
Structure and Dynamics of Spider Silk Studied with Solid-State Nuclear Magnetic Resonance and Molecular Dynamics Simulation

Department of Biotechnology, Tokyo University of Agriculture and Technology, 2-24-16 Nakacho, Koganei, Tokyo 184-8588, Japan
Molecules 2020, 25(11), 2634; https://doi.org/10.3390/molecules25112634
Submission received: 29 April 2020
/
Revised: 1 June 2020
/
Accepted: 1 June 2020
/
Published: 5 June 2020
(This article belongs to the Special Issue Silk Fibroin Materials)
Abstract
:This review will introduce very recent studies using solid-state nuclear magnetic resonance (NMR) and molecular dynamics (MD) simulation on the structure and dynamics of spider dragline silks conducted by the author’s research group. Spider dragline silks possess extraordinary mechanical properties by combining high tensile strength with outstanding elongation before breaking, and therefore continue to attract attention of researchers in biology, biochemistry, biophysics, analytical chemistry, polymer technology, textile technology, and tissue engineering. However, the inherently non-crystalline structure means that X-ray diffraction and electron diffraction methods provide only limited information because it is difficult to study the molecular structure of the amorphous region. The most detailed picture of the structure and dynamics of the silks in the solid state experimentally have come from solid-state NMR measurements coupled with stable isotope labeling of the silks and the related silk peptides. In addition, combination of solid-state NMR and MD simulation was very powerful analytical tools to understand the local conformation and dynamics of the spider dragline silk in atomic resolution. In this review, the author will emphasize how solid-state NMR and MD simulation have contributed to a better understanding of the structure and dynamics in the spider dragline silks.
1. Introduction
There are a variety of silkworms and spiders, each producing silk with unique primary and higher order structures [1,2]. Over nearly 400 million years, orb-web weaving spiders have evolved the ability to produce up to six different silks and one glue-like substance that possesses amazing, yet unique, mechanical properties as summarized in Figure 1 [2,3,4,5]. Among the spider silks, spider dragline silks have very excellent mechanical properties such as the combination of high strength and high toughness [1,2,3,4,5,6,7]. Another unique characteristic of the spider silks is supercontraction, which occurs in the process of hydration. Interaction with water causes spider dragline silk fiber to contract up to 50% in length and swell in diameter [8,9,10]. In addition, recent developments in biotechnology have opened the possibility to reproduce the recombinant spider silk proteins [11]. These appealing backgrounds have attracted researchers in diverse fields, such as biology, biochemistry, biophysics, analytical chemistry, polymer technology, textile technology, and biomaterials. The unique properties of spider silks are essentially originated from the structure and dynamics, and therefore, the atomic resolution information on the structure gives us the answer to why the silk has such unique and excellent properties [1,7,12].
A variety of techniques such as X-ray diffraction, Fourier transform infrared/Raman spectroscopies, and transmission electron microscopy have been applied to clarify the structure of the spider dragline silk [13,14,15,16,17,18,19,20,21,22,23,24,25]. By using these techniques, the structures have been studied from secondary structures to molecular arrangement to hierarchical structure. For example, the X-ray diffraction technique has been used extensively to clarify the structure of silk at the atomic/molecular level, including the size and orientation of nano-crystallites, and also molecular chain arrangement. However, because of the semi-crystalline nature of silk, it is quite difficult to study the atomic/molecular structure of the amorphous region in the silk by X-ray diffraction. To replace or complement this technique, NMR spectroscopy has been applied recently and has been demonstrated as a very effective method to clarify the structure and dynamics of the dragline spider silk [26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65]. The solid-state NMR studies used a combination of approaches [66]. Spiders can be fed on an isotopically labeled diet, allowing isotopic labeling of silk within the silk gland: either uniformly or by feeding amino acids labeled at individual positions [27,28,29,30,31,32,34,35,36,37,40,42,43,44,45,47,48,49,50,53,54,55,56,59,61]. In addition, synthetic peptides with isotopically labelings in specific positions have proved very useful model systems permitting site-specific structural information [33,38,39,41,46,51,52,60,62,63,64,65]. The conformation-dependent 13C chemical shift in the 13C cross polarization and magic angle spinning (CP/MAS) NMR spectra of silks and the model peptides coupled with selective 13C labeling can be used effectively for determination of local structure at secondary structure level in an amino acid-specific manner [66,67,68,69,70,71,72]. In particular, the combination of 13C labeling of specific sites in amino acids, and 13C Cα and Cβ chemical shift contour maps of these sites is very useful for the structural study. Moreover, many kinds of advanced solid-state NMR techniques have been applied to obtain local structure and dynamics of spider silk at atomic resolution as will be shown below [27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,61].
On the other hand, molecular dynamics (MD) simulation is very effective to understand the local conformation and dynamics of Nephila clavipes (N. clavipes) dragline silks theoretically and to verify the experimental results [56,58,62,63,65]. MD simulation can play a critical role in connecting experimental restraints with potentially plausible molecular structures. Therefore, it has been also used to understand the structure and structural changes of silk fibroins including silk fiber formation mechanism and the chain dynamics [73,74,75,76,77,78,79,80,81,82,83,84,85]. Especially, combination of solid-state NMR and MD simulations was very strong analytical tools to understand the local conformation and dynamics of the spider dragline silk in atomic resolution [56,58,62,63,65].
We previously reviewed about NMR studies of silk several times [86,87,88,89,90,91] and most recently, an excellent review about NMR characterization of silk including spider silk was published by Guo and Yarger [92]. Therefore, the author hopes to avoid duplication of contents here.
In this review, recent studies about the structure and dynamics of the dragline silks of two kinds of spiders, N. clavipes and Nephila clavata (N. clavata) in the dry and hydrated states using solid-state NMR and MD simulation were introduced because the dragline silk of the golden-orb weaver, N. clavipes, has become the benchmark and N. clavata can be easily obtained in Japan.
2. Major Ampullate Silk Fiber of N. clavipes and Their 13C Solid-State NMR Spectra
Spider dragline silks are synthesized in ampullate glands located in the abdomen and spun through a series of orifices (spinnerets) [2,3,4,5]. The silk fiber originating in the major ampullate gland has been the most studied. The major ampullate silk consists of two structural proteins, designated as spidron 1 (MaSp1) and spidron 2 (MaSp2) [93]. The dominant MaSp1 protein can be described as a block copolymer consisting of polyalanine (poly-A) domain as the crystalline region and Gly-rich regions as the less structurally ordered non-crystalline region [1,2,3,4,5,6,7]. The former region has been associated with high fiber strength, and the latter is a source of the high elasticity observed for the spider silk fiber. They assemble into fibers with the excellent mechanical properties.
The dragline silk of N. clavipes has become the benchmark and therefore, let us start by introducing NMR studies of this spider dragline silk. The 13C CP/MAS NMR spectrum of the major ampullate silk fibers of N. clavipes reported by Simmons et al. [26] is shown in Figure 2 together with the assignment.
The integrated areas of peaks from Gly, Ala, Glu, and Tyr were 42%, 46%, 9%, and 4%, compared to values of 43%, 30%, 7%, and 4% obtained by amino acid analysis, which means that both the crystalline and amorphous regions of silk are detected by 13C CP/MAS NMR. The Ala Cα and Ala Cβ chemical shifts were 49.0 and 20.6 ppm, respectively, indicating that the spider silk fiber took antiparallel β (AP-β) sheet structure judging from the 13C conformation-dependent chemical shift data [67,68,69,70,71,72]. On the other hand, the structural analysis of the Gly-rich region is difficult because the region is heterogeneous and amorphous. Therefore, the combination of solid-state NMR and MD simulation together with use of appropriate stable-isotope labeled model peptides of the typical sequences in the Gly-rich region is very effective to clarify the structure and dynamics.
3. Structure of the Gly-Rich Region of N. clavipes Dragline Silk
The typical amino acid sequence including poly-A and Gly-rich regions of N. clavipes dragline silk reported by Xu and Lewis [4] is partially shown in Figure 3.
It is difficult to analyze the structure of the Gly-rich region directly from simple 13C CP/MAS NMR spectrum of the dragline silk as shown in Figure 2 because of the heterogeneity of the primary structure. Instead of this, the use of synthetic peptides with isotopically labelings in specific positions provides a sequential model of the Gly-rich region permitting site-specific structural information. Therefore, the selectively stable-isotope labeled 47-mer peptides flanked by (Ala)6 at both ends were synthesized as summarized in Table 1 [46,60,62]. The sequence was selected in Figure 3 as underlined sequence, a and poly-Glu blocks were attached at both N- and C-termini to make them water-soluble [94]. In order to determine the local conformation of Gly-rich region, rotational echo double resonance (REDOR) NMR [29,46,95,96] was used to determine the inter-nuclear distance between the 15N and 13C labeled sites as summarized in Table 1 (I).
The inter-nuclear distances between 13C(i) and 15N(i+2) nuclei in the Gly(i)-X-Gly(i+2) motifs were also calculated using the torsion angles of the X amino acid (X = Q, Y, L, and R) taken from the Protein Data Bank (PDB) database [46] and plotted together with the corresponding distances determined by the REDOR experiment in Figure 4 (red circles). The corresponding distances of typical conformations are also shown in Figure 4 (gray hollow circles), i.e., AP-β sheet; 4.6 Å, α-helix and type II β-turn; 3.2 Å and 31-helix; 4.2 Å [46]. The REDOR-determined distances did not match any distances of typical conformations. Therefore, it is concluded that the Gly-rich region took random coil and no well-defined conformation.
However, the presence of 31-helix (Φ and Ψ = −90°, 150°) in the Gly-rich region has been frequently proposed in previous papers [28,32,42,43]. Therefore, it is important to examine the possibility about the presence of 31-helix in detail using these 47-mer peptides.
The 13C chemical shift of Ala Cβ peak, 17.4 ppm is a measure of the presence of 31-helix as reported previously [33] because the value, 17.4 ppm is the 13C chemical shift of the Ala Cβ peak of poly(Ala-Gly-Gly) with the 31-helix form confirmed by X-ray diffraction analysis [97]. Here it is necessary to be careful about the chemical shift reference when comparing the 13C chemical shift-dependent conformations of silks. We used tetramethylsilane, TMS, as the external chemical shift reference in the solution and solid-state NMR of silks [66,70,71]. Three kinds of the 47-mer peptides introduced three successive 13C-labeled amino acid residues, [2-13C]Gly, [3-13C]Ala, and [1-13C]Gly at different positions were synthesized as listed in Table 1(II) [62]. The 13C CP/MAS NMR spectra of three Ala Cβ peaks of the Ala13, Ala26, and Ala36 residues in the [2-13C]Gly[3-13C]Ala[1-13C]Gly motifs at the different three positions are shown in Figure 5. Judging from the 13C chemical shifts of Ala Cα, Cβ, and C = O peaks of (Ala)6 residues at the both ends, these residues took on the AP-β sheet structure. It was clear that the Ala13, Ala26, and Ala36 residues in the Gly-Ala-Gly motifs in the Gly-rich region did not adopt the 31-helix because the Cβ chemical shifts of these three Ala residues were not 17.4 ppm, which shows a typical 31-helix. In order to determine the local conformations of the individual Ala and Gly residues in detail, the fractions of individual local conformations were determined from the 13C labeled Gly and Ala residues of these three kinds of peptides by the peak deconvolution analysis. In addition, those of other Gly residues were determined for six kinds of the peptides used for REDOR experiments (Table 1 (I)) and two additional 47-mer peptides with [1-13C]Gly12or [1-13C]Gly35 residues [62].
The fractions of several conformations were plotted against the position of the 13C-labeled nuclei in a 47-mer peptide chain in Figure 6a. Due to the presence of poly-A sequences with AP-β structure at both N- and C-termini, the residues located close to the poly-A sequences tended to adopt the AP-β structure. Actually, the fractions of AP-β structure rapidly decreased toward the center from the edge, in contrast to those of random coil. The fraction of β-turn structure tended to decrease after increasing once towards the center from the edge although the change was relatively small. In order to examine the change in the fractions theoretically, MD simulations were performed as shown in Figure 6b [62]. Thus, the observed results were successfully rationalized through the MD simulation.
4. Packing Structure of the Polyalanine Region of N. clavipes Dragline Silk
It is important to clarify the packing structure of poly-A region with AP-β sheet structure to clarify the origin of higher fiber strength in the atomic level, but there have been no detailed reports about the packing structure so far. Therefore, the packing structure of poly-A region was started to analyze for a series of alanine oligopeptides, (Ala)n (n = 4–8 and 12) with AP-β sheet structures using both solid-state NMR spectroscopy and X-ray crystallography as shown in Figure 7 [52]. Here (Ala)12 is the typical number of the Ala residue used for the model of poly-A region in Samia cynthia ricini (S. c. ricini) silk fibroin whose primary structure is similar to that of the spider dragline silk [88]. The 13C chemical shifts and line shapes of the Ala Cβ carbons were used together with X-ray diffraction patterns for the structural analyses. The atomic co-ordinates of (Ala)4 were determined using a single crystal X-ray diffraction analysis of the single crystal (Ala)4 sample [52]. It is noted that both the line shapes of the Ala Cβ carbons and X-ray diffraction powder patterns of (Ala)n (n = 7, 8, and 12) were markedly different from those for short alanine oligopeptides, (Ala)n (n = 4–6). Namely, the chemical shifts of the central peaks with highest intensity were 20.4 ppm and the same among the corresponding chemical shifts of the Ala Cβ carbons of (Ala)n (n = 4–6). On the other hand, the Ala Cβ peaks had two peaks at 22.7 ppm and 19.6 ppm together with a broad peak at 21 ppm.
In the X-ray diffraction spectra, the second peak 2 shifted from 2θ = 19.2° to 2θ = 20.4° remarkably [52]. These data indicate that the packing arrangement of alanine oligopeptides, (Ala)n with the AP-β sheet structure changed from rectangular (n = 4–6) to staggered (n = 7, 8, and 12) arrangement.
