
The Effect of Fermentation with Kefir Grains on the Physicochemical and Antioxidant Properties of Beverages from Blue Lupin (Lupinus angustifolius L.) Seeds

 ,
,  ,
, 
 ,
,
Abstract
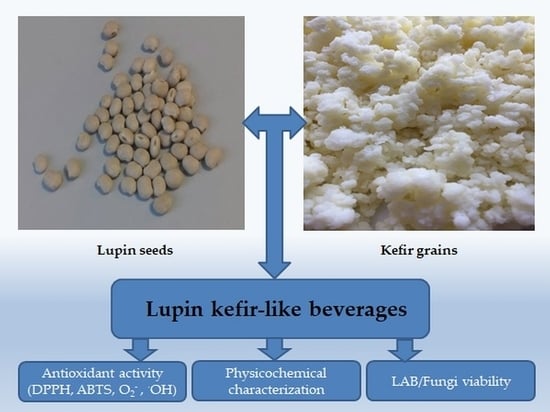
:
1. Introduction
2. Results and Discussion
2.1. The Lactic Acid Bacteria (LAB) and Yeast Survivability during Cold Storage
2.2. The Changes of pH, Reducing Sugars and Free Amino Acids Level
2.3. The Changes of Total Polyphenolics, Flavonoids, and Ascorbic Acid Contents
2.4. The Changes of Reducing Power and Radical Scavenging Activties
2.5. The Changes of Color
2.6. Viscosity and Textural Changes
3. Materials and Methods
3.1. Materials and Reagents
3.2. Samples Preparation and Fermentation
3.3. Microbiological Analyses and pH Determination
3.4. Supernatants Preparation
3.5. Determination of Total Polyphenolic Content (TPC), Total Flavonoid Content (TFC), Reducing Sugars Content (RCS), Total Free Amino Acids Level (TFAAL), and Ascorbic Acid Content (AAC)
3.6. Determination of Reducing Power and Radical Scavenging Activity
3.7. Texture Profile Analysis and Viscosity Measurements
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aspri, M.; Papademas, P.; Tsaltas, D. Review on non-dairy probiotics and their use in non-dairy based products. Fermentation 2020, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Väkeväinen, K.; Ludena-Urquizo, F.; Korkala, E.; Lapveteläinen, A.; Peräniemi, S.; von Wright, A.; Plumed-Ferrer, C. Potential of quinoa in the development of fermented spoonable vegan products. LWT Food Sci. Technol. 2020, 120, 108912. [Google Scholar] [CrossRef]
- Aiello, F.; Restuccia, D.; Spizzirri, U.G.; Carullo, G.; Leporini, M.; Loizzo, M.R. Improving Kefir Bioactive Properties by Functional Enrichment with Plant and Agro-Food Waste Extracts. Fermentation 2020, 6, 83. [Google Scholar] [CrossRef]
- Duranti, M.; Consonni, A.; Magni, C.; Sessa, F.; Scarafoni, A. The major proteins of lupin seed: Characterisation and molecular properties for use as functional and nutraceutical ingredients. Trends Food Sci. Technol. 2008, 19, 624–633. [Google Scholar] [CrossRef]
- Demir, H.; Simsek, M.; Yıldırım, G. Effect of oat milk pasteurization type on the characteristics of yogurt. LWT Food Sci. Technol. 2021, 135, 110271. [Google Scholar] [CrossRef]
- Farag, M.; Jomaa, S.; El-Wahed, A.; El-Seedi, H. The Many Faces of Kefir Fermented Dairy Products. Nutrients 2020, 12, 346. [Google Scholar] [CrossRef] [Green Version]
