
8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development

 ,
,
Abstract
:
1. Introduction
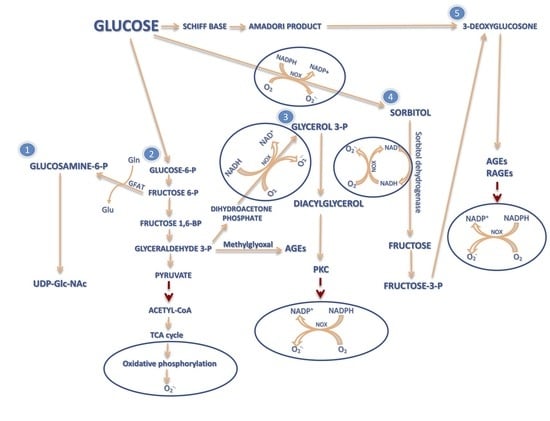
2. The Influence of ROS Overproduction on GDM Development
3. Types of the Antioxidant Defense Mechanisms
4. Biological Markers of Oxidative Stress in GDM Women
4.1. Influence of Oxidative Stress on Lipids
4.2. Influence of Oxidative Stress on Proteins
4.3. Influence of Oxidative Stress on Enzymatic and Non-Enzymatic Antioxidants
4.4. Influence of Oxidative Stress on DNA
5. Generation of the Most Abundant Lesions Observed in GDM: 8-OHdG and 8-oxodG
6. Repair Mechanism of 8-OHdG and 8-oxodG in Diabetes Mellitus
7. Concentration of 8-OHdG/8-oxodG and Risk of GDM Development
8. Maintenance of Genomic Integrity during GDM
9. 8-OHdG/8-oxodG as a Potential Biomarker for GDM—Summary
Author Contributions
Funding
Conflicts of Interest
References
- Badakhsh, M.; Balouchi, A.; Amirshahi, M.; Hashemi, Z. Gestational diabetes and its maternal and neonatal complications: A review article. IJPT 2016, 8, 18868–18878. [Google Scholar]
- Pikee, S.; Sakshi, M.; Aruna, N. Screening and Diagnosis of Gestational Diabetes Mellitus: From Controversy to Consensus. CRDOJ 2017, 2, 555600. [Google Scholar]
- Roglic, G. WHO Global report on diabetes: A summary. Int. J. Non-Commun. Dis. 2016, 1, 3–8. [Google Scholar] [CrossRef]
- Law, K.P.; Zhang, H. The pathogenesis and pathophysiology of gestational diabetes mellitus: Deductions from a three-part longitudinal metabolomics study in China. Clin. Chim. Acta 2017, 468, 60–70. [Google Scholar] [CrossRef]
- Ben-Haroush, A.; Yogev, Y.; Hod, M. Epidemiology of gestational diabetes mellitus and its association with Type 2 diabetes. Diabet. Med. 2004, 21, 103–113. [Google Scholar] [CrossRef]
- Kampmann, U.; Madsen, L.R.; Skajaa, G.O.; Iversen, D.S.; Moeller, N.; Ovesen, P. Gestational diabetes: A clinical update. World J. Diabetes 2015, 6, 1065–1072. [Google Scholar] [CrossRef]
- HAPO Study Cooperative Research Group; Metzger, B.E.; Lowe, L.P.; Dyer, A.R.; Trimble, E.R.; Chaovarindr, U.; Coustan, D.R.; Hadden, D.R.; McCance, D.R.; Hod, M.; et al. Hyperglycemia and adverse pregnancy outcomes. N. Engl. J. Med. 2008, 358, 1991–2002. [Google Scholar]
- Cypryk, K.; Szymczak, W.; Czupryniak, L.; Sobczak, M.; Lewiński, A. Gestational diabetes mellitus—An analysis of risk factors. Endokrynol. Pol. 2008, 59, 393–397. [Google Scholar]
- Kaaja, R.; Rönnemaa, T. Gestational diabetes: Pathogenesis and consequences to mother and offspring. Rev. Diabet. Stud. 2008, 5, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Shah, B.R.; Retnakaran, R.; Booth, G.L. Increased risk of cardiovascular disease in young women following gestational diabetes mellitus. Diabetes Care 2008, 31, 1668–1669. [Google Scholar] [CrossRef] [Green Version]
- Sathiamma, P.K.; Karunakaran, L. A prospective study on maternal and perinatal outcome of gestational diabetes mellitus. Int. J. Reprod. Contracept. Obstet. Gynecol. 2017, 6, 2933–2938. [Google Scholar]