Later, the transition from the rectangular to the staggered arrangement in (Ala)6 was observed from the change in the Ala Cβ peak through heat treatment at 200 °C for 4 h and therefore the rectangular structure was stable for (Ala)6 and shorter poly-A chains when the stabilization of the intermolecular hydrogen bonding with bound water molecules occurred at the end groups of the terminal residues [58]. However, poly-A regions existed between both ends of Gly-rich sequences in spider dragline silks and therefore, the packing states of poly-A regions should be examined by taking into account these sequences again.
In Figure 3, the underlined sequence (b) was selected and a series of the sequential peptides where the poly-A regions exist between both ends, i.e., (E)4GGLGGQGAG(A)nGGAGQGGYGG(E)4 (n = 3–8) were studied for analyzing the packing structure of the poly-A region. The Gr(A)nGr* (n = 3–8) samples were soluble in water because of the presence of two (Glu)4 blocks at both termini. Here, Gr = (E)4GGLGGQGAG and Gr* = GGAGQGGYGG(E)4. After lowering the pH to 2, the Gr(A)nGr* (n = 3–5) samples were still soluble, but the samples (n = 6–8) were precipitated and took AP-β sheet structures. On the other hand, the Gr(A)nGr* (n = 4–8) samples were precipitated after adding methanol (MeOH) to the formic acid (FA) solutions.
Figure 8 shows the Ala Cβ peaks in the dipolar dephasing 13C CP / MAS NMR spectra of Gr(Ala)nGr* (n = 4 and 6–8) after FA and MeOH treatment to change them to the AP-β sheet structure. Both the ① 22.7 ppm and ③ 19.8 ppm peaks are assigned to the AP-β sheet structure with a staggered packing arrangement and the ④ 16.8 ppm peak is assigned to random coil. The peak ② at 21.2 ppm is assigned to the general AP-β sheet structure rather than rectangular packing arrangement because the chemical shift shows the typical AP-β sheet structure and the poly-A sequence is not necessary to consider the possibility of the inter-molecular hydrogen bonding with bound waters. The MD simulation was performed for the sequence GGLGGQGAG(A)6GGAGQGGYGG without two (E)4 blocks to confirm the observed results as shown in Figure 9 [63].
Almost all Ala residues in the central sequence, (Ala)6, were in the AP-β sheet structure (96–98%); this finding was similar to the observed data for Gr(Ala)6Gr* (94.5%).
In addition, the staggered packing arrangement was preserved even after the annealing and replica exchange molecular dynamics (REMD) simulations, suggesting that staggered packing is a reasonably stable structure [63]. The 13C CP/MAS NMR spectra of a series of AP-β sheet poly-A samples were reported by Lee and Ramamoorthy [98] and Wildman et al. [99]. In their papers, a similar Ala Cβ spectral pattern-like staggered arrangement was reported, that is, an asymmetric peak at 19.9 ppm and a small peak at 22.9 ppm for AP-β sheet poly-A samples with high molecular weight. In general, X-ray diffraction is a very powerful technique used to study the packing structure, but such studies have been difficult to conduct because of the large amounts of amorphous fraction in dragline silk fiber as shown in Figure 7 for N. clavata dragline silk fibers [59].
5. Dynamical Behavior and Hydration of N. clavipes Dragline Silk
The unique characteristic of the spider silks, supercontraction, which occurs in the process of hydration [8,9,10], has been the interest of many NMR researchers studying the spider dragline silk [27,30,31,34,35,37,43,45,47,48,49,53,54,57,59,60,61]. For example, Figure 10 shows the aliphatic region from fully relaxed 13C directly detected MAS NMR spectrum of N. clavipes major ampullate silk in the hydrated state reported by Jenkins et al. [48]. Compared with Figure 2, it is noted that all peaks became shaper, i.e., three Ala Cβ, Glu Cβ and Glu Cγ, and two Ser Cβ peaks could be observed separately. Two Ala Cβ peaks at the AP-β sheet region were also observed as discussed in the previous section in detail.
Holland and Yarger groups studied the structure and dynamics of stable-isotope labeled spider dragline silks in the dry and hydrated states very actively using several advanced two-dimensional (2D) solid-state NMR, i.e., 2D 13C-13C correlation spectrum with dipolar assisted rotational resonance, 2D 1H-13C or 2D 13C-15N Heteronuclear correlation NMR, Homonuclear double quantum NMR and 2D incredible natural abundance double quantum transfer experiment as reviewed previously [89,90,92].
The structure and dynamics of N. clavipes dragline silks was also studied by using the selectively stable-isotope labeled 47-mer peptides flanked by (Ala)6 at both ends listed in Table 1 (II)[60]. Figure 11 is the expanded regions (10−70 ppm) of the 13C-labeled 47 mer peptides with three 13C-labeled amino acid motifs, (A) [2-13C]Gly25[3-13C]Ala26[1-13C]Gly27 or (B) [2-13C]Gly35[3-13C]Ala36[1-13C]Gly37, respectively in the dry and hydrated states [60]. Here, (a) 13C refocused insensitive nuclei enhanced by polarization transfer (r-INEPT) [100], (b) 13C dipolar decoupled magic angle spinning (DD/MAS) [101,102], and (c) 13C CP/MAS NMR spectra in the hydrated states, and (d) 13C CP/MAS spectrum in the dry state. The 13C r-INEPT, where the pulse sequence was developed for solution NMR, is sensitive to the mobile component of the peptide with fast isotropic motion (>105 Hz) [103]. In contrast, 13C CP/MAS NMR can observe selectively immobile components or those with very slow motion (<104 Hz) [103]. If the penetration of water molecules causes an increase in the chain mobility, a loss in the CP signal of the amino acid residues occurs and consequently such a mobile domain cannot be observed in the 13C CP/MAS NMR spectra [43]. 13C DD/MAS NMR can be used to detect the mobile domains as well as the immobile domains [102]. Thus, these three kinds of 13C NMR techniques, 13C r-INEPT, 13C CP/MAS, and 13C DD/MAS NMR, provide different perspectives on the dynamic behavior of hydrated peptides and can be used together to characterize their local structure and conformations [57,59,60,104,105]. The 13C CP/MAS NMR spectra of two peptides in the dry state are the same as those in Figure 5. With hydration, sharp peaks in the Gly Cα and Ala Cβ carbons at 43.0 and 16.6 ppm, respectively assigned to the random coil conformation with high mobilities can be observed in the 13C CP/MAS NMR spectrum (c) of the peptide (A). Additionally, these sharp peaks became dominant in the 13C DD/MAS NMR (b) and 13C r-INEPT (a) spectra.
Similar changes in the spectra were also observed for another 47-mer peptide with three 13C-labeled amino acid motifs, [2- 13C]Gly12 [3-13C]Ala13[1-13C]Gly14 (not shown). In contrast to these observations, the effect of the hydration of the peptide (B), [2-13C]Gly35 [3-13C]Ala36 [1-13C]Gly37-47 mer peptide was quite different. Only a small difference in the 13C CP/MAS NMR spectra was observed between the dry (d) and hydrated states (c). In addition, the 13C DD/MAS NMR and 13C r-INEPT spectra ((b) and (a), respectively) showed very small sharp peaks with random coil chemical shifts in the hydrated state. Thus, the effect of hydration is very small for the motif, [2-13C]Gly35 [3-13C]Ala36 [1-13C]Gly37 because the motif is adjacent to the (Ala)6(Glu)4 sequences with AP-β structure. These results indicate that the hydration occurs heterogeneously along the Gly-rich region. The MD simulations are useful in examining changes in the local conformations of the individual residues of silks in the dry and hydrated states from the theoretical perspective [42,52,62,68,76,77,78,79,80,81,82]. Figure 12 shows the distributions of the Φ-Ψ torsion angles of the 13C labeled residues in the motifs, Gly12Ala13Gly14, Gly25Ala26Gly27, and Gly35Ala36Gly37 of the Gly-rich region in the dry and hydrated states, respectively, as calculated by MD simulation [60]. As is expected, the distributions of Φ-Ψ torsion angles of the residues become generally larger due to partial breaking of the hydrogen bonds in peptides by water molecules. Changes in the Φ-Ψ torsion angles of the motif, Gly25Ala26Gly27, shown in Figure 12 follow such a prediction. Thus, broader distributions of Φ-Ψ torsion angles by hydration indicate the occurrence of fast exchange among several conformations in water. This supports the appearance of the sharp peaks in the 13C DD/MAS and 13C r-INEPT NMR spectra as shown in Figure 11A. On the other hand, this is not the case for the motif, Gly35Ala36Gly37, where with hydration the change in the Φ-Ψ torsion angles was small, especially for the residues, Ala36Gly37, and the distribution of the Φ-Ψ torsion angles was confined to the AP-β sheet region. Thus, the residues close the C-terminal poly-Ala sequence, the Gly37 and Ala36 residues, were structured and stable. This is in agreement with the experimental results (Figure 11B).
6. Dynamical Behavior and Hydration of N. clavata Dragline Silk
N. clavata is often known as the Joro spider in East Asia (Japan, China, Korea, and Taiwan) and can be easily obtained in Japan [106,107]. Therefore, the structural and dynamical studies of dragline silk were performed for N. clavata. Figure 13 is the expanded regions (10−70 ppm) of four kinds of 13C sold-state NMR spectra of N. clavata dragline silk fibers in the dry and hydrated states, i.e., (a) 13C r-INEPT, (b) 13C DD/MAS, and (c) 13C CP/MAS NMR spectra in the hydrated states, and (d) 13C CP/MAS spectrum in the dry state [59]. The intensities of the 13C CP/MAS NMR peaks except for Ala Cα and Cβ peaks decreased significantly in the hydrated state (c), which was due to a significant loss in CP signals of the carbons whose mobility increased by hydration as mentioned above. Actually, several small peaks for the other amino acid residues in the 13C CP/MAS NMR spectra in the hydrated state (c) were observed as peaks with increased intensities in the 13C DD/MAS NMR spectrum (b) and as sharp peaks in the 13C r-INEPT spectrum (a) in the hydrated state. Only the random coil peak of the Ala residue was observed in the 13C r-INEPT spectrum, and the Ala peaks in the poly-A sequences with AP-β sheet structure were no longer observed.
This meant that water molecules could penetrate inside the random coil domains of N. clavata dragline silk fibers, but hardly penetrated through the hydrophobic β-sheet domains of poly-A sequences. The Ala Cβ peak region was analyzed in detail for [3-13C]Ala- N. clavata dragline silk fibers. It is speculated that there are also contributions from the peaks of the Leu and Arg side chains, especially at the lower field region, 20-24 ppm in the chemical shift range from 14 to 24 ppm of the Ala Cβ peak in the 13C natural abundant spectrum of N. clavata dragline silk as shown in Figure 13. However, these additional contributions to the Ala Cβ peak can be neglected by [3-13C]Ala labeling, which makes it possible to analyze these Cβ spectral region quantitatively.
Figure 14 shows Ala Cβ peaks in the 13C CP/MAS NMR spectra of [3-13C]Ala-N. clavata dragline silk fiber in the dry and hydrated states [59]. The percentages of random coil changed from 23% in the dry state to 10% in the hydrated state in the 13C CP/MAS NMR spectra (A). As mentioned above, a significant loss in CP signals occurred for the Ala Cβ carbons with random coil because of increased mobility by penetration of water molecules in the hydrated state. In addition, the line shape became sharper in the hydrated state. From the Ala Cβ peaks (B) with exclusively AP-β sheet structures, the percentage of the general AP-β sheet and staggered packing arrangement were determined as 49% and 51%, respectively in the dry state, and 40% and 60%, respectively in the hydrated state.
Solid-state NMR has the advantage that it can provide detailed structural information and can also give the dynamical information at different motional levels from the side-chain fast motion to the slower motion of the backbone chain of the spider dragline silks.
Thus, the relaxation behavior of Ala residues in [3-13C]Ala-N. clavata dragline silk fiber was studied in relation to the difference in the packing structures in the dry and hydrated states [61]. Figure 15 shows a series of partially relaxed 13C CP/MAS NMR spectra of Ala Cβ peaks of [3-13C]Ala-N. clavata dragline silk fiber in the dry state (A) and in the hydrated state (B) as a function of delay time τ for T1 determinations at 25 °C. The T1 values were 0.61 ± 0.02 s for 16.6 ppm (random coil), 0.57 ± 0.02 s for AP-β sheet (main peak at 20.2 ppm: mixture of rectangular and staggered peaks), and 0.93 ± 0.02 s for AP-β sheet (22.9 ppm: lower field staggered peak). On the other hand, the T1 values in the hydrated state were 0.48 ± 0.01 s for 17.2 ppm peak, 0.44 ± 0.02 s for 20.2 ppm peak, and 0.88 ± 0.02 s for 22.9 ppm, which are slightly shorter than those of the corresponding peaks in the dry state. Thus, it is noted that the T1 value of the 22.9 ppm was significantly longer than other two peaks in both dry and hydrated states. The longer T1 value means a faster motion, which was obtained from the temperature dependent T1 experiment [61]. Thus, the motional behavior of the lowest field peak assigned exclusively staggered packing structure shows the fastest hopping motion of the Cβ carbon about the C3 axis of Ala Cβ group in the dry and hydrated states [108,109].
The origin of these long T1 values in the staggered packing structure was already studied from the 13C solid-state NMR relaxation work of S. c. ricini silk fibroin fiber as shown in Figure 16 [110,111,112,113]. Especially, the MD simulation was performed to interpret the relationship between the packing structure and the dynamical behavior of these Ala Cβ groups in the poly-A region of S. c. ricini silk fibers [110]. Namely, two of the Ala Cβ carbons out of eight existing in the unit cell of the staggered packing structure exhibited the fastest hopping motion in spite of the shortest Cβ−Cβ distance. The unusual motion of the Ala methyl groups is named as a geared hopping motion, as shown in Figure 16.
