
Transport Oligonucleotides—A Novel System for Intracellular Delivery of Antisense Therapeutics

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Design and Synthesis of Transport Oligonucleotides
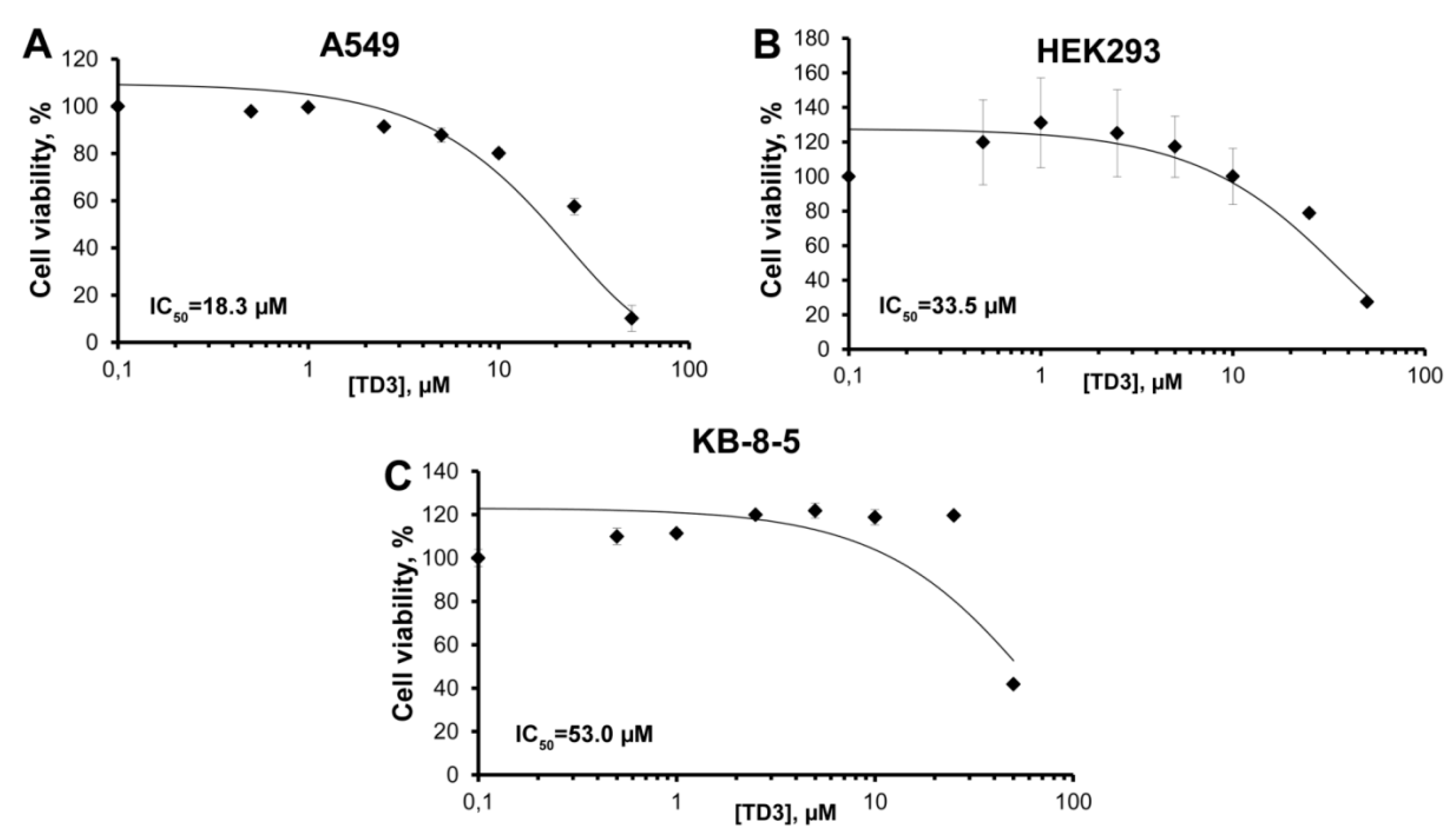
2.2. Cell Viability Assay
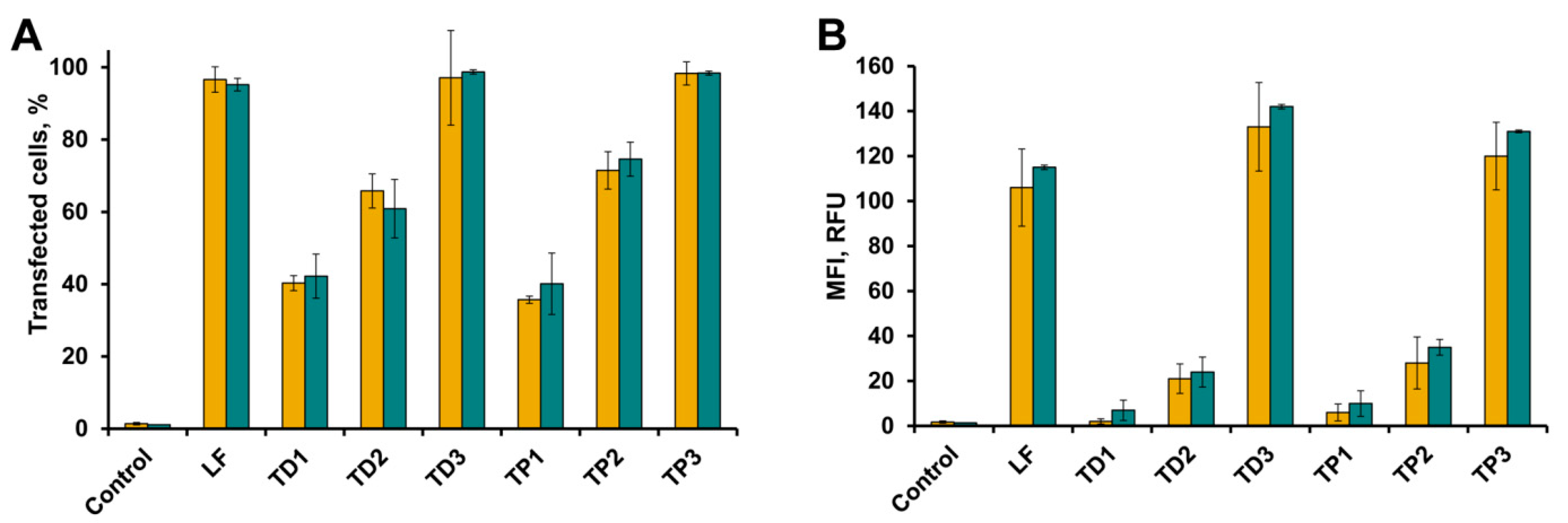
2.3. Intracellular Accumulation of tON in A549 and HEK293 Cells
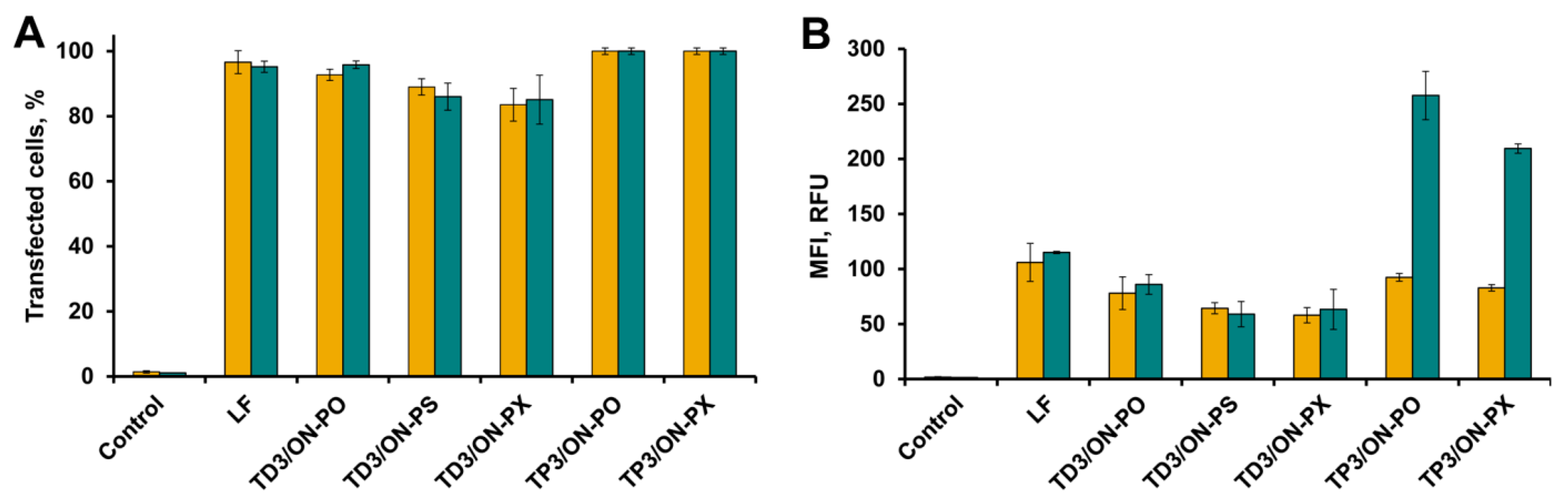
2.4. Accumulation of tON/dON Duplexes in A549 and HEK293 Cells
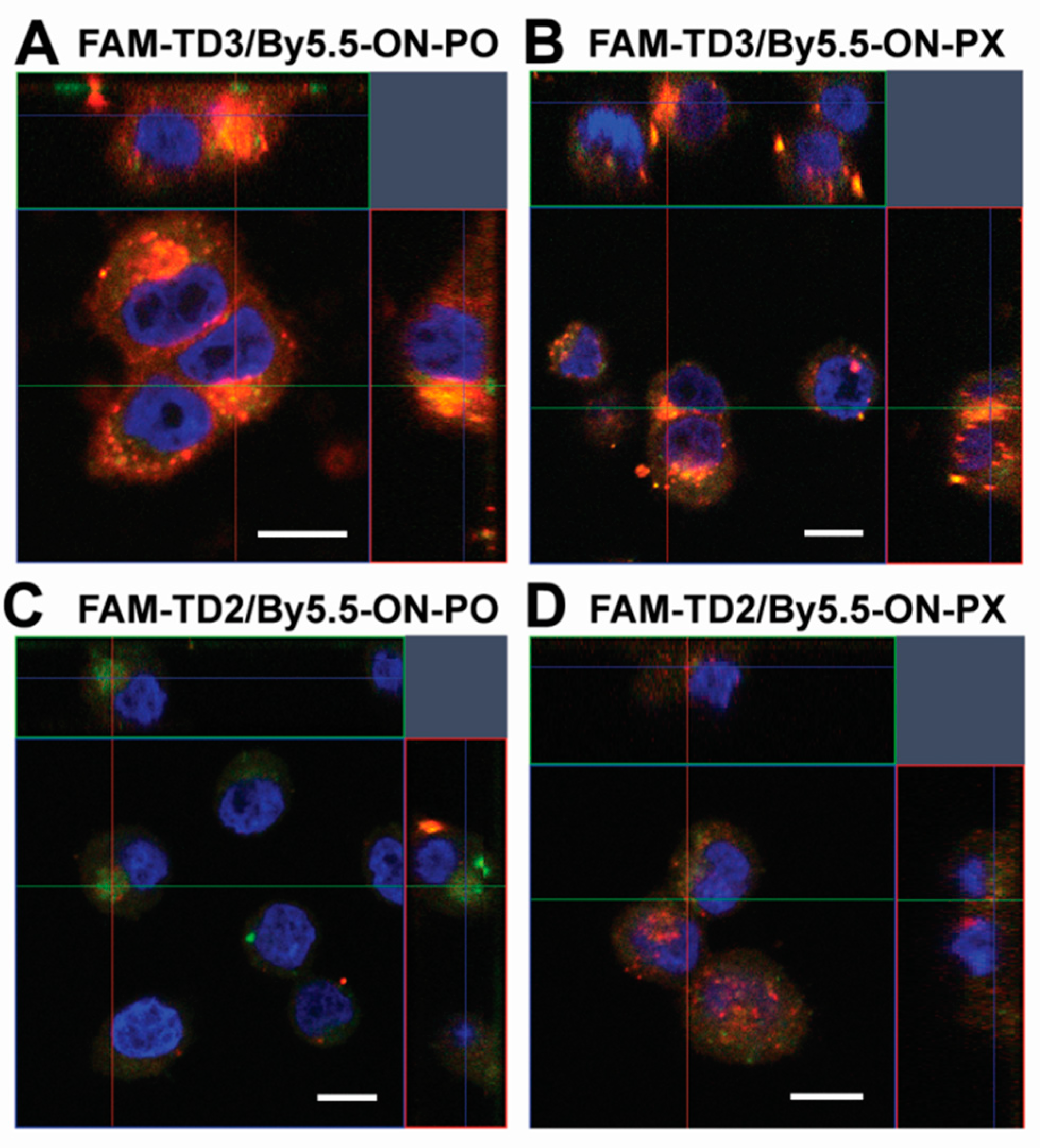
2.5. Intracellular Localization of tON/dON Duplexes
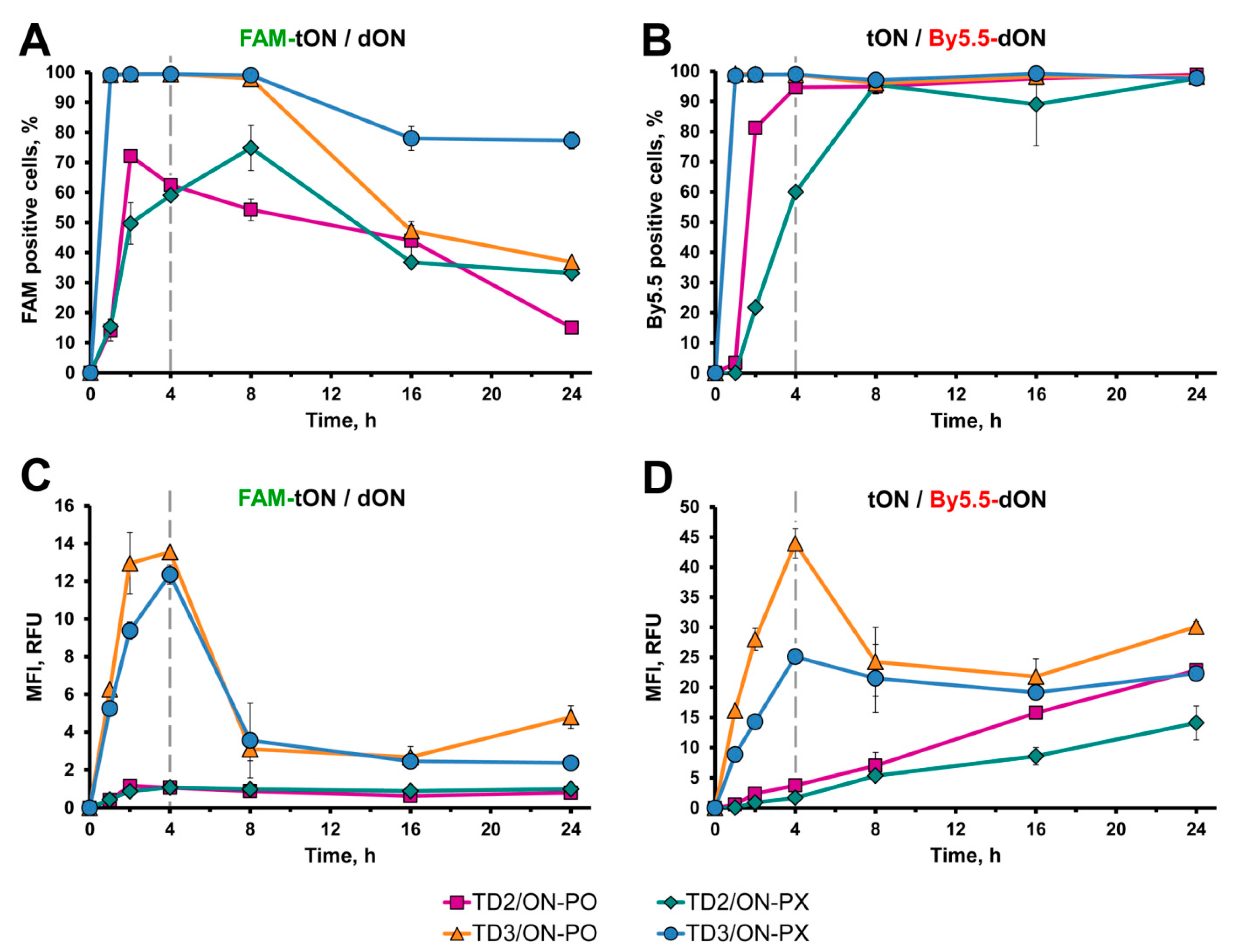
2.6. Kinetics of tON/dON Duplex Accumulation in KB-8-5 Cells
2.6.1. Flow Cytometry
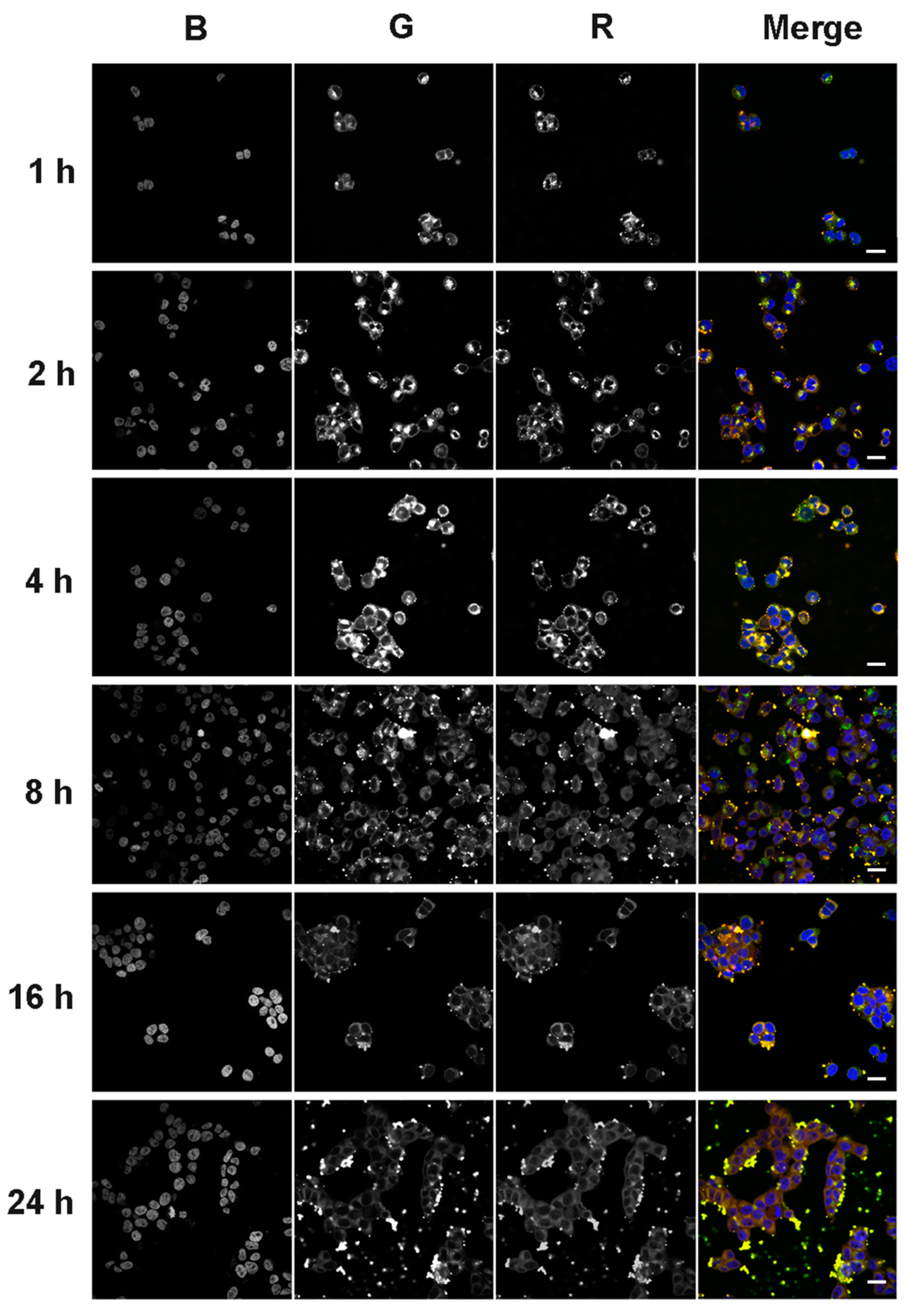
2.6.2. Confocal Fluorescent Microscopy
2.7. On the Mechanism of Release of TD3/dON Duplexes from Cells
2.8. Biological Activity of TD3/asON Duplexes
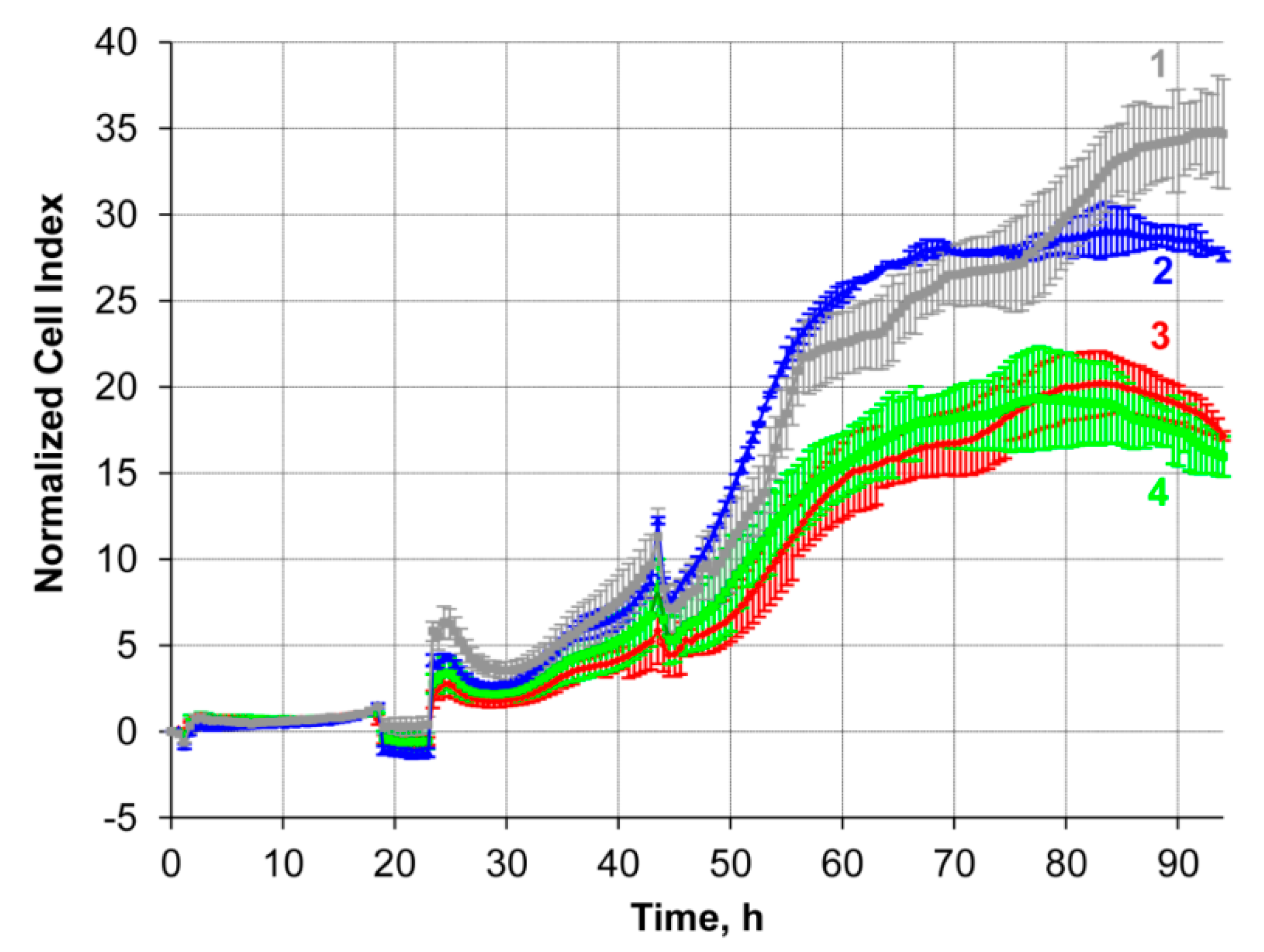
2.8.1. Silencing of P-Glycoprotein Expression in KB-8-5 Cells by Duplexes TD3/ON-MDR1
2.8.2. Reversion of Multiple Drug Resistance Phenotype of KB-8-5 Cells by Antisense Oligonucleotides Delivered with TD3
3. Discussion
4. Materials and Methods
4.1. Oligonucleotide Preparation
4.2. Oligonucleotide Purification and Identification
4.3. Duplex Thermal Denaturation Experiments
4.4. Cell Lines
4.5. Duplex Formation
4.6. Cell Viability Assay
4.7. Effects of tON/asON Duplexes on Apoptosis Induction, Cell Cycle and Mitochondrial Potential
4.8. Analysis of Cellular Accumulation of tON or Duplexes of tON/dON
4.9. Flow Cytometry
4.10. Confocal Microscopy
4.11. Transmission Electron Microscopy of Extracellular Vesicles Containing Duplexes
4.12. Western Blotting
4.13. Evaluation of Sensitivity of KB-8-5 Cells to Vinblastine
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Setten, R.L.; Rossi, J.J.; Han, S. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Corey, D.R. Chemistry, mechanism and clinical status of antisense oligonucleotides and duplex RNAs. Nucleic Acids Res. 2018, 46, 1584–1600. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.A.; Castanotto, D. FDA-Approved Oligonucleotide Therapies in 2017. Mol. Ther. 2017, 25, 1069–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Hoy, S.M. Nusinersen: First Global Approval. Drugs 2017, 77, 473–479. [Google Scholar] [CrossRef]
- Paik, J.; Duggan, S. Volanesorsen: First Global Approval. Drugs 2019, 79, 1349–1354. [Google Scholar] [CrossRef]
- Chernikov, I.V.; Vlassov, V.V.; Chernolovskaya, E.L. Current development of siRNA bioconjugates: From research to the clinic. Front. Pharmacol. 2019, 10, 444. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Hashiba, K.; Sasaki, K.; Maeki, M.; Tokeshi, M.; Harashima, H. Understanding structure-activity relationships of pH-sensitive cationic lipids facilitates the rational identification of promising lipid nanoparticles for delivering siRNAs in vivo. J. Control. Release 2019, 295, 140–152. [Google Scholar] [CrossRef]