- Saisho, Y.; Miyakoshi, K.; Tanaka, M.; Shimada, A.; Ikenoue, S.; Kadohira, I.; Yoshimura, Y.; Itoh, H. Beta cell dysfunction and its clinical significance in gestational diabetes. Endocr. J. 2010, 57, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmicki, M.; Telejko, B.; Zonenberg, A.; Szamatowicz, J.; Kretowski, A.; Nikolajuk, A.; Laudanski, P.; Gorska, M. Circulating pro- and anti-inflammatory cytokines in Polish women with gestational diabetes. Horm. Metab. Res. 2008, 40, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30 (Suppl. 2), S112–S119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik, M.; Chmielewska-Kassassir, M.; Grzywnowicz, K.; Woźniak, L.; Cypryk, K. The relationship between adipose tissue-derived hormones and gestational diabetes mellitus (GDM). Endokrynol. Pol. 2014, 65, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Lappas, M.; Hiden, U.; Desoye, G.; Froehlich, J.; Hauguel-de Mouzon, S.; Jawerbaum, A. The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid. Redox Signal. 2011, 15, 3061–3100. [Google Scholar] [CrossRef]
- Cadet, J.; Wagner, J.R.; Shafirovich, V.; Geacintov, N.E. One-electron oxidation reactions of purine and pyrimidine bases in cellular DNA. Int. J. Radiat. Biol. 2014, 90, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2’ -deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Qiu, C.; Hevner, K.; Abetew, D.; Enquobahrie, D.A.; Williams, M.A. Oxidative DNA damage in early pregnancy and risk of gestational diabetes mellitus: A pilot study. Clin. Biochem. 2011, 44, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Qiu, C.; Hevner, K.; Abetew, D.; Sedensky, M.; Morgan, P.; Enquobahrie, D.A.; Williams, M.A. Mitochondrial DNA copy number and oxidative DNA damage in placental tissues from gestational diabetes and control pregnancies: A pilot study. Clin. Lab. 2013, 59, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarniak, A.; Lipińska, J.; Tytman, K.; Lipińska, S. Endogenne mechanizmy powstawania reaktywnych form tlenu (ROS). Postepy Hig. Med. Dosw. 2016, 70, 1150–1164. [Google Scholar] [CrossRef] [PubMed]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Kalisz, O.; Wolski, T.; Gerkowicz, M.; Smorawski, M. Reaktywne formy tlenu (RFT) oraz ich rola w patogenezie niektórych chorób. Ann. Univ. Mariae Curie-Skłodowska Sect. DD Med. Vet. 2007, 62, 87–99. [Google Scholar]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Turek, I.A.; Wozniak, L.A.; Cypryk, K.; Wojcik, M. Hyperglycaemia-induced oxidative stress in gestational diabetes mellitus (GDM). Diabetol. Prakt. 2015, 4, 189–198. [Google Scholar]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [Green Version]
- Unoki, H.; Yamagishi, S. Advanced glycation end products and insulin resistance. Curr. Pharm. Des. 2008, 14, 987–989. [Google Scholar] [CrossRef]
- Yan, S.D.; Schmidt, A.M.; Anderson, G.M.; Zhang, J.; Brett, J.; Zou, Y.S.; Pinsky, D.; Stern, D. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J. Biol. Chem. 1994, 269, 9889–9897. [Google Scholar]
- Kuboki, K.; Jiang, Z.Y.; Takahara, N.; Ha, S.W.; Igarashi, M.; Yamauchi, T.; Feener, E.P.; Herbert, T.P.; Rhodes, C.J.; King, G.L. Regulation of endothelial constitutive nitric oxide synthase gene expression in endothelial cells and In Vivo: A specific vascular action of insulin. Circulation 2000, 101, 676–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Yu, Y.; Haigh, S.; Johnson, J.; Lucas, R.; Stepp, D.W.; Fulton, D.J. Regulation of NADPH oxidase 5 by protein kinase C isoforms. PLoS ONE 2014, 9, e88405. [Google Scholar] [CrossRef] [PubMed]
