
Bark Extract of the Amazonian Tree Endopleura uchi (Humiriaceae) Extends Lifespan and Enhances Stress Resistance in Caenorhabditis elegans

 , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Extract
2.2. Antioxidant Activity
2.3. Total Phenolic Content
2.4. Chemical Characterization and Quantification of Bergenin
2.5. C. elegans Strains and Maintenance
2.6. Survival Assay under Oxidative Stress
2.7. Intracellular ROS Accumulation
2.8. Quantification of Gene Expression Using GFP Reporter
2.9. Longevity Assay
2.10. Quantification of PolyQ40::GFP Aggregate Formation
2.11. Pharyngeal Pumping Rate
2.12. Body Length
2.13. Antimicrobial Activity
3. Results
3.1. Antioxidant Activity in Vitro and Chemical Characterization of the Bark Extract
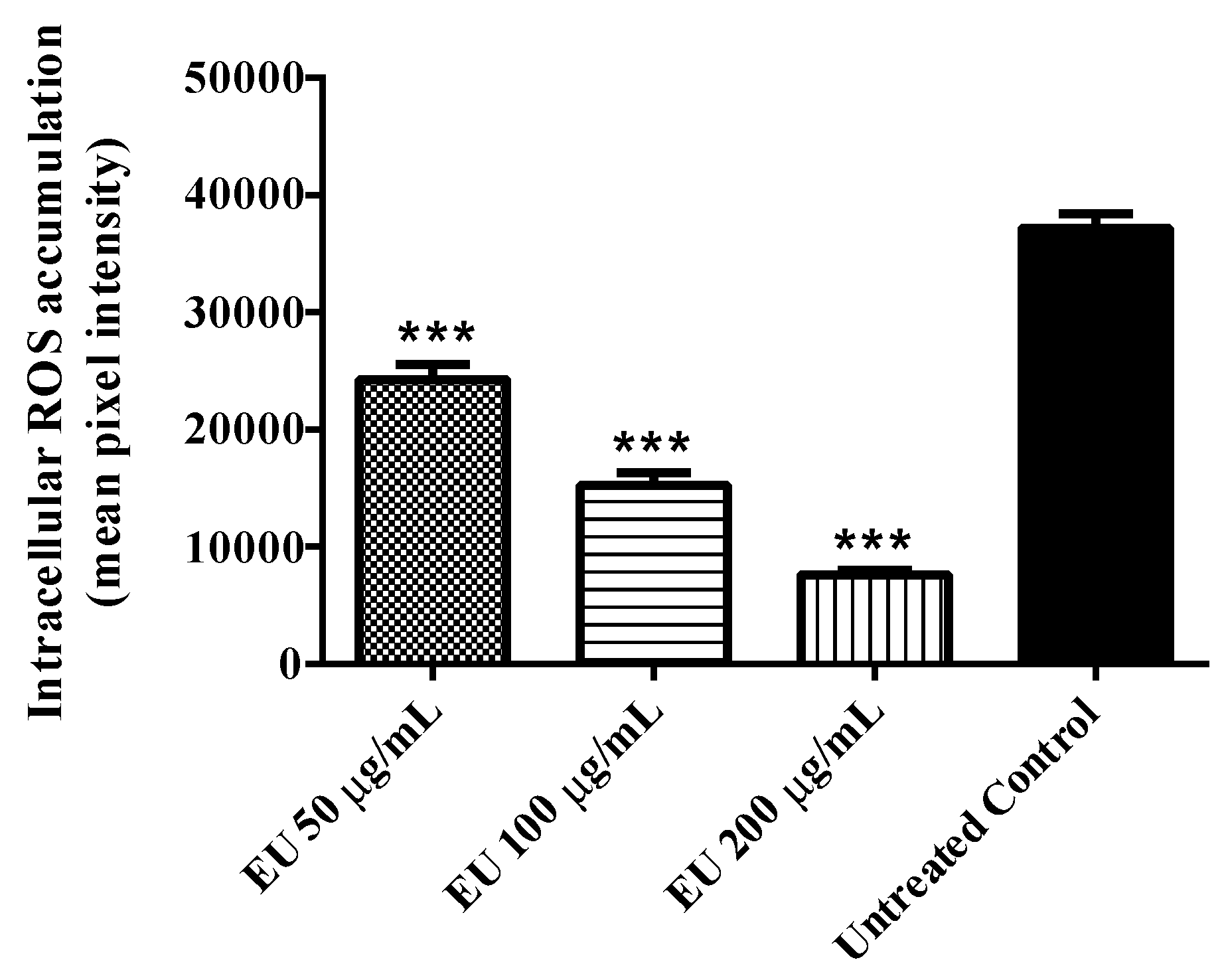
3.2. Effect of the Extract on Intracellular ROS Accumulation
3.3. Protection against Oxidative Stress
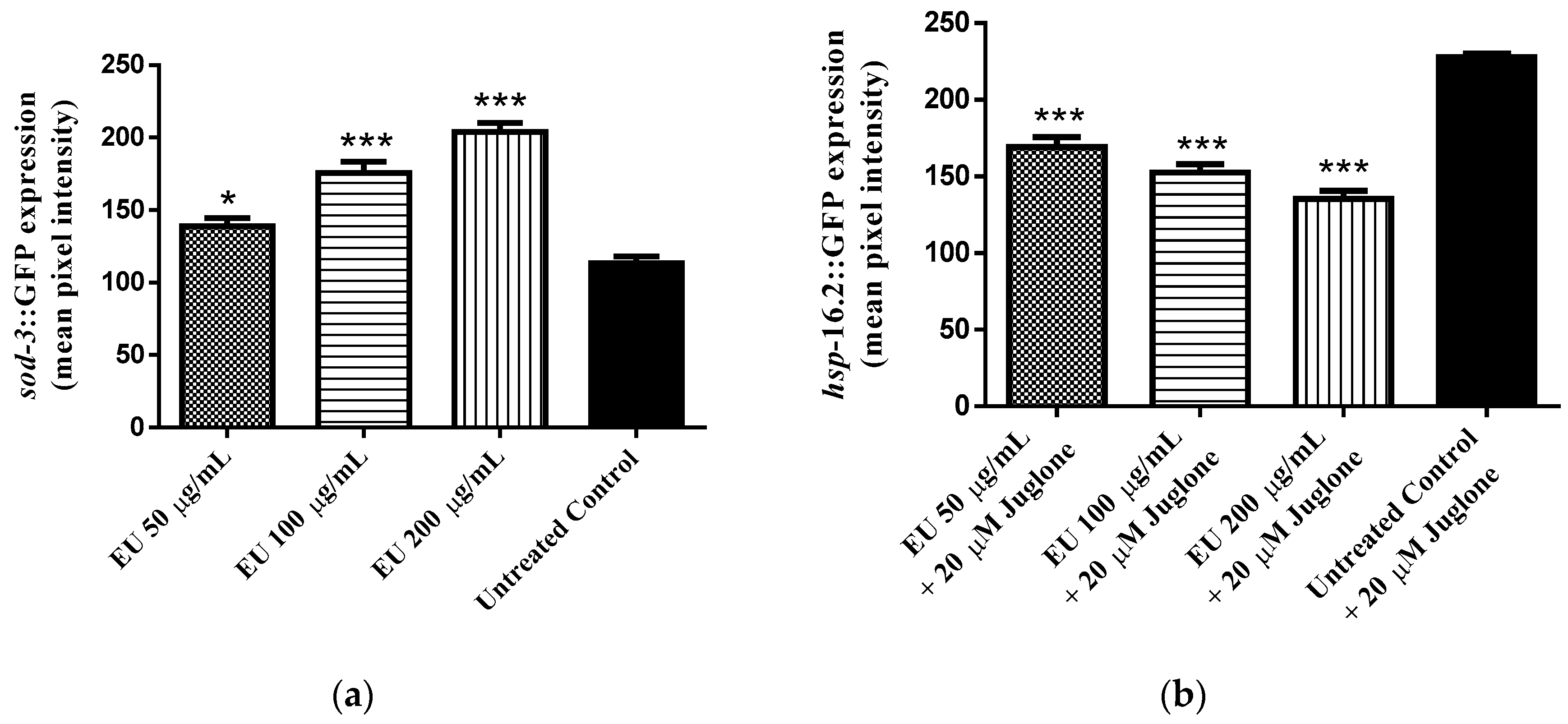
3.4. Effect of the Extract on the Expression of Stress Response Genes (hsp-16.2::GFP and sod-3::GFP)
3.5. Effect of the Extract on Longevity
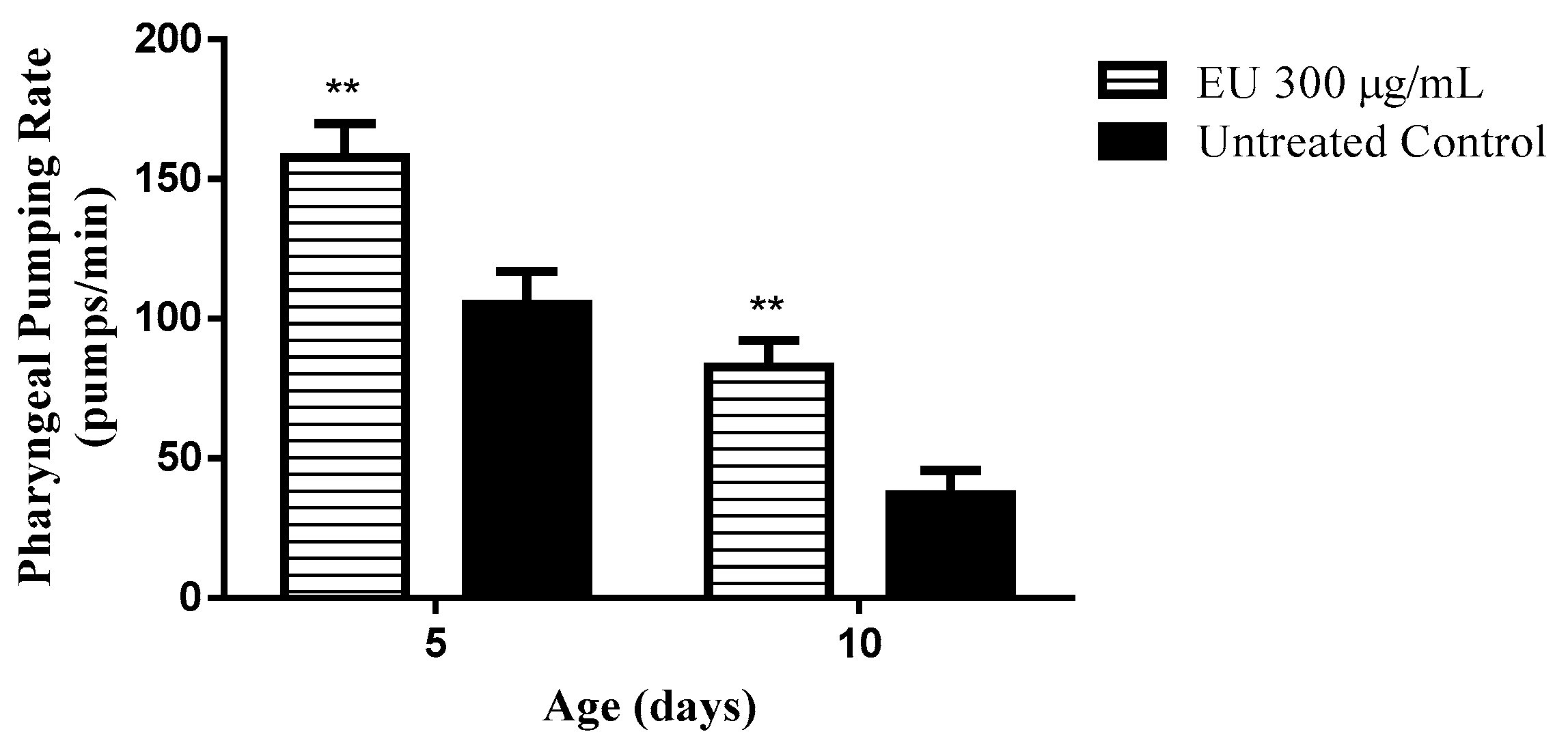
