
Microbiological Study in Petrol-Spiked Soil

Department of Soil Science and Microbiology, University of Warmia and Mazury in Olsztyn, 10-727 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(9), 2664; https://doi.org/10.3390/molecules26092664
Submission received: 9 April 2021
/
Revised: 27 April 2021
/
Accepted: 30 April 2021
/
Published: 1 May 2021
(This article belongs to the Topic Extreme Environments: Microbial and Biochemical Diversity)
Abstract
:The pollution of arable lands and water with petroleum-derived products is still a valid problem, mainly due the extensive works aimed to improve their production technology to reduce fuel consumption and protect engines. An example of the upgraded fuels is the BP 98 unleaded petrol with Active technology. A pot experiment was carried out in which Eutric Cambisol soil was polluted with petrol to determine its effect on the microbiological and biochemical properties of this soil. Analyses were carried out to determine soil microbiome composition—with the incubation and metagenomic methods, the activity of seven enzymes, and cocksfoot effect on hydrocarbon degradation. The following indices were determined: colony development index (CD); ecophysiological diversity index (EP); index of cocksfoot effect on soil microorganisms and enzymes (IFG); index of petrol effect on soil microorganisms and enzymes (IFP); index of the resistance of microorganisms, enzymes, and cocksfoot to soil pollution with petrol (RS); Shannon–Weaver’s index of bacterial taxa diversity (H); and Shannon–Weaver’s index of hydrocarbon degradation (IDH). The soil pollution with petrol was found to increase population numbers of bacteria and fungi, and Protebacteria phylum abundance as well as to decrease the abundance of Actinobacteria and Acidobacteria phyla. The cultivation of cocksfoot on the petrol-polluted soil had an especially beneficial effect mainly on the bacteria belonging to the Ramlibacter, Pseudoxanthomonas, Mycoplana, and Sphingobium genera. The least susceptible to the soil pollution with petrol and cocksfoot cultivation were the bacteria of the following genera: Kaistobacter, Rhodoplanes, Bacillus, Streptomyces, Paenibacillus, Phenylobacterium, and Terracoccus. Cocksfoot proved effective in the phytoremediation of petrol-polluted soil, as it accelerated hydrocarbon degradation and increased the genetic diversity of bacteria. It additionally enhanced the activities of soil enzymes.
1. Introduction
The following four elements are the main determinants of human and animal health: soil fertility, water quality, air purity, and technologies employed in the agri-food processing [1,2,3]. Each of these elements is crucial to the sustainable development of populations, with soil playing a significant role in this chain due to its quality being the main driver of the dietary value of agricultural crops produced [4,5]. The physical and chemical devastation of soil, including both the area-related one affected by the widely understood industrialization, and the point-related one associated with incidental events, urges for the continuous upgrade of various reclamation technologies [6,7,8]. An increasingly important role is ascribed in this regard to microorganisms, as evidenced by the bioaugmentation-aided reclamation methods [9,10,11,12].
The soil microbiome is highly diversified [13,14]. According to Nesme [15], one gram of soil can provide a habitat for over 10,000 various bacterial species. However, the assessment of soil ecosystem functioning is usually based on the physical soil indicators related to its texture, aggregation, porosity, and humidity [16,17,18,19], as well as the chemical ones, including the contents of organic carbon [20,21], total nitrogen [22], available phosphorus and potassium [23,24], and also soil pH value [25,26]. The methods used for soil quality assessment usually take no account of the microbiological processes ongoing in the soil, such as e.g., ISO standards [27]. Furthermore, Bünemann et al. [28] have underlined the underestimation of biological indicators in the soil functioning evaluation. It is an extremely valuable observation considering the meaningful role the soil microorganisms play in the structure and functioning of ecosystems [29]. The life on Earth would not be possible without them [28,30]. The communities of rhizospheric microorganisms are involved in, i.e., soil structure formation [18] and organic matter degradation [31,32], as well as increase nutrient availability and plant productivity [33,34]. Root secretions are both perfect substrates and signalling molecules for microorganisms, establishing complex interactions between roots, soil, and microorganisms [35,36,37]. This complex community of microorganisms associated with plants is also called the second plant genome. Plants can influence the development of their rhizospheric microbiome [38]. Hence, the soil microorganisms are of key importance to plant health and to the biogeochemical cycles of biogenic elements [20,22,32,39,40].
Both the soil microorganisms and the enzymes they produce are sensitive to varying environmental conditions [41,42,43,44]. They faithfully reproduce the condition of the soil environment; therefore, the microbiological and biochemical indicators cannot be neglected in soil quality evaluation [6,13,30,45].
The rapid development of molecular methods observed in the last decade has caused the appearance of an increasing number of indicators based on genotypic and phenotypic diversity, next to the microbiological ones determined with conventional methods [46,47]. They allow for the immediate assessment of the composition and functioning of a microbial community at various trophic levels [14,48]. Thus, the molecular methods complement the conventional microbiological and biochemical indicators of soil quality [49]. Nevertheless, the molecular techniques are also burdened with some errors resulting from sample contamination, primer selection, or taxonomic classification techniques [15,50]. Moreover, a large proportion of soil organisms has not yet been characterized in terms of their taxonomy and functions [30,46]. Therefore, there has long been a debate about the best choice of bioindicators that can objectively be used to monitor soil quality and thus provide early warning of a potential loss of its multifunctionality [30]. It is important to apply a holistic approach to soil quality assessment, taking into account different ecosystems. However, this assessment may be distorted by the specific, often contradictory, response of microorganisms to heavy metals [51,52] and organic pollutants [6,12,53,54,55].
Many functions performed by soil microorganisms are currently under threat due to the degradation of soil ecosystems by petroleum-derived products, which is a global problem [56]. The stability of microorganisms is disturbed in such an environment [41,57]. Soil pollution with petroleum-derived products induces stress in microbial communities that require long adaptation periods [43,58]. According to Borowik et al. [59], diesel oil triggered greater changes in the soil microbiome than unleaded petrol. Soil pollution with diesel oil reduced the counts of all bacterial taxa except for species, while soil pollution with petrol decreased the bacterial diversity only at the class, order, and family level. The composition of soil microbiota in the soil polluted with petrol hydrocarbons evolves [53,60]. The soils exposed to the pressure of PAHs are mainly predominated by representatives of β-, γ-Proteobacteria, Actinobacteria, and Bacteroidia [61,62,63]. As Tejeda-Agredano et al. [64] claim, the prevailing genera of the soils polluted with petroleum-derived products include Sphingomonas, Commamonas, Oxalobacteria, and Xhanthomonas, whereas according to Kumar et al. [65] these are the representatives of Alcanivorax and Aequorivita. Soil pollution with petroleum-derived products not only causes changes in the microbial diversity but also, through the food chain, contributes to the induction of chronic diseases of immunosuppressed men and animals. These products pose a significant threat to the environment and ecological safety of the global population [4], thereby reducing the agricultural productivity of soils [5]. Therefore, restoring the soil biological homeostasis is a priority to maintain the social stability and sustainable development [66,67], and to ensure food safety [3,53]. This restoration will be feasible owing to the more comprehensive understanding of the interactions of soil microorganisms under various stress conditions, which will additionally enable predicting responses of the soil microbial communities and activities of enzymes in the environment exposed to the pressure of petroleum-based products. The above premises have prompted a research aimed to determine the impact of soil pollution with petrol on its microbiome and enzymatic activity, on the response of cocksfoot to the soil pollution, and the role of this plant in degradation of petrol hydrocarbons.
2. Results
2.1. Microbiological Properties of Soil
Cocksfoot was found to exert a beneficial effect on organotrophic bacteria (Org), actinobacteria (Act), and fungi (Fun), as indicated by the positive values of the IFG index, ranging from 0.162 (Act) to 0.629 (Table S1, Figure 1a). Intermediate values of IFG were determined for Org. The pollution of non-sown soil and soil sown with cocksfoot with petrol significantly promoted the proliferation of all tested groups of microorganisms, as evidenced by positive IFP values. Petrol had a stronger effect on the organotrophic bacteria and actinobacteria in the soil planted with cocksfoot, whereas its impact on fungi in this soil was weaker. These effects were reflected in the values of the index of microorganism resistance (RS) to petrol (Figure 1a–c).
Cocksfoot cultivation in the soil tested contributed to a decrease in the colony development (CD) index of organotrophic bacteria and actinobacteria, and to an increase in the CD index of fungi. The CD index values were similarly affected by soil pollution with petrol. In the non-sown soil, petrol caused an increase in the ecophysiological diversity (EP) index of organotrophic bacteria and fungi, whereas in the soil sown with cocksfoot, it decreased EP index values of all microorganisms tested (Figure 2a,b).
