
Thermostable Tannase from Aspergillus Niger and Its Application in the Enzymatic Extraction of Green Tea

Abstract
:1. Introduction
2. Results and Discussion
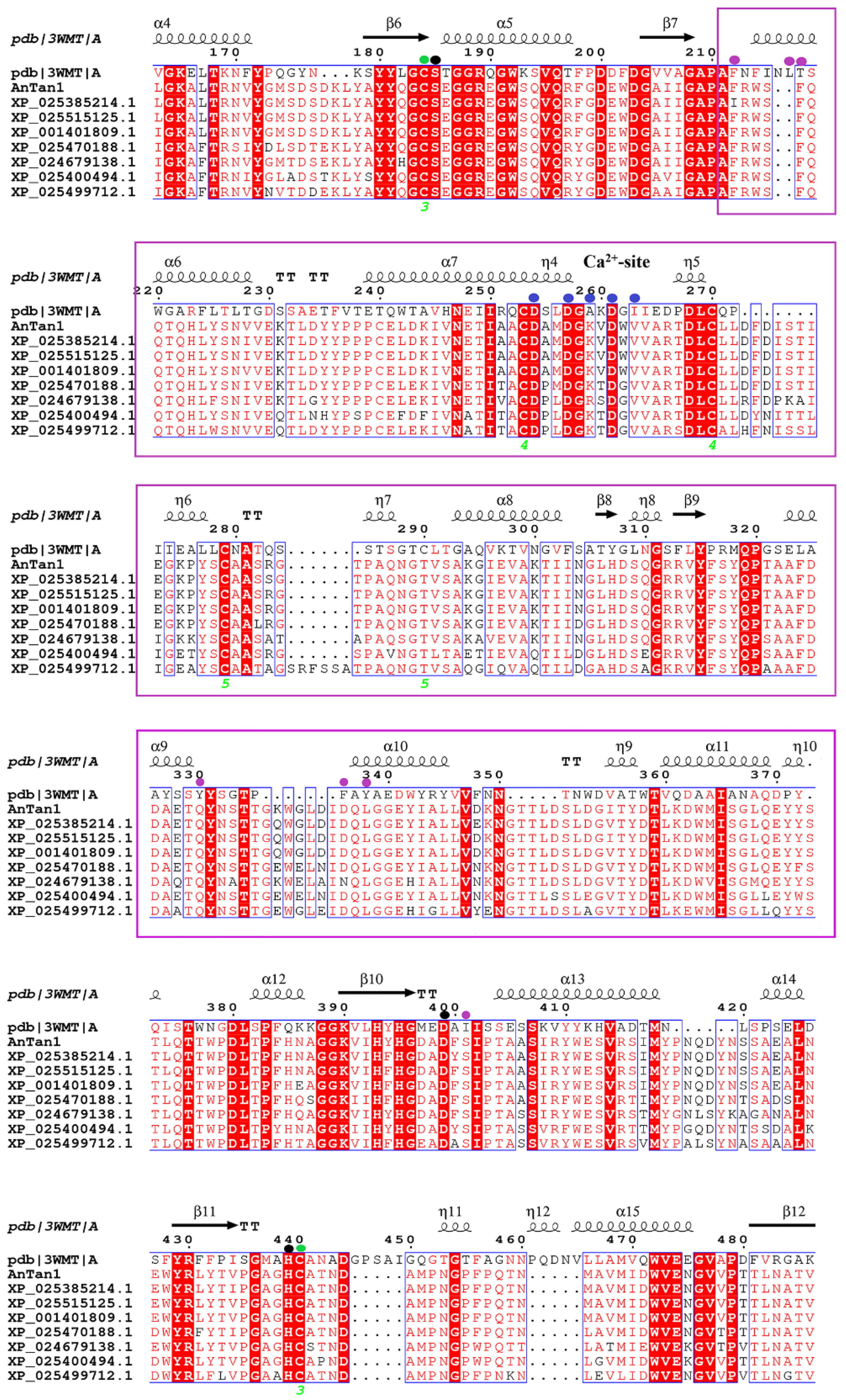
2.1. Analysis of Bioinformatics and Cloning of A. Niger Tannase
2.2. Tannase Production
2.3. Enzymatic Characteristics
2.3.1. Effects of Temperature and pH on Enzymatic Activity and Stability
2.3.2. Effects of Metal Ions, Surfactants, Enzyme Inhibitors, and Solvents on Enzyme Activity
2.3.3. Substrate Specificity
2.3.4. Kinetic Parameters
2.4. Application of Tannase in Tea Extraction
3. Materials and Methods
3.1. Microorganism
3.2. Construction of Plasmid
3.3. Production of Tannase
3.4. Activity Assay of Tannase
3.5. Enzyme Purification
3.6. Characterization of Tannase
3.6.1. Effects of Temperature and pH
3.6.2. Effect of Metal Ions and Other Additives on Enzyme Activity
3.6.3. Effects of Organic Solvents on Enzyme Activity
3.6.4. Substrate Specificity and Kinetic Constants of Tannase
3.7. Application of Tannase
3.7.1. Enzymatic Extraction of Green Tea
3.7.2. Determination of Catechins and Caffeine by HPLC
3.7.3. Total Polyphenols Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Y.; Yin, J.; Chen, J.; Wang, F.; Du, Q.; Jiang, Y.; Xu, Y. Improving the sweet aftertaste of green tea infusion with tannase. Food Chem. 2016, 192, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Chu, S.; Yan, L.; Chen, C. Effect of tannase treatment on protein–tannin aggregation and sensory attributes of green tea infusion. Lwt - Food Sci. Technol. 2009, 42, 338–342. [Google Scholar] [CrossRef]
- Ananingsih, V.K.; Sharma, A.; Zhou, W. Green tea catechins during food processing and storage: A review on stability and detection. Food Res. Int. 2013, 50, 469–479. [Google Scholar] [CrossRef]
- Perva-Uzunalić, A.; Škerget, M.; Knez, Ž.; Weinreich, B.; Otto, F.; Grüner, S. Extraction of active ingredients from green tea (Camellia sinensis): Extraction efficiency of major catechins and caffeine. Food Chem. 2006, 96, 597–605. [Google Scholar] [CrossRef]
- Chandini, S.K.; Rao, L.J.; Gowthaman, M.K.; Haware, D.J.; Subramanian, R. Enzymatic treatment to improve the quality of black tea extracts. Food Chem. 2011, 127, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Dubey, K.K.; Janve, M.; Ray, A.; Singhal, R.S. Ready-to-Drink Tea. In Trends in Non-alcoholic Beverages; Academic Press: Cambridge, MA, USA, 2020; pp. 101–140. [Google Scholar]
