; Eisenman, R. The MYC transcription factor network: Balancing metabolism, proliferation and oncogenesis. Front. Med. 2018, 12, 412–425. [Google Scholar] [CrossRef]
- Wierstra, I.; Alves, J. The c-myc promoter: Still MysterY and challenge. Adv. Cancer Res. 2008, 99, 113–333. [Google Scholar]
- Battey, J.; Moulding, C.; Taub, R.; Murphy, W.; Stewart, T.; Potter, H.; Lenoir, G.; Leder, P. The human c-myc oncogene: Structural consequences of translocation into the igh locus in Burkitt lymphoma. Cell 1983, 34, 779–787. [Google Scholar] [CrossRef]
- Saito, H.; Hayday, A.C.; Wiman, K.; Hayward, W.S.; Tonegawa, S. Activation of the c- myc gene by translocation: A model for translational control. Proc. Natl. Acad. Sci. USA 1983, 80, 7476. [Google Scholar] [CrossRef]
- Siebenlist, U.; Hennighausen, L.; Battey, J.; Leder, P. Chromatin structure and protein binding in the putative regulatory region of the c- myc gene in burkitt lymphoma. Cell 1984, 37, 381–391. [Google Scholar] [CrossRef]
- Kinniburgh, A.J. A cis-acting transcription element of the c-myc gene can assume an H- DNA conformation. Nucleic Acids Res. 1989, 17, 8412. [Google Scholar] [CrossRef]
- Krasilnikov, A.S.; Panyutin, I.G.; Samadashwily, G.M.; Cox, R.; Lazurkin, Y.S.; Mirkin, S.M. Mechanisms of triplex-caused polymerization arrest. Nucleic Acids Res. 1997, 25, 1339. [Google Scholar] [CrossRef]
- Wang, G.; Vasquez, K. Effects of Replication and Transcription on DNA Structure-Related Genetic Instability. Genes 2017, 8, 17. [Google Scholar] [CrossRef]
- Valton, A.-L.; Prioleau, M.-N. G-Quadruplexes in DNA Replication: A Problem or a Necessity? Trends Genet. 2016, 32, 697–706. [Google Scholar] [CrossRef]
- Wilda, M.; Busch, K.; Klose, I.; Keller, T.; Woessmann, W.; Kreuder, J.; Harbott, J.; Borkhardt, A. Level of MYC overexpression in pediatric Burkitt’s lymphoma is strongly dependent on genomic breakpoint location within the MYC locus. Genes Chromosomes Cancer 2004, 41, 178–182. [Google Scholar] [CrossRef]
- Ralf, K.; Riccardo, D.-F. Mechanisms of chromosomal translocations in B cell lymphomas. Oncogene 2001, 20, 5580–5594. [Google Scholar]
- Gazin, C.; Dupont de Dinechin, S.; Hampe, A.; M Masson, J.; Martin, P.; Stehelin, D.; Galibert, F. Nucleotide sequence of the human c-myc locus: Provocative open reading frame within the first exon. Embo J. 1984, 3, 383–387. [Google Scholar] [CrossRef]
- Mirkin, S.M.; Frank-Kamenetskii, M.D. H-DNA and Related Structures. Annu. Rev. Biophys. Biomol. Struct. 1994, 23, 541–576. [Google Scholar] [CrossRef]
- Zain, R.; Sun, J.S. Do natural DNA triple-helical structures occur and function in vivo? CMLS 2003, 60, 862–870. [Google Scholar] [CrossRef]
- Palecek, E. Local supercoil-stabilized DNA structures. Crit. Rev. Biochem. Mol. Biol. 1991, 26, 151–226. [Google Scholar] [CrossRef]
- Davis, J.T. G-quartets 40 years later: From 5′-GMP to molecular biology and supramolecular chemistry. Angew. Chem. Int. Ed. 2004, 43, 668–698. [Google Scholar] [CrossRef]
- Matthew, L.B.; Katrin, P.; Virginia, A.Z. DNA secondary structures: Stability and function of G-quadruplex structures. Nat. Rev. Genet. 2012, 13, 770–780. [Google Scholar]