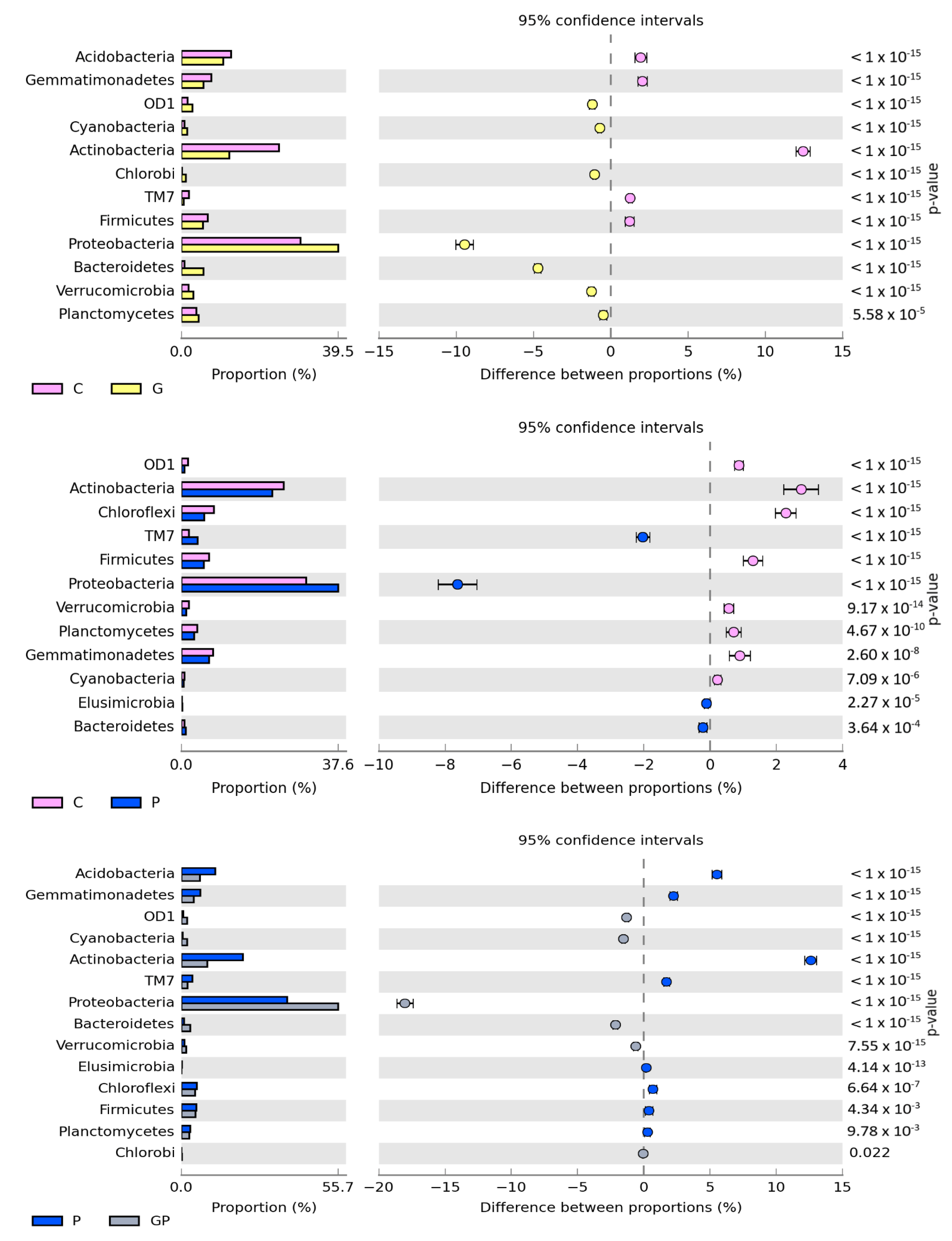
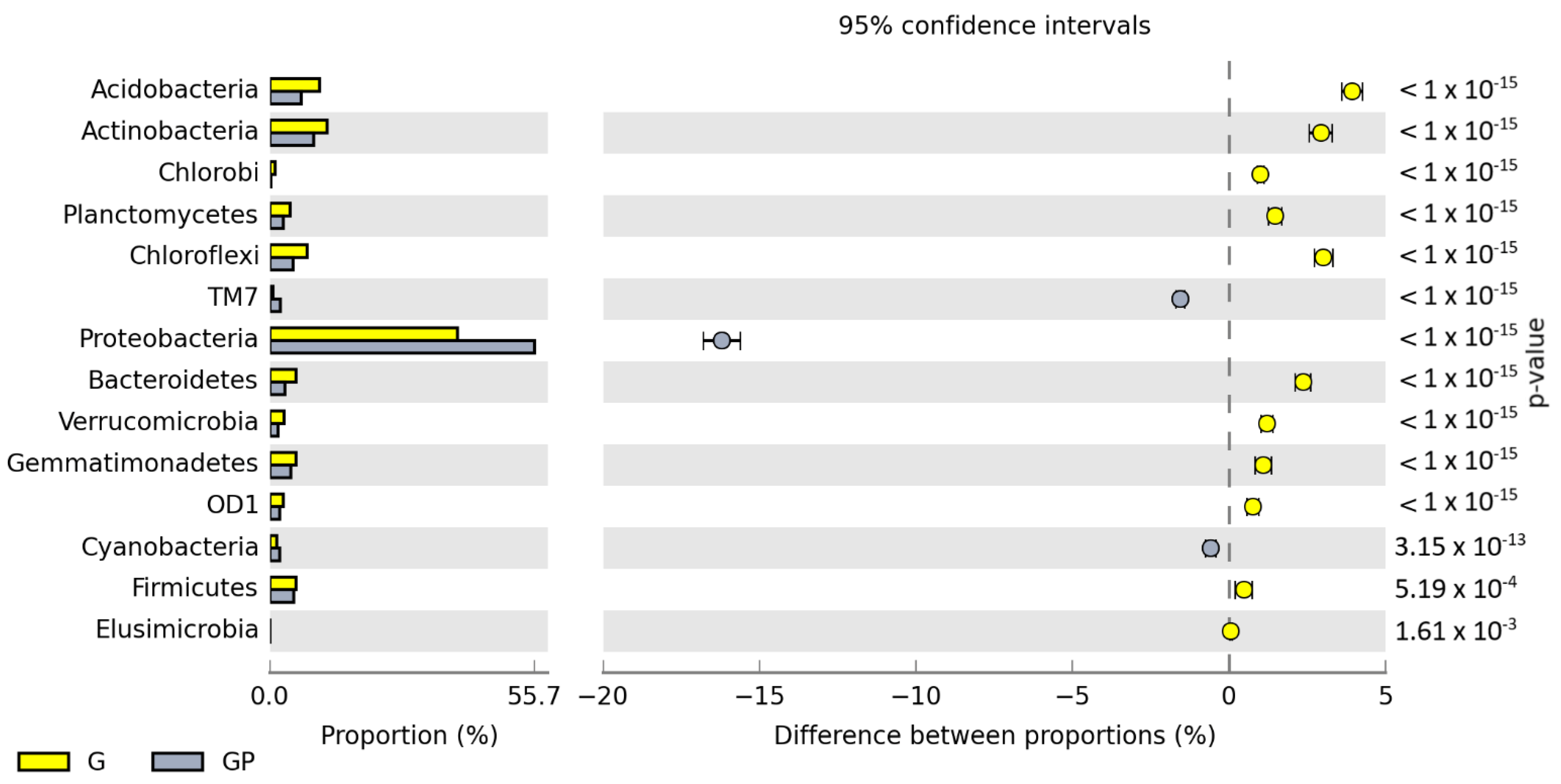
The number of isolated OTUs reached 56,726 in the non-sown and non-polluted soil and 49,640 in the non-sown and petrol-polluted soil. After cocksfoot sowing, OTU numbers reached 59,104 and 54,338 in the non-polluted and polluted soil. Regardless of soil cultivation type and pollution with petrol, the prevailing phyla were: Proteobacteria, Actinobacteria, and Acidobacteria (Figure 3). The pollution of non-sown soil increased the OTU number of Proteobacteria by 7.67% and that of TM7 by 2.03%, but decreased OTU numbers of Actinobacteria and Chloroflexi by 2.73% and 2.28%, respectively. In turn, cocksfoot sowing increased the OUT number of Proteobacteria by 16.18% and decreased the OUT number of Acidobacteria by 3.95%, that of Chloroflexi by 3.02%, that of Actinobacteria by 2.94%, and that of Bacteroidetes by 2.37%, compared to the soil sown with cocksfoot but not polluted with petrol. The greatest differences in OTU numbers of the prevailing phyla were found when comparing the non-sown petrol-polluted soil with the soil sown with cocksfoot but not polluted with petrol (P vs. GP). Cocksfoot sowing in the petrol-polluted soil increased OTU numbers of Proteobacteria by 17.97% and Bacteroidetes by 2.14%, whereas it decreased those of Actinobacteria by 12.66%, Acidobacteria by 5.54%, and Gemmatimonadetes by 2.27%.