- Nejati, F.; Junne, S.; Neubauer, P. A big world in small grain: A review of natural milk Kefir starters. Microorganisms 2020, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-J.; Huang, W.-C.; Lin, J.-S.; Chen, Y.-M.; Ho, S.-T.; Huang, C.-C.; Tung, Y.-T. Kefir Supplementation Modifies Gut Microbiota Composition, Reduces Physical Fatigue, and Improves Exercise Performance in Mice. Nutrients 2018, 10, 862. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.R.A.; Silva, M.M.N.; Ribeiro, B.D. Health issues and technological aspects of plant-based alternative milk. Food Res. Int. 2020, 131, 108972. [Google Scholar] [CrossRef]
- Łopusiewicz, Ł.; Drozłowska, E.; Siedlecka, P.; Mężyńska, M.; Bartkowiak, A.; Sienkiewicz, M.; Zielińska-Bliźniewska, H.; Kwiatkowski, P. Development, Characterization, and Bioactivity of Non-Dairy Kefir-Like Fermented Beverage Based on Flaxseed Oil Cake. Foods 2019, 8, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łopusiewicz, Ł.; Drozłowska, E.; Siedlecka, P.; Mężyńska, M.; Bartkowiak, A. Preparation and characterization of novel flaxseed oil cake yogurt-like plant milk fortified with inulin. J. Food Nutr. Res. 2020, 59, 61–70. [Google Scholar]
- Nazhand, A.; Souto, E.B.; Lucarini, M.; Souto, S.B.; Durazzo, A.; Santini, A. Ready to Use Therapeutical Beverages: Focus on Functional Beverages Containing Probiotics, Prebiotics and Synbiotics. Beverages 2020, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Łopusiewicz, Ł.; Drozłowska, E.; Tarnowiecka-Kuca, A.; Bartkowiak, A.; Mazurkiewicz-Zapałowicz, K.; Salachna, P. Biotransformation of flaxseed oil cake into bioactive camembert-analogue using lactic acid bacteria, Penicillium camemberti and Geotrichum candidum. Microorganisms 2020, 8, 1266. [Google Scholar] [CrossRef]
- Mäkinen, O.E.; Wanhalinna, V.; Zannini, E.; Arendt, E.K. Foods for Special Dietary Needs: Non-dairy Plant-based Milk Substitutes and Fermented Dairy-type Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 339–349. [Google Scholar] [CrossRef]
- Jiménez-Martínez, C.; Hernández-Sánchez, H.; Dávila-Ortiz, G. Production of a yogurt-like product from Lupinus campestris seeds. J. Sci. Food Agric. 2003, 83, 515–522. [Google Scholar] [CrossRef]
- Karina, T.M.-G.; Uéllina, S.S.; Marcia, R.S.; Ferlando, L.S.; Itaciara, L.N. Production of rice cereal-based Kefir beverage. Afr. J. Biotechnol. 2018, 17, 322–327. [Google Scholar] [CrossRef]
- Atalar, I. Functional kefir production from high pressure homogenized hazelnut milk. LWT Food Sci. Technol. 2019, 107, 256–263. [Google Scholar] [CrossRef]
- Bensmira, M.; Jiang, B. Total phenolic compounds and antioxidant activity of a novel peanut based kefir. Food Sci. Biotechnol. 2015, 24, 1055–1060. [Google Scholar] [CrossRef]