- Jayaraman, M.; Ansell, S.M.; Mui, B.L.; Tam, Y.K.; Chen, J.; Du, X.; Butler, D.; Eltepu, L.; Matsuda, S.; Narayanannair, J.K.; et al. Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew. Chemie-Int. Ed. 2012, 51, 8529–8533. [Google Scholar] [CrossRef]
- Khan, O.F.; Kowalski, P.S.; Doloff, J.C.; Tsosie, J.K.; Bakthavatchalu, V.; Winn, C.B.; Haupt, J.; Jamiel, M.; Langer, R.; Anderson, D.G. Endothelial siRNA delivery in nonhuman primates using ionizable low–molecular weight polymeric nanoparticles. Sci. Adv. 2018, 4, eaar8409. [Google Scholar] [CrossRef] [Green Version]
- Dahlman, J.E.; Barnes, C.; Khan, O.F.; Thiriot, A.; Jhunjunwala, S.; Shaw, T.E.; Xing, Y.; Sager, H.B.; Sahay, G.; Speciner, L.; et al. In vivo endothelial siRNA delivery using polymeric nanoparticles with low molecular weight. Nat. Nanotechnol. 2014, 9, 648–655. [Google Scholar] [PubMed]
- Kim, B.; Park, J.H.; Sailor, M.J. Rekindling RNAi Therapy: Materials Design Requirements for In Vivo siRNA Delivery. Adv. Mater. 2019, 31, e1903637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossen, M.N.; Rao, G.; Dey, A.; Robertson, J.D.; Bhattacharya, R.; Mukherjee, P. Gold Nanoparticle Transforms Activated Cancer-Associated Fibroblasts to Quiescence. ACS Appl. Mater. Interfaces 2019, 11, 26060–26068. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Baker, B.F.; Xia, S.; Yu, R.Z.; Viney, N.J.; Wang, Y.; Tsimikas, S.; Geary, R.S. Integrated Assessment of the Clinical Performance of GalNAc 3-Conjugated 2′-O-Methoxyethyl Chimeric Antisense Oligonucleotides: I. Human Volunteer Experience. Nucleic Acid Ther. 2019, 29, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.Z.; Graham, M.J.; Post, N.; Riney, S.; Zanardi, T.; Hall, S.; Burkey, J.; Shemesh, C.S.; Prakash, T.P.; Seth, P.P.; et al. Disposition and Pharmacology of a GalNAc3-conjugated ASO Targeting Human Lipoprotein (a) in Mice. Mol. Ther. Nucleic Acids 2016, 5, e317. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J. Givosiran: First Approval. Drugs 2020, 80, 335–339. [Google Scholar] [CrossRef]
- Huang, Y. Preclinical and Clinical Advances of GalNAc-Decorated Nucleic Acid Therapeutics. Mol. Ther. Nucleic Acids 2017, 6, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Biscans, A.; Coles, A.; Haraszti, R.; Echeverria, D.; Hassler, M.; Osborn, M.; Khvorova, A. Diverse lipid conjugates for functional extra-hepatic siRNA delivery in vivo. Nucleic Acids Res. 2019, 47, 1082–1096. [Google Scholar] [CrossRef] [Green Version]
- Prakash, T.P.; Mullick, A.E.; Lee, R.G.; Yu, J.; Yeh, S.T.; Low, A.; Chappell, A.E.; Østergaard, M.E.; Murray, S.; Gaus, H.J.; et al. Fatty acid conjugation enhances potency of antisense oligonucleotides in muscle. Nucleic Acids Res. 2019, 47, 6029–6044. [Google Scholar] [CrossRef]