The bacterial structure was observed to change successively depending on soil cultivation and pollution with petrol. In total, 42 genera were identified in the soil samples that were represented by at least 1% of total assigned sequences (Figure 4). Twenty of them (Arenimonas, Burkholderia, Dechloromonas, Devosia, Gallionella, Geobacter, HB2-32-21, Hyphomicrobium, Kaistobacter, Lysobacter, Methylibium, Methylotenera, Mycoplana, Phenylobacterium, Pseudomonas, Pseudoxanthomonas, Ramlibacter, Rhodanobacter, Rhodoplanes, and Sphingobium) were classified to the phylum Proteobacteria, seven of them (Arthrobacter, Iamia, Nocardioides, Pseudonocardia, Rhodococcus, Streptomyces, and Terracoccus) to Actinobacteria, five (Alicyclobacillus, Bacillus, Clostridium, Paenibacillus, and Planifilum) to Firmicutes, three (Flavobacterium, Flavisolibacter, and Sporocytophaga) to Bacteroidetes, two (DA101 and Opitutus) to Verrucomicrobia, two (Candidatus Koribacter and Candidatus Solibacter) to Acidobacteria, two (Gemmata and Planctomyces) to Planctomycetes, and one (Fimbriimonas) to Armatimonadetes. In the non-sown soil polluted and not polluted with petrol, the most abundant genus turned out to be Kaistobacter, which accounted for 17.55% and 21.74% of all assigned sequences, respectively. In turn, HB2-32-21 turned out to be the most abundant genus in the soil sown with cocksfoot, accounting for 8.59% and for as much as 31.40% of total OTUs in the non-polluted and polluted soils, respectively. The structure of individual identified genera varied depending on soil cultivation type and its pollution with petrol. The pollution of non-sown soil was found to increase the relative abundance of Kaistobacter, Burkholderia, and Phenylobacterium, whereas the pollution of soil sown with cocksfoot—to increase the relative abundance of HB2-32-21, Devosia, Bacillus, Pseudomonas, Arenimonas, and Paenibacillus.
The Venn diagram (Figure 5a), depicting the number of all bacterial genera identified in particular pots, demonstrated that as many as 121 genera were common for all soil samples, regardless of soil cultivation and pollution. The highest number of specific bacterial genera was identified in the petrol-polluted soil sown with cocksfoot (19 genera), a slightly lower one in the soil sown with cocksfoot but non-polluted (11 genera), and the lowest one in the non-sown and non-polluted soil (1 genus) and in the non-sown but petrol-polluted soil (3 genera). After excluding bacterial genera with OTU lower than 1% of the assigned sequences, it can be undoubtedly concluded that Kaistobacter, Rhodococcus, Bacillus, Streptomyces, Paenibacillus, Phenylobacterium, and Terracoccus were the endogenous genera, hence common for oil soil types. In turn, Arthrobacter, Rhodanobacter, and Rhodococcus were found typical of the now-sown soil polluted with petrol, whereas Ramlibacter, Iamia, Mycoplana, Pseudoxanthomonas, and Sphingobium for the petrol-polluted soil sown with cocksfoot (Figure 5b).
Taking into account the types of bacterial genera with OTU higher than 1% of the assigned sequences, bacteria belonging to 10 genera were identified to the species (Figure 6). Although the Proteobacteria z bacteria were found to prevail, the highest number of sequences assigned to the species was identified for the phylum Firmicutes bacteria, which represented a common microbiome for all analyzed objects, both the non-polluted and the petrol-polluted ones.
2.2. Biochemical Properties of Soil
In the non-polluted soil (Table S2, Figure 8a) cocksfoot stimulated activities of dehydrogenases (Deh), catalase (Cat), urease (Ure), alkaline phosphatase (Pal), and arylsulfatase (Aryl), but inhibited activities of acid phosphatase (Pac), and β-glucosidase (Glu). In the petrol-polluted soil, it enhanced activities of most enzymes, but usually to a significantly lesser extent than in the petrol-polluted soil. The impact of petrol on the enzymatic activity was significantly lower compared to cocksfoot. In the non-sown soil, negative values of the petrol effect index were noted for all enzymes except for dehydrogenases and β-glucosidase, whereas in the cocksfoot-sown soil, the negative IFP values were noted for Deh, Ure, Pac, and Aryl, and the positive ones for Cat and Glu (Figure 8b). The values of the resistance index (RS) showed that Pal was the most stable enzyme in the petrol-polluted soil that was sown and non-sown with cocksfoot, followed by Cat and Glue, and that Deh were the least stable (Figure 8c).
2.3. Degradation of Petrol
Cocksfoot significantly accelerated degradation of the following hydrocarbons in soil: mineral oils (C12–C35), benzene (Ben), naphthalene (Nap), anthracene (Ant), chrysene (Chr), benzo[a]anthracene (BaA), benzo(a)pyrene (BaP), benzo[b]fluoranthene (BbF), and benzo(k)fluoranthene (BkF) (Table 1). The transformation of gasoline fractions (C6–C12), ethylbenzene (EtB), toluene (Tol), and xylene (Xyl) in the study period reached almost 100% and did not depend on soil cultivation type. In turn, the degradation of such hydrocarbons as Ant, Chr, BaA, BaP, BbF, BkF, and IP was much slower and, except for IP, was intensified by cocksfoot cultivation. Furthermore, cocksfoot itself was susceptible to the pollution with petrol; however, its resistance to the pollution increased with time. The resistance index (RS) of the first regrowth reached 0.154, that of the second regrowth reached 0.295, and that of the third regrowth reached 0.652 (Figure 9).
3. Discussion
3.1. Response of Microorganisms and Enzymes to Soil Pollution with Petrol
The soil microorganisms quite faithfully respond to ongoing changes in the soil; hence, they can be used as highly sensitive biosensors [30]. They influence the trends and magnitude of changes, leading to a specific homeostasis reached in ecosystems [68]. Environmental pollution can modify the microbiological succession and change its direction, debilitating ecosystem’s resistance [21,66]. In the present study, soil pollution with petrol significantly promoted the proliferation of all tested groups of microorganisms, as evidenced by positive IFP values. In the soil non-sown with cocksfoot, petrol increased the count of bacteria from the Proteobacteria and TM7 phyla, and reduced counts of those from Actinobacteria and Chloroflexi phyla. Furthermore, Galitskaya et al. [69] demonstrated that 12% soil pollution with petroleum for 120 days contributed to the succession of bacterial communities. The polluted soil was predominated by Actinobacteria (from 35% to 58%), Proteobacteria (from 25% to 30%), and TM7 (from 15% to 35%).
In our experiment, the structure of communities has changed as well, i.e., the contribution of k strategist bacteria (slowly-growing ones) and r strategist fungi (rapidly growing ones) increased. The changes triggered by petrol in the structure of soil bacteria communities stem from the different resistance of individual species to its toxic effect [12,43,66,70] and from the diversified possibilities of utilizing its hydrocarbons as sources of carbon, hydrogen, and energy [71].
Our study indicated that, regardless of the stress triggered by soil pollution with petrol, the stability of the diversity of bacterial communities was highly reliably described by the autochthonous microbiome being common for the non-polluted and petrol-polluted soils, represented by Kaistobacter, Rhodococcus, Bacillus, Streptomyces, Paenibacillus, Phenylobacterium, and Terracoccus. It is the bacteria of these genera that are active in the degradation of petrol products, which can be effective in the bio-augmentation of polluted areas. Galitskaya et al. [69] demonstrated that the prevailing OTUs in the soil exposed to petrol belonged to the Rhodococcus and Mycobacterium genera, whereas according to de la Cueva et al. [66], the most abundant OTUs were these of: Acinetobacter, Pedomicrobium, Halomonas, Rhizobium, Cryobacterium, Pseudomonas, Lysobacter, Thermomonas, and Stenotrophomonas. Thus, even the soils strongly polluted with petroleum-based products reveal a potential for bio-remediation [58,72].
Because it is feasible to only cultivate a fraction of soil microbiota [15], it seems essential to determine the biological activity of soil by assaying its enzymatic activity [73,74]. The soil enzymes are perceived as reliable soil quality indicators [17,75,76]. They are used to construct models for the assessment of potential soil fertility [28,77,78,79,80]. Due to their ability to signalize changes triggered under the pressure of stress factors, they also perfectly mirror the effects of pollutants on soil [81,82,83]. Among the soil enzymes, dehydrogenases prove best in reflecting the rate of changes in the soil environment. They indicate levels of physiologically-active microorganisms in the soil, the oxidation rate of organic matter, and the availability of nutrients in the soil [71,76]. Therefore, in the present study, the petrol pollution of soil not-sown with cocksfoot caused an over 100% increase in the activity of dehydrogenases. Although more rarely used to determine pollution’s effect on the soil ecosystem’s stability, also such enzymes as catalase, β-glucosidase, urease, phosphatases, and arylsulfatase can provide valuable information on the metabolic capability of soil [73,74,83]. Their indicatory potential is due to the strong association of their activities with biogeochemical cycles of C, N, P, and S [71,75]. Our study has emphasized the significant effect of cocksfoot and petrol on C, N, and P metabolism in the soil, as evidenced by the changes in the population numbers and diversity of microorganisms as well as in activities of the soil enzymes tested. The present study proved alkaline phosphatase followed by catalase and β-glucosidase to be relatively the most stable enzymes, while dehydrogenases to be the least stable ones in the petrol-polluted soil.