- Corona, O.; Randazzo, W.; Miceli, A.; Guarcello, R.; Francesca, N.; Erten, H.; Moschetti, G.; Settanni, L. Characterization of kefir-like beverages produced from vegetable juices. LWT Food Sci. Technol. 2016, 66, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, W.; Corona, O.; Guarcello, R.; Francesca, N.; Germanà, M.A.; Erten, H.; Moschetti, G.; Settanni, L. Development of new non-dairy beverages from Mediterranean fruit juices fermented with water kefir microorganisms. Food Microbiol. 2016, 54, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Plessas, S.; Nouska, C.; Mantzourani, I.; Kourkoutas, Y.; Alexopoulos, A.; Bezirtzoglou, E. Microbiological Exploration of Different Types of Kefir Grains. Fermentation 2016, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Sardjono; Zhu, Y.; Knol, W. Comparison of Fermentation Profiles between Lupine and Soybean by Aspergillus oryzae and Aspergillus sojae in Solid-State Culture Systems. J. Agric. Food Chem. 1998, 46, 3376–3380. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Bez, J.; Petersen, I.L.; Joehnke, M.S.; Detzel, A.; Busch, M.; Krueger, M.; Ispiryan, L.; O’Mahony, J.A.; Arendt, E.K.; et al. Techno-functional, nutritional and environmental performance of protein isolates from blue lupin and white lupin. Foods 2020, 9, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkiene, E.; Sakiene, V.; Bartkevics, V.; Juodeikiene, G.; Lele, V.; Wiacek, C.; Braun, P.G. Modulation of the nutritional value of lupine wholemeal and protein isolates using submerged and solid-state fermentation with Pediococcus pentosaceus strains. Int. J. Food Sci. Technol. 2018, 53, 1896–1905. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Rusko, J.; Starkute, V.; Bendoraitiene, E.; Zadeike, D.; Juodeikiene, G. The effect of Pediococcus acidilactici and Lactobacillus sakei on biogenic amines formation and free amino acid profile in different lupin during fermentation. LWT Food Sci. Technol. 2016, 74, 40–47. [Google Scholar] [CrossRef]
- Klupsaite, D.; Juodeikiene, G.; Zadeike, D.; Bartkiene, E.; Maknickiene, Z.; Liutkute, G. The influence of lactic acid fermentation on functional properties of narrow-leaved lupine protein as functional additive for higher value wheat bread. LWT Food Sci. Technol. 2017, 75, 180–186. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Zieliński, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. Antioxidant capacity and polyphenolic content of high-protein lupin products. Food Chem. 2009, 112, 84–88. [Google Scholar] [CrossRef]
- Karamać, M.; Orak, H.H.; Amarowicz, R.; Orak, A.; Piekoszewski, W. Phenolic contents and antioxidant capacities of wild and cultivated white lupin (Lupinus albus L.) seeds. Food Chem. 2018, 258, 1–7. [Google Scholar] [CrossRef]