- Osborn, M.F.; Coles, A.H.; Biscans, A.; Haraszti, R.A.; Roux, L.; Davis, S.; Ly, S.; Echeverria, D.; Hassler, M.R.; Godinho, B.M.D.C.; et al. Hydrophobicity drives the systemic distribution of lipid-conjugated siRNAs via lipid transport pathways. Nucleic Acids Res. 2019, 47, 1070–1081. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, F. Phosphorothioates, essential components of therapeutic oligonucleotides. Nucleic Acid Ther. 2014, 24, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.H.S.; Wan, J.; Spesock, A.; Sergueeva, Z.; Shaw, B.R.; Alexander, K.A. High potency silencing by single-stranded boranophosphate siRNA. Nucleic Acids Res. 2006, 34, 2773–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gryaznov, S.M. Oligonucleotide N3′→P5′ phosphoramidates and thio-phoshoramidates as potential therapeutic agents. Chem. Biodivers. 2010, 7, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Vlaho, D.; Fakhoury, J.F.; Damha, M.J. Structural studies and gene silencing activity of siRNAs containing cationic phosphoramidate linkages. Nucleic Acid Ther. 2018, 28, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.; Lunstad, B.; Yamada, C.M.; Stell, B.G.; Caruthers, M.H.; Dellinger, D.J. Biochemical properties of phosphonoacetate and thiophosphonoacetate oligodeoxyribonucleotides. Nucleic Acids Res. 2003, 31, 4109–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meade, B.R.; Gogoi, K.; Hamil, A.S.; Palm-Apergi, C.; Van Den Berg, A.; Hagopian, J.C.; Springer, A.D.; Eguchi, A.; Kacsinta, A.D.; Dowdy, C.F.; et al. Efficient delivery of RNAi prodrugs containing reversible charge-neutralizing phosphotriester backbone modifications. Nat. Biotechnol. 2014, 32, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Kupryushkin, M.S.; Pyshnyi, D.V.; Stetsenko, D.A. Phosphoryl guanidines: A new type of nucleic acid analogues. Acta Naturae 2014, 6, 116–118. [Google Scholar] [CrossRef]
- Miroshnichenko, S.K.; Patutina, O.A.; Burakova, E.A.; Chelobanov, B.P.; Fokina, A.A.; Vlassov, V.V.; Altman, S.; Zenkova, M.A.; Stetsenko, D.A. Mesyl phosphoramidate antisense oligonucleotides as an alternative to phosphorothioates with improved biochemical and biological properties. Proc. Natl. Acad. Sci. USA 2019, 116, 1229–1234. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; Rigo, F.; Bennett, C.F.; Krainer, A.R.; Hua, Y. Comparison of the efficacy of MOE and PMO modifications of systemic antisense oligonucleotides in a severe SMA mouse model. Nucleic Acids Res. 2020, 48, 2853–2865. [Google Scholar] [CrossRef]
- Muangkaew, P.; Vilaivan, T. Modulation of DNA and RNA by PNA. Bioorg. Med. Chem. Lett. 2020, 30, 127064. [Google Scholar] [CrossRef]