7. Conclusions and Future Aspects
In this review, the author showed how solid-state NMR coupled with stable isotope labeling of the silks and the related silk peptides, and combination of solid-state NMR and MD simulation have contributed to a better understanding the structure and dynamics of the spider dragline silks in the dry and hydrated states. Recently, the excellent mechanical properties of spider silk fiber have been challenged to predict on the basis of the atomic resolution structure of the crystalline region embedded in a softer-amorphous region using MD simulations [73,74,75,76,77,78,79,80,81,82,83,84,85]. In the future, such a prediction of the mechanical properties of natural and recombinant spider silks [11,57,59,61] is expected to increase more and more for molecular design of new materials with excellent mechanical properties. Therefore, a more detailed investigation about the atomic resolution of spider silk fiber seems to be required experimentally. Moreover, it is important to accumulate a lot of knowledge about the structures and dynamics of other silks and/or silks of other species, including smaller orb weavers and non-orb weavers, and also change in the structure and dynamics of the silks to explain spider silk property variations in ecological and evolutionary contexts other than hydration [114]. NMR spectroscopy, especially solid-state NMR including dynamic nuclear polarization (DNP) [115] will undoubtedly use more frequently for the purpose.
Funding
This research was funded by Impulsing Paradigm Change through Disruptive Technologies Program (ImPACT) and a JSPS KAKENHI, Grant-in-Aid for Scientific Research (C), Grant Number JP19K05609.
Acknowledgments
Tetsuo Asakura acknowledges support by Impulsing Paradigm Change through Disruptive Technologies Program (ImPACT) and a JSPS KAKENHI, Grant-in-Aid for Scientific Research (C), Grant Number JP19K05609a JSPS KAKENHI, Grant-in-Aid for Scientific Research (C), Grant Number JP19K05609.
Conflicts of Interest
There is no conflict of interest associated with the author of this paper.
References
- Fu, C.; Shao, Z.; Fritz, V. Animal silks: Their structures, properties, and artificial production. Chem. Commun. 2009, 37, 6515. [Google Scholar] [CrossRef] [PubMed]
- Asakura, T.; Suzuki, Y.; Kametani, S. Silk. Encycl. Polym. Sci. Technol. 2018, 124, 8794–8795. [Google Scholar]
- Gosline, J.M.; Demont, M.; Denny, M.W. The structure and properties of spider silk. Endeavour 1986, 10, 37–43. [Google Scholar] [CrossRef]
- Xu, M.; Lewis, R.V. Structure of a protein superfiber: Spider dragline silk. Proc. Natl. Acad. Sci. USA 1990, 87, 7120–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisoldt, L.; Smith, A.; Scheibel, T. Decoding the secrets of spider silk. Mater. Today 2011, 14, 80–86. [Google Scholar] [CrossRef]
- Tokareva, O.; Jacobsen, M.M.; Buehler, M.; Wong, J.; Kaplan, D. Structure-function-property-design interplay in biopolymers: Spider silk. Acta Biomater. 2013, 10, 1612–1626. [Google Scholar] [CrossRef] [Green Version]
- Koh, L.-D.; Cheng, Y.; Teng, C.-P.; Khin, Y.-W.; Loh, X.-J.; Tee, S.-Y.; Low, M.; Ye, E.; Yu, H.-D.; Zhang, Y.-W.; et al. Structures, mechanical properties and applications of silk fibroin materials. Prog. Polym. Sci. 2015, 46, 86–110. [Google Scholar] [CrossRef]
- Work, R.W. Viscoelastic Behaviour and Wet Supercontraction of Major Ampullate Silk Fibres of Certain Orb-Web-Building Spiders (Araneae). J. Exp. Biol. 1985, 118, 379–404. [Google Scholar]
- Savage, K.N.; Gosline, J. The effect of proline on the network structure of major ampullate silks as inferred from their mechanical and optical properties. J. Exp. Boil. 2008, 211, 1937–1947. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sponner, A.; Porter, D.; Vollrath, F. Proline and Processing of Spider Silks. Biomacromolecules 2008, 9, 116–121. [Google Scholar] [CrossRef]
- Aigner, T.B.; de Simone, E.; Scheibel, T. Biomedical Applications of Recombinant Silk-Based Materials. Adv. Mater. 2018, 30, 1704636. [Google Scholar] [CrossRef] [PubMed]
- Asakura, T.; Miller, T. Biotechnology of Silk. In Biologically-Inspired Systems; Asakura, T., Miller, T., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-007-7118-5. [Google Scholar]
- Lucas, F.; Shaw, J.T.; Smith, S.G. Comparative studies of fibroins I. The amino acid composition of various fibroins and its significance in relation to their crystal structure and taxonomy. J. Mol. Boil. 1960, 2, 339–349. [Google Scholar] [CrossRef]
- Work, R.W.; Young, C.T. The Amino Acid Compositions of Major and Minor Ampullate Silks of Certain Orb-Web-Building Spiders (Araneae, Araneidae). J. Arachnol. 1987, 15, 65–80. [Google Scholar]
- Grubb, D.T.; Jelinski, L.W. Fiber Morphology of Spider Silk: The Effects of Tensile Deformation. Macromolecules 1997, 30, 2860–2867. [Google Scholar] [CrossRef]
- Riekel, C.; Müller, M.; Vollrath, F. In Situ X-ray Diffraction during Forced Silking of Spider Silk. Macromolecules 1999, 32, 4464–4466. [Google Scholar] [CrossRef]
- Riekel, C.; Madsen, B.; Knight, D.; Vollrath, F. X-ray diffraction on spider silk during controlled extrusion under a synchrotron radiation X-ray beam. Biomacromolecules 2000, 1, 622–626. [Google Scholar] [CrossRef]
- Trancik, J.; Czernuszka, J.; Bell, F.; Viney, C. Nanostructural features of a spider dragline silk as revealed by electron and X-ray diffraction studies. Polymer 2006, 47, 5633–5642. [Google Scholar] [CrossRef]
- Glišović, A.; Salditt, T. Temperature dependent structure of spider silk by X-ray diffraction. Appl. Phys. A 2007, 87, 63–69. [Google Scholar] [CrossRef]
- Rousseau, M.-E.; Hernández-Cruz, D.; West, M.M.; Hitchcock, A.P.; Pézolet, M. Nephila clavipes Spider Dragline Silk Microstructure Studied by Scanning Transmission X-ray Microscopy. J. Am. Chem. Soc. 2007, 129, 3897–3905. [Google Scholar] [CrossRef]
- Sampath, S.; Isdebski, T.; Jenkins, J.E.; Ayon, J.V.; Henning, R.W.; Orgel, J.P.R.O.; Antipoa, O.; Yarger, J.L. X-ray diffraction study of nanocrystalline and amorphous structure within major and minor ampullate dragline spider silks. Soft Matter 2012, 8, 6713–6722. [Google Scholar] [CrossRef] [Green Version]
- Benmore, C.J.; Izdebski, T.; Yarger, J.L. Total X-Ray Scattering of Spider Dragline Silk. Phys. Rev. Lett. 2012, 108, 178102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madurga, R.; Blackledge, T.A.; Perea, B.; Plaza, G.R.; Riekel, C.; Burghammer, M.; Elices, M.; Guinea, G.; Pérez-Rigueiro, J. Persistence and variation in microstructural design during the evolution of spider silk. Sci. Rep. 2015, 5, 14820. [Google Scholar] [CrossRef] [Green Version]
- Riekel, C.; Burghammer, M.; Dane, T.; Ferrero, C.; Rosenthal, M. Nanoscale Structural Features in Major Ampullate Spider Silk. Biomacromolecules 2016, 18, 231–241. [Google Scholar] [CrossRef]
- Yazawa, K.; Malay, A.D.; Masunaga, H.; Norma-Rashid, Y.; Numata, K. Simultaneous effect of strain rate and humidity on the structure and mechanical behavior of spider silk. Commun. Mater. 2020, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Simmons, A.; Ray, E.; Jelinski, L.W. Solid-State 13C NMR of Nephila clavipes Dragline Silk Establishes Structure and Identity of Crystalline Regions. Macromolecules 1994, 27, 5235–5237. [Google Scholar] [CrossRef]
- Simmons, A.H.; Michal, C.A.; Jelinski, L.W. Molecular Orientation and Two-Component Nature of the Crystalline Fraction of Spider Dragline Silk. Science 1996, 271, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Kümmerlen, J.; van Beek, J.D.; Vollrath, F.; Meier, B.H. Local Structure in Spider Dragline Silk Investigated by Two-Dimensional Spin-Diffusion Nuclear Magnetic Resonance. Macromolecules 1996, 29, 2920–2928. [Google Scholar] [CrossRef] [Green Version]
- Michal, C.A.; Jelinski, L.W. Rotational-echo double-resonance in complex biopolymers: A study of Nephila clavipes dragline silk. J. Biomol. NMR 1998, 12, 231–241. [Google Scholar] [CrossRef]
- Yang, Z.; Liivak, O.; Seidel, A.; LaVerde, G.; Zax, D.B.; Jelinski, L.W. Supercontraction and Backbone Dynamics in Spider Silk: 13C and 2H NMR Studies. J. Am. Chem. Soc. 2000, 122, 9019–9025. [Google Scholar] [CrossRef]
- Kishore, A.I.; Herberstein, M.; Craig, C.L.; Separovic, F. Solid-state NMR relaxation studies of Australian spider silks. Biopolymers 2002, 61, 287–297. [Google Scholar] [CrossRef]
- Van Beek, J.D.; Hess, S.; Vollrath, F.; Meier, B.H. The molecular structure of spider dragline silk: Folding and orientation of the protein backbone. Proc. Natl. Acad. Sci. USA 2002, 99, 10266–10271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, J.; Ohgo, K.; Komatsu, K.; Kubota, A.; Asakura, T. Determination of the torsion angles of alanine and glycine residues of model compounds of spider silk (AGG) 10 using solid-state NMR methods. J. Biomol. NMR 2003, 25, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Eles, P.T.; Michal, C.A. A Decoder NMR Study of Backbone Orientation in Nephila clavipes Dragline Silk under Varying Strain and Draw Rate. Biomacromolecules 2004, 5, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Eles, P.T.; Michal, C.A. Strain Dependent Local Phase Transitions Observed during Controlled Supercontraction Reveal Mechanisms in Spider Silk. Macromolecules 2004, 37, 1342–1345. [Google Scholar] [CrossRef]
- Hronska, M.; van Beek, J.D.; Williamson, P.T.F.; Vollrath, F.; Meier, B.H. NMR Characterization of Native Liquid Spider Dragline Silk from Nephila edulis. Biomacromolecules 2004, 5, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Lewis, R.V.; Yarger, J.L. Wise NMR Characterization of Nanoscale Heterogeneity and Mobility in Supercontracted Nephila clavipes Spider Dragline Silk. J. Am. Chem. Soc. 2004, 126, 5867–5872. [Google Scholar] [CrossRef]
- Asakura, T.; Yang, M.; Kawase, T.; Nakazawa, Y. 13C Solid-State NMR Study of Structural Heterogeneity in Peptides Containing Both Polyalanine and Repeated GGA Sequences as a Local Structural Model of Nephila clavipes Dragline Silk (Spidroin 1). Macromolecules 2005, 38, 3356–3363. [Google Scholar] [CrossRef]
- Yang, M.; Nakazawa, Y.; Yamauchi, K.; Knight, D.P.; Asakura, T. Structure of model peptides based on Nephila clavipes dragline silk spidroin (MaSp1) studied by 13C cross polarization/magic angle spinning NMR. Biomacromolecules 2005, 6, 3220–3226. [Google Scholar] [CrossRef]