- Awad, R.A.; Salama, W.M.; Farahat, A.M. Effect of lupine as cheese base substitution on technological and nutritional properties of processed cheese analogue. Acta Sci. Pol. Technol. Aliment. 2014, 13, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Abdel–Salam, A.; Ali, J.; Zayan, A. Effect of Lupine Powder on Rheological, Chemical and Microbiological Properties of Yoghurt. J. Food Dairy Sci. 2015, 6, 531–546. [Google Scholar] [CrossRef]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.-E.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Technol. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]
- Mohamed, S.; Awad, R.; Elbatawy, O.; Salama, W. Production of vegetable yoghurt like from lupin milk. Arab Univ. J. Agric. Sci. 2019, 27, 2155–2165. [Google Scholar] [CrossRef]
- Al-Saedi, N.; Agarwal, M.; Ma, W.; Islam, S.; Ren, Y. Proteomic Characterisation of Lupin (Lupinus angustifolius) Milk as Influenced by Extraction Techniques, Seed Coat and Cultivars. Molecules 2020, 25, 1782. [Google Scholar] [CrossRef]
- Zaouadi, N.; Hadj Ziane-Zafour, A.; Ouazib, M.; Arab, Y.; Hacini, K.; Aslan, S.S. Formulation and Optimization by Experimental Design of Dairy Dessert Based on Lupinus albus L. and Stevia rebaudiana Extracts. Asian J. Dairy Food Res. 2019, 38, 281–287. [Google Scholar]
- Tomczak, A.; Zielińska-Dawidziak, M.; Piasecka-Kwiatkowska, D.; Lampart-Szczapa, E. Blue lupine seeds protein content and amino acids composition. Plant Soil Environ. 2018, 64, 147–155. [Google Scholar]
- Hickisch, A.; Bindl, K.; Vogel, R.F.; Toelstede, S. Thermal treatment of lupin-based milk alternatives—Impact on lupin proteins and the network of respective lupin-based yogurt alternatives. Food Res. Int. 2016, 89, 850–859. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [Green Version]
- Kasprowicz-Potocka, M.; Borowczyk, P.; Zaworska, A.; Nowak, W.; Frankiewicz, A.; Gulewicz, P. The effect of dry yeast fermentation on chemical composition and protein characteristics of blue lupin seeds. Food Technol. Biotechnol. 2016, 54, 360–366. [Google Scholar] [CrossRef]
- Zaworska, A.; Kasprowicz-Potocka, M.; Frankiewicz, A.; Zduńczyk, Z.; Juśkiewicz, J. Effects of fermentation of narrow-leafed lupine (L. angustifolius) seeds on their chemical composition and physiological parameters in rats. J. Anim. Feed Sci. 2016, 25, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Camacho, L.; Sierra, C.; Marcus, D.; Guzmán, E.; Campos, R.; von Bäer, D.; Trugo, L. Nutritional quality of lupine (Lupinus albus cv. Multolupa) as affected by lactic acid fermentation. Int. J. Food Microbiol. 1991, 14, 277–286. [Google Scholar] [CrossRef]