- Shen, W.; De Hoyos, C.L.; Sun, H.; Vickers, T.A.; Liang, X.H.; Crooke, S.T. Acute hepatotoxicity of 2 fluoro-modified 5–10–5 gapmer phosphorothioate oligonucleotides in mice correlates with intracellular protein binding and the loss of DBHS proteins. Nucleic Acids Res. 2018, 46, 2204–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; De Hoyos, C.L.; Migawa, M.T.; Vickers, T.A.; Sun, H.; Low, A.; Bell, T.A.; Rahdar, M.; Mukhopadhyay, S.; Hart, C.E.; et al. Chemical modification of PS-ASO therapeutics reduces cellular protein-binding and improves the therapeutic index. Nat. Biotechnol. 2019, 37, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, T.; Morales-Medina, J.; Palmieri, B. Phosphorothioate Oligonucleotides: Effectiveness and Toxicity. Curr. Drug Targets 2014, 15, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, M.E.; De Hoyos, C.L.; Wan, W.B.; Shen, W.; Low, A.; Berdeja, A.; Vasquez, G.; Murray, S.; Migawa, M.T.; Liang, X.H.; et al. Understanding the effect of controlling phosphorothioate chirality in the DNA gap on the potency and safety of gapmer antisense oligonucleotides. Nucleic Acids Res. 2020, 48, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Migawa, M.T.; Shen, W.; Brad Wan, W.; Vasquez, G.; Oestergaard, M.E.; Low, A.; De Hoyos, C.L.; Gupta, R.; Murray, S.; Tanowitz, M.; et al. Site-specific replacement of phosphorothioate with alkyl phosphonate linkages enhances the therapeutic profile of gapmer ASOs by modulating interactions with cellular proteins. Nucleic Acids Res. 2019, 47, 5465–5479. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Yasuhara, H.; Wada, F.; Sawamura, M.; Waki, R.; Yamamoto, T.; Harada-Shiba, M.; Obika, S. Evaluation of the effects of chemically different linkers on hepatic accumulations, cell tropism and gene silencing ability of cholesterol-conjugated antisense oligonucleotides. J. Control. Release 2016, 226, 57–65. [Google Scholar] [CrossRef]
- Hagiwara, K.; Kurihara, K.; Honma, M.; Yamamoto, J.; Shinohara, F. PEG-modification on the endo-position of an antisense oligonucleotide increases tumor accumulation via the EPR effect. J. Biomater. Sci. Polym. Ed. 2018, 29, 448–459. [Google Scholar] [CrossRef]
- Shokrzadeh, N.; Winkler, A.M.; Dirin, M.; Winkler, J. Oligonucleotides conjugated with short chemically defined polyethylene glycol chains are efficient antisense agents. Bioorg. Med. Chem. Lett. 2014, 24, 5758–5761. [Google Scholar] [CrossRef] [Green Version]
- Tepel, J.; March, C.; Ketterer, T.; Kapischke, M.; Arlt, A.; Kremer, B.; Kalthoff, H.; Kruse, M.L. A modified random oligonucleotide-based combination therapy for adjuvant treatment of pancreatic ductal adenocarcinoma. Int. J. Oncol. 2006, 28, 1105–1112. [Google Scholar] [CrossRef] [Green Version]
- Nishina, T.; Numata, J.; Nishina, K.; Yoshida-Tanaka, K.; Nitta, K.; Piao, W.; Iwata, R.; Ito, S.; Kuwahara, H.; Wada, T.; et al. Chimeric antisense oligonucleotide conjugated to α-Tocopherol. Mol. Ther. Nucleic Acids 2015, 4, e220. [Google Scholar] [CrossRef]