- Bonev, B.B.; Grieve, S.M.; Herberstein, M.; Kishore, A.I.; Watts, A.; Separovic, F. Orientational order of Australian spider silks as determined by solid-state NMR. Biopolymer 2006, 82, 134–143. [Google Scholar] [CrossRef]
- Ohgo, K.; Kawase, T.; Ashida, J.; Asakura, T. Solid-State NMR Analysis of a Peptide (Gly-Pro-Gly-Gly-Ala)6-Gly Derived from a Flagelliform Silk Sequence of Nephila clavipes. Biomacromolecules 2006, 7, 1210–1214. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, I.; van Beek, J.D.; Meier, B.H. Molecular Disorder and Structure of Spider Dragline Silk Investigated by Two-Dimensional Solid-State NMR Spectroscopy. Macromolecules 2007, 40, 1995–2001. [Google Scholar] [CrossRef]
- Holland, G.P.; Jenkins, J.E.; Creager, M.S.; Lewis, R.V.; Yarger, J.L. Solid-State NMR Investigation of Major and Minor Ampullate Spider Silk in the Native and Hydrated States. Biomacromolecules 2008, 9, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Creager, M.S.; Jenkins, J.E.; Lewis, R.V.; Yarger, J.L. Determining Secondary Structure in Spider Dragline Silk by Carbon−Carbon Correlation Solid-State NMR Spectroscopy. J. Am. Chem. Soc. 2008, 130, 9871–9877. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Jenkins, J.E.; Creager, M.S.; Lewis, R.V.; Yarger, J.L. Quantifying the fraction of glycine and alanine in β-sheet and helical conformations in spider dragline silk using solid-state NMR. Chem. Commun. 2008, 5568–5570. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, E.; Yamauchi, K.; Gullion, T.; Asakura, T. Structural analysis of the Gly-rich region in spider dragline silk using stable-isotope labeled sequential model peptides and solid-state NMR. Chem. Commun. 2009, 4176. [Google Scholar] [CrossRef]
- Jenkins, J.E.; Creager, M.S.; Butler, E.B.; Lewis, R.V.; Yarger, J.L.; Holland, G.P. Solid-state NMR evidence for elastin-like β-turn structure in spider dragline silk. Chem. Commun. 2010, 46, 6714. [Google Scholar] [CrossRef]
- Jenkins, J.E.; Creager, M.S.; Lewis, R.V.; Holland, G.P.; Yarger, J.L. Quantitative Correlation between the Protein Primary Sequences and Secondary Structures in Spider Dragline Silks. Biomacromolecules 2010, 11, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Izdebski, T.; Akhenblit, P.; Jenkins, J.E.; Yarger, J.L.; Holland, G.P. Structure and Dynamics of Aromatic Residues in Spider Silk: 2D Carbon Correlation NMR of Dragline Fibers. Biomacromolecules 2010, 11, 168–174. [Google Scholar] [CrossRef]
- Holland, G.P.; Cherry, B.R.; Jenkins, J.E.; Yarger, J.L. Proton-detected heteronuclear single quantum correlation NMR spectroscopy in rigid solids with ultra-fast MAS. J. Magn. Reson. 2009, 202, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Yazawa, K.; Yamaguchi, E.; Knight, D.; Asakura, T. 13C solid-state NMR study of the 13C-labeled peptide, (E)8GGLGGQGAG(A)6GGAGQGGYGG as a model for the local structure of Nephila clavipes dragline silk (MaSp1) before and after spinning. Biopolymers 2011, 97, 347–354. [Google Scholar] [CrossRef]
- Asakura, T.; Okonogi, M.; Horiguchi, K.; Aoki, A.; Saitô, H.; Knight, D.P.; Williamson, M.P. Two Different Packing Arrangements of Antiparallel Polyalanine. Angew. Chem. Int. Ed. 2012, 51, 1212–1215. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.E.; Sampath, S.; Butler, E.; Kim, J.; Henning, R.W.; Holland, G.P.; Yarger, J.L. Characterizing the Secondary Protein Structure of Black Widow Dragline Silk Using Solid-State NMR and X-ray Diffraction. Biomacromolecules 2013, 14, 3472–3483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Yarger, J.L.; Holland, G.P. Elucidating proline dynamics in spider dragline silk fibre using 2H-13C HETCOR MAS NMR. Chem. Commun. 2014, 50, 4856–4859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Shi, X.; Thompson, F.; Weber, W.S.; Mou, Q.; Yarger, J.L. Protein secondary structure of Green Lynx spider dragline silk investigated by solid-state NMR and X-ray diffraction. Int. J. Boil. Macromol. 2015, 81, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Gray, G.M.; van der Vaart, A.; Guo, C.; Jones, J.; Onofrei, D.; Cherry, B.R.; Lewis, R.V.; Yarger, J.L.; Holland, G.P. Secondary Structure Adopted by the Gly-Gly-X Repetitive Regions of Dragline Spider Silk. Int. J. Mol. Sci. 2016, 17, 2023. [Google Scholar] [CrossRef] [Green Version]
- Tasei, Y.; Nishimura, A.; Suzuki, Y.; Sato, T.K.; Sugahara, J.; Asakura, T. NMR Investigation about Heterogeneous Structure and Dynamics of Recombinant Spider Silk in the Dry and Hydrated States. Macromolecules 2017, 50, 8117–8128. [Google Scholar] [CrossRef]
- Kametani, S.; Tasei, Y.; Nishimura, A.; Asakura, T. Distinct solvent- and temperature-dependent packing arrangements of anti-parallel β-Sheet polyalanines studied with solid-state 13 C NMR and MD simulation. Phys. Chem. Chem. Phys. 2017, 19, 20829–20838. [Google Scholar] [CrossRef]
- Asakura, T.; Tasei, Y.; Aoki, A.; Nishimura, A. Mixture of Rectangular and Staggered Packing Arrangements of Polyalanine Region in Spider Dragline Silk in Dry and Hydrated States as Revealed by 13C NMR and X-ray Diffraction. Macromolecules 2018, 51, 1058–1068. [Google Scholar] [CrossRef]
- Asakura, T.; Matsuda, H.; Kataoka, N.; Imai, A. Changes in the Local Structure of Nephila clavipes Dragline Silk Model Peptides upon Trifluoroacetic Acid, Low pH, Freeze-Drying, and Hydration Treatments Studied by 13C Solid-State NMR. Biomacromolecules 2018, 19, 4396–4410. [Google Scholar] [CrossRef]
- Asakura, T.; Tasei, Y.; Matsuda, H.; Naito, A. Dynamics of Alanine Methyl Groups in Alanine Oligopeptides and Spider Dragline Silks with Different Packing Structures as Studied by 13C Solid-State NMR Relaxation. Macromolecules 2018, 51, 6746–6756. [Google Scholar] [CrossRef]
- Asakura, T.; Nishimura, A.; Tasei, Y. Determination of Local Structure of 13C Selectively Labeled 47-mer Peptides as a Model for Gly-Rich Region of Nephila clavipes Dragline Silk Using a Combination of 13C Solid-State NMR and MD Simulation. Macromolecules 2018, 51, 3608–3619. [Google Scholar] [CrossRef]
- Asakura, T.; Nishimura, A.; Aoki, A.; Naito, A. Packing Structure of Antiparallel β-Sheet Polyalanine Region in a Sequential Model Peptide of Nephila clavipes Dragline Silk Studied Using 13C Solid-State NMR and MD Simulation. Biomacromolecules 2019, 20, 3884–3894. [Google Scholar] [CrossRef]
- Asakura, T.; Okonogi, M.; Naito, A. Toward Understanding the Silk Fiber Structure: 13C Solid-State NMR Studies of the Packing Structures of Alanine Oligomers before and after Trifluoroacetic Acid Treatment. J. Phys. Chem. B 2019, 123, 6716–6727. [Google Scholar] [CrossRef] [PubMed]
- Asakura, T.; Matsuda, H.; Aoki, A.; Kataoka, N.; Imai, A. Conformational change of 13C-labeled 47-mer model peptides of Nephila clavipes dragline silk in poly(vinyl alcohol) film by stretching studied by 13C solid-state NMR and molecular dynamics simulation. Int. J. Boil. Macromol. 2019, 131, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Asakura, T.; Okushita, K.; Williamson, M.P. Analysis of the Structure of Bombyx mori Silk Fibroin by NMR. Macromolecules 2015, 48, 2345–2357. [Google Scholar] [CrossRef]
- Saitô, H. Conformation-dependent 13C chemical shifts: A new means of conformational characterization as obtained by high-resolution solid-state 13C NMR. Magn. Reson. Chem. 1986, 24, 835–852. [Google Scholar] [CrossRef]
- Saitô, H.; Ando, I. High-Resolution Solid-State NMR Studies of Synthetic and Biological Macromolecules. Ann. Rep. NMR Spectrosc. 1989, 21, 209–290. [Google Scholar] [CrossRef]
- Spera, S.; Bax, A. Empirical correlation between protein backbone conformation and Cα and Cβ 13C nuclear magnetic resonance chemical shifts. J. Am. Chem. Soc. 1991, 113, 5490–5492. [Google Scholar] [CrossRef]
- Iwadate, M.; Asakura, T.; Williamson, M.P. Cα and Cβ Carbon-13 Chemical Shifts in Proteins from an Empirical Database. J. Biomol. NMR 1999, 13, 199–211. [Google Scholar] [CrossRef]
- Asakura, T.; Iwadate, M.; Demura, M.; Williamson, M.P. Structural analysis of silk with 13C NMR chemical shift contour plots. Int. J. Boil. Macromol. 1999, 24, 167–171. [Google Scholar] [CrossRef]
- Wishart, D.S. Interpreting protein chemical shift data. Prog. Nucl. Magn. Reson. Spectrosc. 2011, 58, 62–87. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Umemura, K.; Asakura, T. The Structural Characteristics of Bombyx mori Silk Fibroin before Spinning as Studied with Molecular Dynamics Simulation. Macromolecules 2002, 35, 8831–8838. [Google Scholar] [CrossRef]
- Yamane, T.; Umemura, K.; Nakazawa, Y.; Asakura, T. Molecular dynamics simulation of conformational change of poly (Ala-Gly) from silk I to silk II in relation to fiber formation mechanism of Bombyx mori silk fibroin. Macromolecules 2003, 36, 6766–6772. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, M. Molecular dynamics simulations of alanine rich β-sheet oligomers: Insight into amyloid formation. Protein Sci. 2009, 11, 2335–2350. [Google Scholar] [CrossRef]
- Keten, S.; Buehler, M.J. Nanostructure, and molecular mechanics of spider dragline silk protein assemblies. J. R. Soc. Interface 2010, 7, 1709–1721. [Google Scholar] [CrossRef] [Green Version]
- Bratzel, G.; Buehler, M.J. Sequence-structure correlations in silk: Poly-Ala repeat of N. clavipes MaSp1 is naturally optimized at a critical length scale. J. Mech. Behav. Biomed. Mater. 2012, 7, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.P.; Markert, B.; Gräter, F. Rate-Dependent Behavior of the Amorphous Phase of Spider Dragline Silk. Biophys. J. 2014, 106, 2511–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Koh, L.-D.; Li, D.; Ji, B.; Han, M.-Y.; Zhang, Y.-W. On the strength of β-sheet crystallites of Bombyx mori silk fibroin. J. R. Soc. Interface 2014, 11, 20140305. [Google Scholar] [CrossRef] [Green Version]