3.2. The Role of Cocksfoot in Restoring the Biological Homeostasis of Petrol-Polluted Soil
The diversity of microorganisms and the activity of enzymes in the soil affect the functioning of ecosystems and, hence, the growth and health of plants [68]. For this reason, the petrol-induced disorders in the soil microbiome could deteriorate the physical properties of soil [35,36,37], leading to the poorer development of the root system and aerial parts of cocksfoot. Growth and development of the plants were hampered despite soil fertilization with nitrogen, phosphorus, potassium, and magnesium in doses meeting the cocksfoot demands for nutrients.
The present study proved that the negative response of cocksfoot to petrol diminished over time, which proves the progressing microbiological degradation of petrol hydrocarbons with time. Camacho-Montealegre et al. [42] demonstrated that, under favourable conditions, from 66% to 75% of hydrocarbons contained in petroleum products can be mineralized within 60 days depending on the species used for phytoremediation. In our study, cocksfoot proved effective in petrol degradation, and this effect was mainly due to its stimulating impact on soil microorganisms. The microbiological degradation of petroleum hydrocarbons in the soil is a natural process, owing to which organic pollutants can be mineralized by autochthonous microorganisms [41,84]. The cultivation of cocksfoot enhanced their activities as a result of the stimulating effect of root secretions on their development [42,60].
Petroleum hydrocarbons exhibit various susceptibilities to the attacks of microorganisms. The rate of their degradation decreases along with the elongation of the hydrocarbon chain [58,69,85]. Furthermore, in the present study, a higher degradation rate was observed for di-, tri-, and four-ring PAHs, followed by the five- and six-ring ones. According to Ite and Ibok [53], the effectiveness of biodegradability of oil components is as follows: n-alkanes > branched alkenes > low molecular weight n-alkyl aromatics > monoaromatics > cyclic alkanes > polycyclic aromatic hydrocarbons > asphaltene. Degradation of hydrocarbons in the soil is strongly associated with root secretions that ensure the rhizodegradation of organic pollutants [85,86,87].
Cocksfoot significantly intensified the degradation of all PAHs, except for IP, because plant roots improve the physical [35,36,37] and chemical [75] properties of soil, thereby creating more favourable conditions for the proliferation of microorganisms, including the hydrocarbon-degrading ones [87]. These abilities of plants are particularly important in the soils polluted with petroleum products [41,85], especially that plant roots create a specific habitat for rhizospheric microbial communities, which can be used for rhizoremediation [42,60]. The cultivation of cocksfoot on the petrol-polluted soil had an especially beneficial effect on the bacteria belonging to the phylum Proteobacteria, mainly those from the genus Ramlibacter and the class β-Proteobacteria, those from the genus Pseudoxanthomonas and the class γ-Proteobacteria, as well as those from Mycoplana and Sphingobium genera from the class α-Proteobacteria. Cocksfoot increased the bacterial diversity at each taxonomic level.
The results of our study demonstrate that petrol disturbed the stability of a soil ecosystem and confirm the hypothesis that cocksfoot is utile in restoring the biological homeostasis of soil contaminated with this pollutant. These findings suggest the need for continuous search, development, and implementation of bioremediation strategies for petrol-polluted soils.
4. Materials and Methods
4.1. Petrol
The study was conducted with petrol available at BP fuel stations [88] under a commercial name BP 98 unleaded petrol with Active technology. Apart from volatile hydrocarbons, it contains paraffins, naphtenes, olephins, and aromatic compounds. It is enriched with dirt-capturing substances forming a protective layer of the engine. Petrol characteristics is available at BP’s website [89].
4.2. Field Studies
The first stage of the study involved soil selection and evaluation of its properties (Table S3). Soils of arable lands with a little degree of anthropogenic transformations were in the focus of interest. Top quality soils with natural contents of elements and organic compounds were found in the north-eastern Poland. The soil to be used in the greenhouse experiment was collected from an arable field located near the city of Olsztyn (53.7167° N, 20.4167° E). This was Eutric Cambisol type soil with the following composition: 2.22% of the loam fraction, 22.85% of the dust fraction, and 74.93% of the sand fraction. Its samples were collected from the topsoil (0–20 cm). Its cation exchange capacity was 60.40 mmol (+) kg−1 d.m. soil, and it had pHKCl 6.7. The analysed soil contained 9.3 g Corg kg−1 d.m. soil, 0.62 g N kg−1 d.m. soil, 93.68 mg available P kg−1 d.m. soil, 141.10 mg available K kg−1 d.m. soil, and 42 mg available Mg kg−1 d.m. soil. It had a very low content of PAHs, in mg kg−1 d.m. soil: gasoline fractions (C6–C12)—0.8; mineral oil (C12–C35)—6; benzene (Ben)—0.01; ethylbenzene (EtB)—0.01; toluene (Tol)—0.01; xylene (X)—0.03; naphthalene (Nap)—0.005; anthracene (Ant)—0.005; chrysene (Chr)—0.013; benzo[a]anthracene (BaA)—0.009; benzo(a)pyrene (BaP)—0.011; benzo[b]fluoranthene (BbF)—0.009; benzo(k)fluoranthene (BkF)—0.008; and indeno(1.2.3-cd)pyrene (IP)—0.006.
4.3. Greenhouse Experiment
Samples of soil (Eutric Cambisol) were transported to a greenhouse, thoroughly mixed, and sieved through a screen with a mesh diameter of 5 mm. The experiment was conducted in 7 dm3 Kick-Brauckman polyethylene pots, filled with 9 kg of soil. Four independent variables were tested in four replications, i.e., non-sown soil (C), non-sown soil polluted with petrol (P), soil sown with cocksfoot (G), and soil sown with cocksfoot and polluted with petrol (GP). Cocksfoot (Dactylis glomerata) was selected for this study as it easily adapts to unfavourable environmental conditions [90,91,92]. It is a fast-growing, loose-cluster species with a well-developed bundle root system, common to Europe, Asia, and North Africa. It is resistant to low temperatures, drought, and diseases [93,94]. It is also an energy grass, with biomass production reaching from 11 to 13 Mg d.m. per 1 ha [95]. The additional advantage of this grass species, speaking for its use in the phytoremediation of petrol-polluted soil, is its high resistance to the pollution with polycyclic aromatic hydrocarbons [96]. The same mineral fertilization was applied in each experimental series, in mg kg−1 d.m. soil: N—80, P—20, K—40, and Mg—10. Nitrogen was used in the form of urea, phosphorus in the form of potassium dihydrophosphate, while potassium as potassium dihydrophosphate and potassium chloride, and magnesium as magnesium sulfate heptahydrate. In P and GP pots, the soil was polluted with BP 98 unleaded petrol with Active technology in a dose of 7 cm3 kg−1 d.m. soil. The mineral fertilizer and petrol were applied in a single dose before pot filling with the soil, after their separate mixing with each soil portion (9 kg) intended for one pot. Afterward, soil moisture content was brought to 60% of the water capillary capacity using distilled water. One week after the experiment had been established, 24 seeds of cocksfoot (Dactylis glomerata) were sown in pots G and GP. The experiment was continued for 105 days. On day 45, 75, and 105, cocksfoot was cut, and its dry matter yield was determined. The soil moisture content was kept stable throughout the experiment at 60% of the water capillary capacity. During the last cut, soil samples were collected for microbiological and biochemical analyses. The samples were sifted through a screen with mesh diameter of 2 mm.
4.4. Microbiological Analyses
4.4.1. Determination of Population Numbers of Microorganisms
A serial dilution method was used to determine population numbers of organotrophic bacteria, actinobacteria, and fungi in the soil samples. To this end, serial dilutions from 10−2 to 10−6 were prepared from the samples of non-polluted and petrol-polluted soil. One cm3 of a respective soil dilution (10−5 and 10−6 for organotrophic bacteria and actinobacteria, and 10−3 and 10−4 for fungi) was measured onto Petri dishes. Then, 18 cm3 of a selective medium (organotrophic bacteria—Bunt and Rovira medium, actinomycetes—Parkinson medium, and fungi—Martin medium) were added onto the dishes. The medium’s composition was presented in our earlier work [97]. Incubation was performed at a temperature of 28 °C for 10 days. The colony forming units (cfu) of microorganisms were counted every 24 h for 10 consecutive days with a colony counter. The microbiological analyses were performed in 4 replications.
4.4.2. DNA Isolation
Genomic DNA was isolated from the samples of non-polluted and petrol-polluted soil using a Genomic Minix X Soil kit, following producer’s guidelines. To check isolated DNA quality and its bacterial origin, a Real-Time PCR was conducted in the Mx3000P thermocycler (Stratagene) using the SYBR Green dye as a fluorochrome. Two universal primers were used, i.e., 1055F and 1392R.
4.4.3. 16S rRNA Gene Amplicon Sequencing
Bacterial communities colonizing the non-polluted and petrol-polluted soils were analysed by sequencing the hypervariable region V3–V4 of the 16S rRNA gene. The mass sequencing of 16S rRNA amplicons were performed by Genomed SA, Poland, following the Illumina protocol. A fragment of the 16S rRNA gene was amplified with PCR primers recommended for the Illumina technique, i.e., 341F and 785R. DNA was sequenced on the Illumina MiSeq apparatus in the paired-end 2 × 250 protocol, using the Miseq Reagent Kit v2 (Illumina, San Diego, CA, USA).