3.6. Effect of the Extract on the Pharyngeal Pumping Rate
3.7. Body Length
3.8. Antimicrobial Activity
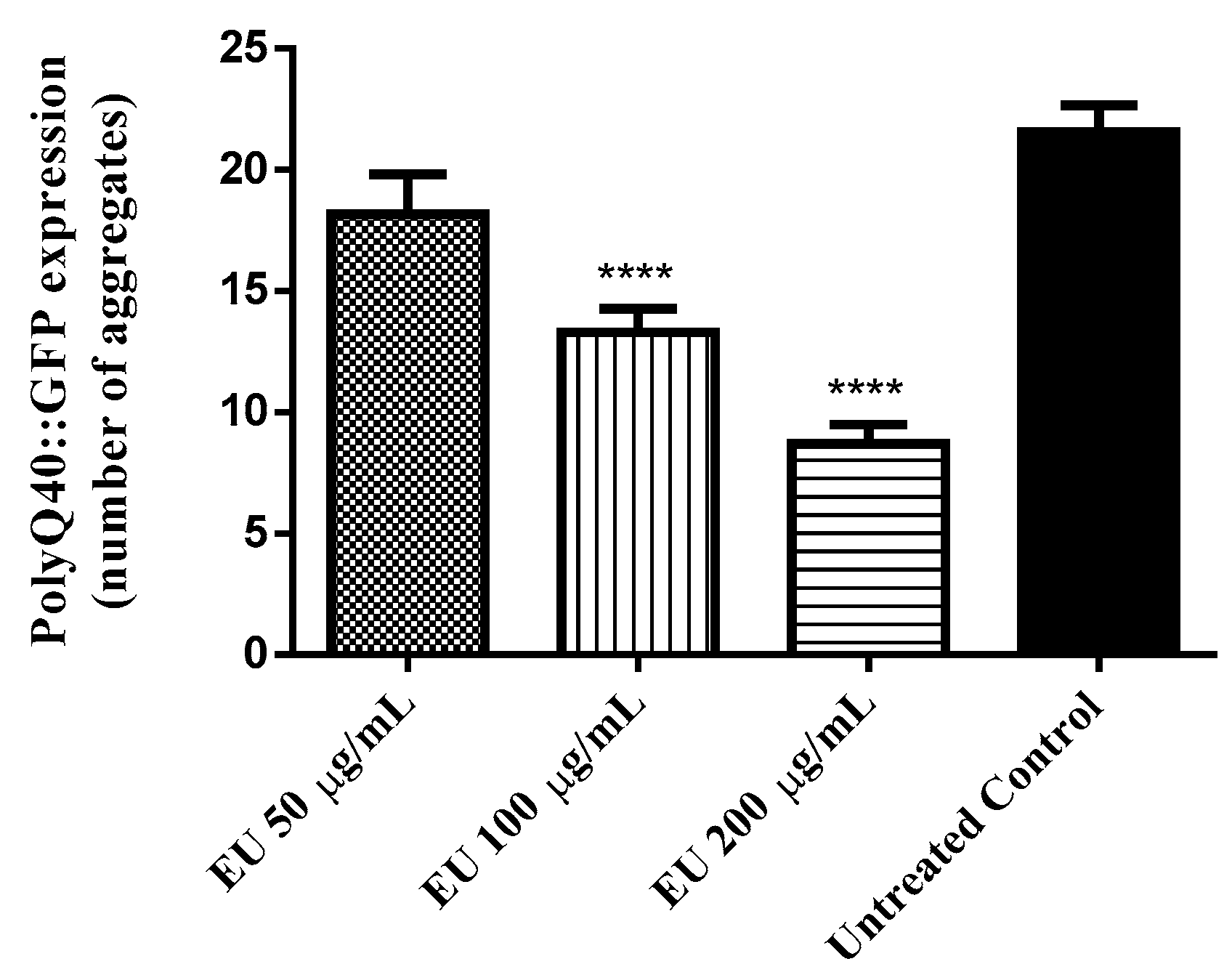
3.9. Effect of the Bark Extract on the Formation of polyQ40 Aggregates
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Food and Fruit-Bearing Forest Species, 3: Examples from Latin America/F2911; Food and Agriculture Organization: Rome, Italy, 1986; p. 44.