- Fritsch, C.; Vogel, R.F.; Toelstede, S. Fermentation performance of lactic acid bacteria in different lupin substrates-influence and degradation ability of antinutritives and secondary plant metabolites. J. Appl. Microbiol. 2015, 119, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, K.; Leidigkeit, A.; Eisner, P.; Schweiggert-Weisz, U. Technofunctional and sensory properties of fermented lupin protein isolates. Foods 2019, 8, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos, D.C.; de Oliveira Filho, J.G.; Santana, A.C.A.; de Freitas, B.S.M.; Silva, F.G.; Takeuchi, K.P.; Egea, M.B. Optimization of soymilk fermentation with kefir and the addition of inulin: Physicochemical, sensory and technological characteristics. LWT Food Sci. Technol. 2019, 104, 30–37. [Google Scholar] [CrossRef]
- Tsafrakidou, P.; Michaelidou, A.-M.; Biliaderis, C.G. Fermented Cereal-based Products: Nutritional Aspects, Possible Impact on Gut Microbiota and Health Implications. Foods 2020, 9, 734. [Google Scholar] [CrossRef]
- Romero-Espinoza, A.M.; Serna-Saldivar, S.O.; Vintimilla-Alvarez, M.C.; Briones-García, M.; Lazo-Vélez, M.A. Effects of fermentation with probiotics on anti-nutritional factors and proximate composition of lupin (Lupinus mutabilis sweet). LWT Food Sci. Technol. 2020, 130, 109658. [Google Scholar] [CrossRef]
- Van Vo, B.; Bui, D.P.; Nguyen, H.Q.; Fotedar, R. Optimized fermented lupin (Lupinus angustifolius) inclusion in juvenile barramundi (Lates calcarifer) diets. Aquaculture 2015, 444, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Villacrés, E.; Quelal, M.B.; Jácome, X.; Cueva, G.; Rosell, C.M. Effect of debittering and solid-state fermentation processes on the nutritional content of lupine (Lupinus mutabilis Sweet). Int. J. Food Sci. Technol. 2020, 55, 2589–2598. [Google Scholar] [CrossRef]
- Siger, A.; Czubinski, J.; Kachlicki, P.; Dwiecki, K.; Lampart-Szczapa, E.; Nogala-Kalucka, M. Antioxidant activity and phenolic content in three lupin species. J. Food Compos. Anal. 2012, 25, 190–197. [Google Scholar] [CrossRef]
- Zhong, L.; Wu, G.; Fang, Z.; Wahlqvist, M.L.; Hodgson, J.M.; Clarke, M.W.; Junaldi, E.; Johnson, S.K. Characterization of polyphenols in Australian sweet lupin (Lupinus angustifolius) seed coat by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2019, 116, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-López, M.A.; Barrientos-Ramírez, L.; García-López, P.M.; Valdés-Miramontes, E.H.; Zamora-Natera, J.F.; Rodríguez-Macias, R.; Salcedo-Pérez, E.; Bañuelos-Pineda, J.; Vargas-Radillo, J.J. Nutritional and bioactive compounds in mexican lupin beans species: A mini-review. Nutrients 2019, 11, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnoldi, A.; Boschin, G.; Zanoni, C.; Lammi, C. The health benefits of sweet lupin seed flours and isolated proteins. J. Funct. Foods 2015, 18, 550–563. [Google Scholar] [CrossRef]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Antioxidant activities and characterisation of polysaccharides isolated from the seeds of Lupinus angustifolius. Ind. Crops Prod. 2015, 74, 950–956. [Google Scholar] [CrossRef]