- Ukpebor, O.T.; Shah, A.; Bazov, E.; Boutis, G.S.; Bazov, E. Inverse temperature transition of elastin like motifs in major ampullate dragline silk: MD simulations of short peptides and NMR studies of water dynamics. Soft Matter 2014, 10, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Giesa, T.; Perry, C.C.; Buehler, M.J. Secondary Structure Transition and Critical Stress for a Model of Spider Silk Assembly. Biomacromolecules 2016, 17, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Sintya, E.; Alam, P. Self-assembled semi-crystallinity at parallel β -sheet nanocrystal interfaces in clustered MaSp1 (spider silk) proteins. Mater. Sci. Eng. C 2016, 58, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Giesa, T.; Schuetz, R.; Fratzl, P.; Buehler, M.J.; Masic, A. Unraveling the Molecular Requirements for Macroscopic Silk Supercontraction. ACS Nano 2017, 11, 9750–9758. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Tarakanova, A.; Hsu, C.C.; Yu, M.; Zheng, S.; Yu, L.; Liu, J.; He, Y.; Dunstan, D.J.; Buehler, M.J. Spider dragline silk as torsional actuator driven by humidity. Sci. Adv. 2019, 5, e9183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, M.; Dubey, D.K.; Singh, S.P. Phenomenological models of Bombyx mori silk fibroin and their mechanical behavior using molecular dynamics simulations. Mater. Sci. Eng. C 2020, 108, 110414. [Google Scholar] [CrossRef]
- Zhao, C.; Han, S. Structure of Silk studied with NMR. Prog. Nucl. Magn. Reson. Spectrosc. 2001, 39, 301–352. [Google Scholar] [CrossRef]
- Kameda, T.; Asakura, T. Dynamics of silk fibroin studied with NMR spectroscopy. Ann. Rep. NMR Spectrosc. 2002, 46, 101–149. [Google Scholar] [CrossRef]
- Asakura, T.; Nakazawa, Y. Structure and Structural Changes of the Silk Fibroin from Samia cynthia ricini Using Nuclear Magnetic Resonance Spectroscopy. Macromol. Biosci. 2004, 4, 175–185. [Google Scholar] [CrossRef]
- Asakura, T.; Suzuki, Y.; Nakazawa, Y.; Yazawa, K.; Holland, G.P.; Yarger, J.L. Silk structure studied with nuclear magnetic resonance. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 69, 23–68. [Google Scholar] [CrossRef]
- Asakura, T.; Suzuki, Y.; Nakazawa, Y.; Holland, G.P.; Yarger, J.L. Elucidating silk structure using solid-state NMR. Soft Matter 2013, 9, 11440. [Google Scholar] [CrossRef]
- Asakura, T.; Tasei, Y. NMR Studies on Silk Materials. In Experimental Approaches of NMR Spectroscopy; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017; pp. 297–312. [Google Scholar]
- Guo, C.; Yarger, J.L. Chapter 18. NMR Characterization of Silk. In New Developments in NMR; Royal Society of Chemistry (RSC): London, UK, 2019; pp. 420–456. [Google Scholar]
- Hinman, M.B.; Lewis, R. V Isolation of a clone encoding a second dragline silk fibroin. Nephila clavipes dragline silk is a two-protein fiber. J. Biol. Chem. 1992, 267, 19320–19324. [Google Scholar]
- Nagano, A.; Sato, H.; Tanioka, Y.; Nakazawa, Y.; Knight, D.; Asakura, T. Characterization of a Ca binding-amphipathic silk-like protein and peptide with the sequence (Glu)8(Ala-Gly-Ser-Gly-Ala-Gly)4 with potential for bone repair. Soft Matter 2012, 8, 741–748. [Google Scholar] [CrossRef]
- Gullion, T.; Schaefer, J. Rotational-echo double-resonance NMR. J. Magn. Reson. 1989, 81, 196–200. [Google Scholar] [CrossRef]
- Pan, Y.; Gullion, T.; Schaefer, J. Determination of CN internuclear distances by rotational-echo double-resonance NMR of solids. J. Magn. Reson. 1990, 90, 330–340. [Google Scholar] [CrossRef]
- Lotz, B.; Keith, H. The crystal structures of poly (lAla-Gly-Gly-Gly) II and poly (lAla-Gly-Gly) II. J. Mol. Boil. 1971, 61, 195–200. [Google Scholar] [CrossRef]
- Lee, N.-K.; Ramamoorthy, A. Determination of the Solid-State Conformations of Polyalanine Using Magic-Angle Spinning NMR Spectroscopy. J. Phys. Chem. B 1999, 103, 271–275. [Google Scholar] [CrossRef]
- Henzler-Wildman, K.; Lee, N.-K.; Ramamoorthy, A. Determination of α-helix and β-sheet stability in the solid state: A solid-state NMR investigation of poly (L-alanine). Biopolymer 2002, 64, 246–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burum, D.; Ernst, R. Net polarization transfer via a J-ordered state for signal enhancement of low-sensitivity nuclei. J. Magn. Reson. 1980, 39, 163–168. [Google Scholar] [CrossRef]
- Asakura, T.; Sato, Y.; Aoki, A. Stretching-Induced Conformational Transition of the Crystalline and Noncrystalline Domains of 13C-Labeled Bombyx mori Silk Fibroin Monitored by Solid State NMR. Macromolecules 2015, 48, 5761–5769. [Google Scholar] [CrossRef]
- Asakura, T.; Isobe, K.; Aoki, A.; Kametani, S. Conformation of Crystalline and Noncrystalline Domains of [3-13C]Ala-, [3-13C]Ser-, and [3-13C] Tyr-Bombyx mori Silk Fibroin in a Hydrated State Studied with 13C DD/MAS NMR. Macromolecules 2015, 48, 8062–8069. [Google Scholar] [CrossRef]
- Saitô, H.; Tuzi, S.; Tanio, M.; Naito, A. Dynamic aspects of membrane proteins and membrane-associated peptides as revealed by 13C NMR: Lessons from bacteriorhodopsin as an intact protein. In Annual Reports on NMR Spectroscopy; Elsevier BV: Amsterdam, The Netherlands, 2002; pp. 39–108. [Google Scholar]
- Asakura, T.; Endo, M.; Tasei, Y.; Ohkubo, T.; Hiraoki, T. Hydration of Bombyx mori silk cocoon, silk sericin and silk fibroin and their interactions with water as studied by 13C NMR and 2H NMR relaxation. J. Mater. Chem. B 2017, 5, 1624–1632. [Google Scholar] [CrossRef]
- Asakura, T.; Endo, M.; Fukuhara, R.; Tasei, Y. 13C NMR characterization of hydrated 13C labeled Bombyx mori silk fibroin sponges prepared using glycerin, poly(ethylene glycol diglycidyl ether) and poly(ethylene glycol) as porogens. J. Mater. Chem. B 2017, 5, 2152–2160. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Zhao, T.; Sima, Y.; Zhang, Y.; Nakagaki, K.; Miao, Y.; Shiomi, K.; Kajiura, Z.; Nagata, Y.; Nakagaki, M. Unique Molecular Architecture of Egg Case Silk Protein in a Spider, Nephila clavata. J. Biochem. 2005, 138, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shimizu, K.; Shiomi, K.; Kajiura, Z.; Nakagawa, M. cDNA cloning of Nephila clavata dragline silk (MaSp1) gene and comparison with the sequence of Bombyx mori fibroin heavy chain. Sanshi-Konchu Biotec. 2008, 77, 39–46. [Google Scholar] [CrossRef]
- Naito, A.; Ganapathy, S.; Akasaka, K.; McDowell, C. Spin-lattice relaxation of 13C in solid amino acids using the CP-MAS technique. J. Magn. Reson. 1983, 54, 226–235. [Google Scholar] [CrossRef]
- Naito, A.; Fukutani, A.; Uitdehaag, M.; Tuzi, S.; Saitô, H. Backbone dynamics of polycrystalline peptides studied by measurements of 15N NMR lineshapes and 13C transverse relaxation times. J. Mol. Struct. 1998, 441, 231–241. [Google Scholar] [CrossRef]
- Naito, A.; Tasei, Y.; Nishimura, A.; Asakura, T. Unusual Dynamics of Alanine Residues in Polyalanine Regions with Staggered Packing Structure of Samia cynthia ricini Silk Fiber in Dry and Hydrated States Studied by 13C Solid-State NMR and Molecular Dynamics Simulation. J. Phys. Chem. B 2018, 122, 6511–6520. [Google Scholar] [CrossRef]
- Arnott, S.; Dover, S.; Elliott, A. Structure of β-poly-l-alanine: Refined atomic co-ordinates for an anti-parallel beta-pleated sheet. J. Mol. Boil. 1967, 30, 201–208. [Google Scholar] [CrossRef]
- Asakura, T.; Nishimura, A.; Kametani, S.; Kawanishi, S.; Aoki, A.; Suzuki, F.; Kaji, H.; Naito, A. Refined Crystal Structure of Samia cynthia ricini Silk Fibroin Revealed by Solid-State NMR Investigations. Biomacromolecules 2017, 18, 1965–1974. [Google Scholar] [CrossRef]
- Asakura, T.; Miyazawa, K.; Tasei, Y.; Kametani, S.; Nakazawa, Y.; Aoki, A.; Naito, A. Packing arrangement of 13 C selectively labeled sequence model peptides of Samia cynthia ricini silk fibroin fibers studied by solid-state NMR. Phys. Chem. Chem. Phys. 2017, 19, 13379–13386. [Google Scholar] [CrossRef]
- Blamires, S.J.; Blackledge, T.A.; Tso, I.-M. Physicochemical Property Variation in Spider Silk: Ecology, Evolution, and Synthetic Production. Annu. Rev. Èntomol. 2017, 62, 443–460. [Google Scholar] [CrossRef]
- Craig, H.C.; Blamires, S.J.; Sani, M.-A.; Kasumovic, M.M.; Rawal, A.; Hook, J.M. DNP NMR spectroscopy reveals new structures, residues, and interactions in wild spider silks. Chem. Commun. 2019, 55, 4687–4690. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic overview of seven types of spider silks.

Figure 2.
13C CP/MAS NMR spectrum of dragline silk fiber from N. clavipes spider together with the assignment [26]. The chemical shifts are represented from tetramethylsilane (TMS).
Figure 2.
13C CP/MAS NMR spectrum of dragline silk fiber from N. clavipes spider together with the assignment [26]. The chemical shifts are represented from tetramethylsilane (TMS).
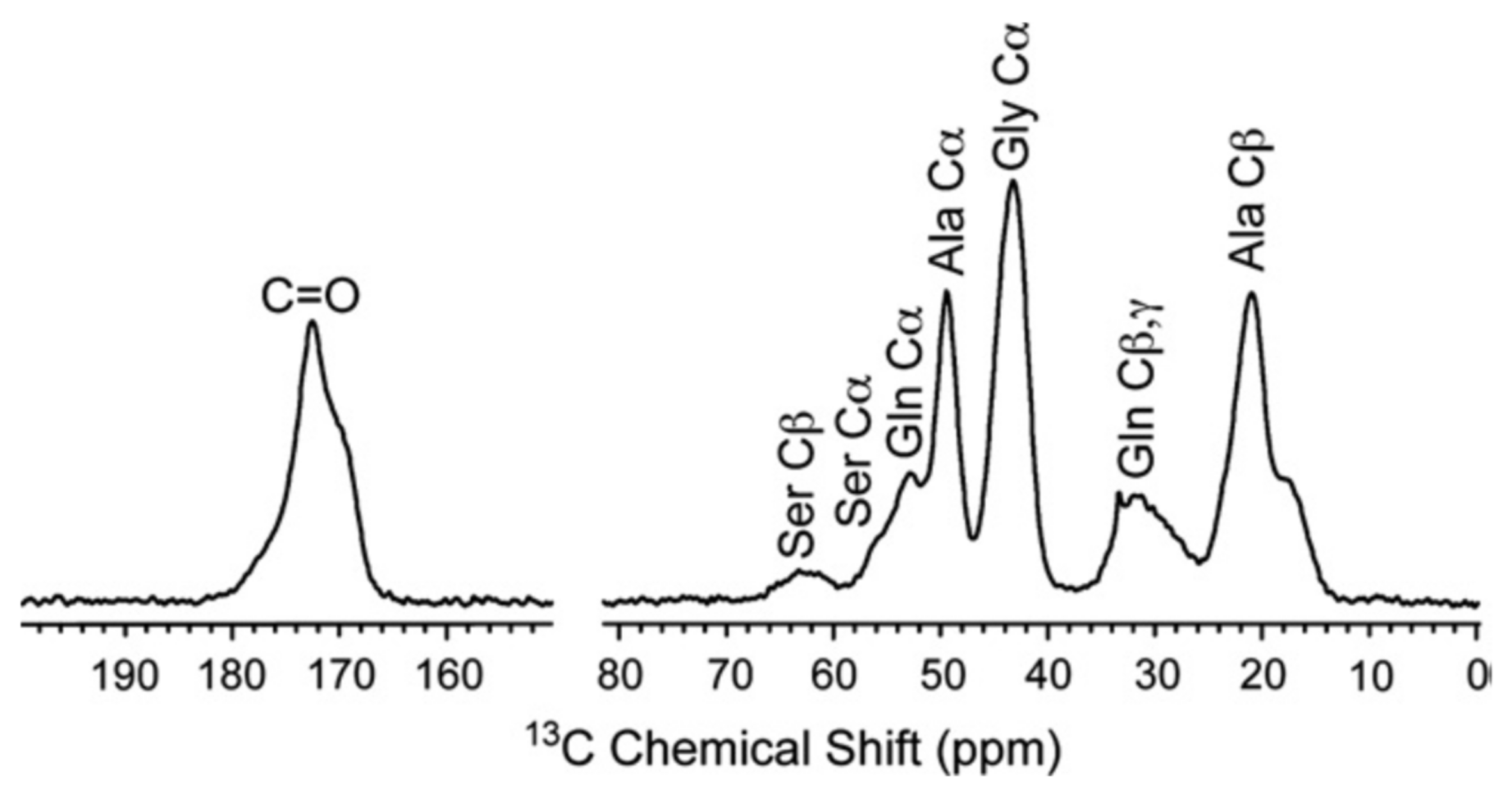
Figure 3.
Partial amino acid sequence of MaSp1 spider silk protein. Polyalanine sequences (red) forms antiparallel β-sheet structure [4]. The underlined sequence, a, is the selected sequence to study the conformations of amino acid residues in the Gly-rich region sandwiched by poly-A sequence on both sides reported previously [46,60,62]. The underlined sequence, b, is selected to study the conformation and packing structure of poly-A sequence sandwiched by partial Gly-rich sequences [63].
Figure 3.
Partial amino acid sequence of MaSp1 spider silk protein. Polyalanine sequences (red) forms antiparallel β-sheet structure [4]. The underlined sequence, a, is the selected sequence to study the conformations of amino acid residues in the Gly-rich region sandwiched by poly-A sequence on both sides reported previously [46,60,62]. The underlined sequence, b, is selected to study the conformation and packing structure of poly-A sequence sandwiched by partial Gly-rich sequences [63].

Figure 4.
Plot of the REDOR-determined inter-nuclear distances of the 13C–15N nuclei in Table 1(I) vs. the corresponding distances calculated from the PDB database. The red circles indicate the inter-nuclear distances between [1-13C]Gly and [15N]Gly nuclei in the [1-13C]Gly-X-[15N]Gly (X = Q,Y,L, and R) motifs determined by REDOR experiment in Table 1(I) and those calculated from the PDB database. The grey hollow circles are corresponding distances of typical conformations: anti-parallel β-sheet; 4.6 Å, α-helix and type II β-turn; 3.2 Å and 31-helix; 4.2 Å [46].
Figure 4.
Plot of the REDOR-determined inter-nuclear distances of the 13C–15N nuclei in Table 1(I) vs. the corresponding distances calculated from the PDB database. The red circles indicate the inter-nuclear distances between [1-13C]Gly and [15N]Gly nuclei in the [1-13C]Gly-X-[15N]Gly (X = Q,Y,L, and R) motifs determined by REDOR experiment in Table 1(I) and those calculated from the PDB database. The grey hollow circles are corresponding distances of typical conformations: anti-parallel β-sheet; 4.6 Å, α-helix and type II β-turn; 3.2 Å and 31-helix; 4.2 Å [46].
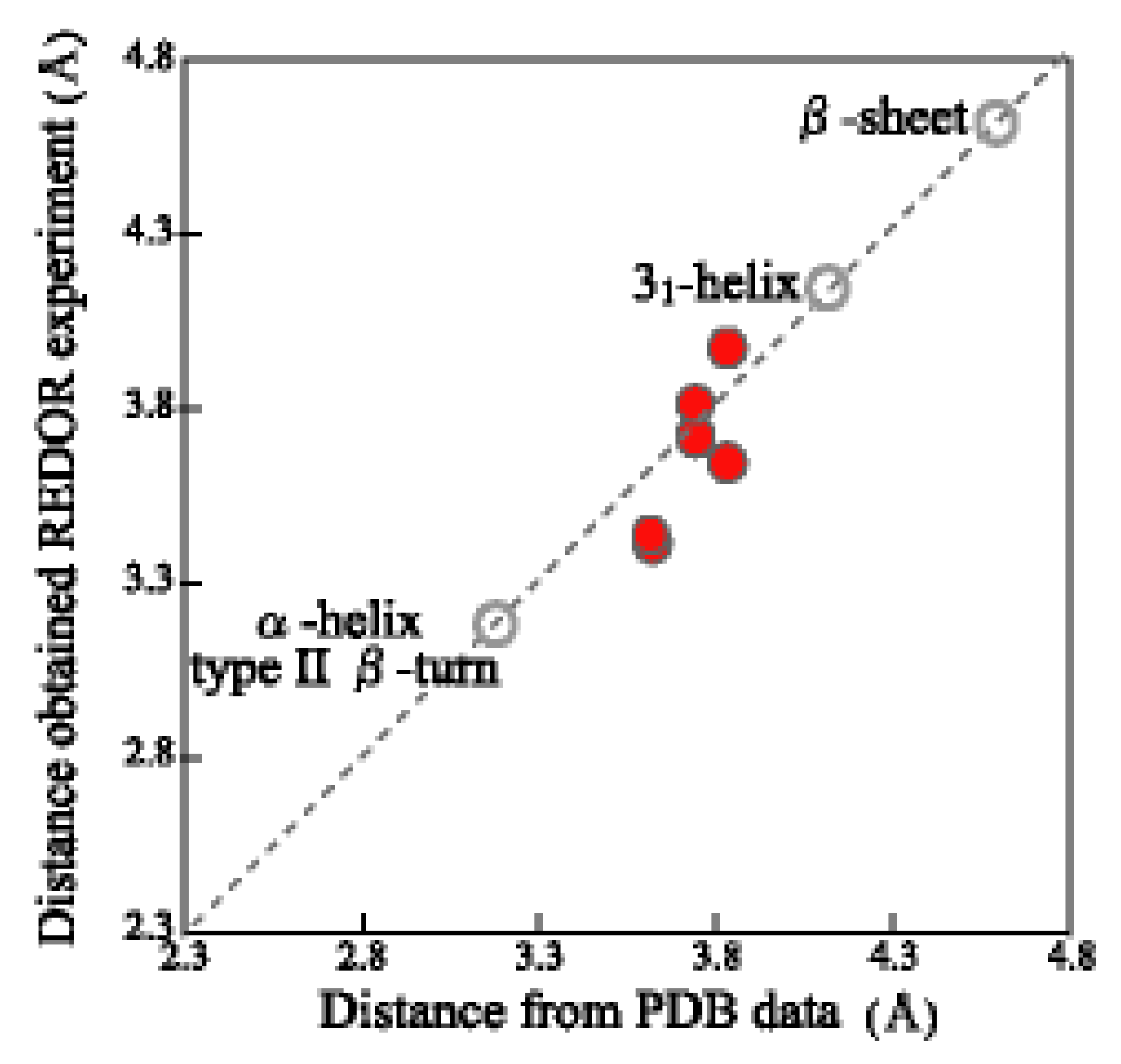
Figure 5.