- Zeraati, M.; Langley, D.B.; Schofield, P.; Moye, A.L.; Rouet, R.; Hughes, W.E.; Bryan, T.M.; Dinger, M.E.; Christ, D. I-motif DNA structures are formed in the nuclei of human cells. Nat. Chem. 2018, 10, 631–637. [Google Scholar] [CrossRef]
- Cui, Y.; Kong, D.; Ghimire, C.; Cuixia, X.; Hanbin, M. Mutually exclusive formation of G-quadruplex and i-motif is a general phenomenon governed by steric hindrance in duplex DNA. Biochemistry 2016, 55, 2291–2299. [Google Scholar] [CrossRef]
- Jain, A.; Wang, G.; Vasquez, K.M. DNA triple helices: Biological consequences and therapeutic potential. Biochimie 2008, 90, 1117–1130. [Google Scholar] [CrossRef]
- Shalaby, T.; Fiaschetti, G.; Nagasawa, K.; Shin-Ya, K.; Baumgartner, M.; Grotzer, M. G-Quadruplexes as Potential Therapeutic Targets for Embryonal Tumors. Molecules 2013, 18, 12500–12537. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Wynand, P.R.; Adam, D.T.; Bernd, K. DNA damage and the balance between survival and death in cancer biology. Nat. Rev. Cancer 2015, 16, 20–33. [Google Scholar]
- Kendrick, S.; Muranyi, A.; Gokhale, V.; Hurley, L.H.; Rimsza, L.M. Simultaneous Drug Targeting of the Promoter MYC G-Quadruplex and BCL2 i-Motif in Diffuse Large B-Cell Lymphoma Delays Tumor Growth. J. Med. Chem. 2017, 60, 6587–6597. [Google Scholar] [CrossRef]
- Neidle, S. The structures of quadruplex nucleic acids and their drug complexes. Curr. Opin. Struct. Biol. 2009, 19, 239–250. [Google Scholar] [CrossRef]
- Maolin, W.; Yuanyuan, Y.; Chao, L.; Aiping, L.; Ge, Z. Recent Advances in Developing Small Molecules Targeting Nucleic Acid. Int. J. Mol. Sci. 2016, 17, 779. [Google Scholar]
- Napoli, S.; Negri, U.; Arcamone, F.; Capobianco, M.L.; Carbone, G.M.; Catapano, C.V. Growth inhibition and apoptosis induced by daunomycin-conjugated triplex-forming oligonucleotides targeting the c-myc gene in prostate cancer cells. Nucleic Acids Res. 2006, 34, 734–744. [Google Scholar] [CrossRef]
- McGuffie, E.M.; Catapano, C.V. Design of a novel triple helix-forming oligodeoxyribonucleotide directed to the major promoter of the c-myc gene. Nucleic Acids Res. 2002, 30, 2701–2709. [Google Scholar] [CrossRef]
- Giovanna, C.; Elisabetta, M.C.; Massimo, U.; Silvio, R.; Olfert, L.; Manlio, F.; Lidia, C.B. Effects in live cells of a c-myc anti-gene PNA linked to a nuclear localization signal. Nat. Biotechnol. 2000, 18, 300–303. [Google Scholar]
- Matis, S.; Mariani, M.R.; Cutrona, G.; Cilli, M.; Piccardi, F.; Daga, A.; Damonte, G.; Millo, E.; Moroni, M.; Roncella, S.; et al. PNAEmu can significantly reduce Burkitt’s lymphoma tumor burden in a SCID mice model: Cells dissemination similar to the human disease. Cancer Gene Ther. 2009, 16, 786–793. [Google Scholar] [CrossRef]
- Cutrona, G.; Carpaneto, E.M.; Ponzanelli, A.; Ulivi, M.; Millo, E.; Scarfì, S.; Roncella, S.; Benatti, U.; Boffa, L.C.; Ferrarini, M. Inhibition of the translocated c-myc in Burkitt’s lymphoma by a PNA complementary to the E mu enhancer. Cancer Res. 2003, 63, 6144–6148. [Google Scholar]
