
Non-Saccharomyces as Biotools to Control the Production of Off-Flavors in Wines



enotecUPM, Departamento de Química y Tecnología de Alimentos, Universidad Politécnica de Madrid, Avenida Puerta de Hierro 2, 28040 Madrid, Spain
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(15), 4571; https://doi.org/10.3390/molecules26154571
Submission received: 7 July 2021
/
Revised: 22 July 2021
/
Accepted: 26 July 2021
/
Published: 28 July 2021
(This article belongs to the Special Issue Wine Sensory Faults: Origin, Prevention and Removal)
Abstract
:Off-flavors produced by undesirable microbial spoilage are a major concern in wineries, as they affect wine quality. This situation is worse in warm areas affected by global warming because of the resulting higher pHs in wines. Natural biotechnologies can aid in effectively controlling these processes, while reducing the use of chemical preservatives such as SO2. Bioacidification reduces the development of spoilage yeasts and bacteria, but also increases the amount of molecular SO2, which allows for lower total levels. The use of non-Saccharomyces yeasts, such as Lachancea thermotolerans, results in effective acidification through the production of lactic acid from sugars. Furthermore, high lactic acid contents (>4 g/L) inhibit lactic acid bacteria and have some effect on Brettanomyces. Additionally, the use of yeasts with hydroxycinnamate decarboxylase (HCDC) activity can be useful to promote the fermentative formation of stable vinylphenolic pyranoanthocyanins, reducing the amount of ethylphenol precursors. This biotechnology increases the amount of stable pigments and simultaneously prevents the formation of high contents of ethylphenols, even when the wine is contaminated by Brettanomyces.
1. Introduction
Wine quality is strongly and negatively affected by some microbial metabolites with low sensory thresholds and negative olfactory impact, including reduced sulfur compounds [1,2], volatile acidity [3], ethylphenols [4,5], and acetaldehyde [6]. The sensory impact is quite variable because sensory thresholds can range from very low values (H2S, 1.6 μg/L) to very high concentrations (volatile acidity 0.3–0.6 g/L), so the range is about 1 million times (Table 1). This makes the analytical approach very specific and makes the use of sensitive and reproducible techniques based mainly on gas chromatography–mass spectrometry (GC-MS or GC-MS/MS) instruments essential for their determination [7]. These analytical methods often require specific sample preparations and concentrations: headspace (HS), dynamic headspace (DHS), solid-phase microextraction (SPME), and Twister [7].
The control of fermentative purity as well as the development of wild spoilage microorganisms in wines are related to pH and sulfur dioxide contents. In warm areas affected by global warming, pH values have been increasing in recent years, which are associated with higher alcoholic strength and intense phenolic ripening [12,13,14], especially in varieties that accumulate high potassium contents in berries [15,16]. A high pH produces wines that are more chemically and microbiologically unstable, and therefore are more susceptible to microbial spoilage, including off-flavor formation. Wine pH can range from 2.8 to 4.5, although most wines are in the 3 to 4 range. However, wines below 3.5 are very stable and usually less affected by microbial developments, while wines with pHs close to 4 are very risky as many spoilage bacteria and yeasts can easily develop in them during processing and especially during aging and storage.
Yeast species can help in the biocontrol of off-flavor formation via bioprotection as a result of competition with or the elimination of wild spoilage microorganisms [17,18,19], by acidity production [20,21,22,23], by nutrient competition and depletion [24,25], by the depletion of off-flavor precursors [26], or by the adsorption of defective molecules on cell walls [27,28] (Figure 1).
This review is focused on the elimination of off-flavors by using non-Saccharomyces yeasts that are able to control pH by bioacidification or to decrease the concentration of precursors of molecules responsible for sensory defects.
2. Bioprotection
Bioprotection is a current concept, so its definition is still under discussion. However, it can be considered the active or passive use of some microorganisms to preserve foods and beverages and to exclude other spoilage microorganisms, thus avoiding the production of off-flavors, sensory alterations, or even the formation of toxic molecules. Bioprotection is a hot topic in enology and foods; several reviews have recently been published [29,30,31]. Bioprotection can be achieved by the production of molecules or metabolites with antimicrobial effects such as organic acids [32]; toxins such as killer factors [33,34,35]; deleterious chelates such as pulcherrimin [25,36]; glucanases [30,37]; ethanol produced in the fermentation of sugars; as well as by nutrient depletion [38] (Figure 1).
The application of bioprotective microorganisms can be scheduled at several stages of the winemaking process [31]. During the prefermentative stage or during prefermentative maceration, they can be applied directly on the grapes (harvesting machine) to control wild yeast [39,40], mold, and bacteria populations [19,41]. During fermentation, they can be applied to control the development of spoilage yeasts and bacteria and the oxidative processes, and frequently also to improve the sensory profile of wines by producing flavor compounds [18,42]. Finally, they can be applied after fermentation to protect and stabilize wines during barrel and bottle aging.
Bioprotection has been proposed as an effective biotool to reduce SO2 levels in wines [19,41]. The non-Saccharomyces Torulaspora delbrueckii (Td) and Metschnikowia pulcherrima (Mp) have been used to produce industrial fermentations without added sulfites. These bioprotective non-Saccharomyces may control some spoilage microorganisms and prevent chemical and enzymatic oxidation [41,43].
Regarding off-flavor formation, Metschnikowia fructicola has been successfully used to reduce the production of ethyl acetate by apiculate yeasts such as Hanseniaspora uvarum during cold soak [44].
3. Bioacidification by Lachancea thermotolerans (Lt)
Acidification and pH control are key tools in enology to preserve wine stability and prevent microbial spoilage. Tartaric acid, the strongest acid in grapes, is systematically used in many wines, particularly in warm areas, to improve chemical stability, enhance color and stabilize anthocyanins, increase the levels of active molecular SO2, and improve wine freshness [45]. Additionally, other acids such as malic, lactic, and citric acid can be used in enology, as can alternative physicochemical processes such as exchange resins and electrodialysis [45]. Acidification with up to 4 g/L tartaric acid is allowed in wines [9].
Bioacidification with Saccharomyces cerevisiae (Sc) through malic acid production has been previously studied. Some Sc strains can produce up to 1 g/L [46] when acidification occurs at the beginning of fermentation (days 2–6). However, the production at the highest level takes place in musts with low malic acid content and low acidity. Moreover, the effect on pH from increasing malic acid by 1 g/L is low and can be degraded by lactic acid bacteria, and thus malic acidification by Sc is not an effective biotechnology in winemaking. Other acids such as lactic, fumaric, and citric acids are also produced by Sc, but at low concentrations and with little impact on wine pH.