- Hong, Y.H.; Jung, E.Y.; Park, Y.; Shin, K.S.; Kim, T.Y.; Yu, K.W.; Chang, U.J.; Suh, H.J. Enzymatic improvement in the polyphenol extractability and antioxidant activity of green tea extracts. Biosci. Biotechnol. Biochem. 2013, 77, 22–29. [Google Scholar] [CrossRef]
- Murugesh, C.S.; Subramanian, R. Applications of Enzymes in Processing Green Tea Beverages: Impact on Antioxidants - Processing and Impact on Antioxidants in Beverages. In Processing and Impact on Antioxidants in Beverages; Academic Press: Cambridge, MA, USA, 2014; pp. 99–108. [Google Scholar]
- Ni, H.; Chen, F.; Jiang, Z.D.; Cai, M.Y.; Yang, Y.F.; Xiao, A.F.; Cai, H.N. Biotransformation of tea catechins using Aspergillus niger tannase prepared by solid state fermentation on tea byproduct. Lwt - Food Sci. Technol. 2015, 60, 1206–1213. [Google Scholar] [CrossRef]
- Cao, Q.; Zou, C.; Zhang, Y.; Du, Q.; Yin, J.; Shi, J.; Xue, S.; Xu, Y. Improving the taste of autumn green tea with tannase. Food Chem. 2019, 277, 432–437. [Google Scholar] [CrossRef]
- Baik, J.H.; Shin, K.; Park, Y.; Yu, K.; Suh, H.J.; Choi, H. Biotransformation of catechin and extraction of active polysaccharide from green tea leaves via simultaneous treatment with tannase and pectinase. J. Sci. Food Agr. 2015, 95, 2337–2344. [Google Scholar] [CrossRef]
- Jana, A.; Maity, C.; Halder, S.K.; Das, A.; Pati, B.R.; Mondal, K.C.; Das Mohapatra, P.K. Structural characterization of thermostable, solvent tolerant, cytosafe tannase from Bacillus subtilis PAB2. Biochem. Eng. J. 2013, 77, 161–170. [Google Scholar] [CrossRef]
- Gonçalves, H.B.; Riul, A.J.; Terenzi, H.F.; Jorge, J.A.; Guimarães, L.H.S. Extracellular tannase from Emericella nidulans showing hypertolerance to temperature and organic solvents. J. Mol. Catal. B: Enzym. 2011, 71, 29–35. [Google Scholar] [CrossRef]
- Mata-Gomez, M.; Rodriguez, L.V.; Ramos, E.L.; Renovato, J.; Cruz-Hernandez, M.A.; Rodriguez, R.; Contreras, J.; Aguilar, C.N. A novel tannase from the xerophilic fungus Aspergillus niger GH1. J. Microbiol. Biotechnol. 2009, 19, 987–996. [Google Scholar] [PubMed] [Green Version]
- Ramirez-Coronel, M.A.; Viniegra-Gonzalez, G.; Darvill, A.; Augur, C. A novel tannase from Aspergillus niger with beta-glucosidase activity. Microbiology 2003, 149, 2941–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabu, A.; Kiran, G.S.; Pandey, A. Purification and Characterization of Tannin Acyl Hydrolase from Aspergillus niger ATCC 16620. Food Technol. Biotechnol. 2005, 43, 133–138. [Google Scholar]