- Shanley, P. Fruit Trees and Useful Plants in Amazonian Life; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2011. [Google Scholar]
- Correa, M.; Penna, L.d.A. Dicionario das plantas uteis do Brasil e das exoticas cultivadas: Volume 5. MR. Rio de Janeiro: Instituto Brasiliero de Desenvolvimento Florestal 687p.-illus. Por Icones. Geog 1974, 4. [Google Scholar]
- Rodrigues, E. Plants and animals utilized as medicines in the Jaú National Park (JNP), Brazilian Amazon. Phytother. Res. 2006, 20, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Scudeller, V.V.; Veiga, J.B.d.; Araújo-Jorge, L.d. Etnoconhecimento de plantas de uso medicinal nas comunidades São João do Tupé e Central (Reserva de Desenvolvimento Sustentável do Tupé); UEA: Manaus, Brazil, 2009; pp. 185–199. [Google Scholar]
- Pinto, A.Z.d.L.; De Assis, A.F.S.; Pereira, A.G.; Pasa, M.C. Levantamento etnobotânico de plantas medicinais comercializadas no mercado do porto em Cuiabá, Mato Grosso, Brasil. FLOVET-Boletim do Grupo de Pesquisa da Flora, Vegetação e Etnobotânica 2013, 1. [Google Scholar]
- Caetano, R.S.; de Souza, A.C.R.; Feitoza, L.F. O Uso de plantas medicinais utilizadas por frequentadores dos ambulatórios Santa Marcelina, Porto Velho-RO. Saúde e Pesquisa 2014, 7. [Google Scholar]
- Silva, L.R.; Teixeira, R. Phenolic profile and biological potential of Endopleura uchi extracts. Asian Pac. J. Trop. Med. 2015, 8, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Bento, J.F. Caracterização de polissacarídeos e metabólicos secundários da casca de Endopleura uchi (Huber) cuatrec: Avaliação dos efeitos do decocto e de frações polissacarídicas em células hela e macrófagos. Ph.D. Thesis, Federal University of Paraná, Curitiba, PR, Brazil, November 2013. [Google Scholar]
- Sá, B.M.; Lima, C.S.; Silva, U.D.A.; Carvalho, H.O.; Fernandes, C.P.; Resque, R.L.; de Oliveria, T.T.; Carvalho, J.C.T. Subchronic toxicity evaluation of the hydroethanolic extract from Endopleura uchi (Huber) Cuatrec in Wistar rats. Afr. J. Pharm. Pharmacol. 2015, 9, 223–229. [Google Scholar]
- Politi, F.A.S.; Moreira, R.R.D.; Salgado, H.R.N.; Pietro, R. Preliminary tests on acute oral toxicity and intestinal motility with extract of pulverized bark of Endopleura uchi (Huber) Cuatrec. (Humiriaceae) in mice. Rev. Pan-Amaz. Saude 2010, 1, 187–189. [Google Scholar] [CrossRef]
- Luna, J.; Silva, T.; Bento, E.; Santana, A. Isolamento e identificação estrutural dos constituintes químicos de Endopleura uchi (Humiriaceae). Reunião Anual da Sociedade Brasileira de Química, Minas Gerais, Brasil 2000, 2, 123. [Google Scholar]
- Politi, F.A.S. Estudos farmacognósticos e avaliação de atividades biológicas de extratos obtidos das cascas pulverizadas de Endopleura uchi (Huber) Cuatrec. (Humiriaceae). Master’s Thesis, Universidade Estadual Paulista, Araraquara, SP, Brazil, February 2009. [Google Scholar]
- Abreu, V.G.d.C.; Corrêa, G.M.; Lagos, I.A.d.S.; Silva, R.R.; Alcântara, A.F.d.C. Pentacyclic triterpenes and steroids from the stem bark of uchi (Sacoglottis uchi, Humiriaceae). Acta Amazonica 2013, 43, 525–528. [Google Scholar] [CrossRef]
- De Abreu, H.A.; Lago, I.A.d.S.; Souza, G.P.; Piló-Veloso, D.; Duarte, H.A.; Alcântara, A.F.d.C. Antioxidant activity of (+)-bergenin—a phytoconstituent isolated from the bark of Sacoglottis uchi Huber (Humireaceae). Org. Biomol. Chem. 2008, 6, 2713–2718. [Google Scholar] [CrossRef] [PubMed]