Lactic acid is also used for wine acidification, and the sensory effect is better in postfermentation acidifications. Even when often associated with dairy products, the sensory profile of lactic acid is fresh and citric [47]. Lactic acid is also authorized by the OIV for wine acidification. However, the use of lactic acid bioproduction by Lachancea thermotolerans is a natural and powerful biotool to control wine pH [20,21,22,23,48,49]. Some strains are capable of producing more than 16 g/L [21]. This amount is likely to be excessive in enological applications, and thus the use of strains with yields ranging from 5 to 8 g/L may be more appropriate [50,51,52]. With these conditions, it is easy to achieve pH reductions of 0.4–0.5 units [50,51,52], which is more effective than the usual acidification with tartaric acid. The production of L(+)-lactic acid [53] is done by the metabolization of sugars so that some reduction in alcoholic strength can be obtained, ranging from 0.2 [50] to 0.9% vol. [52]. Additionally, this strong reduction in pH favors higher levels of molecular SO2 under enological conditions (Table 2), which is very effective in controlling undesirable spoilage microorganisms and off-flavor production. The usual total SO2 contents (40–60 mg/L) at the typical high pH of grape juice in warm areas (3.7–4.0) can produce ineffective molecular SO2 contents (<0.5 mg/L) (Table 2). These conditions are suitable for the development of spoilage microorganisms that may increase the content of off-flavors or even allergenic or toxic molecules in wines, such as biogenic amines or ethyl carbamate, which can increase during wine aging. The same levels of total SO2 at pH 3.4–3.5 that can be obtained by Lt acidification during fermentation can easily produce molecular SO2 levels above 0.8 mg/L, resulting in a more protective effect and a safer situation (Table 2).
Therefore, Lt fermentations are a potent biotool to promote wine stability by reducing pH and increasing molecular SO2 levels. The use of Lt in sequential fermentations produces a significant reduction in pH and a concomitant effect on molecular SO2 (Figure 2). To produce complete fermentations without residual sugars, it is necessary to inoculate some non-Saccharomyces yeasts because most Lt strains have fermentative powers ranging from 7% to 9% v/v [20,23].
A typical stage in the stabilization of red wines is the need to carry out malolactic fermentation (MLF) and improve the sensory profile. However, MLF results in a reduction of pH, and in warm areas the sensory perception can be flat with a less crispy acidity sensation in the mouth. In these winemaking regions, it could be interesting to preserve malic acidity by inhibiting MLF and simultaneously lower the pH by producing lactic acid with Lt. It should be noted that lactic acid is a strong inhibitor of MLF at high doses, which occurs in many enzymatic processes and is known as product inhibition. It has been observed that lactic acid concentrations above 4 g/L produce a strong inhibition of MLF [32] and significantly decrease lactic acid bacteria populations. At lower values (2 g/L), a significant delay of MLF is observed [32]. Therefore, in addition to pH control by Lt acidification, effective inhibition of MLF can be obtained when lactic acid production is higher than 4 g/L. Furthermore, other malic-acid-preserving additives such as fumaric acid or chitosan can be used to control MLF [54]; the former is in the final stages of evaluation at the OIV [55] and the latter is also authorized for organic wines. Additionally, it has been observed that fumaric acid production can be increased by engineered Sc to more than 5 g/L [56]. Overproduction is done by overexpression of the RoPYC gene, so perhaps in some countries where the use of engineered yeasts is allowed, it can aid in inhibiting MLF along with Lt.
Another interesting application of Lt is the control of volatile acidity levels and most likely of ethyl acetate contents as well [57]. In addition, several authors have reported low volatile acidity contents (<0.5 g/L) in sequential fermentations with Lt [20,22,50,58], even in ternary fermentations with Lt and other non-Saccharomyces, such as Metschnikowia pulcherrima, Torulaspora delbrueckii or Hanseniaspora vineae [59], and ethyl acetate contents similar to Sc controls [50].
Lactic acid production and pH reduction by Lt also have concomitant effects on color due to the increased amounts of pyrylium cation in the wine, resulting in a hyperchromic effect and color protection [60,61].
Furthermore, Lt fermentations have shown preliminary positive effects on Brettanomyces control, likely due to acidification and the high contents of lactate [52].
4. Apiculate Yeasts and Volatile Acidity
Traditionally, apiculate yeasts (Hanseniaspora/Kloeckera species), usually involved in the early fermentation phases, have been considered overproducers of volatile acidity and ethyl acetate [62]. Pure culture fermentations of Hanseniaspora uvarum and Kloeckera apiculata have been reported to produce up to 0.98 and 1.5 g/L acetic acid, and 408 and 225 mg/L acetoin, respectively. In fact, they usually release high contents of acetate esters during fermentation; such is the case for the accumulation of ethyl acetate with concentrations between 450 and 760 mg/L. However, not all species behave in the same way and some of them, such as Hanseniaspora vineae (Hv), have shown a high ability to decrease volatile acidity in sequential fermentations with Saccharomyces cerevisiae compared to single Sc fermentations [63,64]. In triplicate fermentations of white wines, the Sc control produced 0.45 g/L acetic acid, but the sequential fermentation with Hv/Sc produced 0.36 g/L [65]. Additionally, Hv can produce significant amounts of floral and fruity acetate esters, benzenoids, and terpenes, improving the aroma profile of flat neutral varieties [63,64,66,67]. Furthermore, Hv is better adapted to the fermentation process and it is possible to select strains capable of reaching 10% ethanol [68]. The use of other Hanseniaspora species such as H. opuntiae fermenting Cabernet Sauvignon red grapes has also shown low volatile acidity values together with positive fruity and floral profiles [69,70].
5. Biocompatibility
The use of non-Saccharomyces in ternary cultures (two non-Saccharomyces species and one Saccharomyces species) in sequential or mixed fermentations has several advantages in terms of aroma improvement, control of spoilage microorganisms, and depletion of off-flavors; however, it is very important to ensure the biocompatibility of the strains used. When Lt has been used in co-inoculation with Hv, Td, and Mp, the latter has shown very good compatibility with Lt, reaching even higher levels of acidification than using only Lt in sequential fermentation with Sc [59]. However, the simultaneous use of Lt and Td decreased acidification, and the pH was higher compared to Lt alone, but lower than in the Sc control. The Lt and Hv strains showed the worst effectiveness on pH reduction, despite our high expectations of the complementary effect of both yeasts on acidity and aroma. This may be caused by the extra consumption of thiamine and pantothenate by Hv and the potential depletion of these important micronutrients, particularly of thiamine. The genes for thiamine biosynthesis in Hv and other Hanseniaspora species have not yet been identified, and this may explain the increased requirements of this vitamin in Hv fermentations [68,71,72]. Thiamine consumption and depletion may affect the development of other non-Saccharomyces species when used in co-fermentation.