13C CP/MAS NMR spectra of 13C successively triple-labeled 47-mer peptides incorporated as the [2-13C]Gly [3-13C]Ala[1-13C]Gly motif in different positions (Table 1 (II)): (a) non-labeled 47-mer peptides (Sample 1), (b) [2-13C]Gly12[3-13C]Ala13 [1-13C]Gly14 motif (Sample 8), (c) [2-13C]Gly25[3-13C]Ala26[1-13C]Gly27 motif (Sample 9), and (d) [2-13C]Gly35[3-13C]Ala36 [1-13C]Gly37 motif (Sample 10) [62]. The asterisk,* shows one of the natural abundance Ala Cβ, Cα, and C = O peaks of two (Ala)6 sequences in the 47-mer peptides.
Figure 5.
13C CP/MAS NMR spectra of 13C successively triple-labeled 47-mer peptides incorporated as the [2-13C]Gly [3-13C]Ala[1-13C]Gly motif in different positions (Table 1 (II)): (a) non-labeled 47-mer peptides (Sample 1), (b) [2-13C]Gly12[3-13C]Ala13 [1-13C]Gly14 motif (Sample 8), (c) [2-13C]Gly25[3-13C]Ala26[1-13C]Gly27 motif (Sample 9), and (d) [2-13C]Gly35[3-13C]Ala36 [1-13C]Gly37 motif (Sample 10) [62]. The asterisk,* shows one of the natural abundance Ala Cβ, Cα, and C = O peaks of two (Ala)6 sequences in the 47-mer peptides.
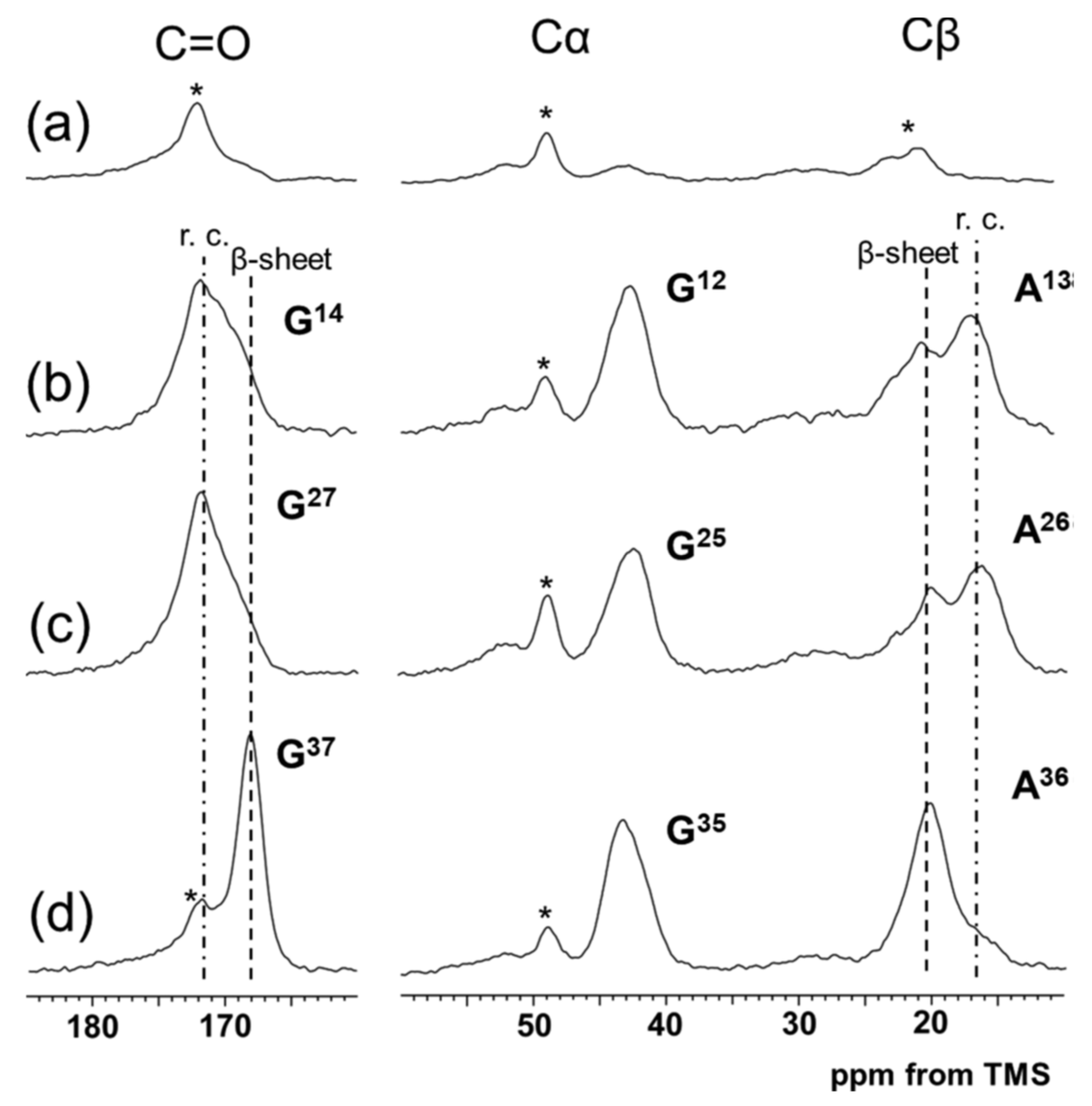
Figure 6.
(a) The fractions of several conformations, i.e., AP-β sheet, random coil and β-turn determined using 13C CP/MAS NMR for the 13C-labeled individual residues in 47-mer peptide, (E)6(A)6GGAGQGGYGGLGSQGAGRGGLGGQGAG(A)6(E)6 (Table 1 (I)) as the sequential model of the Gly-rich region (Figure 3a). The broken, dash-dotted and solid lines indicate change in the fractions of AP-β sheet, random coil and β-turn conformations, respectively as a function of residue number. (b) The fractions of several conformations calculated by the structural identification (STRIDE) algorithm, for the representative structure of the replica exchange molecular dynamics (REMD) simulations with the lowest temperature replica for the individual residues in the sequence, (A)6GGAGQGGYGGLGSQGAGRGGLGGQGAG(A)6 shown for a comparison [62]. The arrangement of the peptide chains with the sequences before MD simulation was as follows. The four extended peptide chains were oriented in antiparallel forms within the planes and parallel between the planes. Three parallel planes were assumed. The intermolecular hydrogen bonding systems of the (A)6 blocks were set in a staggered packing arrangement.
Figure 6.
(a) The fractions of several conformations, i.e., AP-β sheet, random coil and β-turn determined using 13C CP/MAS NMR for the 13C-labeled individual residues in 47-mer peptide, (E)6(A)6GGAGQGGYGGLGSQGAGRGGLGGQGAG(A)6(E)6 (Table 1 (I)) as the sequential model of the Gly-rich region (Figure 3a). The broken, dash-dotted and solid lines indicate change in the fractions of AP-β sheet, random coil and β-turn conformations, respectively as a function of residue number. (b) The fractions of several conformations calculated by the structural identification (STRIDE) algorithm, for the representative structure of the replica exchange molecular dynamics (REMD) simulations with the lowest temperature replica for the individual residues in the sequence, (A)6GGAGQGGYGGLGSQGAGRGGLGGQGAG(A)6 shown for a comparison [62]. The arrangement of the peptide chains with the sequences before MD simulation was as follows. The four extended peptide chains were oriented in antiparallel forms within the planes and parallel between the planes. Three parallel planes were assumed. The intermolecular hydrogen bonding systems of the (A)6 blocks were set in a staggered packing arrangement.
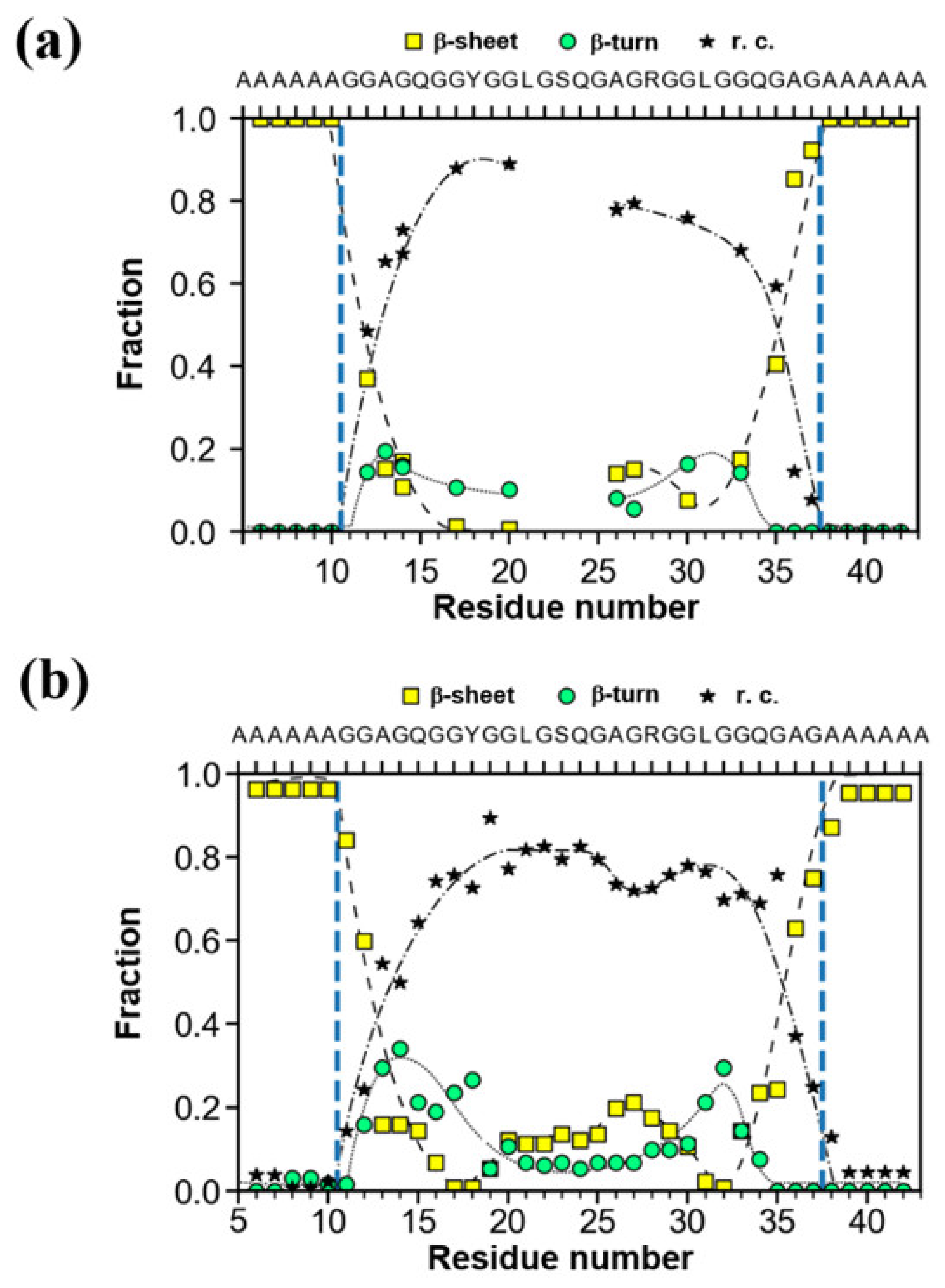
Figure 7.
(a) X-ray diffraction patterns and (b) Ala Cβ peaks in the 13C CP/MAS NMR spectra of alanine oligopeptides, (Ala)n (n = 4−8 and 12), together with those of N. clavata dragline silk fibers [52].
Figure 7.
(a) X-ray diffraction patterns and (b) Ala Cβ peaks in the 13C CP/MAS NMR spectra of alanine oligopeptides, (Ala)n (n = 4−8 and 12), together with those of N. clavata dragline silk fibers [52].

Figure 8.
(Left) Expanded dipolar dephasing 13C DD/MAS NMR spectra of the Ala Cβ regions of Gr(Ala)nGr* (n = 4 and 6−8) after FA/MeOH treatment [63]. The decomposed four spectra shown by broken lines were obtained by the deconvolutions under assuming the chemical shifts as 22.7, 21.2, 19.8 and 16.9 ppm from lower field to higher field, respectively. (Right) Pie charts of the fractions determined by the deconvolutions. The color of the fraction is ① red (22.7 ppm), ② green (21.2 ppm), ③ pink (19.8 ppm), and ④ light blue (16.9 ppm; right).
Figure 8.
(Left) Expanded dipolar dephasing 13C DD/MAS NMR spectra of the Ala Cβ regions of Gr(Ala)nGr* (n = 4 and 6−8) after FA/MeOH treatment [63]. The decomposed four spectra shown by broken lines were obtained by the deconvolutions under assuming the chemical shifts as 22.7, 21.2, 19.8 and 16.9 ppm from lower field to higher field, respectively. (Right) Pie charts of the fractions determined by the deconvolutions. The color of the fraction is ① red (22.7 ppm), ② green (21.2 ppm), ③ pink (19.8 ppm), and ④ light blue (16.9 ppm; right).

Figure 9.
(a) Plot of the fractions determined by the STRIDE algorithm, for the representative structure of the REMD simulation with the lowest temperature replica of the sequence, GGLGGQGAG(A)6G- GAGQGGYGG. (b) Image of the backbone structure after the MD simulation [63].