- Le, T.D.; Nakagawa, O.; Fisher, M.; Juliano, R.L.; Yoo, H. RGD conjugated dendritic polylysine for cellular delivery of antisense oligonucleotide. J. Nanosci. Nanotechnol. 2017, 17, 2353–2357. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.M.; Fadzen, C.M.; Holden, R.L.; Yao, M.; Hanson, G.J.; Pentelute, B.L. Perfluoroaryl Bicyclic Cell-Penetrating Peptides for Delivery of Antisense Oligonucleotides. Angew. Chem. Int. Ed. 2018, 57, 4756–4759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gusachenko, O.N.; Pishnyi, D.V.; Vlassov, V.V.; Zenkova, M.A. Modified concatemeric oligonucleotide complexes: New system for efficient oligonucleotide transfer into mammalian cells. Hum. Gene Ther. 2008, 19, 532–546. [Google Scholar] [CrossRef]
- Simonova, O.N.; Vladimirova, A.V.; Zenkova, M.A.; Vlassov, V.V. Enhanced cellular binding of concatemeric oligonucleotide complexes. Biochim. Biophys. Acta Biomembr. 2006, 1758, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Misu, S.; Kurihara, R.; Kainuma, R.; Sato, R.; Nishihara, T.; Tanabe, K. Hybridizing Oligonucleotides with Hydrophobic Peptide Nucleic Acids Assists Their Cellular Uptake through Aggregate Formation. ChemBioChem 2020, 21, 1140–1143. [Google Scholar] [CrossRef] [PubMed]
- McClorey, G.; Banerjee, S. Cell-penetrating peptides to enhance delivery of oligonucleotide-based therapeutics. Biomedicines 2018, 6, 51. [Google Scholar] [CrossRef] [Green Version]
- Raouane, M.; Desmaële, D.; Urbinati, G.; Massaad-Massade, L.; Couvreur, P. Lipid conjugated oligonucleotides: A useful strategy for delivery. Bioconjug. Chem. 2012, 23, 1091–1104. [Google Scholar] [CrossRef]
- Kupryushkin, M.S.; Nekrasov, M.D.; Stetsenko, D.A.; Pyshnyi, D.V. Efficient functionalization of oligonucleotides by new achiral nonnucleosidic monomers. Org. Lett. 2014, 16, 2842–2845. [Google Scholar] [CrossRef]
- Nielsen, J.; Caruthers, M.H. Directed arbuzov-type reactions of 2-cyano-1,1-dimethylethyl deoxynucleoside phosphites. J. Am. Chem. Soc. 1988, 110, 6275–6276. [Google Scholar] [CrossRef]
- Thomas, P.; Smart, T.G. HEK293 cell line: A vehicle for the expression of recombinant proteins. J. Pharmacol. Toxicol. Methods 2005, 51, 187–200. [Google Scholar] [CrossRef]
- Foreman, H.C.C.; Lalwani, G.; Kalra, J.; Krug, L.T.; Sitharaman, B. Gene delivery to mammalian cells using a graphene nanoribbon platform. J. Mater. Chem. B 2017, 5, 2347–2354. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, D.A.; Maslov, M.A.; Serikov, R.N.; Morozova, N.G.; Serebrenikova, G.A.; Sheglov, D.V.; Latyshev, A.V.; Vlassov, V.V.; Zenkova, M.A. Novel cholesterol-based cationic lipids for gene delivery. J. Med. Chem. 2009, 52, 6558–6568. [Google Scholar]