- Liu, J.-R.; Chen, M.-J.; Lin, C.-W. Antimutagenic and Antioxidant Properties of Milk–Kefir and Soymilk–Kefir. J. Agric. Food Chem. 2005, 53, 2467–2474. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barac, M.; Vucic, T.; Zilic, S.; Pesic, M.; Sokovic, M.; Petrovic, J.; Kostic, A.; Ignjatovic, I.S.; Milincic, D. The effect of in vitro digestion on antioxidant, ACE-inhibitory and antimicrobial potentials of traditional Serbian white-brined cheeses. Foods 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Salachna, P.; Grzeszczuk, M.; Wilas, J. Total phenolic content, photosynthetic pigment concentration and antioxidant activity of leaves and bulbs of selected Eucomis L’Hér. taxa. Fresenius Environ. Bull. 2015, 24, 4220–4225. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Storage (Days) | |||||
|---|---|---|---|---|---|
| 1 | 4 | 7 | 14 | 21 | |
| LAB CFU/g | |||||
| A | 1.47 × 109 ± 0.18 Ac | 7.05 × 107 ± 1.86 Ba | 1.53 × 108 ± 0.08 Ca | 1.92 × 107 ± 0.10 Da | 4.18 × 108 ± 0.39 Ea |
| B | 2.86 × 108 ± 3.02 Ab | 1.25 × 109 ± 0.26 Ba | 7.54 × 107 ± 1.92 Cb | 3.25 × 107 ± 0.41 Da | 7.45 × 108 ± 2.19 Db |
| C | 1.82 × 107 ± 0.00 Aa | 1.53 × 108 ± 0.30 Bb | 1.13 × 108 ± 0.19 Cab | 1.03 × 109 ± 0.38 Db | 5.18 × 108 ± 5.76 Dc |
| Yeast CFU/g | |||||
| A | 1.47 × 109 ± 0.18 Aa | 3.36 × 107 ± 1.29 Ba | 3.45 × 107 ± 0.07 Ca | 5.87 × 106 ± 7.90 Da | 1.26 × 107 ± 0.06 Ea |
| B | 1.68 × 109 ± 0.02 Aa | 8.59 × 107 ± 0.06 Bb | 1.20 × 108 ± 0.22 Cb | 1.40 × 107 ± 0.19 Db | 6.23 × 107 ± 1.61 Eab |
| C | 3.15 × 109 ± 3.32 Ab | 9.49 × 107 ± 0.45 Bc | 2.25 × 108 ± 0.07 Cc | 1.38 × 108 ± 0.18 Dc | 8.41 × 107 ± 1.35 Eb |
| Time of Storage (Days) | ||||||
|---|---|---|---|---|---|---|
| Unfermented | 1 | 4 | 7 | 14 | 21 | |
| TPC (mg GAE/mL) | ||||||
| A | 10.32 ± 0.05 Aa | 11.60 ± 0.03 Ba | 12.71 ± 0.03 Ca | 16.85 ± 0.03 Da | 18.81 ± 0.31 Ea | 19.32 ± 0.03 Fa |
| B | 15.95 ± 0.03 Ab | 16.45 ± 0.00 Bb | 20.35 ± 0.00 Cb | 24.87 ± 0.07 Db | 27.26 ± 0.03 Eb | 30.54 ± 0.08 Fb |
| C | 17.93 ± 0.08 Ac | 18.94 ± 0.00 Bc | 22.28 ± 0.14 Cc | 25.86 ± 0.03 Dc | 28.71 ± 0.04 Ec | 35.86 ± 0.03 Fc |
| TFC (mg QE/mL) | ||||||
| A | 13.75 ± 0.07 Aa | 15.13 ± 0.04 Ba | 18.47 ± 0.04 Ca | 20.30 ± 0.05 Da | 22.54 ± 0.04 Ea | 23.92 ± 0.04 Fa |
| B | 21.27 ± 0.04 Ab | 22.17 ± 0.04 Bb | 23.42 ± 0.04 Cb | 27.75 ± 0.05 Db | 28.29 ± 0.05 Eb | 33.42 ± 0.04 Fb |
| C | 23.90 ± 0.10 Ac | 31.54 ± 0.07 Bc | 31.17 ± 0.05 Cc | 31.48 ± 0.14 Dc | 32.60 ± 0.08 Ec | 33.89 ± 0.08 Fc |
