Isolation and Characterization of a Novel Sialoglycopeptide Promoting Osteogenesis from Gadus morhua Eggs

 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of Gm-SGPP
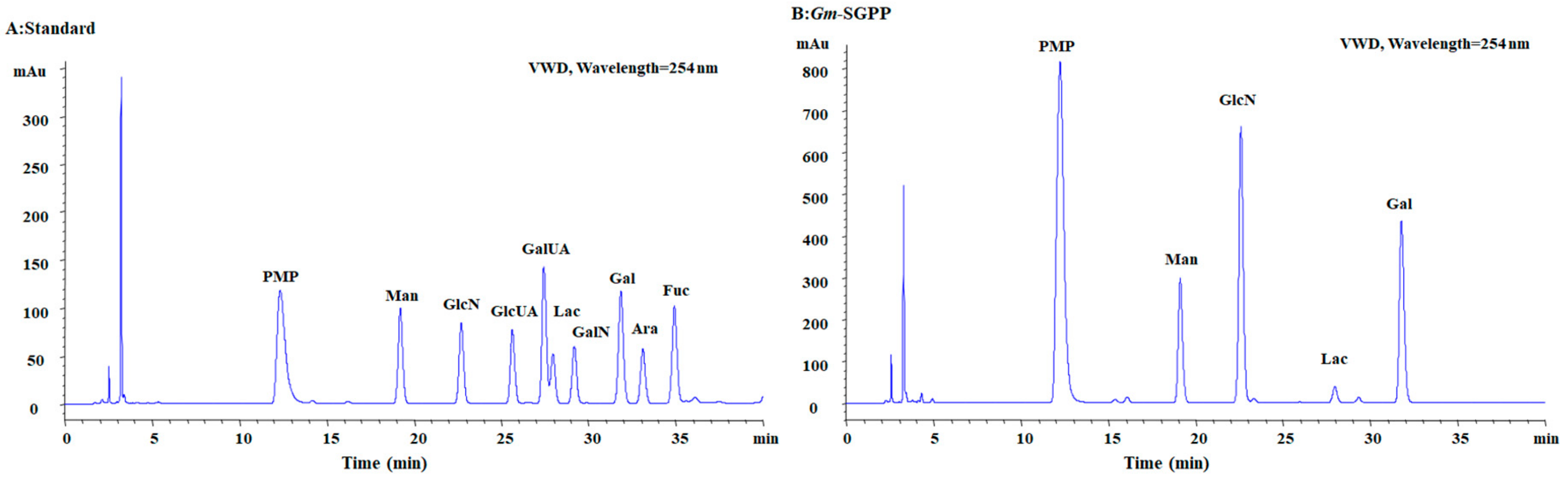
2.2. Chemical Composition
2.3. Determination of Glycosidic Bond Type
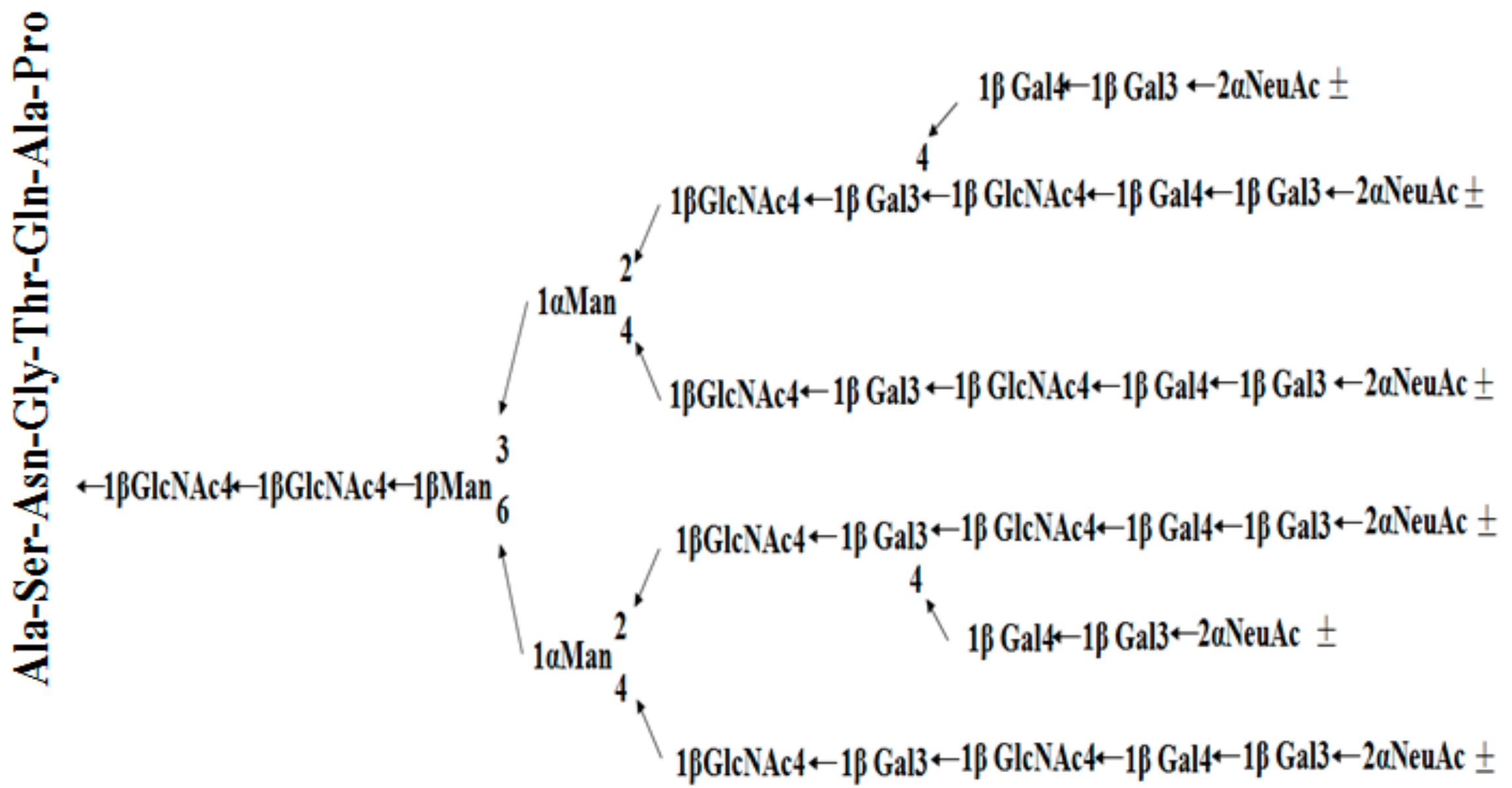
2.4. Amino Acid Composition and Sequence Analysis of Gm-SGPP
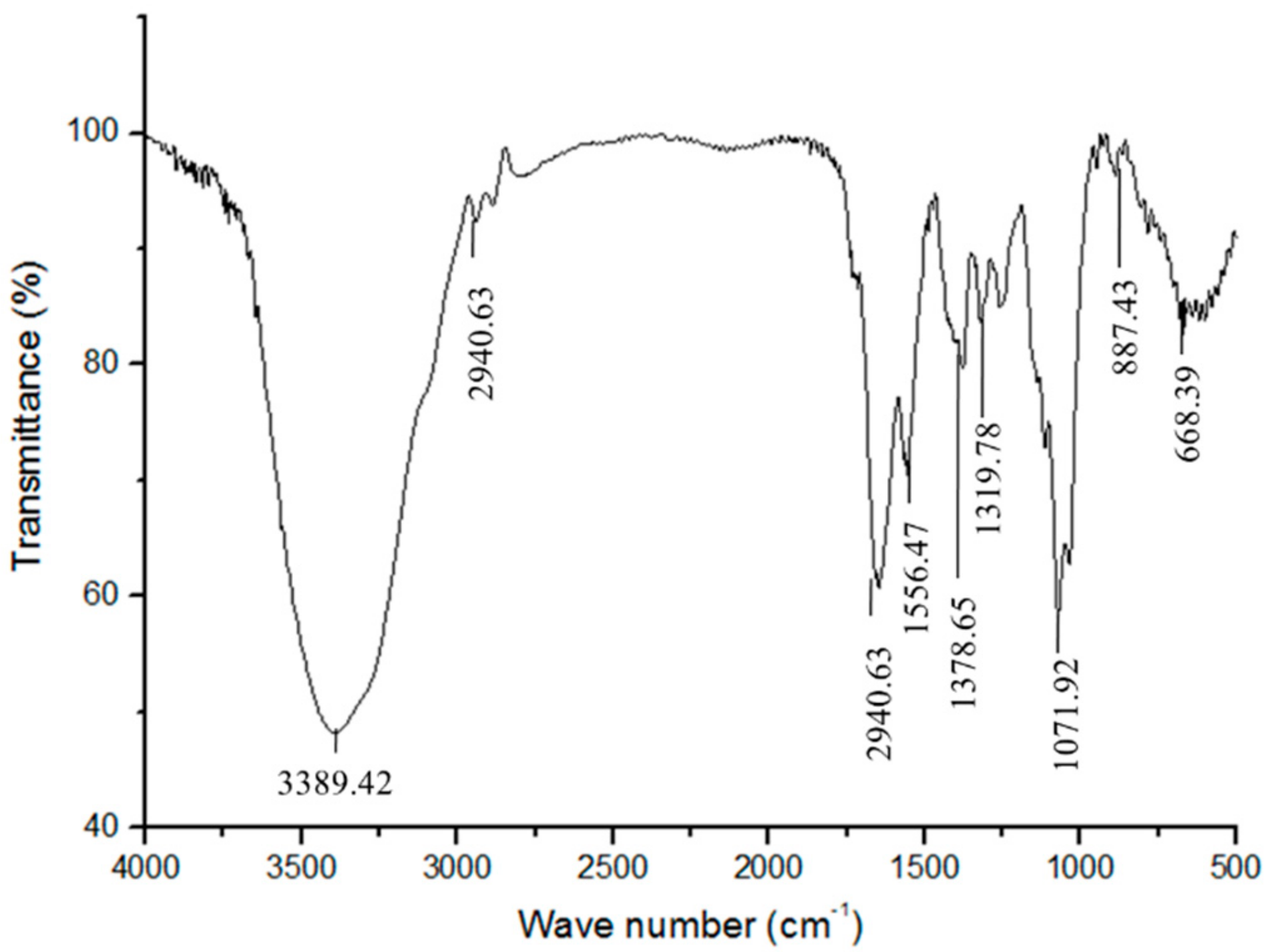
2.5. Fourier Transform-Infrared (FT-IR) Spectroscopy Analysis of Gm-SGPP
2.6. Methylation Analysis of Gm-SGPP
2.7. H-Nuclear Magnetic Resonance (1H-NMR) Spectroscopy of Gm-SGPP
2.8. Gm-SGPP Promoted MC3T3-E1 Cell Proliferation
2.9. Gm-SGPP Promoted Bone Formation in Adolescent Mice
3. Methods
3.1. Materials
3.2. Preparation of Gm-SGPP
3.3. Identification of Gm-SGPP
3.4. Chemical Composition of Gm-SGPP
3.5. Determination of Gm-SGPP Glycosylation Type
3.6. FT-IR Spectroscopy Analysis of Gm-SGPP
3.7. Amino Acid Composition Analysis of Gm-SGPP
3.8. Amino Acid Sequence Analysis of Gm-SGPP
3.9. Methylation Analysis of Gm-SGPP
3.10. 1H-NMR Spectroscopy of Gm-SGPP
3.11. MTT Cell Viability Assay
3.12. Animal Treatments
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Q.; Xue, C.H.; Li, Z.J.; Xu, J. Phosphatidylcholine levels and their fatty acid compositions in squid egg: A comparison study with pollack roe and sturgeon caviar. J. Food Lipids 2008, 15, 222–230. [Google Scholar] [CrossRef]
- Xia, G.; Wang, J.; Sun, S.; Zhao, Y.; Wang, Y.; Yu, Z.; Wang, S.; Xue, C. Sialoglycoproteins prepared from the eggs of Carassius auratus prevent bone loss by inhibiting the NF-kappaB pathway in ovariectomized rats. Food Funct. 2016, 7, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Della Giustina, A.; Goldim, M.P.; Danielski, L.G.; Florentino, D.; Garbossa, L.; Joaquim, L.; Oliveira Junior, A.N.; Mathias, K.; Fileti, M.E.; Zarbato, G.F.; et al. Fish oil–rich lipid emulsion modulates neuroinflammation and prevents long-term cognitive dysfunction after sepsis. Nutrition 2018, 110417. [Google Scholar] [CrossRef] [PubMed]