- Ozturk, B.; Seyhan, F.; Ozdemir, I.S.; Karadeniz, B.; Bahar, B.; Ertas, E.; Ilgaz, S. Change of enzyme activity and quality during the processing of Turkish green tea. Lwt - Food Sci. Technol. 2016, 65, 318–324. [Google Scholar] [CrossRef]
- Lu, M.; Chen, C. Enzymatic modification by tannase increases the antioxidant activity of green tea. Food Res. Int. 2008, 41, 130–137. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Koseki, T.; Ichikawa, K.; Sasaki, K.; Shiono, Y. Characterization of a novel Aspergillus oryzae tannase expressed in Pichia pastoris. J. Biosci. Bioeng. 2018, 126, 553–558. [Google Scholar] [CrossRef]
- Suzuki, K.; Hori, A.; Kawamoto, K.; Thangudu, R.R.; Ishida, T.; Igarashi, K.; Samejima, M.; Yamada, C.; Arakawa, T.; Wakagi, T.; et al. Crystal structure of a feruloyl esterase belonging to the tannase family: A disulfide bond near a catalytic triad. Proteins: Struct. Funct. Bioinform. 2014, 82, 2857–2867. [Google Scholar] [CrossRef]
- Liu, F.; Wang, B.; Ye, Y.; Pan, L. High level expression and characterization of tannase tan7 using Aspergillus niger SH-2 with low-background endogenous secretory proteins as the host. Protein Expres. Purif. 2018, 144, 71–75. [Google Scholar] [CrossRef]
- Amanda, R.D.S.; Gouveia, M.J.; Tonny, C.C.L.; Aparecida Moreira, K.; Sandra, A.D.A. Production, Characterization and Application of a Thermostable Tannase from Pestalotiopsis guepinii URM 7114. Food Technol. Biotechnol. 2014, 52, 459–467. [Google Scholar]
- Selwal, M.K.; Selwal, K.K. High-level tannase production by Penicillium atramentosum KM using agro residues under submerged fermentation. Ann. Microbiol. 2012, 62, 139–148. [Google Scholar] [CrossRef]
- Kumar, M.; Beniwal, V.; Salar, R.K. Purification and characterization of a thermophilic tannase from Klebsiella pneumoniae KP715242. Biocatal. Agric. Biotechnol. 2015, 4, 745–751. [Google Scholar] [CrossRef]
- Abdel-Naby, M.A.; El-Tanash, A.B.; Sherief, A.D.A. Structural characterization, catalytic, kinetic and thermodynamic properties of Aspergillus oryzae tannase. Int. J. Biol. Macromol. 2016, 92, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Gayen, S.; Ghosh, U. Purification and Characterization of Tannin Acyl Hydrolase Produced by Mixed Solid State Fermentation of Wheat Bran and Marigold Flower by Penicillium notatum NCIM 923. Biomed. Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, T.; Shiono, Y.; Koseki, T. Biochemical characterization of Aspergillus oryzae native tannase and the recombinant enzyme expressed in Pichia pastoris. J. Biosci. Bioeng. 2014, 118, 392–395. [Google Scholar] [CrossRef]