6. Depletion of Off-Flavor Precursors
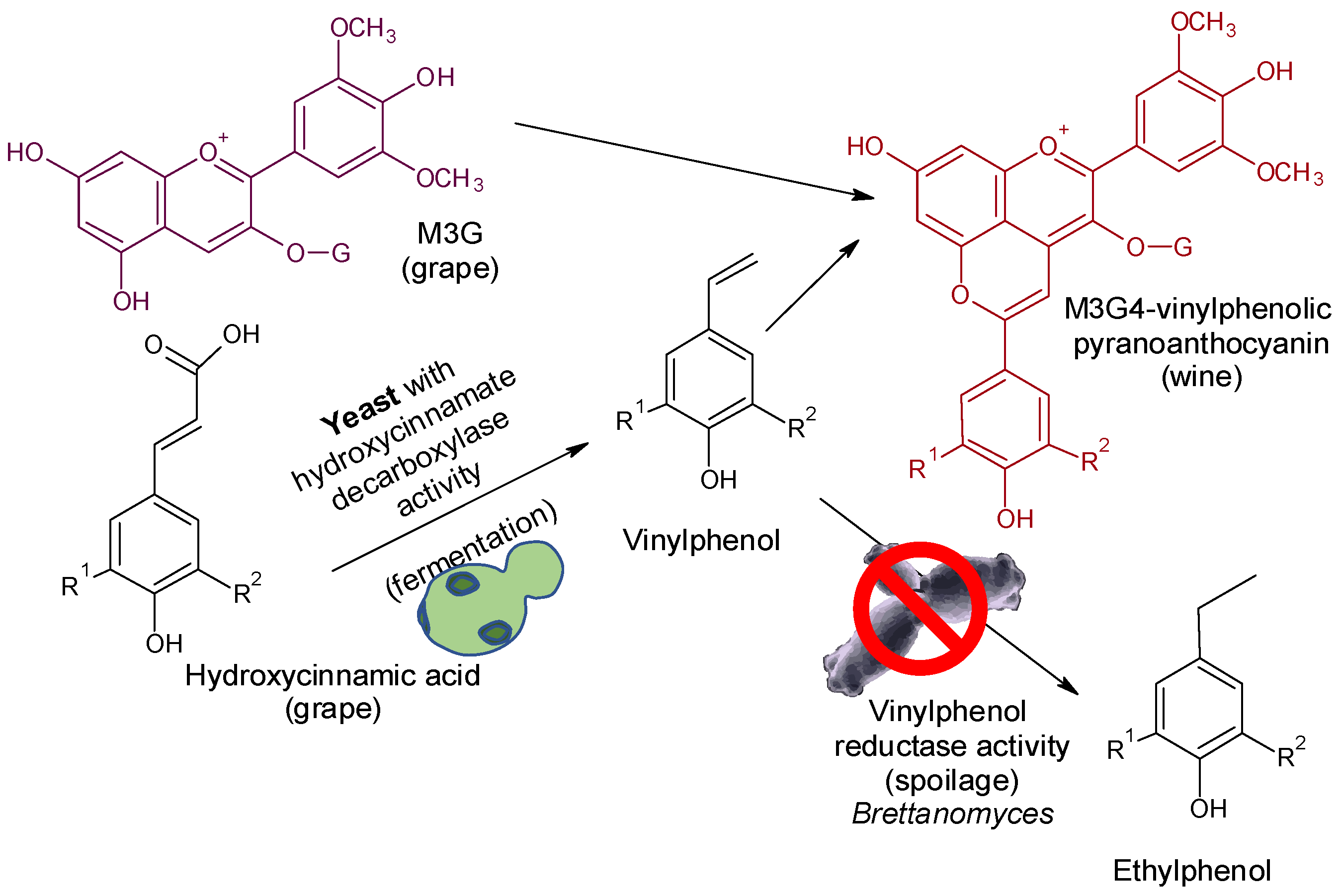
The production of some off-flavors that are extremely deleterious to wine quality, such as ethylphenols (EPs) [4,5], is highly dependent on precursor content. EPs are formed from hydroxycinnamic acids (HCAs) or their tartaric esters (TE-HCAs) by the sequential activities of hydroxycinnamate decarboxylase (HCDC) and vinylphenol reductase (VPR) from Brettanomyces/Dekkera [4,10]. Several technologies have been proposed to control Brettanomyces in wines, including emerging non-thermal technologies, some additives, and biotechnologies [4,73]. Many Sc strains express HCDC activity, but VPR activity has not been described in this species. Some Sc strains express HCDC activity with high intensity with the ability to transform most hydroxycinnamic acids into vinylphenols (VPs) (Figure 3). Moreover, it has been observed that these VPs can spontaneously react with grape anthocyanins to form vinylphenolic pyranoanthocyanins (VPAs) [74,75], which are stable pigments under enological conditions, as they are less affected by pH, oxidations, and sulfur dioxide bleaching than grape anthocyanins [61,76,77,78,79]. The use of Sc with an appropriate expression of HCDC activity is a powerful and natural biotool to favor the enzymatic metabolization of HCAs to VPs and the subsequent reaction with grape anthocyanins to form VPAs. This biological process blocks the EP precursors into stable VPAs, which are positive in terms of color stability, but also preserves the wines from the effect of Brettanomyces/Dekkera. When 10 commercial yeasts with verified HCDC activity were used to ferment red musts and subsequently contaminated with Brettanomyces, the 4EP content ranged from 22 to 498 µg/L, which is below or close to the sensory threshold of 4EP in wines [26]. However, in the control yeast (without HCDC activity), the 4EP content was 1150 µg/L, more than twice the sensory threshold [26]. Furthermore, most of the HCAs in grapes are found as tartaric esters (TE-HCAs); that is, caftaric, coutaric, and fertaric acids are reservoirs of HCAs that can be released by acid hydrolysis during aging. The use of cinnamyl esterase enzymes during fermentation can release the HCAs which, using Sc with HCDC activity, can be transformed into VPs and subsequently into VPAs by condensation with grape anthocyanins [26]. In addition, the use of some non-Saccharomyces yeast strains such as Torulaspora delbrueckii or Metschnikowia pulcherrima can enhance the formation of VPAs [60,80].
7. Increasing the Implantation of Non-Saccharomyces as Bioprotective Tools Using Emerging Non-Thermal Technologies
To achieve a good effectiveness with non-Saccharomyces yeasts in off-flavor control through bioprotection, acidification, and improved sulfur dioxide efficiency or precursor depletion, it is necessary to reach a good implantation of the intended species. One of the main drawbacks of non-Saccharomyces yeasts is the low fermentative power (<10% vol. and in many species <4% vol.) and the low fermentative yield, which generally results in poor implantation compared to Sc. To improve this, the use of non-thermal emerging technologies is compelling and effective because they can effectively eliminate wild yeasts, but also have little impact on the sensory components of the musts, that is, the aroma, pigments, and flavors [73,81,82,83].