- Cooney, M.; Czernuszewicz, G.; Postel, E.H.; Flint, S.J.; Hogan, M.E. Site-specific oligonucleotide binding represses transcription of the human c- myc gene in vitro. Science 1988, 241, 456–459. [Google Scholar] [CrossRef]
- Moreno, P.M.D.; Geny, S.; Pabon, Y.V.; Bergquist, H.; Zaghloul, E.M.; Rocha, C.S.J.; Oprea, I.I.; Bestas, B.; Andaloussi, S.E.; Jorgensen, P.T.; et al. Development of bis- locked nucleic acid (bisLNA) oligonucleotides for efficient invasion of supercoiled duplex DNA. Nucleic Acids Res. 2013, 41, 3257–3273. [Google Scholar] [CrossRef]
- Geny, S.; Moreno, P.M.D.; Krzywkowski, T.; Gissberg, O.; Andersen, N.K.; Isse, A.J.; El-Madani, A.M.; Lou, C.; Pabon, Y.V.; Anderson, B.A.; et al. Next-generation bis-locked nucleic acids with stacking linker and 2′-glycylamino-LNA show enhanced DNA invasion into supercoiled duplexes. Nucleic Acids Res. 2016, 44, 2007–2019. [Google Scholar] [CrossRef]
- Nielsen, P.E.; Egholm, M.; Berg, R.H.; Buchardt, O. Sequence-Selective Recognition of DNA by Strand Displacement with a Thymine-Substituted Polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef]
- Sharma, C.; Awasthi, S.K. Versatility of peptide nucleic acids (PNAs): Role in chemical biology, drug discovery, and origins of life. Chem. Biol. Drug Des. 2017, 89, 16–37. [Google Scholar] [CrossRef]
- Sergey, V.S.; Carla, G.S.; James, C.N.; Teresa, W.W.; David, R.C. Enhancement of strand invasion by oligonucleotides through manipulation of backbone charge. Nat. Biotechnol. 1996, 14, 1700–1704. [Google Scholar]
- Nielsen, P.E.; Egholm, M.; Buchardt, O. Peptide nucleic acid ( PNA). A DNA mimic with a peptide backbone. Bioconjug. Chem. 1994, 5, 3–7. [Google Scholar] [CrossRef]
- Nielsen, P.E.; Christensen, L. Strand displacement binding of a duplex-forming homopurine PNA to a homopyrimidine duplex DNA target. J. Am. Chem. Soc. 1996, 118, 2287–2288. [Google Scholar] [CrossRef]
- Roy, S.; Tanious, F.A.; Wilson, W.D.; Ly, D.H.; Armitage, B.A. High-affinity homologous peptide nucleic acid probes for targeting a quadruplex-forming sequence from a MYC promoter element. Biochemistry 2007, 46, 10433–10443. [Google Scholar] [CrossRef]
- Bergquist, H.; Rocha, C.S.J.; Alvarez-Asencio, R.; Nguyen, C.-H.; Rutland, M.W.; Smith, C.I.E.; Good, L.; Nielsen, P.E.; Zain, R. Disruption of Higher Order DNA Structures in Friedreich’s Ataxia (GAA)n Repeats by PNA or LNA Targeting. PLoS ONE 2016, 11, e0165788. [Google Scholar] [CrossRef]
- Zaid, A.; Sun, J.-S.; Nguyen, C.-H.; Bisagni, E.; Garestier, T.; Grierson, D.S.; Zain, R. Triple-Helix Directed Cleavage of Double-Stranded DNA by Benzoquinoquinoxaline-1,10-phenanthroline Conjugates. ChemBioChem 2004, 5, 1550–1557. [Google Scholar] [CrossRef]
- Amiri, H.; Nekhotiaeva, N.; Sun, J.-S.; Nguyen, C.-H.; Grierson, D.S.; Good, L.; Zain, R. Benzoquinoquinoxaline Derivatives Stabilize and Cleave H-DNA and Repress Transcription Downstream of a Triplex- forming Sequence. J. Mol. Biol. 2005, 351, 776–783. [Google Scholar] [CrossRef]
- Christophe, E.; Chi Hung, N.; Shrikant, K.; Yves, J.; Jian-Sheng, S.; Emile, B.; Thérèse, G.; Claude, H. Rational design of a triple helix-specific intercalating ligand. Proc. Natl. Acad. Sci. USA 1998, 95, 3591. [Google Scholar]