- Ogata, N.; Yamamoto, H.; Kugiyama, K.; Yasue, H.; Miyamoto, E. Involvement of protein kinase C in superoxide anion-induced activation of nuclear factor-kappa B in human endothelial cells. Cardiovasc. Res. 2000, 5, 513–521. [Google Scholar] [CrossRef]
- Eitel, K.; Staiger, H.; Rieger, J.; Mischak, H.; Brandhorst, H.; Brendel, M.D.; Bretzel, R.G.; Häring, H.U.; Kellerer, M. Protein kinase C delta activation and translocation to the nucleus are required for fatty acid-induced apoptosis of insulin-secreting cells. Diabetes 2003, 52, 991–997. [Google Scholar] [CrossRef] [Green Version]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S. Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. BioMed Res. Int. 2014, 2014, 801269. [Google Scholar] [CrossRef] [Green Version]
- Mathebula, S.D. Polyol pathway: A possible mechanism of diabetes complications in the eye. Afr. Vis. Eye Health 2015, 74, 5. [Google Scholar] [CrossRef]
- Schleicher, E.D.; Weigert, C. Role of the hexosamine biosynthetic pathway in diabetic nephropathy. Kidney Int. Suppl. 2000, 77, S13–S18. [Google Scholar] [CrossRef] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Bernea, E.G.; Antohe, F.; Mihai, A.; Ionecsu-Tirgoviste, C. Oxidative stress and gestational diabetes mellitus. The effects of supplements on oxidative stress. Proc. Rom. Acad. Ser. B 2018, 20, 121. [Google Scholar]
- Ho, E.; Galougahi, K.K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girotti, A.W. Mechanisms of lipid peroxidation. J. Free Radic. Biol. Med. 1985, 1, 87–95. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [PubMed]
- Biri, A.; Onana, A.; Devrimb, E.; Babacanc, F.; Kavutcua, M.; Durakb, I. Oxidant Status in Maternal and Cord Plasma and Placental Tissue in Gestational Diabetes. Placenta 2006, 27, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Grissa, O.; Ategbo, J.M.; Yessoufou, A.; Tabka, Z.; Miled, A.; Jerbi, M.; Dramane, K.L.; Moutairou, K.; Prost, J.; Hichami, A.; et al. Antioxidant status and circulating lipids are altered in human gestational diabetes and macrosomia. Transl. Res. 2007, 150, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Bis-Głuchowska, M.; Marciniak, B.; Szpringer-Boguń, E.; Rola, R.; Leszczyńska-Gorzelak, B.; Oleszczuk, J. Determination of antioxidative-peroxidative balance in the cord blood of newborns delivered to mothers with diabetes type G1. Ginekol. Pol. 2001, 72, 1255–1258. [Google Scholar]
- Vural, M.; Camuzcuoglu, H.; Toy, H.; Cece, H.; Aydin, H.; Eren, M.A.; Kocyigit, A.; Aksoy, N. Evaluation of the future atherosclerotic heart disease with oxidative stress and carotid artery intima media thickness in gestational diabetes mellitus. Endocr. Res. 2012, 37, 145–153. [Google Scholar] [CrossRef]
- Coughlan, M.T.; Vervaart, P.P.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Altered placental oxidative stress status in gestational diabetes mellitus. Placenta 2004, 25, 78–84. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Karacay, O.; Sepici-Dincel, A.; Karcaaltincaba, D.; Sahin, D.; Yalvaç, S.; Akyol, M.; Kandemir, O.; Altan, N. A quantitative evaluation of total antioxidant status and oxidative stress markers in preeclampsia and gestational diabetic patients in 24-36 weeks of gestation. Diabetes Res. Clin. Pract. 2010, 89, 231–238. [Google Scholar] [CrossRef]