| AAC (mg/mL) | ||||||
| A | 0.05 ± 0.01 ABa | 0.05 ± 0.00 ABa | 0.04 ± 0.01 Aa | 0.04 ± 0.01 Aa | 0.05 ± 0.01 ABa | 0.06 ± 0.01 BCa |
| B | 0.06 ± 0.01 Aab | 0.04 ± 0.01 ABa | 0.05 ± 0.02 ABa | 0.06 ± 0.01 ABa | 0.06 ± 0.01 Aab | 0.06 ± 0.01 Aa |
| C | 0.07 ± 0.00 Ab | 0.05 ± 0.01 Bb | 0.07 ± 0.00 Ab | 0.07 ± 0.01 Ab | 0.07 ± 0.00 Ab | 0.11 ± 0.01 Cb |
| Time of Storage (Days) | ||||||
|---|---|---|---|---|---|---|
| Unfermented | 1 | 4 | 7 | 14 | 21 | |
| RP (700 nm) | ||||||
| A | 0.449 ± 0.01 Aa | 0.630 ± 0.02 Bb | 0.635 ± 0.02 Cb | 0.422 ± 0.05 Da | 0.628 ± 0.02 Ba | 0.600 ± 0.01 Ea |
| B | 0.513 ± 0.02 Ab | 0.593 ± 0.02 Ba | 0.587 ± 0.01 Ca | 0.753 ± 0.01 Db | 0.732 ± 0.01 Eb | 0.754 ± 0.01 Fb |
| C | 0.621 ± 0.01 Ac | 0.643 ± 0.01 Bc | 0.681 ± 0.01 Cc | 0.782 ± 0.01 Dc | 0.825 ± 0.01 Ec | 0.754 ± 0.01 Fb |
| DPPH inhibition (%) | ||||||
| A | 65.50 ± 0.27 Aa | 67.98 ± 0.22 Ba | 69.79 ± 0.00 Ca | 85.54 ± 0.89 Da | 87.91 ± 0.36 Ea | 91.48 ± 0.06 Fa |
| B | 65.32 ± 0.06 Aa | 87.27 ± 0.07 Bb | 90.35 ± 0.23 Cb | 90.10 ± 0.15 Db | 90.74 ± 0.00 Eb | 92.90 ± 0.27 Fb |
| C | 75.06 ± 0.57 Ab | 88.63 ± 0.06 Bc | 90.79 ± 0.00 Cb | 91.99 ± 0.12 Dc | 93.06 ± 0.00 Ec | 94.27 ± 0.06 Fc |
| ABTS inhibition (%) | ||||||
| A | 42.60 ± 0.06 Aa | 49.53 ± 0.37 Ba | 57.76 ± 0.31 Ca | 64.77 ± 0.39 Da | 70.86 ± 0.34 Ea | 82.05 ± 0.40 Fa |
| B | 44.06 ± 0.14 Ab | 60.23 ± 0.33 Bb | 61.57 ± 0.13 Cb | 72.40 ± 0.13 Db | 78.23 ± 0.05 Eb | 86.35 ± 0.07 Fb |
| C | 47.41 ± 0.10 Ac | 69.28 ± 0.27 Bc | 73.88 ± 0.49 Cc | 79.34 ± 0.21 Dc | 88.72 ± 0.07 Ec | 92.37 ± 0.11 Fc |
| O2− inhibition (%) | ||||||
| A | 32.49 ± 0.09 Aa | 66.67 ± 0.00 Ba | 66.70± 0.03 Ba | 70.32 ± 0.00 Ca | 85.71 ± 0.00 Ca | 87.62 ± 0.00 Da |
| B | 44.29 ± 0.05 Ab | 76.19 ± 0.01 Bb | 78.68 ± 0.18 Cb | 83.92 ± 0.01 Db | 88.41 ± 0.05 Eb | 95.47 ± 0.08 Fc |
| C | 56.40 ± 0.09 Ac | 88.68 ± 0.10 Bc | 88.57 ± 0.05 Bc | 89.42 ± 0.37 Cc | 91.43 ±0.03 Dc | 92.85 ± 0.07 Eb |
| OH inhibition (%) | ||||||
| A | 22.57 ± 0.96 Aa | 27.53 ± 0.67 Ba | 46.87 ± 0.29 Ca | 62.22 ± 0.92 Da | 66.87 ± 0.83 Ea | 52.98 ± 0.41 Fa |
| B | 37.82 ± 0.40 Ab | 33.48 ± 0.57 Bb | 65.10 ± 0.06 Cb | 71.62 ± 0.22 Db | 85.06 ± 0.05 Eb | 79.09 ± 0.06 Fb |
| C | 44.95 ± 0.50 Ac | 41.57 ± 0.89 Bc | 70.56 ± 0.44 Cc | 72.87 ± 0.36 Dc | 89.07 ± 0.20 Ec | 83.67 ± 0.06 Fc |
| Time of Storage (Days) | ||||||
|---|---|---|---|---|---|---|
| Unfermented | 1 | 4 | 7 | 14 | 21 | |
| L* | ||||||
| A | 77.45 ± 0.01 Aa | 81.83 ± 0.01 Ba | 80.03 ± 0.01 Ca | 81.55 ± 0.01 Db | 82.21 ± 0.12 Eb | 81.37 ± 0.02 Fa |
| B | 77.23 ± 0.02 Ab | 82.41 ± 0.01 Bc | 82.18 ± 0.02 Cc | 82.26 ± 0.01 Dc | 82.30 ± 0.02 Ec | 82.52 ± 0.04 Fc |