4.4.4. Bioinformatic Analysis
The quality of the obtained sequences was controlled, and incomplete or chimeric ones were rejected. The identified sequences were grouped using the uclust algorithm. The taxonomic identification was performed with the QIIME package using a database of reference sequences GreenGenes v13_8. The sequencing data were deposited in the GenBank NCBI [98] under accession numbers of MW266821-MW266860; MW380430-MW380501; MW579372-MW579410; MW686950-MW687007.
4.4.5. Biochemical, Chemical, and Physicochemical Analyses of Soil
Activities of the following soil enzymes were determined in the study: dehydrogenases (Deh), catalase (Cat), urease (Ure), alkaline phosphatase (Pal), acid phosphatase (Pac), arylsulfatase (Aryl), and β-glucosidase (Glu). All determinations were carried out with standard methods described in our previous works [97] in three replications, under controlled conditions The enzymatic activity was expressed in product units per 1 kg d.m. soil per 1 h. The activity of Deh was expressed in μmol TFF (triphenyl formazane), that of Cat in mol O2, that of Ure in mmol N-NH4+, and these of Pal, Pac, Aryl, and Glu in mmol PN (4-nitrophenol).
In addition, the soil samples were determined for the contents of: gasoline fractions (C6–C12), mineral oils (C12–C35), volatile aromatic hydrocarbons (BETX), naphthalene (Nap), anthracene (Ant), chrysene (Chr), benzo(a)antracene (BaA), benzo(a)pyrene (BaP), benzo(b)fluoranthene (BbF), benzo(k)fluoranthene (BkF), and indeno(123-cd)pyrene (IP), following procedures provided in the following standards: EN ISO 18287 [99], EN ISO 16703 [100], and EN ISO 22155 [101]. The above determinations were conducted using an Agilent 7890A-5975C mas spectrometer equipped in EI/CI ion source.
The soils samples were also analysed for their granulometric composition; pH value; contents of total nitrogen, organic carbon, available P, K, and Mg; exchangeable cations Ca2+, Mg2+, K+, and Na+, and ion exchange capacity. All determinations were carried out with standard methods described in our previous work [70].
4.4.6. Calculations and Statistical Analysis
The following indices were determined to establish cocksfoot and petrol effects on soil condition:s
(1) colony development index (CD) of organotrophic bacteria, actinobacteria, and fungi according to Leij et al. [102]:
where: N1, N2, N3,...N10—sum of the quotients of colony numbers of microorganisms identified in particular days of the study (1, 2, 3,...10) and the sum of all colonies in the entire study period;
CD = [N1/1 + N2/2 + N3/3….. N10/10] · 100,
(2) ecophysiological diversity index (EP) of organotrophic bacteria, actinobacteria, and fungi according to Leij et al. [102]:
where: pi—the quotient of the number of colonies of microorganisms from particular days of the study and the sum of all colonies from the entire study period;
EP = −Σ(π·log10 π),
(3) index of cocksfoot effect (IFG) on soil microorganisms and enzymes:
where: AG—the number of microorganisms or the activity of enzymes in the soil sown with cocksfoot, A0—the number of microorganisms or the activity of enzymes in the non-sown soil;
(4) index of petrol effect (IFP) on soil microorganisms and enzymes:
where: Ap—the number of microorganisms or the activity of enzymes in the soil polluted with petrol, A0—the number of microorganisms or the activity of enzymes in the non-polluted soil;
(5) resistance (RS) of microorganisms, enzymes, and cocksfoot to soil pollution with petrol [103]:
where: D0 = C0—P0, C0—parameter value determined for the control soil, P0—parameter value determined for the petrol-polluted soil;
(6) index of hydrocarbon degradation (IDH):
where AH0—the content of hydrocarbons in the petrol-polluted soil on the day of experiment establishment, AH—the content of hydrocarbons on the day of experiment termination.
(7) Shannon–Weaver index (H) determining bacterial diversity:
where: p is the ratio of OTU numbers of one representative of the tested taxon to the total OTU number of the entire taxon.
H = −Σpi(lnpi),
The normality of data distribution was verified with the Kruskal–Wallis and Shapiro–Wilk tests. The post-hoc Duncan test was used for further analyses. One-way significance tests (Table S2) were performed using the analysis of variance (ANOVA). Statistical calculations were performed with the Statistica 13.3 package [104]. The results of the metagenomic analysis were elaborated and interpreted using: STAMP 2.1.3. software [105], RStudio v1.2.5033 software [106], R v3.6.2 system [107], gplots libraries [108], and cluster analysis [109]. The STAMP 2.1.3. software was used to present significant differences between sequence proportions for the phylum in the analysed soil samples, at a confidence interval of 95%. The diversity of bacteria at the genus level was presented using the heatmap with a dendrogram of their similarities, generated using RStudio v1.2.5033 software [106], R v3.6.2 system [107] and gplots library [108] as well as a Venn diagram plotted using InteractiVenn [109].
5. Conclusions
The microbiological and biochemical homeostasis was disturbed in the soil polluted with petrol. The pollution resulted in increased counts of bacteria and fungi as well as enhanced enzymatic activity. It also caused the increased contribution of k strategist bacteria and r strategist fungi in the structure of respective communities. Soil sowing with cocksfoot increased the genetic diversity of bacteria and accelerated the degradation of petrol hydrocarbons, which predisposes this plant for phytoremediation purposes. Proteobacteria, Actinobacteria, and Acidobacteria were the prevailing phyla in the soil tested. Its pollution with petrol and sowing with cocksfoot increased the abundance of Proteobacteria and decreased the abundance of Actinobacteria and Acidobacteria. The ones least susceptible to the soil pollution with petrol and cocksfoot cultivation were the bacteria of the following genera: Kaistobacter, Rhodoplanes, Bacillus, Streptomyces, Paenibacillus, Phenylobacterium, and Terracoccus.
Supplementary Materials
The following are available online. Table S1: Effect of petroleum (P) and cocksfoot (G) on the count of organotrophic bacteria (Org), actinobacteria (Act), and fungi (Fun) in the soil, 10n cfu kg−1 soil d.m. n—for Org and Act = 9, and for Fun = 7, Table S2: Effect of petroleum (P) and cocksfoot (G) on the activity of enzymes in 1 kg of soil d.m. within 1 h, Table S3: Main properties of soil used in the study, Figure S1: Effect of petroleum on the yield of cocksfoot, d.m. in g per pot.
Author Contributions
A.B. and J.W. conceived and designed the ideas and wrote the manuscript with the help of J.K.; A.B. she did bio-informatic analysis and visualization of the data, conducted the experiments, collected and analysed the data with the help of J.W. and J.K.; all authors contributed to the final version of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The results presented in this paper were obtained as part of a comprehensive study financed by the University of Warmia and Mazury in Olsztyn, Faculty of Agriculture and Forestry, Department of Soil Science and Microbiology (grant No. 30.610.006-110) and the project was financially supported by Minister of Science and Higher Education in the range of the programme entitled “Regional Initiative of Excellence” for the years 2019–2022, Project No. 010/RID/2018/19, amount of funding 12,000,000 PLN.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Sample Availability
Not available.