- Gelisgen, R.; Genc, H.; Kayali, R.; Oncul, M.; Benian, A.; Guralp, O.; Uludag, S.; Cakatay, U.; Albayrak, M.; Uzun, H. Protein oxidation markers in women with and without gestational diabetes mellitus: A possible relation with paraoxonase activity. Diabetes Res. Clin. Pract. 2011, 94, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, H.M.; Lappas, M.; Georgiou, G.M.; Marita, A.; Bryant, V.J.; Hiscock, R.; Permezel, M.; Khalil, Z.; Rice, G.E. Screening for biomarkers predictive of gestational diabetes mellitus. Acta Diabetol. 2008, 45, 157–165. [Google Scholar] [CrossRef] [PubMed]
- López-Tinoco, C.; Roca, M.; García-Valero, A.; Murri, M.; Tinahones, F.J.; Segundo, C.; Bartha, J.L.; Aguilar-Diosdado, M. Oxidative stress and antioxidant status in patients with late-onset gestational diabetes mellitus. Acta Diabetol. 2013, 50, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Suhail, M.; Patil, S.; Khan, S.; Siddiqui, S. Antioxidant Vitamins and Lipoperoxidation in Non-pregnant, Pregnant, and Gestational Diabetic Women: Erythrocytes Osmotic Fragility Profiles. J. Clin. Med. Res. 2010, 2, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santra, D.; Sawhney, H.; Aggarwal, N.; Majumdar, S.; Vasishta, K. Lipid peroxidation and vitamin E status in gestational diabetes mellitus. J. Obstet. Gynaecol. Res. 2003, 29, 300–304. [Google Scholar] [CrossRef]
- Ba, X.; Boldogh, I. 8-Oxoguanine DNA glycosylase 1: Beyond repair of the oxidatively modified base lesions. Redox Biol. 2018, 14, 669–678. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated base damage to cellular DNA. Free Radic. Biol. Med. 2010, 49, 9–21. [Google Scholar] [CrossRef]
- Ghosh, R.; Mitchell, D.L. Effect of oxidative DNA damage in promoter elements on transcription factor binding. Nucleic Acids Res. 1999, 27, 3213–3218. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Pfeifer, G.P. Are there specific readers of oxidized 5-methylcytosine bases? BioEssays 2016, 38, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Marietta, C.; Gulam, H.; Brooks, P.J. A single 8,5’-cyclo-2’-deoxyadenosine lesion in a TATA box prevents binding of the TATA binding protein and strongly reduces transcription in vivo. DNA Repair 2002, 1, 967–975. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. The human genome as a target of oxidative modification: Damage to nucleic acids. In Redox-Genome Interactions in Health and Disease; Fuchs, J., Podda, M., Packer Marcel Dekker, L., Eds.; CRC Press: New York, NY, USA, 2003; pp. 145–192. [Google Scholar]
- Simone, S.; Gorin, Y.; Velagapudi, C.; Abboud, H.E.; Habib, S.L. Mechanism of oxidative DNA damage in diabetes: Tuberin inactivation and downregulation of DNA repair enzyme 8-oxo-7,8-dihydro-2’-deoxyguanosine-DNA glycosylase. Diabetes 2008, 57, 2626–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, T. An N-glycosidase from Escherichia coli that releases free uracil from DNA containing deaminated cytosine residues. Proc. Natl. Acad. Sci. USA 1974, 71, 3649–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, M.; Maki, H.; Sekiguchi, M.; Horiuchi, T. A specific role of MutT protein: To prevent dG.dA mispairing in DNA replication. Proc. Natl. Acad. Sci. USA 1989, 86, 3949–3952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakabeppu, Y.; Ohta, E.; Abolhassani, N. MTH1 as a nucleotide pool sanitizing enzyme: Friend or foe? Free Radic. Biol. Med. 2017, 107, 151–158. [Google Scholar] [CrossRef]