- Qiu, Y.; Niu, H.; Huang, W.; He, Y.; Wu, X. Properties and secondary structure of tannase from Penicillium herquei. Biotechnol. Bioproc. E 2011, 16, 858–866. [Google Scholar] [CrossRef]
- Riul, A.J.; Gonçalves, H.B.; Jorge, J.A.; Guimarães, L.H.S. Characterization of a glucose- and solvent-tolerant extracellular tannase from Aspergillus phoenicis. J. Mol. Catal. B: Enzym. 2013, 85, 126–133. [Google Scholar] [CrossRef]
- Mahmoud, A.E.; Fathy, S.A.; Rashad, M.M.; Ezz, M.K.; Mohammed, A.T. Purification and characterization of a novel tannase produced by Kluyveromyces marxianus using olive pomace as solid support, and its promising role in gallic acid production. Int. J. Biol. Macromol. 2018, 107, 2342–2350. [Google Scholar] [CrossRef]
- El-Tanash, A.B.; Sherief, A.A.; Nour, A. Catalytic properties of immobilized tannase produced from Aspergillus aculeatus compared with the free enzyme. Braz. J. Chem. Eng. 2011, 28, 381–391. [Google Scholar] [CrossRef]
- Chhokar, V.; Sangwan, M.; Beniwal, V.; Nehra, K.; Nehra, K.S. Effect of Additives on the Activity of Tannase from Aspergillus awamori MTCC9299. Appl. Biochem. Biotech. 2010, 160, 2256–2264. [Google Scholar] [CrossRef]
- Castiglioni, S.; Damiani, E.; Astolfi, P.; Carloni, P. Influence of steeping conditions (time, temperature, and particle size) on antioxidant properties and sensory attributes of some white and green teas. Int. J. Food Sci. Nutr. 2015, 66, 491–497. [Google Scholar] [CrossRef]
- Kim, E.S.; Liang, Y.R.; Jin, J.; Sun, Q.F.; Lu, J.L.; Du, Y.Y.; Lin, C. Impact of heating on chemical compositions of green tea liquor. Food Chem. 2007, 103, 1263–1267. [Google Scholar] [CrossRef]
- Xu, Y.; Ji, W.; Yu, P.; Chen, J.; Wang, F.; Yin, J. Effect of extraction methods on the chemical components and taste quality of green tea extract. Food Chem. 2018, 248, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Venditti, E.; Bacchetti, T.; Tiano, L.; Carloni, P.; Greci, L.; Damiani, E. Hot vs. cold water steeping of different teas: Do they affect antioxidant activity? Food Chem. 2010, 119, 1597–1604. [Google Scholar] [CrossRef]
- Labbé, D.; Tremblay, A.; Bazinet, L. Effect of brewing temperature and duration on green tea catechin solubilization: Basis for production of EGC and EGCG-enriched fractions. Sep. Purif. Technol. 2006, 49, 1–9. [Google Scholar] [CrossRef]
- Yang, D.; Hwang, L.S.; Lin, J. Effects of different steeping methods and storage on caffeine, catechins and gallic acid in bag tea infusions. J. Chromatogr. A 2007, 1156, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xiao, Q.; Huang, Y.; Ni, H.; Wu, C.; Xiao, A. Tannase application in secondary enzymatic processing of inferior Tieguanyin oolong tea. Electron. J. Biotechn. 2017, 28, 87–94. [Google Scholar] [CrossRef]