- Ju, J.; Huan, M.L.; Wan, N.; Qiu, H.; Zhou, S.Y.; Zhang, B. Le Novel cholesterol-based cationic lipids as transfecting agents of DNA for efficient gene delivery. Int. J. Mol. Sci. 2015, 16, 5666–5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svoboda, O.; Fohlerova, Z.; Baiazitova, L.; Mlynek, P.; Samouylov, K.; Provaznik, I.; Hubalek, J. Transfection by polyethyleneimine-coated magnetic nanoparticles: Fine-tuning the condition for electrophysiological experiments. J. Biomed. Nanotechnol. 2018, 14, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, S.; Haghighipour, N.; Soleymani, S.; Nadji, S.A.; Bolhassani, A. Delivery of molecular cargoes in normal and cancer cell lines using non-viral delivery systems. Biotechnol. Lett. 2018, 40, 923–931. [Google Scholar] [CrossRef]
- Kostenko, E.V.; Laktionov, P.P.; Vlassov, V.V.; Zenkova, M.A. Downregulation of PGY1/MDR1 mRNA level in human KB cells by antisense oligonucleotide conjugates. RNA accessibility in vitro and intracellular antisense activity. Biochim. Biophys. Acta 2002, 1576, 143–147. [Google Scholar] [CrossRef]
- Logashenko, E.B.; Vladimirova, A.V.; Repkova, M.N.; Venyaminova, A.G.; Chernolovskaya, E.L.; Vlassov, V.V. Silencing of MDR 1 gene in cancer cells by siRNA. Nucleosides Nucleotides Nucleic Acids 2004, 23, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.S.; Chernikov, I.V.; Meschaninova, M.I.; Dovydenko, I.S.; Venyaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Carrier-free cellular uptake and the gene-silencing activity of the lipophilic siRNAs is strongly affected by the length of the linker between siRNA and lipophilic group. Nucleic Acids Res. 2012, 40, 2330–2344. [Google Scholar] [CrossRef] [Green Version]
- Chernikov, I.V.; Gladkikh, D.V.; Meschaninova, M.I.; Ven’yaminova, A.G.; Zenkova, M.A.; Vlassov, V.V.; Chernolovskaya, E.L. Cholesterol-Containing Nuclease-Resistant siRNA Accumulates in Tumors in a Carrier-free Mode and Silences MDR1 Gene. Mol. Ther. Nucleic Acids 2017, 6, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Chernikov, I.V.; Gladkikh, D.V.; Karelina, U.A.; Meschaninova, M.I.; Ven’yaminova, A.G.; Vlassov, V.V.; Chernolovskaya, E.L. Trimeric small interfering RNAs and their cholesterol-containing conjugates exhibit improved accumulation in tumors, but dramatically reduced silencing activity. Molecules 2020, 25, 1877. [Google Scholar] [CrossRef]
- Cardarelli, F.; Digiacomo, L.; Marchini, C.; Amici, A.; Salomone, F.; Fiume, G.; Rossetta, A.; Gratton, E.; Pozzi, D.; Caracciolo, G. The intracellular trafficking mechanism of Lipofectamine-based transfection reagents and its implication for gene delivery. Sci. Rep. 2016, 6, 25879. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doughty, M.J. PH dependent spectral properties of sodium fluorescein ophthalmic solutions revisited. Ophthalmic Physiol. Opt. 2010, 30, 167–174. [Google Scholar] [CrossRef]