- Gelaleti, R.B.; Damasceno, D.C.; Lima, P.H.; Salvadori, D.M.; Calderon, I.M.; Peraçoli, J.C.; Rudge, M.V. Oxidative DNA damage in diabetic and mild gestational hyperglycemic pregnant women. Diabetol. Metab. Syndr. 2015, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R.; Raslová, K.; Smorovská, M.P.; Petrovská, H.; Ondrusová, A.; Vohnout, B.; Fábry, R.; Dusinská, M. DNA damage in diabetes: Correlation with a clinical marker. Free Radic. Biol. Med. 1998, 25, 373–377. [Google Scholar] [CrossRef]
- Lima, P.H.; Sinzato, Y.K.; Gelaleti, R.B.; Calderon, I.M.; Rudge, M.V.; Damasceno, D.C. Genotoxicity evaluation in severe or mild diabetic pregnancy in laboratory animals. Exp. Clin. Endocrinol. Diabetes 2012, 120, 303–307. [Google Scholar] [CrossRef]
- Lima, P.H.; Sinzato, Y.K.; de Souza Mda, S.; Braz, M.G.; Rudge, M.V.; Damasceno, D.C. Evaluation of level of DNA damage in blood leukocytes of non-diabetic and diabetic rat exposed to cigarette smoke. Mutat. Res. 2007, 628, 117–122. [Google Scholar] [CrossRef]
- Lima, P.H.; Damasceno, D.C.; Sinzato, Y.K.; de Souza Mda, S.; Salvadori, D.M.; Calderon, I.M.; Rudge, M.V. Levels of DNA damage in blood leukocyte samples from non-diabetic and diabetic female rats and their fetuses exposed to air or cigarette smoke. Mutat. Res. 2008, 653, 44–49. [Google Scholar] [CrossRef]
- Moreli, J.B.; Santos, J.H.; Lorenzon-Ojea, A.R.; Corrêa-Silva, S.; Fortunato, R.S.; Rocha, C.R.; Rudge, M.V.; Damasceno, D.C.; Bevilacqua, E.; Calderon, I.M. Hyperglycemia Differentially Affects Maternal and Fetal DNA Integrity and DNA Damage Response. Int. J. Biol. Sci. 2016, 12, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Giglia-Mari, G.; Zotter, A.; Vermeulen, W. DNA damage response. Cold Spring Harb. Perspect. Biol. 2011, 3, a000745. [Google Scholar] [CrossRef] [PubMed]
- Moslehi, R.; Signore, C.; Tamura, D.; Mills, J.L.; Digiovanna, J.J.; Tucker, M.A.; Troendle, J.; Ueda, T.; Boyle, J.; Khan, S.G.; et al. Adverse effects of trichothiodystrophy DNA repair and transcription gene disorder on human fetal development. Clin. Genet. 2010, 77, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pácal, L.; Varvařovská, J.; Rušavý, Z.; Lacigová, S.; Stětina, R.; Racek, J.; Pomahačová, R.; Tanhäuserová, V.; Kaňková, K. Parameters of oxidative stress, DNA damage and DNA repair in type 1 and type 2 diabetes mellitus. Arch Physiol. Biochem. 2011, 117, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Metzger, B.E.; Buchanan, T.A.; Coustan, D.R.; de Leiva, A.; Dunger, D.B.; Hadden, D.R.; Hod, M.; Kitzmiller, J.L.; Kjos, S.L.; Oats, J.N.; et al. Summary and recommendations of the Fifth International Workshop-Conference on Gestational Diabetes Mellitus. Diabetes Care 2007, 30 (Suppl. 2), S251–S260. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association (ADA). Standards of medical care in Diabetes. Diabetes Care 2014, 37, S14–S80. [Google Scholar] [CrossRef] [Green Version]