- Skórkowska-Telichowska, K.; Kosińska, J.; Chwojnicka, M.; Tuchendler, D.; Tabin, M.; Tuchendler, R.; Bobak, Ł.; Trziszka, T.; Szuba, A. Positive effects of egg-derived phospholipids in patients with metabolic syndrome. Adv. Med. Sci. Pol. 2016, 61, 169–174. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, S.; Leem, K.H. Protective effect of egg yolk peptide on bone metabolism. Menopause 2011, 18, 307–313. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, L.; Zhang, J.; Xiao, J.; Wei, X. Study on the purification and characterization of a polysaccharide conjugate from tea flowers. Int. J. Blol. Macromol. 2010, 47, 266–270. [Google Scholar] [CrossRef]
- Zhang, G.; Han, L.; Zhang, G.; Zhang, X.; Feng, J. Purification and characterization of a novel glycoprotein from Streptomyces sp. ZX01. Int. J. Blol. Macromol. 2015, 78, 195–201. [Google Scholar] [CrossRef]
- Velkova, L.; Voelter, W.; Dolashki, A.; Dolashka, P. Structure and antibacterial activity of isolated peptides from the mucus of garden snail Cornu aspersum. Bulg. Chem. Commun. 2018, 50, 195–200. [Google Scholar]
- Dolashki, A.; Dolashka, P.; Stevanovic, S.; Molin, L.; Traldi, P.; Velkova, L.; Van Beeumen, J.; Devreese, B.; Voelter, W. Antimicrobial Activity of Peptides Isolated from the Hemolymph of Helix Lucorum Snails. J. Pept. Sci. 2014, 20, S268–S269. [Google Scholar]
- Yu, J.; Li, X.M.; Li, H.M.; Zhang, Q.H.; Li, X.Q.; Shao, S.L. Analysis of Glycosylation Sites and Glycan Structure of Glycoprotein by Multiple Mass Spectrometric Techniques. Chinese J. Anal. Chem. 2015, 43, 536–541. [Google Scholar]