- Crooke, S.T.; Wang, S.; Vickers, T.A.; Shen, W.; Liang, X.H. Cellular uptake and trafficking of antisense oligonucleotides. Nat. Biotechnol. 2017, 35, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun, H.; Tanowitz, M.; Liang, X.H.; Crooke, S.T. Annexin A2 facilitates endocytic trafficking of antisense oligonucleotides. Nucleic Acids Res. 2016, 44, 7314–7330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilleron, J.; Paramasivam, P.; Zeigerer, A.; Querbes, W.; Marsico, G.; Andree, C.; Seifert, S.; Amaya, P.; Stöter, M.; Koteliansky, V.; et al. Identification of siRNA delivery enhancers by a chemical library screen. Nucleic Acids Res. 2015, 43, 7984–8001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, S.; Navaroli, D.M.; Didiot, M.C.; Cardia, J.; Pandarinathan, L.; Alterman, J.F.; Fogarty, K.; Standley, C.; Lifshitz, L.M.; Bellve, K.D.; et al. Visualization of self-delivering hydrophobically modified siRNA cellular internalization. Nucleic Acids Res. 2017, 45, 15–25. [Google Scholar] [CrossRef]
- Shukla, R.S.; Jain, A.; Zhao, Z.; Cheng, K. Intracellular trafficking and exocytosis of a multi-component siRNA nanocomplex. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1323–1334. [Google Scholar] [CrossRef] [Green Version]
- Zenkov, A.N.; Scvortsova, N.V.; Chernolovskaya, E.L.; Pospelova, T.I.; Vlassov, V.V. Expression of the MDR1 and MRP genes in patients with lymphoma with primary bone marrow involvement. Nucleosides Nucleotides Nucleic Acids 2004, 23, 843–847. [Google Scholar] [CrossRef]
- Yang, H.; Ding, R.; Tong, Z.; Huang, J.; Shen, L.; Sun, Y.; Liao, J.; Yang, Z.; Hoffman, R.M.; Wang, C.; et al. SiRNA targeting of MDR1 reverses multidrug resistance in a nude mouse model of doxorubicin-resistant human hepatocellular carcinoma. Anticancer Res. 2016, 36, 2675–2682. [Google Scholar]
- Essex, S.; Navarro, G.; Sabhachandani, P.; Chordia, A.; Trivedi, M.; Movassaghian, S.; Torchilin, V.P. Phospholipid-modified PEI-based nanocarriers for in vivo siRNA therapeutics against multidrug-resistant tumors. Gene Ther. 2015, 22, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yhee, J.Y.; Song, S.; Lee, S.J.; Park, S.G.; Kim, K.S.; Kim, M.G.; Son, S.; Koo, H.; Kwon, I.C.; Jeong, J.H.; et al. Cancer-targeted MDR-1 siRNA delivery using self-cross-linked glycol chitosan nanoparticles to overcome drug resistance. J. Control. Release 2015, 198, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Du, Z.; Pan, S.; Shi, M.; Li, J.; Yang, C.; Hu, H.; Qiao, M.; Chen, D.; Zhao, X. Overcoming Multidrug Resistance by Codelivery of MDR1-Targeting siRNA and Doxorubicin Using EphA10-Mediated pH-Sensitive Lipoplexes: In Vitro and in Vivo Evaluation. ACS Appl. Mater. Interfaces 2018, 10, 21590–21600. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jia, L.; Wang, Q.; Hu, H.; Zhao, X.; Chen, D.; Qiao, M. PH/Redox Dual-Responsive Polyplex with Effective Endosomal Escape for Codelivery of siRNA and Doxorubicin against Drug-Resistant Cancer Cells. ACS Appl. Mater. Interfaces 2019, 11, 16296–16310. [Google Scholar] [CrossRef] [PubMed]
- Stetsenko, D.A.; Kupryushkin, M.S.; Pyshnyi, D.V. Modified Oligonucleotides and Methods for Their Synthesis. International Patent No. WO2016028187A1, 22 June 2014. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Oligonucleotide | Name | Sequence and Modifications 1 | Mr (Theor)/Mr (Exp) |