- Zain, R.; Marchand, C.; Sun, J.-S.; Nguyen, C.H.; Bisagni, E.; Garestier, T.; Hélène, C. Design of a triple-helix-specific cleaving reagent. Chem. Biol. 1999, 6, 771–777. [Google Scholar] [CrossRef]
- del Mundo, I.M.A.; Zewail-Foote, M.; Kerwin, S.M.; Vasquez, K.M. Alternative DNA structure formation in the mutagenic human c-MYC promoter. Nucleic Acids Res. 2017, 45, 4929–4943. [Google Scholar] [CrossRef]
- Smith, C.I.E.; Zain, R. Therapeutic Oligonucleotides: State of the Art. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 605–630. [Google Scholar] [CrossRef]
- Firulli, A.B.; Maibenco, D.C.; Kinniburgh, A.J. The identification of a tandem H-DNA structure in the c-MYC nuclease sensitive promoter element. Biochem. Biophys. Res. Commun. 1992, 185, 264–270. [Google Scholar] [CrossRef]
- Simonsson, T.; Pecinka, P.; Kubista, M. DNA tetraplex formation in the control region of c-myc. Nucleic Acids Res. 1998, 26, 1167–1172. [Google Scholar] [CrossRef]
- Adam, S.-J.; Cory, L.G.; David, J.B.; Laurence, H.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar]
- Zaghloul, E.M.; Madsen, A.S.; Moreno, P.M.D.; Oprea, I.I.; El-Andaloussi, S.; Bestas, B.; Gupta, P.; Pedersen, E.B.; Lundin, K.E.; Wengel, J.; et al. Optimizing anti-gene oligonucleotide ‘ Zorro- LNA’ for improved strand invasion into duplex DNA. Nucleic Acids Res. 2011, 39, 1142–1154. [Google Scholar] [CrossRef]
- Burmeister, T.; Molkentin, M.; Schwartz, S.; Gökbuget, N.; Hoelzer, D.; Thiel, E.; Reinhardt, R. Erroneous class switching and false VDJ recombination: Molecular dissection of t(8;14)/MYC-IGHtranslocations in Burkitt-type lymphoblastic leukemia/B-cell lymphoma. Mol. Oncol. 2013, 7, 850–858. [Google Scholar] [CrossRef]
Sample Availability: Samples of the plasmids are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | 5′GTCTCCTCCCCACCTTCGCGACCCTCCCCACCCTCCCCATAAGCGCCCCTCCCGGGTTCC3′ 3′CAGAGGAGGGGTGGAAGCGCTGGGAGGGGTGGGAGGGGTATTCGCGGGGAGGGCCCAAGG5′ |
| Pu TFO | 5′GGGAGGGGTGGGAGGGG 3′ |
| Py TFO | 3′CCCTCCCCACCCTCCCC 5′ |
| ON | Sequence 5′–3′ |
|---|---|
| Cy3-bis-m44 | Cy3-CcTtTtCtTtTtTcT-tctct-tCtTtTtTcTtTtCcCccAcgCccTctGc |
| Cy3-bis-m44-M3 | Cy3-CcTtTtCtTtTtTcT-M3-tCtTtTtTcTtTtCcCccAcgCccTctGc |
| WC-m44 | tCtTtTtTcTtTtCcCccAcgCccTctGc |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umek, T.; Sollander, K.; Bergquist, H.; Wengel, J.; Lundin, K.E.; Smith, C.I.E.; Zain, R. Oligonucleotide Binding to Non-B-DNA in MYC. Molecules 2019, 24, 1000. https://doi.org/10.3390/molecules24051000
Umek T, Sollander K, Bergquist H, Wengel J, Lundin KE, Smith CIE, Zain R. Oligonucleotide Binding to Non-B-DNA in MYC. Molecules. 2019; 24(5):1000. https://doi.org/10.3390/molecules24051000
Chicago/Turabian StyleUmek, Tea, Karin Sollander, Helen Bergquist, Jesper Wengel, Karin E. Lundin, C.I. Edvard Smith, and Rula Zain. 2019. "Oligonucleotide Binding to Non-B-DNA in MYC" Molecules 24, no. 5: 1000. https://doi.org/10.3390/molecules24051000
APA StyleUmek, T., Sollander, K., Bergquist, H., Wengel, J., Lundin, K. E., Smith, C. I. E., & Zain, R. (2019). Oligonucleotide Binding to Non-B-DNA in MYC. Molecules, 24(5), 1000. https://doi.org/10.3390/molecules24051000