| C | 76.76 ± 0.02 Ac | 81.65 ± 0.02 Bb | 81.23 ± 0.02 Cb | 80.15 ± 0.02 Da | 81.71 ± 0.03 Ea | 81.64 ± 0.07 Eb |
| a* | ||||||
| A | 2.51 ± 0.01 Aa | 2.68 ± 0.01 Ba | 3.55 ± 0.01 Cc | 2.26 ± 0.01 Da | 2.57 ± 0.01 Ea | 2.38 ± 0.01 Fa |
| B | 2.80 ± 0.01 Ab | 2.66 ± 0.01 Ba | 2.64 ± 0.01 Ca | 2.50 ± 0.01 Dc | 2.63 ± 0.02 Cb | 2.68 ± 0.02 Bb |
| C | 3.18 ± 0.02 Ac | 3.12 ± 0.01 Bb | 3.05 ± 0.01 Cb | 2.35 ± 0.01 Db | 3.21 ± 0.01 Ec | 3.07 ± 0.02 Cc |
| b* | ||||||
| A | 37.15 ± 0.01 Aa | 31.93 ± 0.01 Ba | 35.85 ± 0.01 Cc | 30.97 ± 0.01 Da | 31.08 ± 0.07 Ea | 30.31 ± 0.01 Fa |
| B | 37.09 ± 0.01 Ab | 32.00 ± 0.02 Bb | 31.61 ± 0.01 Cb | 32.15 ± 0.01 Dc | 31.09 ± 0.02 Ea | 31.34 ± 0.05 Fb |
| C | 37.13 ± 0.02 Aa | 32.04 ± 0.02 Bb | 31.35 ± 0.01 Ca | 31.12 ± 0.02 Db | 31.84 ± 0.01 Eb | 31.48 ± 0.02 Fc |
| Time of Storage (Days) | ||||||
|---|---|---|---|---|---|---|
| Unfermented | 1 | 4 | 7 | 14 | 21 | |
| Viscosity (MPa·s) | ||||||
| A | 422.57 ± 0.05 Aa | 433.33 ± 0.40 Ba | 948.00 ± 0.67 Ca | 481.00 ± 0.58 Da | 399.50 ± 1.00 Ea | 399.50 ± 0.28 Ea |
| B | 766.82 ± 0.40 Ab | 811.00 ± 0.21 Bb | 1030.00 ± 5.77 Cb | 829.00 ± 2.00 Db | 707.00 ± 1.73 Eb | 704.00 ± 4.04 Fb |
| C | 1044.95 ± 0.50 Ac | 1038.67 ± 0.11 Bc | 2846.00 ± 1.33 Cc | 1182.50 ± 0.50 Dc | 1172.50 ± 0.86 Ec | 1083.67 ± 0.06 Fc |
| Firmness (N) | ||||||
| A | 0.19 ± 0.07 Aa | 0.27 ± 0.07 Ba | 0.39 ± 0.07 Ca | 0.26 ± 0.05 Da | 0.19 ± 0.06 Eb | 0.11 ± 0.04 Fa |
| B | 0.47 ± 0.04 Ab | 0.55 ± 0.04 Bb | 0.65 ± 0.07 Cb | 0.35 ± 0.03 Db | 0.18 ± 0.02 Ea | 0.11 ± 0.03 Fa |
| C | 0.82 ± 0.10 Ac | 0.90 ± 0.10 Bc | 1.72 ± 0.23 Cc | 1.14 ± 0.14 Dc | 1.12 ± 0.26 Ec | 0.70 ± 0.08 Fb |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łopusiewicz, Ł.; Drozłowska, E.; Trocer, P.; Kwiatkowski, P.; Bartkowiak, A.; Gefrom, A.; Sienkiewicz, M. The Effect of Fermentation with Kefir Grains on the Physicochemical and Antioxidant Properties of Beverages from Blue Lupin (Lupinus angustifolius L.) Seeds. Molecules 2020, 25, 5791. https://doi.org/10.3390/molecules25245791
Łopusiewicz Ł, Drozłowska E, Trocer P, Kwiatkowski P, Bartkowiak A, Gefrom A, Sienkiewicz M. The Effect of Fermentation with Kefir Grains on the Physicochemical and Antioxidant Properties of Beverages from Blue Lupin (Lupinus angustifolius L.) Seeds. Molecules. 2020; 25(24):5791. https://doi.org/10.3390/molecules25245791
Chicago/Turabian StyleŁopusiewicz, Łukasz, Emilia Drozłowska, Paulina Trocer, Paweł Kwiatkowski, Artur Bartkowiak, Annett Gefrom, and Monika Sienkiewicz. 2020. "The Effect of Fermentation with Kefir Grains on the Physicochemical and Antioxidant Properties of Beverages from Blue Lupin (Lupinus angustifolius L.) Seeds" Molecules 25, no. 24: 5791. https://doi.org/10.3390/molecules25245791