The emerging non-thermal technologies include pressurization technologies (high hydrostatic pressure, HHP [84], and ultra-high-pressure homogenization, UHPH [85]), pulsed electric fields (PEFs) [86], pulsed light (PL) [87], irradiation (βI) [88], cold plasma (CD) [89], and ultrasound (US) [90]. All of them except US have demonstrated a good capacity to inactivate wild yeasts and even bacteria in grapes and musts, and preserve sensory and nutritional quality. HHP can produce reductions in wild yeast populations of more than 4-log [91,92], but residual bacterial counts can remain. UHPH is capable of producing sterilization with the elimination of yeast, bacteria, and even spores, depending on the in-valve temperature [85,93]. Pulsed technologies (i.e., PEFs and PL) have shown an inactivation capacity around or above 2-log for wild yeasts in grapes [94,95,96]. The antimicrobial performance of PEFs can be greatly enhanced in combination with mild temperatures (50 °C) [97].
The inactivation of wild yeasts by emerging non-thermal technologies in grapes or grape must is a useful technology to facilitate the implantation of non-Saccharomyces starters that can be used to control off-flavor formation. Several non-thermal technologies have shown high efficiency in increasing the implantation of non-Saccharomyces yeasts, such as HHP [92] and PEFs [95]. The high effectiveness of UHPH also makes it a leading technology not only for improving yeast implantation, but also for reducing SO2 levels due to its ability to inactivate oxidative enzymes [85].
In addition, when used on grapes, several of these non-thermal technologies are able to increase the extraction of phenolic compounds, thus improving the tannin and anthocyanin content of the wine. An increase in anthocyanin extraction ranging from 23% to 63% by HHP [98,99], 21% to 29% by PEFs [100], and the same contents but with reductions of more than 50% in maceration time by US [101] have been published.
8. Conclusions
The use of non-Saccharomyces in wine fermentation is a verified biotechnology to improve the sensory profile, and is also a powerful biotool to control off-flavor formation by the biocontrol of spoilage microorganisms, by pH control and the improvement of molecular SO2 contents by acidification, and by the depletion of precursors, among many other potential future possibilities.
Author Contributions
Conceptualization, A.M.; writing—original draft preparation, A.M. and I.L.; writing—review and editing, A.M., I.L., C.G. and C.E.; funding acquisition, A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Ministerio de Ciencia, Innovación y Universidades, project: RTI2018-096626-B-I00.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The review study did not report any data.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds ... are available from the authors.
References
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef]
- Ugliano, M.; Kolouchova, R.; Henschke, P.A. Occurrence of hydrogen sulfide in wine and in fermentation: Influence of yeast strain and supplementation of yeast available nitrogen. J. Ind. Microbiol. Biotechnol. 2011, 38, 423–429. [Google Scholar] [CrossRef]
- Bely, M.; Rinaldi, A.; Dubourdieu, D. Influence of assimilable nitrogen on volatile acidity production by Saccharomyces cerevisiae during high sugar fermentation. J. Biosci. Bioeng. 2003, 96, 507–512. [Google Scholar] [CrossRef]
- Suárez, R.; Suárez-Lepe, J.A.; Morata, A.; Calderón, F. The production of ethylphenols in wine by yeasts of the genera Brettanomyces and Dekkera: A review. Food Chem. 2007, 102, 10–21. [Google Scholar] [CrossRef]
- Malfeito-Ferreira, M. Two decades of “Horse Sweat” taint and Brettanomyces yeasts in wine: Where do we stand now? Beverages 2018, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.Q.; Pilone, G.J. An overview of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar] [CrossRef]
- Culleré, L.; López, R.; Ferreira, V. Chapter 20—The instrumental analysis of aroma-active compounds for explaining the flavor of red wines. In Red Wine Technology; Morata, A., Ed.; Elsevier, Academic Press: Amsterdam, The Netherlands, 2018; pp. 283–307. ISBN 9780128144008. [Google Scholar]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Alcohols and other volatile compounds. In Handbook of Enology: The Chemistry of Wine Stabilization and Treatments (Volume 2); John Wiley & Sons: Chichester, UK, 2006; pp. 51–64. [Google Scholar]
- OIV International Code of Oenological Practice. Available online: https://www.oiv.int/public/medias/3741/e-code-annex-maximum-acceptable-limits.pdf (accessed on 15 June 2021).
- Chatonnet, P.; Dubourdie, D.; Boidron, J.-n.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Curtin, C.; Bramley, B.; Cowey, G.; Holdstock, M.; Kennedy, E.; Lattey, K.; Coulter, A.; Henschke, P.; Francis, L.; Godden, P. Sensory perceptions of ‘Brett’ and relationship to consumer preference. In Proceedings of the Thirteenth Australian Wine Industry Technical Conference, Adelaide, Australia, 29 July–2 August 2007; Blair, R.J., Williams, P.J., Pretorius, I.S., Eds.; Australian Wine Industry Technical Conference Inc.: Adelaide, Australia, 2007; pp. 207–211. [Google Scholar]
- Mira de Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Sadras, V.O.; Petrie, P.R.; Moran, M.A. Effects of elevated temperature in grapevine. II juice pH, titratable acidity and wine sensory attributes. Aust. J. Grape Wine Res. 2013, 19, 107–115. [Google Scholar] [CrossRef]