- Velkova, L.; Dolashka, P.; Beeumen, J.V.; Devreese, B. N-glycan structures of β-HH subunit of Helix lucorum hemocyanin. Carbohyd. Res. 2017, 488, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Xue, C.; Chang, Y.; Hu, Y.; Xu, X.; Ge, L.; Liu, G. Structure and rheological characteristics of fucoidan from sea cucumber Apostichopus japonicus. Food Chem. 2015, 180, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Hurum, D.C.; Rohrer, J.S. Five-minute glycoprotein sialic acid determination by high-performance anion exchange chromatography with pulsed amperometric detection. Anal. Biochem. 2011, 419, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Barrabes, S.; Sarrats, A.; Fort, E.; De Llorens, R.; Rudd, P.M.; Peracaula, R. Effect of sialic acid content on glycoprotein pI analyzed by two-dimensional electrophoresis. Electrophoresis 2010, 31, 2903–2912. [Google Scholar] [CrossRef]
- Nasir ud, D.; Hoessli, D.C.; Rungger-Brändle, E.; Hussain, S.A.; Walker-Nasir, E. Role of sialic acid and sulfate groups in cervical mucus physiological functions: Study of Macaca radiata glycoproteins. BBA Gen. Subj. 2003, 1623, 53–61. [Google Scholar] [CrossRef]
- Ricci, G.; Ambrosi, A.D.; Resca, D.; Masotti, M.; Alvisi, V. Comparison of serum total sialic acid, C-reactive protein, α1-acid glycoprotein and β2-microglobulin in patients with non-malignant bowel diseases. Biomed. Pharmacother 1995, 49, 259–262. [Google Scholar] [CrossRef]
- Inoue, S.; Iwasaki, M. Isolation of a novel glycoprotein from the eggs of rainbow trout: Occurrence of disialosyl groups on all carbohydrate chains. Biochem. Biophys. Res. Commun. 1978, 83, 1018–2103. [Google Scholar] [CrossRef]
- Inoue, S.; Iwasaki, M. Sialoglycoproteins from the eggs of Pacific herring. Isolation and characterization. Eur. J. Biochem. 1980, 111, 131–135. [Google Scholar] [CrossRef]
- Inoue, S.; Kitajima, K.; Inoue, Y.; Kudo, S. Localization of polysialoglycoprotein as a major glycoprotein component in cortical alveoli of the unfertilized eggs of Salmo gairdneri. Dev. Biol. 1987, 123, 442–454. [Google Scholar] [CrossRef]
- Kitajima, K.; Inoue, Y.; Inoue, S. Polysialoglycoproteins of Salmonidae fish eggs. Complete structure of 200-kDa polysialoglycoprotein from the unfertilized eggs of rainbow trout (Salmo gairdneri). J. Biol. Chem. 1986, 261, 5262–5269. [Google Scholar]