References
- Erisman, J.W.; van Eekeren, N.; De Wit, J.; Koopmans, C.J.; Cuijpers, W.J.M.; Oerlemans, N.; Koks, B. Agriculture and biodiversity: A better balance benefits both. AIMS Agric. Food 2016, 1, 157–174. [Google Scholar] [CrossRef]
- Huera-Lucero, T.; Labrador-Moreno, J.; Blanco-Salas, J.; Ruiz-Téllez, T. A framework to incorporate biological soil quality indicators into assessing the sustainability of territories in the Ecuadorian Amazon. Sustainability 2020, 12, 3007. [Google Scholar] [CrossRef] [Green Version]
- Odukoya, J.; Lambert, R.; Sakrabani, R. Understanding the impacts of crude oil and its induced abiotic stresses on agrifood production: A Review. Horticulturae 2019, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; da Silva Cabral Pinto, M.-M.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; et al. Lead toxicity: Health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health 2020, 17, 2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garousin, H.; Pourbabaee, A.A.; Alikhani, H.A.; Yazdanfar, N. A combinational strategy mitigated old-aged petroleum contaminants: Ineffectiveness of biostimulation as a bioremediation technique. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Ławniczak, Ł.; Woźniak-Karczewska, M.; Loibner, A.P.; Heipieper, H.J.; Chrzanowski, Ł. Microbial degradation of hydrocarbons—basic principles for bioremediation: A review. Molecules 2020, 25, 856. [Google Scholar] [CrossRef] [Green Version]
- Varjani, S.; Upasani, V.N. Influence of abiotic factors, natural attenuation, bioaugmentation and nutrient supplementation on bioremediation of petroleum crude contaminated agricultural soil. J. Environ. Manag. 2019, 245, 358–366. [Google Scholar] [CrossRef]
- Hernández-Adame, N.M.; López-Miranda, J.; Martínez-Prado, M.A.; Cisneros-de la Cueva, S.; Rojas-Contreras, J.A.; Medrano-Roldán, H. Increase in total petroleum hydrocarbons removal rate in contaminated mining soil through bioaugmentation with autochthonous fungi during the slow bioremediation stage. Water Air Soil Pollut. 2021, 232, 95. [Google Scholar] [CrossRef]
- Jeyasundar, P.G.S.A.; Ali, A.; Azeem, M.; Li, Y.; Guo, D.; Sikdar, A.; Abdelrahman, H.; Kwon, E.; Antoniadis, V.; Mani, V.M.; et al. Green remediation of toxic metals contaminated mining soil using bacterial consortium and Brassica juncea. Environ. Pollut. 2021, 277, 116789. [Google Scholar] [CrossRef]
- Lipińska, A.; Wyszkowska, J.; Kucharski, J. Microbiological and biochemical activity in soil contaminated with pyrene subjected to bioaugmentation. Water Air Soil Pollut. 2021, 232, 45. [Google Scholar] [CrossRef]
- Varjani, S.J.; Gnansounou, E.; Pandey, A. Comprehensive review on toxicity of persistent organic pollutants from petroleum refinery waste and their degradation by microorganisms. Chemosphere 2017, 188, 280–291. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Vestergaard, G.; Schulz, S.; Schöler, A.; Schloter, M. Making Big Data Smart—How to use metagenomics to understand soil quality. Biol. Fertil. Soils 2017, 53, 479–484. [Google Scholar] [CrossRef]
- Nesme, J.; Achouak, W.; Agathos, S.N.; Bailey, M.; Baldrian, P.; Brunel, D.; Frostegård, Å.; Heulin, T.; Jansson, J.K.; Jurkevitch, E.; et al. Back to the future of soil metagenomics. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Amorim, H.C.S.; Ashworth, A.J.; Wienhold, B.J.; Savin, M.C.; Allen, F.L.; Saxton, A.M.; Owens, P.R.; Curi, N. Soil quality indices based on long-term conservation cropping systems management. Agrosyst. Geosci. Environ. 2020, 3, e20036. [Google Scholar] [CrossRef] [Green Version]
- Andrea, F.; Bini, C.; Amaducci, S. Soil and ecosystem services: Current knowledge and evidences from Italian case studies. Appl. Soil Ecol. 2018, 123, 693–698. [Google Scholar] [CrossRef]
- Cardoso, E.J.B.N.; Vasconcellos, R.L.F.; Bini, D.; Miyauchi, M.Y.H.; dos Santos, C.A.; Alves, P.R.L.; de Paula, A.M.; Nakatani, A.S.; de Moraes Pereira, J.; Nogueira, M.A. Soil health: Looking for suitable indicators. What should be considered to assess the effects of use and management on soil health? Sci. Agric. 2013, 70, 274–289. [Google Scholar] [CrossRef] [Green Version]
- Juhos, K.; Czigány, S.; Madarász, B.; Ladányi, M. Interpretation of soil quality indicators for land suitability assessment—A multivariate approach for Central European arable soils. Ecol. Indic. 2019, 99, 261–272. [Google Scholar] [CrossRef]
- Cederlund, H.; Wessén, E.; Enwall, K.; Jones, C.; Juhanson, J.; Pell, M.; Philippot, L.; Hallin, S. Soil carbon quality and nitrogen fertilization structure bacterial communities with predictable responses of major bacterial phyla. Appl. Soil Ecol. 2014, 84, 62–68. [Google Scholar] [CrossRef]
- Deru, J.G.C.; Bloem, J.; de Goede, R.; Keidel, H.; Kloen, H.; Rutgers, M.; van den Akker, J.; Brussaard, L.; van Eekeren, N. Soil ecology and ecosystem services of dairy and semi-natural grasslands on peat. Appl. Soil Ecol. 2018, 125, 26–34. [Google Scholar] [CrossRef]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B.H. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Wang, S.; Wang, Z.; Wang, C.; Mo, H.; Mo, J.; Lu, X. Testing potassium limitation on soil microbial activity in a sub-tropical forest. J. For. Res. 2019, 30, 2341–2347. [Google Scholar] [CrossRef]
- Pereira, D.G.C.; Santana, I.A.; Megda, M.M.; Megda, M.X.V.; Pereira, D.G.C.; Santana, I.A.; Megda, M.M.; Megda, M.X.V. Potassium chloride: Impacts on soil microbial activity and nitrogen mineralization. Ciência Rural 2019, 49. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on changbai mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Biological Characterization ISO Standards. Available online: https://www.iso.org/committee/54366/x/catalogue/completed (accessed on 23 February 2021).
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Perez-Mon, C.; Frey, B.; Frossard, A. Functional and structural responses of arctic and alpine soil prokaryotic and fungal communities under freeze-thaw cycles of different frequencies. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Schloter, M.; Nannipieri, P.; Sørensen, S.J.; van Elsas, J.D. Microbial indicators for soil quality. Biol. Fertil. Soils 2018, 54, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gunina, A.; Dippold, M.A.; Glaser, B.; Kuzyakov, Y. Fate of low molecular weight organic substances in an arable soil: From microbial uptake to utilisation and stabilisation. Soil Biol. Biochem. 2014, 77, 304–313. [Google Scholar] [CrossRef]
- Praeg, N.; Pauli, H.; Illmer, P. Microbial diversity in bulk and rhizosphere soil of ranunculus glacialis along a high-alpine altitudinal gradient. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Huang, C.; Han, X.; Yang, Z.; Chen, Y.; Rengel, Z. Sowing methods influence soil bacterial diversity and community composition in a winter wheat-summer maize rotation system on the Loess Plateau. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Bing, H.; Fang, L.; Wu, Y.; Yu, J.; Shen, G.; Jiang, M.; Wang, X.; Zhang, X. Diversity patterns of the rhizosphere and bulk soil microbial communities along an altitudinal gradient in an alpine ecosystem of the Eastern Tibetan Plateau. Geoderma 2019, 338, 118–127. [Google Scholar] [CrossRef]
- Dos Santos, J.J.; Maranho, L.T. Rhizospheric microorganisms as a solution for the recovery of soils contaminated by petroleum: A review. J. Environ. Manag. 2018, 210, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Adams, C.A.; Yang, W.; Sun, Y.; Shi, Z. Benefits of arbuscular mycorrhizal fungi in reducing organic contaminant residues in crops: Implications for cleaner agricultural production. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1580–1612. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Gumiere, T.; Rousseau, A.N.; da Costa, D.P.; Cassetari, A.; Cotta, S.R.; Andreote, F.D.; Gumiere, S.J.; Pavinato, P.S. Phosphorus source driving the soil microbial interactions and improving sugarcane development. Sci. Rep. 2019, 9, 4400. [Google Scholar] [CrossRef]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N. The role of phosphorus limitation in shaping soil bacterial communities and their metabolic capabilities. mBio 2020, 11. [Google Scholar] [CrossRef]
- Bidja Abena, M.T.; Li, T.; Shah, M.N.; Zhong, W. Biodegradation of total petroleum hydrocarbons (TPH) in highly contaminated soils by natural attenuation and bioaugmentation. Chemosphere 2019, 234, 864–874. [Google Scholar] [CrossRef]
- Camacho-Montealegre, C.M.; Rodrigues, E.M.; Tótola, M.R. Microbial diversity and bioremediation of rhizospheric soils from Trindade Island—Brazil. J. Environ. Manag. 2019, 236, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Mafiana, M.O.; Kang, X.-H.; Leng, Y.; He, L.-F.; Li, S.-W. Petroleum contamination significantly changes soil microbial communities in three oilfield locations in Delta State, Nigeria. Environ. Sci. Pollut. Res. Int. 2021. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Tsai, S.M.; Navarrete, A.A.; de Hollander, M.; van Veen, J.A.; Kuramae, E.E. Soil-borne microbiome: Linking diversity to function. Microb. Ecol. 2015, 70, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Allamin, I.A.; Halmi, M.I.E.; Yasid, N.A.; Ahmad, S.A.; Abdullah, S.R.S.; Shukor, Y. Rhizodegradation of petroleum oily sludge-contaminated soil using cajanus cajan increases the diversity of soil microbial community. Sci. Rep. 2020, 10, 4094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchez, T.; Blieux, A.L.; Dequiedt, S.; Domaizon, I.; Dufresne, A.; Ferreira, S.; Godon, J.J.; Hellal, J.; Joulian, C.; Quaiser, A.; et al. Molecular microbiology methods for environmental diagnosis. Environ. Chem. Lett. 2016, 14, 423–441. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowik, A.; Wyszkowska, J.; Oszust, K. Changes in the functional diversity of bacterial communities in soil contaminated with diesel oil. J. Elem. 2018, 23, 1099–1117. [Google Scholar] [CrossRef]