- Mozell, M.R.; Thachn, L. The impact of climate change on the global wine industry: Challenges & solutions. Wine Econ. Policy 2014, 3, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Mpelasoka, B.S.; Schachtman, D.P.; Treeby, M.T.; Thomas, M.R. A review of potassium nutrition in grapevines with special emphasis on berry accumulation. Aust. J. Grape Wine Res. 2003, 9, 154–168. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Coetzee, Z.A.; Walker, R.R.; Deloire, A.; Tyerman, S.D. Potassium in the grape (Vitis vinifera L.) berry: Transport and function. Front. Plant Sci. 2017, 8, 1629. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, C.; Spano, G.; Fragasso, M.; Grieco, F.; Russo, P.; Capozzi, V. Starter cultures as biocontrol strategy to prevent Brettanomyces bruxellensis proliferation in wine. Appl. Microbiol. Biotechnol. 2018, 102, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces commercial starter cultures: Scientific trends, recent patents and innovation in the wine sector. Recent Pat. Food. Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-Protection as an alternative to sulphites: Impact on chemical and microbial characteristics of red wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Banilas, G.; Sgouros, G.; Nisiotou, A. Development of microsatellite markers for Lachancea thermotolerans typing and population structure of wine-associated isolates. Microbiol. Res. 2016, 193, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic yeasts: A promising alternative to chemical fungicides for controlling postharvest decay of fruit. J. Fungi 2020, 6, 158. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Morata, A.; Vejarano, R.; Ridolfi, G.; Benito, S.; Palomero, F.; Uthurry, C.; Tesfaye, W.; González, C.; Suárez-Lepe, J.A. Reduction of 4-ethylphenol production in red wines using HCDC+ yeasts and cinnamyl esterases. Enzym. Microb. Technol. 2013, 52, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Chassagne, D.; Guilloux-Benatier, M.; Alexandre, H.; Voilley, A. Sorption of wine volatile phenols by yeast lees. Food Chem. 2005, 91, 39–44. [Google Scholar] [CrossRef]
- Palomero, F.; Ntanos, K.; Morata, A.; Benito, S.; Suárez-Lepe, J.A. Reduction of wine 4-ethylphenol concentration using lyophilised yeast as a bioadsorbent: Influence on anthocyanin content and chromatic variables. Eur. Food Res. Technol. 2011, 232, 971–977. [Google Scholar] [CrossRef]
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [Green Version]
- Escott, C.; Loira, I.; Morata, A.; Bañuelos, M.A.; Suárez-Lepe, J.A. Wine spoilage yeasts: Control strategy. In Yeast—Industrial Applications; Morata, A., Loira, I., Eds.; InTechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Nardi, T. Microbial resources as a tool for enhancing sustainability in winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata, A.; Bañuelos, M.A.; López, C.; Song, C.; Vejarano, R.; Loira, I.; Palomero, F.; Suarez Lepe, J.A. Use of fumaric acid to control pH and inhibit malolactic fermentation in wines. Food Addit. Contam.—Part A Chem. Anal. Control. Expo. Risk Assess. 2020, 37, 228–238. [Google Scholar] [CrossRef]
- Woods, D.R.; Bevan, E.A. Studies on the nature of the killer factor produced by Saccharomyces cerevisiae. J. Gen. Microbiol. 1968, 51, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Young, T.W.; Yagiu, M. A comparison of the killer character in different yeasts and its classification. Antonie Leeuwenhoek 1978, 44, 59–77. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Ciani, M. The zymocidial activity of Tetrapisispora phaffii in the control of Hanseniaspora uvarum during the early stages of winemaking. Lett. Appl. Microbiol. 2010, 50, 50–56. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Rodriguez, L.; Joseph, C.M.L.; Nazaris, B.; Coulon, J.; Richardson, S.; Dycus, D.A. Innovative use of non-Saccharomyces in bio-protection: T. delbrueckii and M. pulcherrima applied to a machine harvester. Catal. Discov. Pract. 2020, 4, 82–90. [Google Scholar] [CrossRef]
- Pelonnier-Magimel, E.; Windholtz, S.; Masneuf-Pomarède, I.; Barbe, J.C. Sensory characterisation of wines without added sulfites via specific and adapted sensory profile. Oeno One 2020, 54, 671–685. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Rubio-Bretón, P.; Gonzalo-Diago, A.; Iribarren, M.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. Bioprotection as a tool to free additives winemaking: Effect on sensorial, anthocyanic and aromatic profile of young red wines. LWT 2018, 98, 458–464. [Google Scholar] [CrossRef]
- Windholtz, S.; Redon, P.; Lacampagne, S.; Farris, L.; Lytra, G.; Cameleyre, M.; Barbe, J.C.; Coulon, J.; Thibon, J.; Masneuf-Pomarède, I. Non-Saccharomyces yeasts as bioprotection in the composition of red wine and in the reduction of sulfur dioxide. LWT 2021, 149, 111781. [Google Scholar] [CrossRef]
- Johnson, J.; Fu, M.; Qian, M.; Curtin, C.; Osborne, J.P. Influence of select non-Saccharomyces yeast on Hanseniaspora uvarum growth during prefermentation cold maceration. Am. J. Enol. Vitic. 2020, 71, 278–287. [Google Scholar] [CrossRef]
- Comuzzo, P.; Battistutta, F. Chapter 2—Acidification and pH Control in Red Wines. In Red Wine Technology; Morata, A., Ed.; Elsevier, Academic Press: Amsterdam, The Netherlands, 2018; pp. 17–34. ISBN 9780128144008. [Google Scholar]
- Yéramian, N.; Chaya, C.; Suárez Lepe, J.A. L-(-)-malic acid production by Saccharomyces spp. during the alcoholic fermentation of wine (1). J. Agric. Food Chem. 2007, 55, 912–919. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; Del Fresno, J.M.; González, C.; Suárez-lepe, J.A. Contribution of non-Saccharomyces yeasts to wine freshness. A review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Bañuelos, M.A.; Vaquero, C.; Loira, I.; Cuerda, R.; Palomero, F.; González, C.; Suárez-Lepe, J.A.; Wang, J.; Han, S.; et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur. Food Res. Technol. 2019, 245, 885–894. [Google Scholar] [CrossRef]
- Vaquero, C.; Loira, I.; Bañuelos, M.A.; Heras, J.M.; Cuerda, R.; Morata, A. Industrial performance of several Lachancea thermotolerans strains for ph control in white wines from warm areas. Microorganisms 2020, 8, 830. [Google Scholar] [CrossRef]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.P.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on chemical composition and sensory profiles of Merlot wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef] [PubMed]
- Gatto, V.; Binati, R.L.; Lemos Junior, W.J.F.; Basile, A.; Treu, L.; de Almeida, O.G.G.; Innocente, G.; Campanaro, S.; Torriani, S. New insights into the variability of lactic acid production in Lachancea thermotolerans at the phenotypic and genomic level. Microbiol. Res. 2020, 238, 126525. [Google Scholar] [CrossRef] [PubMed]
- Castro Marín, A.; Colangelo, D.; Lambri, M.; Riponi, C.; Chinnici, F. Relevance and perspectives of the use of chitosan in winemaking: A review. Crit. Rev. Food Sci. Nutr. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- OIV. Treatment with Fumaric Acid in Wine to Inhibit MLF. Resolution OENO-TECHNO 15-581. 2020. Available online: https://www.oiv.int/en/technical-standards-and-documents/resolutions-of-the-oiv/oenology-resolutions (accessed on 28 July 2021).