- Borges, J.C.; Ripardo Filho, H.d.S.; Guilhon, G.; Carvalho, J.C.; Santos, L.S.; Sousa, P.J. Antinociceptive activity of acetylbergenin in mice. Lat. Am. J. Pharm. 2011, 30, 1303–1308. [Google Scholar]
- Tacon, L.A.; Freitas, L.A. Box-Behnken design to study the bergenin content and antioxidant activity of Endopleura uchi bark extracts obtained by dynamic maceration. Revista Brasileira de Farmacognosia 2013, 23, 65–71. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958. [Google Scholar] [CrossRef]
- Nunomura, R.; Oliveira, V.G.; Da Silva, S.L.; Nunomura, S.M. Characterization of bergenin in Endopleura uchi bark and its anti-inflammatory activity. J. Braz. Chem. Soc. 2009, 20, 1060–1064. [Google Scholar] [CrossRef]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An anthocyanin-rich extract of acai (Euterpe precatoria Mart.) increases stress resistance and retards aging-related markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2005. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Röhrig, T.; Richling, E.; Wang, X.; Wink, M. Anti-Aging and antioxidant potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans indicate a new utilization for roasted seeds of Guarana. Medicines 2017, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Koolen, H.; da Silva, F.; Silva, E.; Braun, M.S.; Wang, X.; Wink, M.J.M. Calycophyllum spruceanum (Benth.), the amazonian “tree of youth” prolongs longevity and enhances stress resistance in Caenorhabditis elegans. Molecules 2018, 23, 534. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.L.; El-Readi, M.Z.; Hamoud, R.; Eid, S.Y.; El Ahmady, S.H.; Nibret, E.; Herrmann, F.; Youns, M.; Tahrani, A.; Kaufmann, D. Anti-infective and cytotoxic properties of Bupleurum marginatum. Chin. Med. 2014, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Piacente, S.; Pizza, C.; De Tommasi, N.; Mahmood, N. Constituents of Ardisia japonica and their in vitro anti-HIV activity. J. Nat. Prod. 1996, 59, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Goel, R.; Maiti, R.; Manickam, M.; Ray, A. Antiulcer activity of naturally occurring pyrano-coumarin and isocoumarins and their effect on prostanoid synthesis using human colonic mucosa. Ind. J. Exp. Biol. 1997, 35, 1080–1083. [Google Scholar]
- Takahashi, H.; Kosaka, M.; Watanabe, Y.; Nakade, K.; Fukuyama, Y. Synthesis and neuroprotective activity of bergenin derivatives with antioxidant activity. Bioorg. Med. Chem. 2003, 11, 1781–1788. [Google Scholar] [CrossRef]
- Lim, H.-K.; Kim, H.-S.; Choi, H.-S.; Oh, S.; Choi, J. Hepatoprotective effects of bergenin, a major constituent of Mallotus japonicus, on carbon tetrachloride-intoxicated rats. J. Ethnopharmacol. 2000, 72, 469–474. [Google Scholar] [CrossRef]
- Nazir, N.; Koul, S.; Qurishi, M.A.; Taneja, S.C.; Ahmad, S.F.; Bani, S.; Qazi, G.N. Immunomodulatory effect of bergenin and norbergenin against adjuvant-induced arthritis—A flow cytometric study. J. Ethnopharmacol. 2007, 112, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Aithal, B.K.; Kumar, M.S.; Rao, B.N.; Udupa, N.; Rao, B.S. Juglone, a naphthoquinone from walnut, exerts cytotoxic and genotoxic effects against cultured melanoma tumor cells. Cell Biol. Int. 2009, 33, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein HSP 16-2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J. 2003, 17, 2305–2307. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R. Heat shock proteins in long-lived worms and mice with insulin/insulin-like signaling mutations. Aging 2009, 1, 573. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Braeckman, B.P.; Vanfleteren, J.R. Genetic control of longevity in C. elegans. Exp. Gerontol. 2007, 42, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Oh, S.W.; Tissenbaum, H.A. Worming pathways to and from DAF-16/FOXO. Exp. Gerontol. 2006, 41, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Wink, M. Epigallocatechin gallate from green tea (Camellia sinensis) increases lifespan and stress resistance in Caenorhabditis elegans. Planta Med. 2009, 75, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Kennedy, B.K. Does longer lifespan mean longer healthspan? Trends Cell Biol. 2016, 26, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Zhu, L.J.; Yen, K.; Tissenbaum, H.A. Uncoupling lifespan and healthspan in Caenorhabditis elegans longevity mutants. Proc. Natl. Acad. Sci. USA 2015, 112, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Kong, Y.; Zhang, H. Oxidative stress, mitochondrial dysfunction, and aging. J. Signal Transduct. 2011, 2012. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Krstin, S.; Wang, X.; Wink, M. Anthocyanin-rich extract of Acai (Euterpe precatoria Mart.) mediates neuroprotective activities in Caenorhabditis elegans. J. Funct. Foods 2016, 26, 385–393. [Google Scholar] [CrossRef]
- Collins, J.J.; Huang, C.; Hughes, S.; Kornfeld, K. The measurement and analysis of age-related changes in Caenorhabditis elegans. WormBook 2005. [Google Scholar] [CrossRef]
- Anton, S.D.; Woods, A.J.; Ashizawa, T.; Barb, D.; Buford, T.W.; Carter, C.S.; Clark, D.J.; Cohen, R.A.; Corbett, D.B.; Cruz-Almeida, Y.; et al. Successful aging: Advancing the science of physical independence in older adults. Ageing Res. Rev. 2015, 24, 304–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span—from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S. Huntington’s disease. Cold Spring Harb Perspect. Biol. 2011, 3, 7476. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Dillin, A. The insulin paradox: Aging, proteotoxicity and neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 759–767. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAMPLE | EC50 (µG/ML) |
|---|---|
| EU | 8.0 |
| EGCG | 1.2 |
| VITAMIN C | 2.1 |
| Ampicillin (256 µg/mL) | Ciprofloxacin (256 µg/mL) | EU (10 mg/mL) | |
|---|---|---|---|
| E. coli OP50 | 27.3 ± 1.2 * | 42.8 ± 0.3 * | NI |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, H.; Roxo, M.; Silva, E.; Valente, K.; Braun, M.; Wang, X.; Wink, M. Bark Extract of the Amazonian Tree Endopleura uchi (Humiriaceae) Extends Lifespan and Enhances Stress Resistance in Caenorhabditis elegans. Molecules 2019, 24, 915. https://doi.org/10.3390/molecules24050915
Peixoto H, Roxo M, Silva E, Valente K, Braun M, Wang X, Wink M. Bark Extract of the Amazonian Tree Endopleura uchi (Humiriaceae) Extends Lifespan and Enhances Stress Resistance in Caenorhabditis elegans. Molecules. 2019; 24(5):915. https://doi.org/10.3390/molecules24050915
Chicago/Turabian StylePeixoto, Herbenya, Mariana Roxo, Emerson Silva, Karla Valente, Markus Braun, Xiaojuan Wang, and Michael Wink. 2019. "Bark Extract of the Amazonian Tree Endopleura uchi (Humiriaceae) Extends Lifespan and Enhances Stress Resistance in Caenorhabditis elegans" Molecules 24, no. 5: 915. https://doi.org/10.3390/molecules24050915