- García-Conesa, M.T.; Østergaard, P.; Kauppinen, S.; Williamson, G. Hydrolysis of diethyl diferulates by a tannase from Aspergillus oryzae. Carbohyd. Polym. 2001, 44, 319–324. [Google Scholar] [CrossRef]
- Sharma, S.; Bhat, T.K.; Dawra, R.K. A Spectrophotometric Method for Assay of Tannase Using Rhodanine. Anal. Biochem. 2000, 279, 85–89. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The Determination of Enzyme Dissociation Constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agr. 1959, 10, 63–68. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Mw | Km (1) Tannic Acid (2) Methyl-Gallate (3) Propyl-Gallate | pH (1) Optima (2) Stability | Temperature (1) Optima (2) Stability (3) Residual Enzyme Activity at 60 °C for 1h | Ref. |
|---|---|---|---|---|---|
| Penicillium notatum | 97 kDa, 43 kDa (Dimeric) | 3.3 mM (1) | (1) 5.0 (2) 3.0–8.0 | (1) 35–40 °C (2) up to 60 °C (3) 60% | Gayen and Ghosh 2013 |
| Emericella nidulans | 302 kDa | 7.69 mM (3) | (1) 5.0 (2) 4.0–5.0 | (1) 45 °C (2) 22–50 °C | Gonçalves et al. 2011 |
| Aspergillus oryzae | 45–80 kDa | n.s. | (1) 4.0–5.0 | (1) 40 °C (2) up to 40 °C | Mizuno et al. 2014 |
| Aspergillus phoenicis | 120 kDa, 93 kDa (Dimeric) | 1.7 mM (1), 14.3 mM (2) and 0.6 mM (3) | (1) 6.0 (2) 2.5–7.0 | (1) 60 °C (2) 40–60 °C (3) 10% | Riul et al. 2013 |
| Penicillium herquei | 72 kDa | 0.62 mM (3) | (1) 6.0 | (1) 30 °C | Qiu et al. 2011 |
| Aspergillus niger SH-2 | 60–70 kDa | n.s. | (1) 7.0 | (1) 40 °C | Liu et al. 2018 |
| Aspergillus oryzae | 90–120 kDa | 1.11 mM (2) | (1) 6.0 | (1) 30–35 °C (2) up to 40 °C (3) 20% | Koseki et al. 2018 |
| Aspergillus niger ATTC 16620 | 168 kDa | n.s. | (1) 6.0 (2) 4.0–8.0 | (1) 40 °C (2) 30–40 °C | Sabu et al. 2005 |
| Aspergillus oryzae | 106 kDa | 3.13 mM (1) | (1) 5.5 (2) 4.5–7.5 | (1) 37 °C | Abdel-Naby et al. 2016 |
| Aspergillus niger | 85 kDa | 0.073 mM (3) | (1) 6.0 (2) 3.0–8.0 | (1) 80 °C (2) 30–60 °C (3) 91.9% | This work |
| Residual Activity (%) * | |||
|---|---|---|---|
| 1 mM (0.1%) | 10 mM (1%) | ||
| Metal ions | Control | 100 ± 3 h | 100 ± 2 h |
| Na+ | 86 ± 6 e,f | 82 ± 1 e,f | |
| Mg2+ | 89 ± 5 f,g | 76 ± 6 d,e | |
| Cu2+ | 74 ± 7 d,e | 34 ± 8 a | |
| Ba2+ | 84 ± 7 e,f | 83 ± 9 e,f | |
| Zn2+ | 85 ± 2 e,f | 86 ± 2 e,f | |
| Cd2+ | 78 ± 1 d,e,f | 88 ± 4 f,g | |
| Ca2+ | 67 ± 6 b,c,d | 75 ± 8 d,e | |