- Xu, G.; Chen, X.; Liu, L.; Jiang, L. Fumaric acid production in Saccharomyces cerevisiae by simultaneous use of oxidative and reductive routes. Bioresour. Technol. 2013, 148, 91–96. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Escott, C.; Morata, A.; Ricardo-Da-Silva, J.M.; Callejo, M.J.; Del Carmen González, M.; Suarez-Lepe, J.A. Effect of Lachancea thermotolerans on the formation of polymeric pigments during sequential fermentation with Schizosaccharosmyces pombe and Saccharomyces cerevisiae. Molecules 2018, 23, 2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero, C.; Loira, I.; Heras, J.M.; Carrau, F.; González, C.; Morata, A. Biocompatibility in ternary fermentations with Lachancea thermotolerans, other non-Saccharomyces and Saccharomyces cerevisiae to control pH and improve the sensory profile of wines from warm areas. Front. Microbiol. 2021, 12, 656262. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Tesfaye, W.; del Carmen González, M.; Suárez-Lepe, J.A. Formation of polymeric pigments in red wines through sequential fermentation of flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018, 239, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Escott, C.; Loira, I.; Manuel Del Fresno, J.; González, C.; Suárez-Lepe, J.A. Influence of Saccharomyces and non-Saccharomyces yeasts in the formation of pyranoanthocyanins and polymeric pigments during red wine making. Molecules 2019, 24, 4490. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Valera, M.J.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Del Fresno, J.M.; Escott, C.; Loira, I.; Herbert-Pucheta, J.E.; Schneider, R.; Carrau, F.; Cuerda, R.; Morata, A. Impact of Hanseniaspora vineae in alcoholic fermentation and ageing on lees of high-quality white wine. Fermentation 2020, 6, 66. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, D.; Duan, C.; Yan, G. Synergistic effect enhances 2-phenylethyl acetate production in the mixed fermentation of Hanseniaspora vineae and Saccharomyces cerevisiae. Process Biochem. 2020, 90, 44–49. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Escott, C.; Loira, I.; Carrau, F.; Cuerda, R.; Schneider, R.; Bañuelos, M.A.; González, C.; Suárez-Lepe, J.A.; Morata, A. The impact of Hanseniaspora vineae fermentation and ageing on lees on the terpenic aromatic profile of white wines of the albillo variety. Int. J. Mol. Sci. 2021, 22, 2195. [Google Scholar] [CrossRef]
- Valera, M.J.; Boido, E.; Dellacassa, E.; Carrau, F. Comparison of the glycolytic and alcoholic fermentation pathways of Hanseniaspora vineae with Saccharomyces cerevisiae wine yeasts. Fermentation 2020, 6, 78. [Google Scholar] [CrossRef]
- Luan, Y.; Zhang, B.Q.; Duan, C.Q.; Yan, G.L. Effects of different pre-fermentation cold maceration time on aroma compounds of Saccharomyces cerevisiae co-fermentation with Hanseniaspora opuntiae or Pichia kudriavzevii. LWT Food Sci. Technol. 2018, 92, 177–186. [Google Scholar] [CrossRef]
- Feng, C.T.; Du, X.; Wee, J. Microbial and chemical analysis of non-Saccharomyces yeasts from Chambourcin hybrid grapes for potential use in winemaking. Fermentation 2021, 7, 15. [Google Scholar] [CrossRef]
- Seixas, I.; Barbosa, C.; Mendes-Faia, A.; Güldener, U.; Tenreiro, R.; Mendes-Ferreira, A.; Mira, N.P. Genome sequence of the non-conventional wine yeast Hanseniaspora guilliermondii UTAD222 unveils relevant traits of this species and of the Hanseniaspora genus in the context of wine fermentation. DNA Res. 2019, 26, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Čadež, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLoS Biol. 2019, 17, e3000255. [Google Scholar] [CrossRef] [Green Version]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging technologies to control Brettanomyces spp. in wine: Recent advances and future trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Calderón, F.; Suárez, J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006, 106, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; González, C.; Suárez-Lepe, J.A. Formation of vinylphenolic pyranoanthocyanins by selected yeasts fermenting red grape musts supplemented with hydroxycinnamic acids. Int. J. Food Microbiol. 2007, 116, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.; Timberlake, C.F. Isolation, identification and characterization of new color-stable anthocyanins occurring in some red wines. J. Agric. Food Chem. 1997, 45, 35–43. [Google Scholar] [CrossRef]
- De Freitas, V.; Mateus, N. Formation of pyranoanthocyanins in red wines: A new and diverse class of anthocyanin derivatives. Anal. Bioanal. Chem. 2011, 401, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Suárez Lepe, J.A. Influence of yeasts in wine colour. In Grape and Wine Biotechnology; InTech: Rijeka, Croatia, 2016. [Google Scholar] [CrossRef] [Green Version]
- Escribano-Bailón, M.T.; Rivas-Gonzalo, J.C.; García-Estévez, I. Wine color evolution and stability. In Red Wine Technology; Elsevier, Academic Press: Amsterdam, The Netherlands, 2018; pp. 195–205. ISBN 9780128144008. [Google Scholar] [CrossRef]
- Escott, C.; Morata, A.; Loira, I.; Tesfaye, W.; Suarez-Lepe, J.A. Characterization of polymeric pigments and pyranoanthocyanins formed in microfermentations of non-Saccharomyces yeasts. J. Appl. Microbiol. 2016, 121, 1346–1356. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Vejarano, R.; González, C.; Callejo, M.J.; Suárez-Lepe, J.A. Emerging preservation technologies in grapes for winemaking. Trends Food Sci. Technol. 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Paniagua-Martínez, I.; Ramírez-Martínez, A.; Serment-Moreno, V.; Rodrigues, S.; Ozuna, C. Non-thermal technologies as alternative methods for Saccharomyces cerevisiae inactivation in liquid media: A review. Food Bioprocess Technol. 2018, 11, 487–510. [Google Scholar] [CrossRef]