- Seko, A.; Kitajima, K.; Iwasaki, M.; Inoue, S.; Inoue, Y. Structural studies of fertilization-associated carbohydrate-rich glycoproteins (hyosophorin) isolated from the fertilized and unfertilized eggs of flounder, Paralichthys olivaceus. Presence of a novel penta-antennary N-linked glycan chain in the tandem repeating glycopeptide unit of hyosophorin. J. Biol. Chem. 1989, 264, 1592–15929. [Google Scholar]
- Xia, G.H.; Wang, S.S.; He, M.; Zhou, X.C.; Zhao, Y.L.; Wang, J.F.; Xue, C.H. Anti-osteoporotic activity of sialoglycoproteins isolated from the eggs of Carassius auratus by promoting osteogenesis and increasing OPG/RANKL ratio. J. Funct. Foods. 2015, 15, 137–150. [Google Scholar] [CrossRef]
- Leem, K.H.; Kim, M.G.; Kim, H.M.; Kim, M.; Lee, Y.J.; Kim, H.K. Effects of egg yolk proteins on the longitudinal bone growth of adolescent male rats. Biosci. Biotechnol. Biochem. 2004, 68, 2388–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seko, A.; Koketsu, M.; Nishizono, M.; Enoki, Y.; Ibrahim, H.R.; Juneja, L.R.; Kim, M.; Yamamoto, T. Occurrence of a sialylglycopeptide and free sialylglycans in hen’s egg yolk. BBA Gen. Subj. 1997, 1335, 23–32. [Google Scholar] [CrossRef]
- Iwasaki, M.; Seko, A.; Kitajima, K.; Inoue, Y.; Inoue, S. Fish Egg Glycophosphoproteins Have Species-Specific N-Linked Glycan Units Previously Found in a Storage Pool of Free Glycan Chains. J. Biol. Chem. 1992, 267, 24287–24296. [Google Scholar]
- Kitajima, K.; Inoue, S.; Inoue, Y. Isolation and characterization of a novel type of sialoglycoproteins (hyosophorin) from the eggs of medaka, Oryzias latipes: Nonapeptide with a large N-linked glycan chain as a tandem repeat unit. Dev. Biol. 1989, 132, 544–553. [Google Scholar] [CrossRef]