Figure 9.
(a) Plot of the fractions determined by the STRIDE algorithm, for the representative structure of the REMD simulation with the lowest temperature replica of the sequence, GGLGGQGAG(A)6G- GAGQGGYGG. (b) Image of the backbone structure after the MD simulation [63].
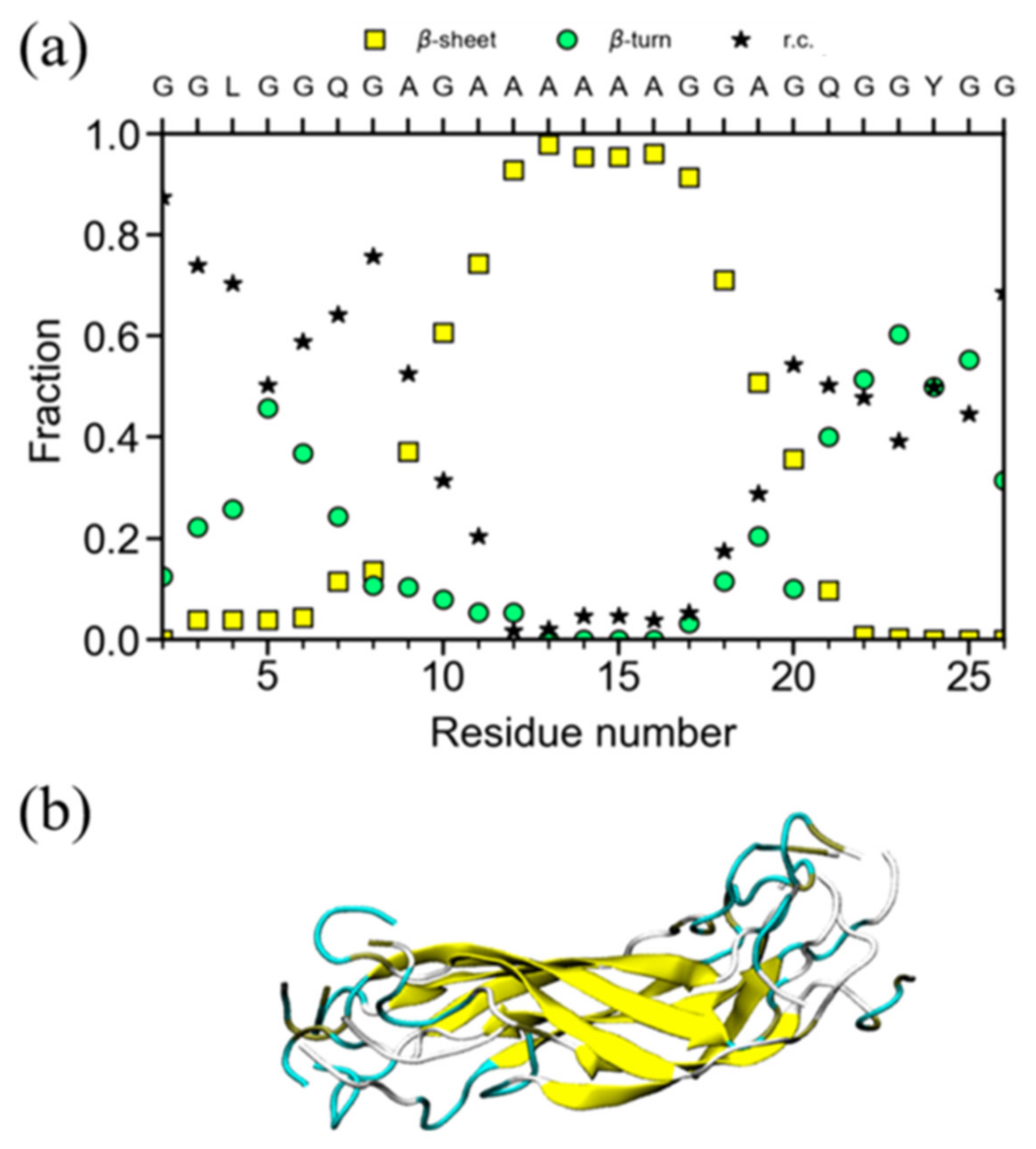
Figure 10.
Expanded aliphatic region from fully relaxed 13C directly detected MAS NMR spectrum of N. clavipes major ampullate silk in the hydrated state. The fraction of each peak was determined from the deconvolution shown by the broken lines [48].
Figure 10.
Expanded aliphatic region from fully relaxed 13C directly detected MAS NMR spectrum of N. clavipes major ampullate silk in the hydrated state. The fraction of each peak was determined from the deconvolution shown by the broken lines [48].

Figure 11.
13C solid-state NMR spectra (a–d) of 47-mer peptides with 13C-labeled motifs; (A) [2-13C]Gly25[3-13C]Ala26 [1-13C]Gly27 and (B) [2-13C]Gly35[3-13C]Ala36 [1-13C]Gly37. (a) 13C r-INEPT, (b) 13C DD/MAS and (c) 13C CP/MAS spectra in the hydrated state, and (d) 13C CP/MAS spectrum in the dry state [60]. The peaks marked by asterisk * were assigned to Ala Cα peaks of non-labeled (Ala)6 in the peptides.
Figure 11.
13C solid-state NMR spectra (a–d) of 47-mer peptides with 13C-labeled motifs; (A) [2-13C]Gly25[3-13C]Ala26 [1-13C]Gly27 and (B) [2-13C]Gly35[3-13C]Ala36 [1-13C]Gly37. (a) 13C r-INEPT, (b) 13C DD/MAS and (c) 13C CP/MAS spectra in the hydrated state, and (d) 13C CP/MAS spectrum in the dry state [60]. The peaks marked by asterisk * were assigned to Ala Cα peaks of non-labeled (Ala)6 in the peptides.

Figure 12.
Ramachandran maps showing the distributions of (Φ,Ψ) torsion angles of the residues in the motifs, Gly12Ala13Gly14, Gly25Ala26Gly27, and Gly35Ala36Gly37 of the Gly-rich region in the dry and hydrated states, respectively, as calculated by MD simulation [60].
Figure 12.
Ramachandran maps showing the distributions of (Φ,Ψ) torsion angles of the residues in the motifs, Gly12Ala13Gly14, Gly25Ala26Gly27, and Gly35Ala36Gly37 of the Gly-rich region in the dry and hydrated states, respectively, as calculated by MD simulation [60].
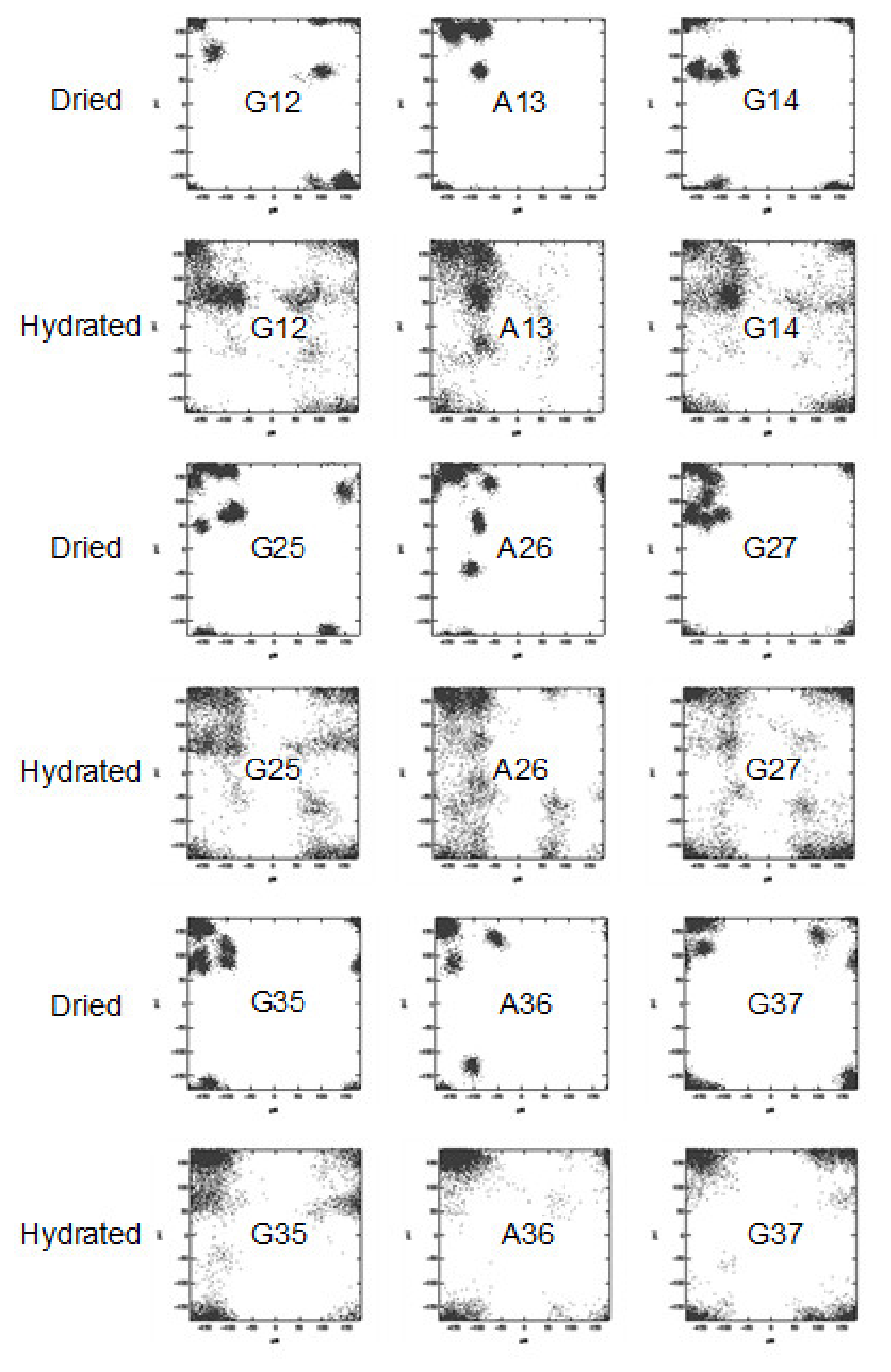
Figure 13.
Expanded 13C solid-state NMR spectra (10−70 ppm) of N. clavata dragline silk fibers together with assignments: (a) 13C refocused INEPT, (b) 13C DD/MAS, and (c) 13C CP/MAS spectra in the hydrated state and (d) 13C CP/MAS spectrum in the dry state [59].
Figure 13.
Expanded 13C solid-state NMR spectra (10−70 ppm) of N. clavata dragline silk fibers together with assignments: (a) 13C refocused INEPT, (b) 13C DD/MAS, and (c) 13C CP/MAS spectra in the hydrated state and (d) 13C CP/MAS spectrum in the dry state [59].
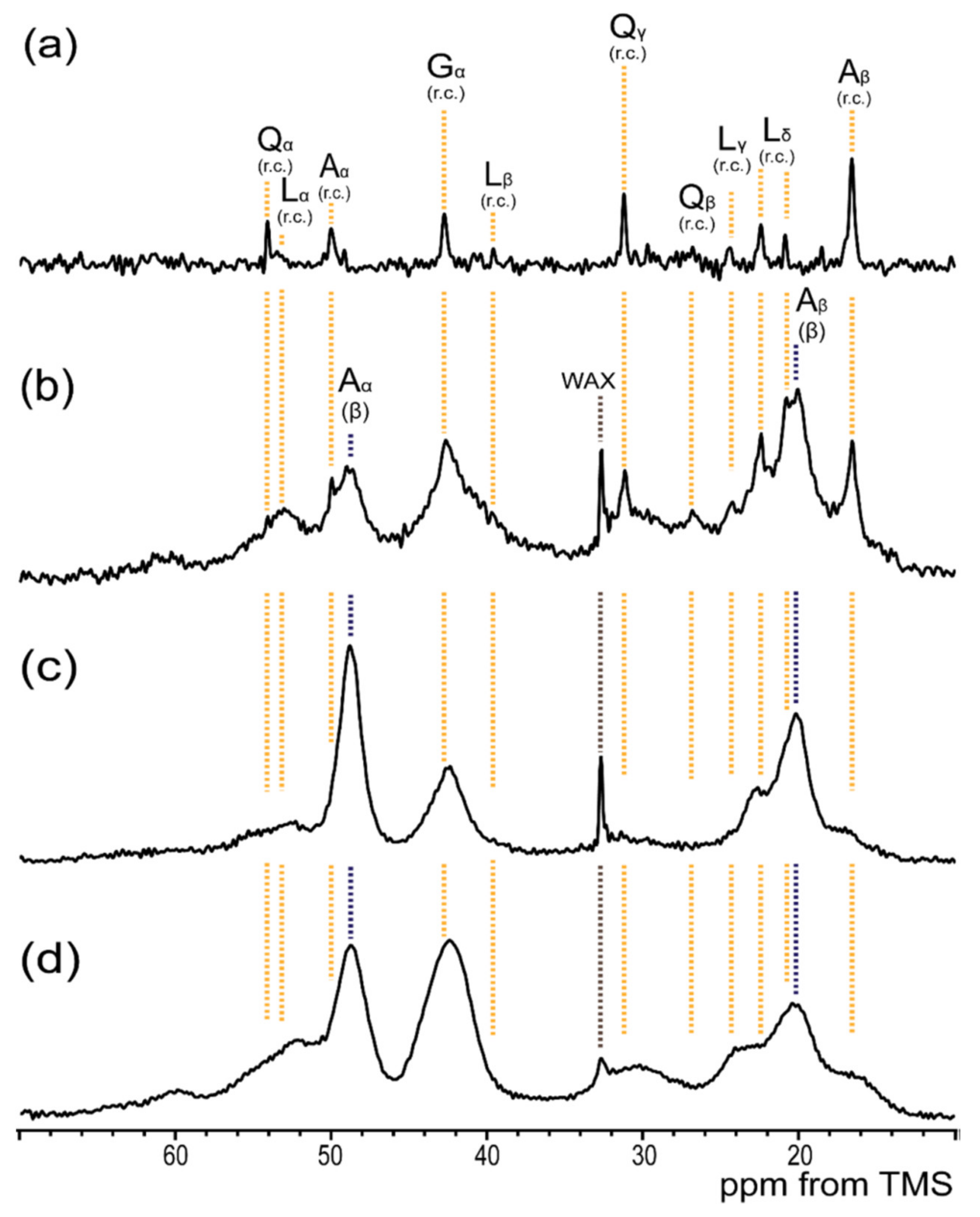
Figure 14.