- Gómez-López, V.M.; Pataro, G.; Tiwari, B.; Gozzi, M.; Meireles, M.Á.A.; Wang, S.; Guamis, B.; Pan, Z.; Ramaswamy, H.; Sastry, S.; et al. Guidelines on reporting treatment conditions for emerging technologies in food processing. Crit. Rev. Food Sci. Nutr. 2021, 1–25. [Google Scholar] [CrossRef] [PubMed]
- San Martín, M.F.; Barbosa-Cánovas, G.V.; Swanson, B.G. Food processing by high hydrostatic pressure. Crit. Rev. Food Sci. Nutr. 2002, 42, 627–645. [Google Scholar] [CrossRef]
- Morata, A.; Guamis, B. Use of UHPH to obtain juices with better nutritional quality and healthier wines with low levels of SO2. Front. Nutr. 2020, 7, 598286. [Google Scholar] [CrossRef] [PubMed]
- Raso, J.; Calderón, M.L.; Góngora, M.; Barbosa-Cánovas, G.V.; Swanson, B.G. Inactivation of Zygosaccharomyces bailii in fruit juices by heat, high hydrostatic pressure and pulsed electric fields. J. Food Sci. 1998, 63, 1042–1044. [Google Scholar] [CrossRef]
- Santamera, A.; Escott, C.; Loira, I.; Del Fresno, J.M.; González, C.; Morata, A. Pulsed light: Challenges of a non-thermal sanitation technology in the winemaking industry. Beverages 2020, 6, 45. [Google Scholar] [CrossRef]
- Morata, A.; Bañuelos, M.A.; Tesfaye, W.; Loira, I.; Palomero, F.; Benito, S.; Callejo, M.J.; Villa, A.; González, M.C.; Suárez-Lepe, J.A. Electron beam irradiation of wine grapes: Effect on microbial populations, phenol extraction and wine quality. Food Bioprocess Technol. 2015, 8, 1845–1853. [Google Scholar] [CrossRef]
- Sainz-García, E.; López-Alfaro, I.; Múgica-Vidal, R.; López, R.; Escribano-Viana, R.; Portu, J.; Alba-Elías, F.; González-Arenzana, L. Effect of the atmospheric pressure cold plasma treatment on Tempranillo red wine quality in batch and flow systems. Beverages 2019, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- García Martín, J.F.; Sun, D.W. Ultrasound and electric fields as novel techniques for assisting the wine ageing process: The state-of-the-art research. Trends Food Sci. Technol. 2013, 33, 40–53. [Google Scholar] [CrossRef]
- Delfini, C.; Conterno, L.; Carpi, G.; Rovere, P.; Tabusso, A.; Cocito, C.; Amati, A. Microbiological stabilisation of grape musts and wines by High Hydrostatic Pressures. J. Wine Res. 1995, 6, 143–151. [Google Scholar] [CrossRef]
- Bañuelos, M.A.; Loira, I.; Escott, C.; Del Fresno, J.M.; Morata, A.; Sanz, P.D.; Otero, L.; Suárez-Lepe, J.A. Grape processing by High Hydrostatic Pressure: Effect on use of non-Saccharomyces in must fermentation. Food Bioprocess Technol. 2016, 9, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Bañuelos, M.A.; Loira, I.; Guamis, B.; Escott, C.; Del Fresno, J.M.; Codina-Torrella, I.; Quevedo, J.M.; Gervilla, R.; Chavarría, J.M.R.; de Lamo, S.; et al. White wine processing by UHPH without SO2. Elimination of microbial populations and effect in oxidative enzymes, colloidal stability and sensory quality. Food Chem. 2020, 332, 127417. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Arias-Gil, M.; Marsellés-Fontanet, A.R.; Ancín-Azpilicueta, C.; Martín-Belloso, O. Effects of thermal and non-thermal processing treatments on fatty acids and free amino acids of grape juice. Food Control 2007, 18, 473–479. [Google Scholar] [CrossRef]
- Vaquero, C.; Loira, I.; Raso, J.; Álvarez, I.; Delso, C.; Morata, A. Pulsed Electric Fields to improve the use of non-Saccharomyces starters in red wines. Foods 2021, 10, 1472. [Google Scholar] [CrossRef] [PubMed]
- Escott, C.; López, C.; Loira, I.; González, C.; Bañuelos, M.A.; Tesfaye, W.; Suárez-Lepe, J.A.; Morata, A. Improvement of must fermentation from late harvest cv. Tempranillo grapes treated with pulsed light. Foods 2021, 10, 1416. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Mittal, G.S.; Griffiths, M.W. Effect of pulsed electric field on the inactivation of microorganisms in grape juices with and without antimicrobials. Biosyst. Eng. 2005, 90, 1–7. [Google Scholar] [CrossRef]
- Corrales, M.; García, A.F.; Butz, P.; Tauscher, B. Extraction of anthocyanins from grape skins assisted by high hydrostatic pressure. J. Food Eng. 2009, 90, 415–421. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Vejarano, R.; Bañuelos, M.A.; Sanz, P.D.; Otero, L.; Suárez-Lepe, J.A. Grape processing by High Hydrostatic Pressure: Effect on microbial populations, phenol extraction and wine quality. Food Bioprocess Technol. 2015, 8, 277–286. [Google Scholar] [CrossRef] [Green Version]
- López, N.; Puértolas, E.; Condón, S.; Álvarez, I.; Raso, J. Effects of pulsed electric fields on the extraction of phenolic compounds during the fermentation of must of Tempranillo grapes. Innov. Food Sci. Emerg. Technol. 2008, 9, 477–482. [Google Scholar] [CrossRef]
- Pérez-Porras, P.; Bautista-Ortín, A.B.; Jurado, R.; Gómez-Plaza, E. Using high-power ultrasounds in red winemaking: Effect of operating conditions on wine physico-chemical and chromatic characteristics. LWT 2021, 138, 110645. [Google Scholar] [CrossRef]
Figure 1.
Summary of the mechanisms of biocontrol of off-flavor formation by yeasts. M3G: malvidin-3-O-glucoside; M3G4-vinylphenolic: malvidin-3-O-glucoside-4-vinylphenolic.
Figure 1.
Summary of the mechanisms of biocontrol of off-flavor formation by yeasts. M3G: malvidin-3-O-glucoside; M3G4-vinylphenolic: malvidin-3-O-glucoside-4-vinylphenolic.

Figure 2.
Sequential fermentations with Lachancea thermotolerans (green yeasts—bottom left) and Saccharomyces cerevisiae (pink yeasts—center). Effect on lactic acid production (green line), alcohol content (blue line), pH, and molecular SO2.
Figure 2.
Sequential fermentations with Lachancea thermotolerans (green yeasts—bottom left) and Saccharomyces cerevisiae (pink yeasts—center). Effect on lactic acid production (green line), alcohol content (blue line), pH, and molecular SO2.