- Cederberg, J.; Eriksson, U.J. Antioxidative treatment of pregnant diabetic rats diminishes embryonic dysmorphogenesis. Birth Defects Res. A Clin. Mol. Teratol. 2005, 73, 498–505. [Google Scholar] [CrossRef]
- Poel, Y.H.; Hummel, P.; Lips, P.; Stam, F.; van der Ploeg, T.; Simsek, S. Vitamin D and gestational diabetes: A systematic review and meta-analysis. Eur. J. Intern. Med. 2012, 23, 465–469. [Google Scholar] [CrossRef]
- Rumbold, A.; Ota, E.; Nagata, C.; Shahrook, S.; Crowther, C.A. Vitamin C supplementation in pregnancy. Cochrane Database Syst. Rev. 2015, 9. [Google Scholar] [CrossRef]
- Rowan, J.A.; Hague, W.M.; Gao, W.; Batin, M.R.; Moore, M.P. Metformin versus insulin for the treatment of gestational diabetes. N. Engl. J. Med. 2008, 358, 2003–2015. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Grzeskowiak, L.; Williamson, K.; Downie, M.R.; Crowther, C.A. Insulin for the treatment of women with gestational diabetes. Cochrane Database Syst. Rev. 2017, 11. [Google Scholar] [CrossRef]
- Park, K.S.; Kim, J.H.; Kim, M.S.; Kim, J.M.; Kim, S.K.; Choi, J.Y.; Chung, M.H.; Han, B.; Kim, S.Y.; Lee, H.K. Effects of insulin and antioxidant on plasma 8-hydroxyguanine and tissue 8-hydroxy-2’-deoxyguanosinein streptozotocin-induced diabetic rats. Diabetes 2001, 50, 2837–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Zhou, M.; Xie, J.; Chao, P.; Feng, Q.; Wu, J. High glucose levels promote the proliferation of breast cancer cells through GTPases. Breast Cancer 2017, 9, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Type of Diabetes | Study Type | Sample | Result | Reference |
|---|---|---|---|---|
| MGH diabetic women with obesity and hypertension | Clinical | Maternal lymphocytes | ↑ Overall oxidative DNA damage | [66] |
| GDM | Clinical | Maternal lymphocytes | ↑ 8-oxoG | |
| MGH | Clinical | Maternal urine | ↑ 8-OHdG | |
| GDM | Clinical | Maternal urine | ↑ 8-OHdG | |
| GDM | Clinical | Maternal lymphocytes | ↑ 8-oxoG | [67] |
| GDM | Clinical | Maternal urine | ↑ 8-OHdG | [20] |
| MGH, induced by streptozotocin | Experimental | Maternal and fetal leukocytes | ↑ 8-OHdG ↑ 8-oxo-dG | [68] |
| Severe, induced by streptozotocin | Experimental | Maternal and fetal leukocytes | ↑ Overall oxidative DNA damage | |
| Severe, induced by streptozotocin | Experimental | Maternal leukocytes | ↑ Overall oxidative DNA damage | [69] |
| Severe, induced by streptozotocin | Experimental | Fetal leukocytes | ↑ Overall oxidative DNA damage | [70] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbaniak, S.K.; Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development. Molecules 2020, 25, 202. https://doi.org/10.3390/molecules25010202
Urbaniak SK, Boguszewska K, Szewczuk M, Kaźmierczak-Barańska J, Karwowski BT. 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development. Molecules. 2020; 25(1):202. https://doi.org/10.3390/molecules25010202
Chicago/Turabian StyleUrbaniak, Sandra K., Karolina Boguszewska, Michał Szewczuk, Julia Kaźmierczak-Barańska, and Bolesław T. Karwowski. 2020. "8-Oxo-7,8-Dihydro-2′-Deoxyguanosine (8-oxodG) and 8-Hydroxy-2′-Deoxyguanosine (8-OHdG) as a Potential Biomarker for Gestational Diabetes Mellitus (GDM) Development" Molecules 25, no. 1: 202. https://doi.org/10.3390/molecules25010202