(A) Expanded Ala Cβ peaks in the 13C CP/MAS spectra (10−30 ppm) of [3-13C]Ala-N. clavata dragline silk fibers in the dry and hydrated states together with peak deconvolution [59]. (A) The observed spectra (black solid lines) were deconvoluted by random coil (blue broken lines) and β-sheet structure (red solid lines). (B) The expanded Ala Cβ peaks with exclusively β-sheet structures were reproduced by superpositions (black broken lines) of the Ala Cβ peak patterns of (Ala)6 (orange broken lines) and (Ala)7 (green broken lines).
Figure 14.
(A) Expanded Ala Cβ peaks in the 13C CP/MAS spectra (10−30 ppm) of [3-13C]Ala-N. clavata dragline silk fibers in the dry and hydrated states together with peak deconvolution [59]. (A) The observed spectra (black solid lines) were deconvoluted by random coil (blue broken lines) and β-sheet structure (red solid lines). (B) The expanded Ala Cβ peaks with exclusively β-sheet structures were reproduced by superpositions (black broken lines) of the Ala Cβ peak patterns of (Ala)6 (orange broken lines) and (Ala)7 (green broken lines).
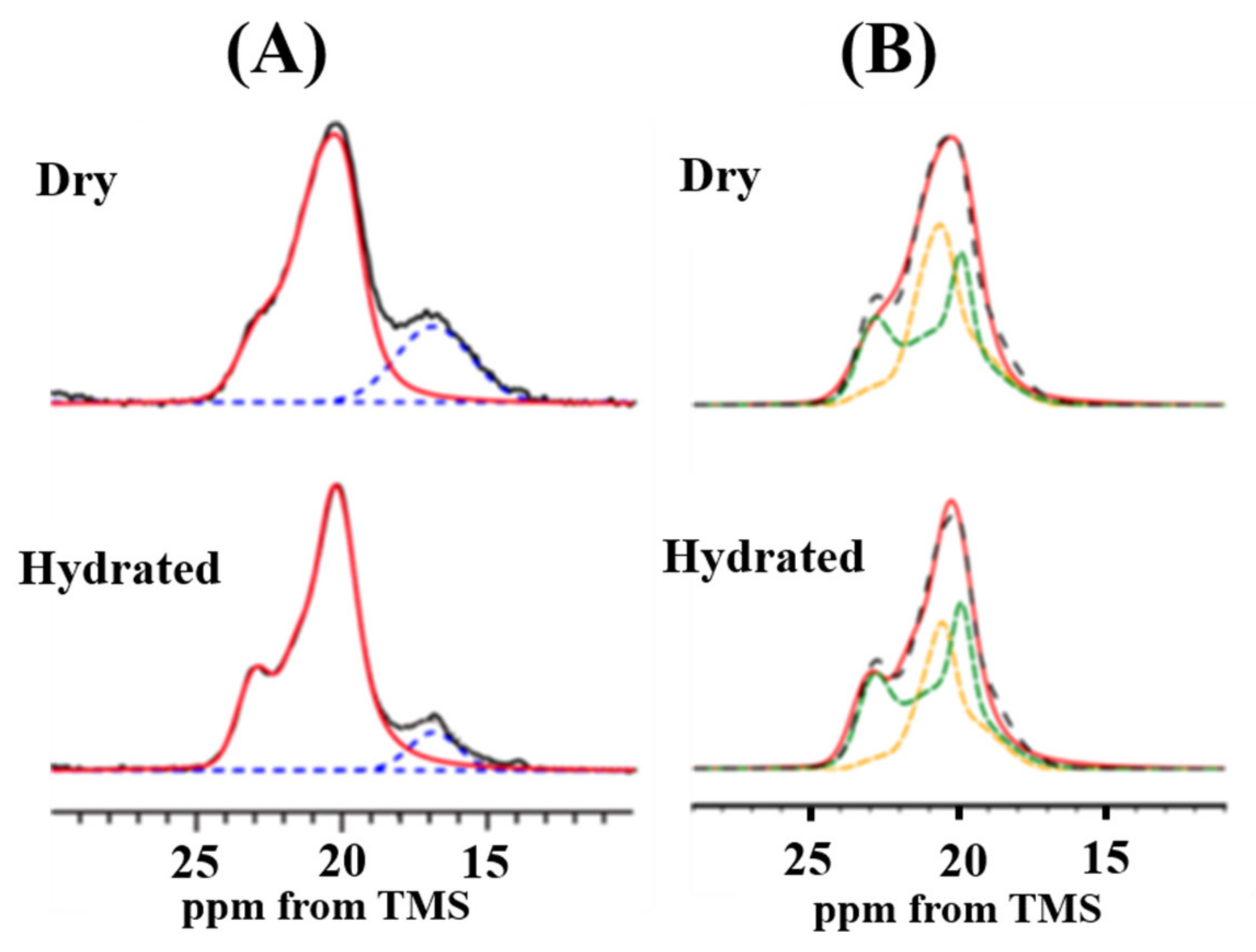
Figure 15.
A series of partially relaxed 13C CP/MAS NMR spectra of Ala Cβ peaks of [3-13C]Ala-N. clavata dragline silk fiber in the dry state (A) and in the hydrated state (B) [61].
Figure 15.
A series of partially relaxed 13C CP/MAS NMR spectra of Ala Cβ peaks of [3-13C]Ala-N. clavata dragline silk fiber in the dry state (A) and in the hydrated state (B) [61].
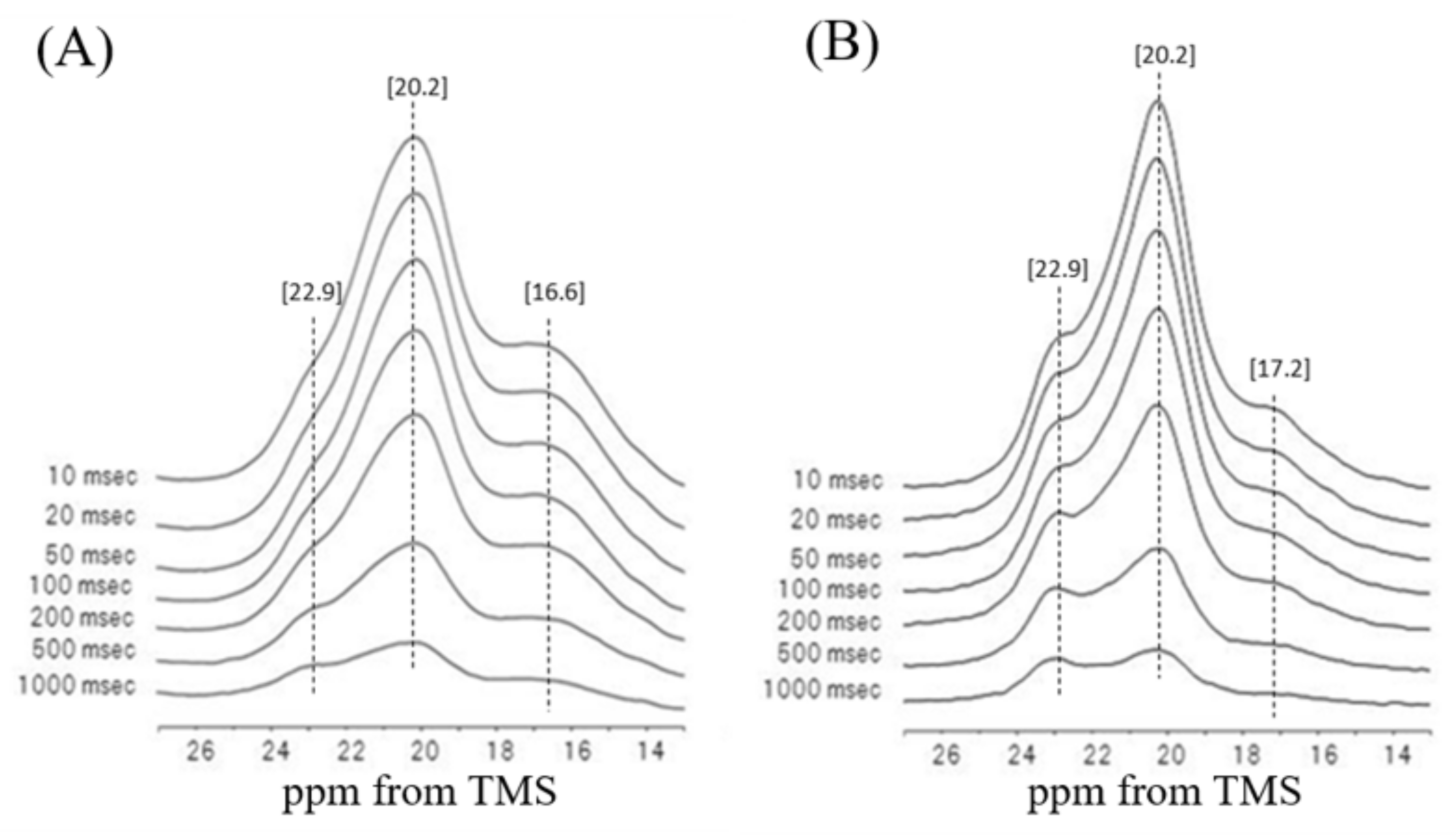
Figure 16.
The packing structure and the dynamical behavior of these Ala Cβ groups in the poly-A region of S. c. ricini silk fibers studied by the MD simulation [110]. Two of the Ala Cβ carbons out of eight existing in the unit cell of the staggered packing structure exhibited the fastest hopping motion in spite of the shortest Cβ−Cβ distance, indicating a geared hopping motion [110].
Figure 16.
The packing structure and the dynamical behavior of these Ala Cβ groups in the poly-A region of S. c. ricini silk fibers studied by the MD simulation [110]. Two of the Ala Cβ carbons out of eight existing in the unit cell of the staggered packing structure exhibited the fastest hopping motion in spite of the shortest Cβ−Cβ distance, indicating a geared hopping motion [110].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
47-mer peptides as the sequential models of the Gly-rich region in Figure 3 as underlined sequence, a flanked by (Ala)6 at both ends [46,60,62]. (I) The peptides, 2–7 were used for determinations of both the inter-nuclear distances of 15N and 13C labeled sites, and the fractions of the local conformations of 13C labeled Gly residues. (II) The peptides, 8–10 were used for determination of the fractions of the local conformations of the 13C labeled Gly-Ala-Gly motifs.
Table 1.
47-mer peptides as the sequential models of the Gly-rich region in Figure 3 as underlined sequence, a flanked by (Ala)6 at both ends [46,60,62]. (I) The peptides, 2–7 were used for determinations of both the inter-nuclear distances of 15N and 13C labeled sites, and the fractions of the local conformations of 13C labeled Gly residues. (II) The peptides, 8–10 were used for determination of the fractions of the local conformations of the 13C labeled Gly-Ala-Gly motifs.
| I: | REDOR and 13C CP/MAS NMR Experiments | Distance * |
|---|---|---|
| 1 | (E)4(A)6GGAGQGGYGGLGSQGAGRGGLGGQGAG(A)6(E)4 | |
| 2 | (E)4(A)6GGA[1-13C]G14Q[15N]G15GYGGLGSQGAGRGGLGGQGAG(A)6(E)4 | 3.71 Å |
| 3 | (E)4(A)6GGAGQG[1-13C]G17Y[15N]G19GLGSQGAGRGGLGGQGAG(A)6(E)4 | 3.97 Å |
| 4 | (E)4(A)6GGAGQGGYG[1-13C]G20L[15N]G22SQGAGRGGLGGQGAG(A)6(E)4 | 3.41 Å |
| 5 | (E)4(A)6GGAGQGGYGGLGSQGA[1-13C]G27R[15N]G29GLGGQGAG(A)6(E)4 | 3.64 Å |
| 6 | (E)4(A)6GGAGQGGYGGLGSQGAGRG[1-13C]G30L[15N]G32GQGAG(A)6(E)4 | 3.43 Å |
| 7 | (E)4(A)6GGAGQGGYGGLGSQGAGRGGLG[1-13C]G33Q[15N]35GAG(A)6(E)4 | 3.81 Å |
| II: | 13C CP/MAS NMR Experiment | |
| 8 | (E)4(A)6G [2-13C]G12 [3-13C]A13 [1-13C]G14QGGYGGLGSQGAGRGGLGGQGAG(A)6(E)4 | |
| 9 | (E)4(A)6GGAGQGGYGGLGSQ [2-13C]G25 [3-13C]A26 [1-13C]G27RGGLGGQGAG(A)6(E)4 | |
| 10 | (E)4(A)6GGAGQGGYGGLGSQGAGRGGLGGQ [2-13C]G35 [3-13C]A36 [1-13C]G37(A)6(E)4 | |
Distance * is the inter-nuclear distance between 15N and 13C labeled sites [46].
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Asakura, T. Structure and Dynamics of Spider Silk Studied with Solid-State Nuclear Magnetic Resonance and Molecular Dynamics Simulation. Molecules 2020, 25, 2634. https://doi.org/10.3390/molecules25112634
AMA Style
Asakura T. Structure and Dynamics of Spider Silk Studied with Solid-State Nuclear Magnetic Resonance and Molecular Dynamics Simulation. Molecules. 2020; 25(11):2634. https://doi.org/10.3390/molecules25112634
Chicago/Turabian StyleAsakura, Tetsuo. 2020. "Structure and Dynamics of Spider Silk Studied with Solid-State Nuclear Magnetic Resonance and Molecular Dynamics Simulation" Molecules 25, no. 11: 2634. https://doi.org/10.3390/molecules25112634