- Cui, F.J.; Zan, X.Y.; Li, Y.H.; Yang, Y.; Sun, W.J.; Zhou, Q.; Yu, S.L.; Dong, Y. Purification and partial characterization of a novel anti-tumor glycoprotein from cultured mycelia of Grifola frondosa. Int. J. Blol. Macromol. 2013, 62, 684–690. [Google Scholar] [CrossRef]
- Cui, F.J.; Li, Y.H.; Zan, X.Y.; Yang, Y.; Sun, W.J.; Qian, J.Y.; Zhou, Q.; Yu, S.L. Purification and partial characterization of a novel hemagglutinating glycoprotein from the cultured mycelia of Hericium erinaceus. Process Biochem. 2014, 49, 1362–1369. [Google Scholar] [CrossRef]
- Du, X.J.; Zhang, Y.; Mu, H.M.; Lv, Z.W.; Yang, Y.; Zhang, J.S. Structural elucidation and antioxidant activity of a novel polysaccharide (TAPB1) from Tremella aurantialba. Food Hydrocoll. 2015, 43, 459–464. [Google Scholar] [CrossRef]
- Heiss, C.; Wang, Z.; Black, I.; Azadi, P.; Fichorova, R.N.; Singh, B.N. Novel structural features of the immunocompetent ceramide phospho-inositol glycan core from Trichomonas vaginalis. Carbohyd. Res. 2016, 419, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Yang, Y.M.; Sanchez, S.; Cui, D.C.; Dang, R.J.; Wang, X.Y.; Lin, Q.X.; Wang, Y.; Wang, C.Y.; Chen, D.F.; et al. Deubiquitinase MYSM1 Is Essential for Normal Bone Formation and Mesenchymal Stem Cell Differentiation. Sci. Rep. 2016, 6, 22211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, M.; Leem, K.H.; Kim, M.; Kim, H.K. Egg yolk soluble protein stimulates the proliferation and differentiation of osteoblastic MC3T3-E1 cells. Biosci. Biotech. Biochem. 2007, 71, 1327–1329. [Google Scholar] [CrossRef] [PubMed]
- Fairbanks, G.; Steck, T.L.; Wallach, D.F. Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 1971, 10, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, S. Purification and characterization of a putative vitellogenin from the ovary of amphioxus (Branchiostoma belcheri tsingtaunese). Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2001, 129, 121–127. [Google Scholar] [CrossRef]