| Mn2+ | 88 ± 7 f,g | 61 ± 1 b,c | |
| K+ | 107 ± 10 h | 89 ± 2 f,g | |
| Al3+ | 70 ± 4 c,d | 58 ± 8 b | |
| Surfactant and inhibitor | Control | 100 ± 1 a | 100 ± 1 a |
| Tween-80 | 89 ± 0 b,c | 88 ± 4 c | |
| Tween-20 | 86 ± 2 d | 98 ± 0 a | |
| Triton X-100 | 92 ± 1 b | 88 ± 2 c,d | |
| β-Mercaptoethanol | 23 ± 1 f | 5 ± 0 g | |
| SDS | 85 ± 1 d | 69 ± 3 e | |
| EDTA | 89 ± 1 c | 91 ± 4 b,c | |
| Solvent | Residual Activity (%) * | ||
|---|---|---|---|
| 20% | 40% | 60% | |
| Control | 100 ± 3 g,h | 100 ± 4 g,h | 100 ± 3 g,h |
| Dimethyl sulfoxide | 98 ± 1 g | 43 ± 1 c | 0 ± 0 a |
| N-hexane | 128 ± 10 k | 156 ± 5 l | 123 ± 9 j,k |
| N-butanol | 62 ± 4 e | 45 ± 2 c,d | 26 ± 0 b |
| Cyclohexane | 124 ± 13 j,k | 257 ± 2 n | 188 ± 27 m |
| N-propanol | 52 ± 3 c,d,e | 4 ± 2 a | 0 ± 0 a |
| Isoamyl alcohol | 106 ± 12 g,h,i | 85 ± 1 f | 56 ± 2 d,e |
| Benzene | 116 ± 4 i,j,k | 112 ± 4 h,I,j | 102 ± 2 g,h |
| Methanol | 52 ± 3 c,d,e | 23 ± 4 b | 11 ± 0 a |
| Trichloromethane | 86 ± 6 f | 85 ± 2 f | 85 ± 8 f |
| Substrate | Tannase Activity (μmol·mL/min) * |
|---|---|
| PG | 219 ± 2 c |
| EGCG | 103 ± 13 a |
| TA | 159 ± 3 b |
| ECG | 221 ± 2 c |
| CG | 231 ± 6 c |
| Treatment | Contents of Single Phenolic Compounds (mg·L−1) by HPLC | Total Phenolics Contents by Folin-Ciocalteu Method (g·kg−1–GT *) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| GA | Caffeine | EGC | EGCG | EC | ECG | Non-ester | Ester | ||
| Enzymatic extraction | 1906 ± 24 a | 612 ± 8 a | 1951 ± 5 a | 0 ± 0 b | 878 ± 14 a | 0 ± 0 b | 2829 a | 0 b | 291 ± 5 a |
| Water extraction | 136 ± 2 b | 600 ± 9 a | 361 ± 5 b | 2687 ± 11 a | 130 ± 3 b | 980 ± 3 a | 491 b | 3666 a | 137 ± 1 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Zhang, Y.-H.; Zhang, F.; Yang, Q.-M.; Weng, H.-F.; Xiao, Q.; Xiao, A.-F. Thermostable Tannase from Aspergillus Niger and Its Application in the Enzymatic Extraction of Green Tea. Molecules 2020, 25, 952. https://doi.org/10.3390/molecules25040952
Shao Y, Zhang Y-H, Zhang F, Yang Q-M, Weng H-F, Xiao Q, Xiao A-F. Thermostable Tannase from Aspergillus Niger and Its Application in the Enzymatic Extraction of Green Tea. Molecules. 2020; 25(4):952. https://doi.org/10.3390/molecules25040952
Chicago/Turabian StyleShao, Yuan, Yong-Hui Zhang, Feng Zhang, Qiu-Ming Yang, Hui-Fen Weng, Qiong Xiao, and An-Feng Xiao. 2020. "Thermostable Tannase from Aspergillus Niger and Its Application in the Enzymatic Extraction of Green Tea" Molecules 25, no. 4: 952. https://doi.org/10.3390/molecules25040952