- Stone, D.; Ritz, K.; Griffiths, B.G.; Orgiazzi, A.; Creamer, R.E. Selection of biological indicators appropriate for European soil monitoring. Appl. Soil Ecol. 2016, 97, 12–22. [Google Scholar] [CrossRef]
- Lynch, M.D.J.; Neufeld, J.D. Ecology and exploration of the rare biosphere. Nat. Rev. Microbiol. 2015, 13, 217–229. [Google Scholar] [CrossRef]
- Kandziora-Ciupa, M.; Nadgórska-Socha, A.; Barczyk, G. The influence of heavy metals on biological soil quality assessments in the Vaccinium myrtillus L. rhizosphere under different field conditions. Ecotoxicology 2021, 30, 292–310. [Google Scholar] [CrossRef]
- Zhu, H.; Teng, Y.; Wang, X.; Zhao, L.; Ren, W.; Luo, Y.; Christie, P. Changes in clover rhizosphere microbial community and diazotrophs in mercury-contaminated soils. Sci. Total Environ. 2021, 767, 145473. [Google Scholar] [CrossRef] [PubMed]
- Ite, A.E.; Ibok, U.J. Role of plants and microbes in bioremediation of petroleum hydrocarbons contaminated soils. Int. J. Environ. Bioremediat. Biodegrad. 2019, 7, 1–19. [Google Scholar] [CrossRef]
- Shen, H.; Huang, Y.; Wang, R.; Zhu, D.; Li, W.; Shen, G.; Wang, B.; Zhang, Y.; Chen, Y.; Lu, Y.; et al. Global atmospheric emissions of polycyclic aromatic hydrocarbons from 1960 to 2008 and future predictions. Environ Sci Technol. 2013, 47, 6415–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaborowska, M.; Wyszkowska, J.; Borowik, A. Soil microbiome response to contamination with bisphenol A, bisphenol F and bisphenol S. Int. J. Mol. Sci. 2020, 21, 3529. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Dashti, N.; Khanafer, M.; Al-Awadhi, H.; Radwan, S. Bioremediation of soils saturated with spilled crude oil. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Shen, D.; Yuan, X.; Han, Y.; Mo, R.; Zhong, D.; Tang, F.; Liu, Y. Evaluation of Polycyclic Aromatic Hydrocarbons (PAHs) in bamboo shoots from soil. Bull. Environ. Contam. Toxicol. 2021. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of bacterial and fungal communities to high petroleum pollution in different soils. Sci. Rep. 2021, 11, 164. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Kucharski, M.; Kucharski, J. Implications of soil pollution with diesel oil and BP petroleum with active technology for soil health. Int. J. Environ. Res. Public Health 2019, 16, 2474. [Google Scholar] [CrossRef] [Green Version]
- Nebeská, D.; Auer Malinská, H.; Erol, A.; Pidlisnyuk, V.; Kuráň, P.; Medžová, A.; Smaha, M.; Trögl, J. Stress response of miscanthus plants and soil microbial communities: A case study in metals and hydrocarbons contaminated soils. Appl. Sci. 2021, 11, 1866. [Google Scholar] [CrossRef]
- Benedek, T.; Vajna, B.; Táncsics, A.; Márialigeti, K.; Lányi, S.; Máthé, I. Remarkable impact of PAHs and TPHs on the richness and diversity of bacterial species in surface soils exposed to long-term hydrocarbon pollution. World J. Microbiol. Biotechnol. 2013, 29, 1989–2002. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Insights into microbial communities mediating the bioremediation of hydrocarbon-contaminated soil from an Alpine former military site. Appl. Microbiol. Biotechnol. 2018, 102, 4409–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Zhang, X.; Wang, X.; Fu, S.; Wu, S.; Lu, X.; Ren, C.; Han, X.; Yang, G. Soil bacteria and fungi respond differently to plant diversity and plant family composition during the secondary succession of abandoned farmland on the Loess Plateau, China. Plant Soil 2020, 448, 183–200. [Google Scholar] [CrossRef]
- Tejeda-Agredano, M.C.; Gallego, S.; Vila, J.; Grifoll, M.; Ortega-Calvo, J.J.; Cantos, M. Influence of the sunflower rhizosphere on the biodegradation of PAHs in soil. Soil Biol. Biochem. 2013, 57, 830–840. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; AlMomin, S.; Al-Aqeel, H.; Al-Salameen, F.; Nair, S.; Shajan, A. Metagenomic analysis of rhizosphere microflora of oil-contaminated soil planted with barley and alfalfa. PLoS ONE 2018, 13, e0202127. [Google Scholar] [CrossRef]
- De la Cueva, S.C.; Rodríguez, C.H.; Cruz, N.O.S.; Contreras, J.A.R.; Miranda, J.L. Changes in bacterial populations during bioremediation of soil contaminated with petroleum hydrocarbons. Water Air Soil Pollut. 2016, 227, 91. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Zhang, X. Bioremediation of petroleum hydrocarbon-contaminated soil by petroleum-degrading bacteria immobilized on biochar. RSC Adv. 2019, 9, 35304–35311. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, A.; Kukal, S.S.; Saha, D.; Sharma, H.; Kalia, A.; Sharma, S. Potential indicators of soil health degradation in different land use-based ecosystems in the Shiwaliks of Northwestern India. Sustainability 2019, 11, 3908. [Google Scholar] [CrossRef] [Green Version]
- Galitskaya, P.; Biktasheva, L.; Kuryntseva, P.; Selivanovskaya, S. Response of soil bacterial communities to high petroleum content in the absence of remediation procedures. Environ. Sci. Pollut. Res. 2021, 28, 9610–9627. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Wyszkowski, M. Resistance of aerobic microorganisms and soil enzyme response to soil contamination with Ekodiesel Ultra Fuel. Environ. Sci. Pollut. Res. 2017, 24, 24346–24363. [Google Scholar] [CrossRef] [Green Version]
- Ramadass, K.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Ecotoxicity of measured concentrations of soil-applied diesel: Effects on earthworm survival, dehydrogenase, urease and nitrification activities. Appl. Soil Ecol. 2017, 119, 1–7. [Google Scholar] [CrossRef]
- Novakovskiy, A.B.; Kanev, V.A.; Markarova, M.Y. Long-term dynamics of plant communities after biological remediation of oil-contaminated soils in Far North. Sci. Rep. 2021, 11, 4888. [Google Scholar] [CrossRef]
- Alrumman, S.A.; Standing, D.B.; Paton, G.I. Effects of hydrocarbon contamination on soil microbial community and enzyme activity. J. King Saud Univ. Sci. 2015, 27, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Dindar, E.; Topaç Şağban, F.O.; Başkaya, H.S. Variations of soil enzyme activities in petroleum-hydrocarbon contaminated soil. Int. Biodeter. Biodegr. 2015, 105, 268–275. [Google Scholar] [CrossRef]
- Curyło, K.; Telesiński, A.; Jarnuszewski, G.; Krzyśko-Łupicka, T.; Cybulska, K. Analysis of chemical and biochemical parameters of petrol-contaminated soil after biostimulation with an enzyme reagent. Processes 2020, 8, 949. [Google Scholar] [CrossRef]
- Kaczyńska, G.; Borowik, A.; Wyszkowska, J. Soil dehydrogenases as an indicator of contamination of the environment with petroleum products. Water Air Soil Pollut. 2015, 226, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravindh, S.; Chinnadurai, C.; Balachandar, D. Development of a soil biological quality index for soils of semi-arid tropics. Soil 2020, 6, 483–497. [Google Scholar] [CrossRef]
- De la Paz Jimenez, M.; de la Horra, A.; Pruzzo, L.; Palma, M. Soil quality: A new index based on microbiological and biochemical parameters. Biol. Fertil. Soils 2002. [Google Scholar] [CrossRef]
- Klimkowicz-Pawlas, A.; Ukalska-Jaruga, A.; Smreczak, B. Soil quality index for agricultural areas under different levels of anthropopressure. Int. Agrophys. 2019, 33, 455–462. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Kucharski, M.; Kucharski, J. Effect of cadmium, copper and zinc on plants, soil microorganisms and soil enzymes. J. Elem. 2013, 18, 769–796. [Google Scholar] [CrossRef]
- Chakravarty, P.; Deka, H. Enzymatic defense of Cyperus brevifolius in hydrocarbons stress environment and changes in soil properties. Sci. Rep. 2021, 11, 718. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Xu, J.; Kong, X.; Zhao, L.; Zeng, D.-H. Assessing the quality of oil-contaminated saline soil using two composite indices. Ecol. Indic. 2013, 24, 105–112. [Google Scholar] [CrossRef]
- Gospodarek, J.; Rusin, M.; Barczyk, G.; Nadgórska-Socha, A. The effect of petroleum-derived substances and their bioremediation on soil enzymatic activity and soil invertebrates. Agronomy 2021, 11, 80. [Google Scholar] [CrossRef]
- Zuzolo, D.; Guarino, C.; Tartaglia, M.; Sciarrillo, R. Plant-soil-microbiota combination for the removal of total petroleum hydrocarbons (TPH): An in-field experiment. Front. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Al-Sayegh, A.; Al-Wahaibi, Y.; Joshi, S.; Al-Bahry, S.; Elshafie, A.; Al-Bemani, A. Bioremediation of heavy crude oil contamination. Open Biotechnol. J. 2016, 10. [Google Scholar] [CrossRef]
- Correa-García, S.; Rheault, K.; Tremblay, J.; Séguin, A.; Yergeau, E. Soil characteristics constrain the response of microbial communities and associated hydrocarbon degradation genes during phytoremediation. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef] [PubMed]
- Hoang, S.A.; Lamb, D.; Seshadri, B.; Sarkar, B.; Cheng, Y.; Wang, L.; Bolan, N.S. Petroleum hydrocarbon rhizoremediation and soil microbial activity improvement via cluster root formation by wild proteaceae plant species. Chemosphere 2021, 275, 130135. [Google Scholar] [CrossRef]
- BP, p.l.c. Website. Available online: https://www.bp.com/ (accessed on 23 February 2021).