|---|---|---|---|
| Transport Oligonucleotides | TD1 FAM-TD1 | 5′-T[R1]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R1]-GGTAGCAAGTCGAGACT-3′ | n.d. 2 6233.47/6232.40 |
| TD2 FAM-TD2 | 5′-T[R1][R1]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R1][R1]-GGTAGCAAGTCGAGACT-3′ | n.d. 6639.93/6639.00 | |
| TD3 FAM-TD3 | 5′-T[R1][R1][R1]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R1][R1][R1]-GGTAGCAAGTCGAGACT-3′ | 6783.10/6782.00 7046.39/7044.50 | |
| TP1 FAM-TP1 | 5′-[R2]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R2]-GGTAGCAAGTCGAGACT-3′ | 5731.05/5723.16 6298.54/6296.50 | |
| TP2 FAM-TP2 | 5′-[R2][R2]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R2][R2]-GGTAGCAAGTCGAGACT-3′ | 6202.59/6201.14 6770.08/6768.12 | |
| TP3 FAM-TP3 | 5′-[R2][R2][R2]-GGTAGCAAGTCGAGACT-3′ 5′-[FAM]p[R2][R2][R2]-GGTAGCAAGTCGAGACT-3′ | 6674.13/6673.00 7241.62/7241.00 | |
| Delivered Oligonucleotides | ON-PO FAM-ON-PO By5.5-ON-PO | 5′-AGTCTCGACTTGCTACC-3′ 5′-[FAM]pAGTCTCGACTTGCTACC-3′ 5′-[By5.5]pAGTCTCGACTTGCTACC-3′ | 5121.41/5119.43 5688.90/5688.12 5906.42/5905.60 |
| ON-PS FAM-ON-PS | 5′-ASGSTSCSTSCSGSASCSTSTSGSCSTSASCSC-3′ 5′-[FAM]pASGSTSCSTSCSGSASCSTSTSGSCSTSASCSC-3′ | 5378.44/5377.12 5945.94/5944.31 | |
| ON-PX FAM-ON-PX By5.5-ON-PX | 5′-AXGXTXCXTXCXGXAXCXTXTXGXCXTXAXCXC-3′ 5′-[FAM]pAXGXTXCXTXCXGXAXCXTXTXGXCXTXAXCXC-3′ 5′-[By5.5]pAXGXTXCXTXCXGXAXCXTXTXGXCXTXAXCXC-3′ | 6643.78/6641.90 7211.28/7208.50 8037.22/8035.00 | |
| Antisense Oligonucleotides | MDR1-ON | 5′-GTCCAGCCCCATGGA-TTTT-AGTCTCGACTTGCTACC-3′ | |
| MDR1gap-ON | 5′-GXUXCXCAGCCCCAUXGXGXA-TTTT-AGTCTCGACTTGCTACC-3′ | 11721.24/11721.60 | |
| Scramble | 5′-GXUXUXCCTCGCGCUXCXCXA-TTTT-AGTCTCGACTTGCTACC-3′ | 11651.13/11647.80 |

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markov, O.V.; Filatov, A.V.; Kupryushkin, M.S.; Chernikov, I.V.; Patutina, O.A.; Strunov, A.A.; Chernolovskaya, E.L.; Vlassov, V.V.; Pyshnyi, D.V.; Zenkova, M.A. Transport Oligonucleotides—A Novel System for Intracellular Delivery of Antisense Therapeutics. Molecules 2020, 25, 3663. https://doi.org/10.3390/molecules25163663
Markov OV, Filatov AV, Kupryushkin MS, Chernikov IV, Patutina OA, Strunov AA, Chernolovskaya EL, Vlassov VV, Pyshnyi DV, Zenkova MA. Transport Oligonucleotides—A Novel System for Intracellular Delivery of Antisense Therapeutics. Molecules. 2020; 25(16):3663. https://doi.org/10.3390/molecules25163663
Chicago/Turabian StyleMarkov, Oleg V., Anton V. Filatov, Maxim S. Kupryushkin, Ivan V. Chernikov, Olga A. Patutina, Anton A. Strunov, Elena L. Chernolovskaya, Valentin V. Vlassov, Dmitrii V. Pyshnyi, and Marina A. Zenkova. 2020. "Transport Oligonucleotides—A Novel System for Intracellular Delivery of Antisense Therapeutics" Molecules 25, no. 16: 3663. https://doi.org/10.3390/molecules25163663