- De Vlaming, V.L.; Vodicnik, M.J.; Bauer, G.; Murphy, T.; Evans, D. Estradiol-17 beta effects on lipid and carbohydrate metabolism and on the induction of a yolk precursor in goldfish, Carassius auratus. Life Sci. 1977, 20, 1945–1952. [Google Scholar] [CrossRef]
- Wang, X.; Hwang, S.Y.; Cong, W.T.; Jin, L.T.; Choi, J.K. Phosphoprotein staining for sodium dodecyl sulfate-polyacrylamide gel electrophoresis using fluorescent reagent morin hydrate. Anal. Biochem. 2013, 435, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Anumula, K.R. Rapid quantitative determination of sialic acids in glycoproteins by high-performance liquid chromatography with a sensitive fluorescence detection. Anal. Biochem. 1995, 230, 24–30. [Google Scholar] [CrossRef]
- Dunham, N.T.; Kane, E.E.; Rodriguez-Saona, L.E. Quantifying soluble carbohydrates in tropical leaves using a portable mid-infrared sensor: Implications for primate feeding ecology. Am. J. Primatol. 2016, 78, 701–706. [Google Scholar] [CrossRef]
- Zhou, G.F.; Pang, M.X.; Chen, S.H.; Lv, G.Y.; Yan, M.Q. Comparison on polysaccharide content and PMP-HPLC fingerprints of polysaccharide in stems and leaves of Dendrobium officinale. China J. Chin. Mater. Med. 2014, 39, 795–802. [Google Scholar]
- Ke, L.J.; Lin, D.Y.; Huang, X.N.; Huo, Y.S.; Rao, P.F.; Ye, X.Y. Comparison of protein composition and activities of pilose antler processed by different methods. J. Chin. Med. Mater. 2008, 31, 11–14. [Google Scholar]
- Wu, J.S.; Zheng, J.; Xia, X.J.; Kan, J.Q. Purification and Structural Identification of Polysaccharides from Bamboo Shoots (Dendrocalamus latiflorus). Int. J. Mol. Sci. 2015, 16, 15560–15577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edge, A.S.; Faltynek, C.R.; Hof, L.; Reichert, L.E., Jr.; Weber, P. Deglycosylation of glycoproteins by trifluoromethanesulfonic acid. Anal. Biochem. 1981, 118, 131–137. [Google Scholar] [CrossRef]
- Hakomori, S. A Rapid Permethylation of Glycolipid, and Polysaccharide Catalyzed by Methylsulfinyl Carbanion in Dimethyl Sulfoxide. J. Biochem. 1964, 55, 205–208. [Google Scholar] [PubMed]