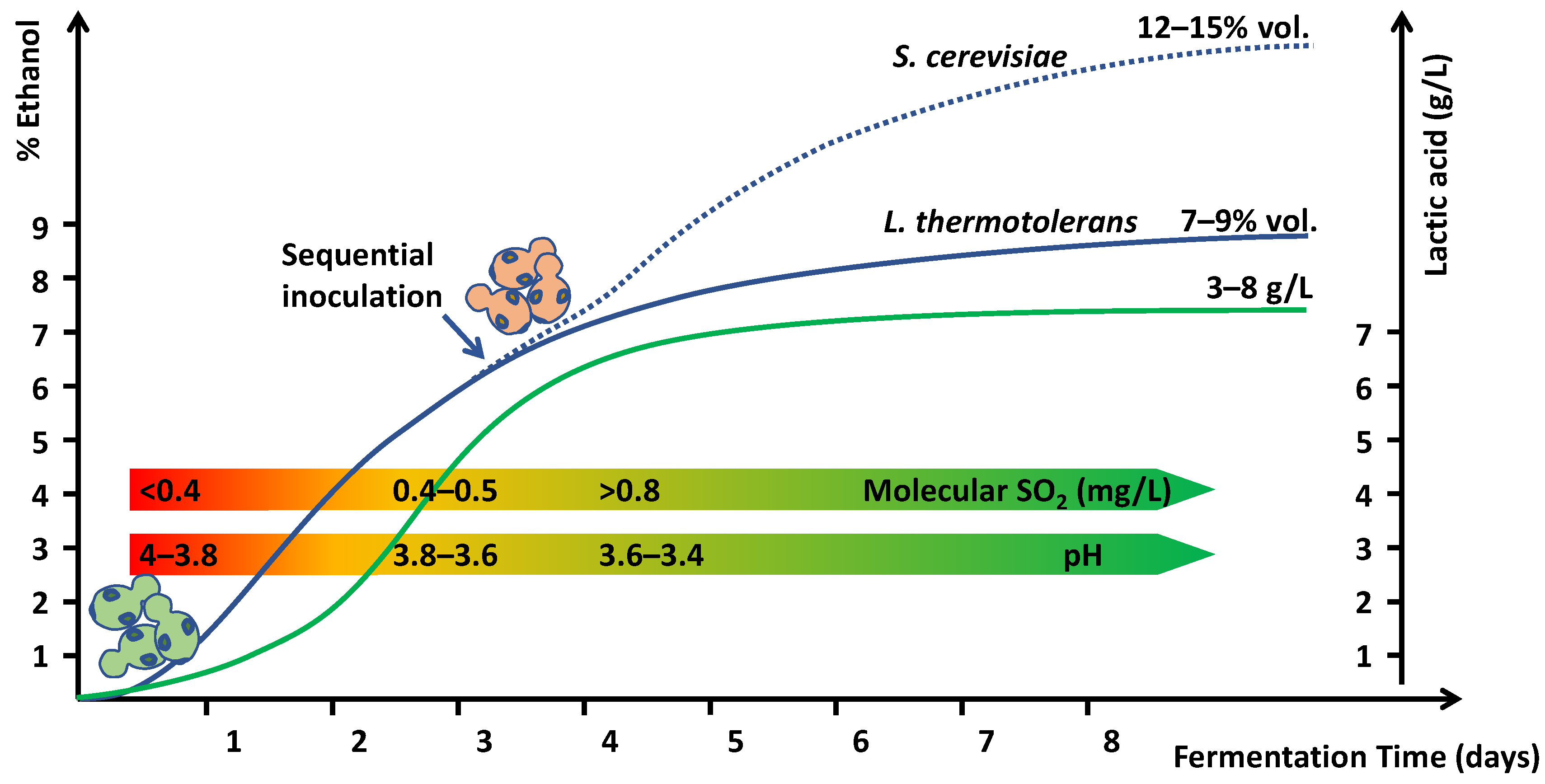
Figure 3.
The depletion of ethylphenol precursors by the metabolization of hydroxycinnamic acids to vinylphenols and the blocking of this by reaction with grape anthocyanins to form vinylphenolic pyranoanthocyanins.
Figure 3.
The depletion of ethylphenol precursors by the metabolization of hydroxycinnamic acids to vinylphenols and the blocking of this by reaction with grape anthocyanins to form vinylphenolic pyranoanthocyanins.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Wine off-flavors produced by microbial metabolites.
| Compound | Sensory Threshold | Off-Flavor Concentration 1 | Descriptor | Reference |
|---|---|---|---|---|
| H2S | 1.6 μg/L | >1.6 μg/L | Rotten eggs/putrefaction | [2] |
| Volatile acidity | 0.3–0.6 g/L | >0.8 g/L legal limit 1.2 g/L | Vinegar | [7,8,9] |
| Ethyl acetate | 12 mg/L | >150 mg/L | Glue, solvent | [7] |
| 4-Ethylphenol | 230 μg/L | >425 μg/L | Phenolic, stable, leather, horse sweat | [5,10,11] |
| Acetaldehyde | 100–125 mg/L | >125 mg/L | Fruity, rotten apples, nut-like, sherry | [6] |
1 The sensory threshold and off-flavor perception can be variable depending on the structure, composition, and sensory buffering effect of the wine.
Table 2.
Bioacidification by Lachancea thermotolerans strain L31 and effect on pH and molecular SO2 in several fermentations with red and white varieties in different Spanish regions. Colors indicate the effectiveness of molecular SO2 from unideal (red) or less than optimal (yellow) to optimal (green) depending on the increase in acidity.
Table 2.
Bioacidification by Lachancea thermotolerans strain L31 and effect on pH and molecular SO2 in several fermentations with red and white varieties in different Spanish regions. Colors indicate the effectiveness of molecular SO2 from unideal (red) or less than optimal (yellow) to optimal (green) depending on the increase in acidity.
| Variety (Region) | Inoculation | Lactic Acid (g/L) and Initial→Final pH | Effect of Acidity on the Molecular SO2 (mg/L) * | Reference |
|---|---|---|---|---|
| Tempranillo (Ribera del Duero) | Sequential with S. cerevisiae | 0.91→6.60 g/L 3.90→3.63 | 0.42→0.77 | [50] |
| Tempranillo (Ribera del Duero) | Mixed with O. oeni and sequential with S. cerevisiae | 0.91→7.50 g/L 3.90→3.31 | 0.42→1.56 | [50] |
| Tempranillo (Mancha) | Sequential with S. cerevisiae | 3.8→3.4 | 0.50→1.22 | Unpublished |
| Albariño (Rias Baixas-O Rosal) | Sequential with S. cerevisiae | 0.05→2.7 g/L 3.12→2.85 | 2.07→3.63 | [50] |
| Airén (La Mancha) | Sequential with S. cerevisiae | 0.05→4.20 g/L 3.75→3.35 | 0.51→1.25 | [51] |
* Comparison for a total content of SO2 of 50 mg/L.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Morata, A.; Loira, I.; González, C.; Escott, C. Non-Saccharomyces as Biotools to Control the Production of Off-Flavors in Wines. Molecules 2021, 26, 4571. https://doi.org/10.3390/molecules26154571
AMA Style
Morata A, Loira I, González C, Escott C. Non-Saccharomyces as Biotools to Control the Production of Off-Flavors in Wines. Molecules. 2021; 26(15):4571. https://doi.org/10.3390/molecules26154571
Chicago/Turabian StyleMorata, Antonio, Iris Loira, Carmen González, and Carlos Escott. 2021. "Non-Saccharomyces as Biotools to Control the Production of Off-Flavors in Wines" Molecules 26, no. 15: 4571. https://doi.org/10.3390/molecules26154571