- Ye, L.; Zhang, J.; Ye, X.; Tang, Q.; Liu, Y.; Gong, C.; Du, X.; Pan, Y. Structural elucidation of the polysaccharide moiety of a glycopeptide (GLPCW-II) from Ganoderma lucidum fruiting bodies. Carbohyd. Res. 2008, 343, 746–752. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Gm-SGPP (mg/g glycopeptide) | Amino Acid | Gm-SGPP (mg/g glycopeptide) |
|---|---|---|---|
| Asp | 15.33 | Gly | 16.42 |
| Thr | 16.29 | Ala | 33.51 |
| Ser | 15.80 | Pro | 14.37 |
| Glu | 13.90 | ||
| Total | 125.62 | ||
| Retention Time | Monosaccharide of Methylation | Characteristic Fragment Ions | Connection Type |
|---|---|---|---|
| 27.11 | 1,2,4,5-Ac4-3,6-Me2-Man | 113,130,131,190,233 | →2,4)Man(1→ |
| 28.23 | 1,5,6-Ac3-2,4-Me2-Man | 57,87,118,129,189 | →3,6)Man(1→ |
| 31.31 | 1,3,4,5-Ac4-2,6-Me2-Gal | 130,190,201,261 | →3,4)Gal(1→ |
| 22.43 | 1,5-Ac2-2,3,4,6-Me4-Gal | 57,118,145,161,205 | Gal(1→ |
| 24.80 | 1,3,5-Ac3-2,4,6-Me3-Gal | 87,101,118,129,161 | →3)Gal(1→ |
| 23.59 | 1,4,5-Ac3-2,3,6-Me3-Gal | 71,118,129,145,161 | →4)Gal(1→ |
| 36.71 | 1,2,4,5-Ac4-2,3,6-Me3-GlcNAc | 117,159,233 | →4)GlcNAc(1→ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hei, Z.; Zhao, M.; Tian, Y.; Chang, H.; Shen, X.; Xia, G.; Wang, J. Isolation and Characterization of a Novel Sialoglycopeptide Promoting Osteogenesis from Gadus morhua Eggs. Molecules 2020, 25, 156. https://doi.org/10.3390/molecules25010156
Hei Z, Zhao M, Tian Y, Chang H, Shen X, Xia G, Wang J. Isolation and Characterization of a Novel Sialoglycopeptide Promoting Osteogenesis from Gadus morhua Eggs. Molecules. 2020; 25(1):156. https://doi.org/10.3390/molecules25010156
Chicago/Turabian StyleHei, Zhiliang, Meihui Zhao, Yingying Tian, Hong Chang, Xuanri Shen, Guanghua Xia, and Jingfeng Wang. 2020. "Isolation and Characterization of a Novel Sialoglycopeptide Promoting Osteogenesis from Gadus morhua Eggs" Molecules 25, no. 1: 156. https://doi.org/10.3390/molecules25010156