- BP’s Fuels Characteristics Cards. Available online: https://www.bp.com/pl_pl/poland/home/produkty_uslugi/hurt_paliw.html (accessed on 23 February 2021).
- Raman, J.K.; Alves, C.M.; Gnansounou, E. A review on moringa tree and vetiver grass—Potential biorefinery feedstocks. Bioresour. Technol. 2018, 249, 1044–1051. [Google Scholar] [CrossRef]
- Visconti, D.; Álvarez-Robles, M.J.; Fiorentino, N.; Fagnano, M.; Clemente, R. Use of Brassica juncea and Dactylis glomerata for the phytostabilization of mine soils amended with compost or biochar. Chemosphere 2020, 260, 127661. [Google Scholar] [CrossRef]
- Yang, Z.; Nie, G.; Feng, G.; Han, J.; Huang, L.; Zhang, X. Genome-wide identification, characterization, and expression analysis of the NAC transcription factor family in orchardgrass (Dactylis glomerata L.). BMC Genom. 2021, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gajić, G.; Djurdjević, L.; Kostić, O.; Jarić, S.; Stevanović, B.; Mitrović, M.; Pavlović, P. Phytoremediation potential, photosynthetic and antioxidant response to arsenic-induced stress of Dactylis glomerata L. sown on fly ash deposits. Plants 2020, 9, 657. [Google Scholar] [CrossRef]
- Li, S.; Nie, Z.; Zhang, D. Competition between Cocksfoot (Dactylis Glomerata L.) and companion species: Evidence for allelopathy. Field Crops Res. 2016, 196, 452–462. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Skinner, R.H.; Elwinger, G.F. Seedling development and field performance of prairiegrass, grazing bromegrass, and orchardgrass. Crop Sci. 2002, 42, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Gałązka, A.; Gałązka, R. Phytoremediation of polycyclic aromatic hydrocarbons in soils artificially polluted using plant-associated-endophytic bacteria and Dactylis glomerata as the bioremediation plant. Pol. J. Microbiol. 2015, 64, 241–252. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Olszewski, J.; Kucharski, J. Soil bacterial community and soil enzyme activity depending on the cultivation of Triticum aestivum, Brassica napus, and Pisum sativum ssp. arvense. Diversity 2019, 11, 246. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 22 March 2021).
- ISO 18287 Soil Quality—Determination of Polycyclic Aromatic Hydrocarbons (PAH)—Gas Chromatographic Method with Mass Spectrometric Detection (GC-MS); International Organization for Standardization: Geneva, Switzerland, 2006.
- EN ISO 16703 Soil Quality—Determination of Content of Hydrocarbon in the Range c10 to c40 by Gas Chromatography; International Organization for Standardization: Geneva, Switzerland, 2004.
- EN ISO 22155 Soil Quality—Gas Chromatographic Determination of Volatile Aromatic and Halogenated Hydrocarbons and Selected Ethers—Static Headspace Method; International Organization for Standardization: Geneva, Switzerland, 2016.
- De Leij, F.A.A.M.; Whipps, J.M.; Lynch, J.M. The use of colony development for the characterization of bacterial communities in soil and on roots. Microb. Ecol. 1993, 27, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Orwin, K.H.; Wardle, D.A. New indices for quantifying the resistance and resilience of soil biota to exogenous disturbances. Soil Biol. Biochem. 2004, 36, 1907–1912. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System), version 13.3; TIBCO Software Inc.: Palo Alto, CA, USA, 2017. [Google Scholar]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 23 February 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 23 February 2020).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, M.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data; R Package Version 2.17.0; 2020; Available online: https://CRAN.R-project.org/package=gplots (accessed on 23 February 2020).
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Indices effect: (a) of cocksfoot on soil microorganisms (IFG), (b) of petrol on soil microorganisms (IFP), and (c) of resistance (RS) of soil microorganisms to the effects of petrol (P). Homogeneous groups denoted with letters (a,b) were calculated separately for each of the microorganisms.
Figure 1.
Indices effect: (a) of cocksfoot on soil microorganisms (IFG), (b) of petrol on soil microorganisms (IFP), and (c) of resistance (RS) of soil microorganisms to the effects of petrol (P). Homogeneous groups denoted with letters (a,b) were calculated separately for each of the microorganisms.
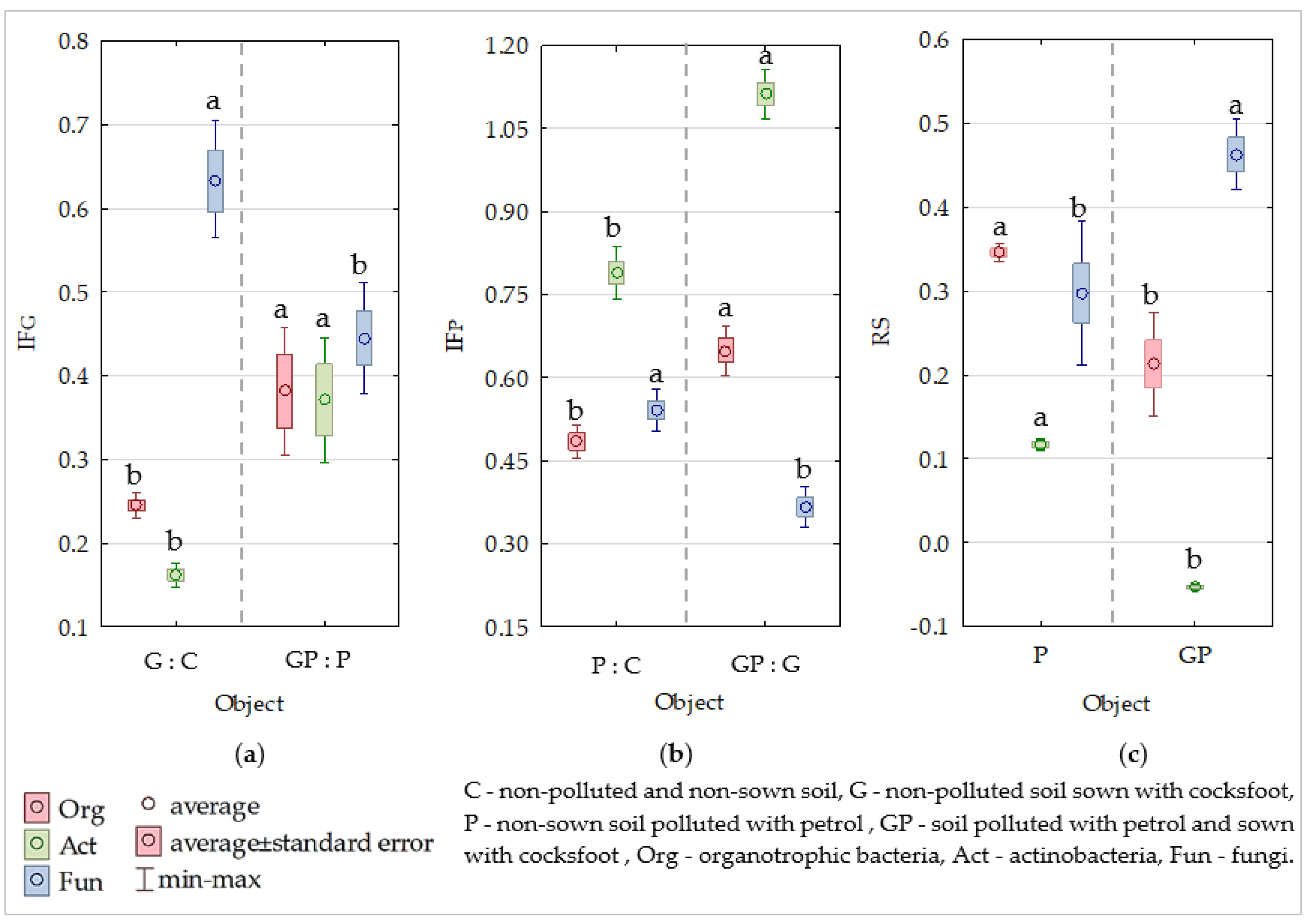
Figure 2.
Effect of petrol (P) and cocksfoot (G) on changes in (a) the colony development (CD) index, and (b) soil microorganisms and the value of their ecophysiological diversity (EP) index. Homogeneous groups denoted with letters (a,b) were calculated separately for each of the microorganisms.
Figure 2.
Effect of petrol (P) and cocksfoot (G) on changes in (a) the colony development (CD) index, and (b) soil microorganisms and the value of their ecophysiological diversity (EP) index. Homogeneous groups denoted with letters (a,b) were calculated separately for each of the microorganisms.

Figure 3.
Comparison of the relative abundance of bacterial phyla in the soil between particular pots, with difference between the proportions at ≥1%. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
Figure 3.
Comparison of the relative abundance of bacterial phyla in the soil between particular pots, with difference between the proportions at ≥1%. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
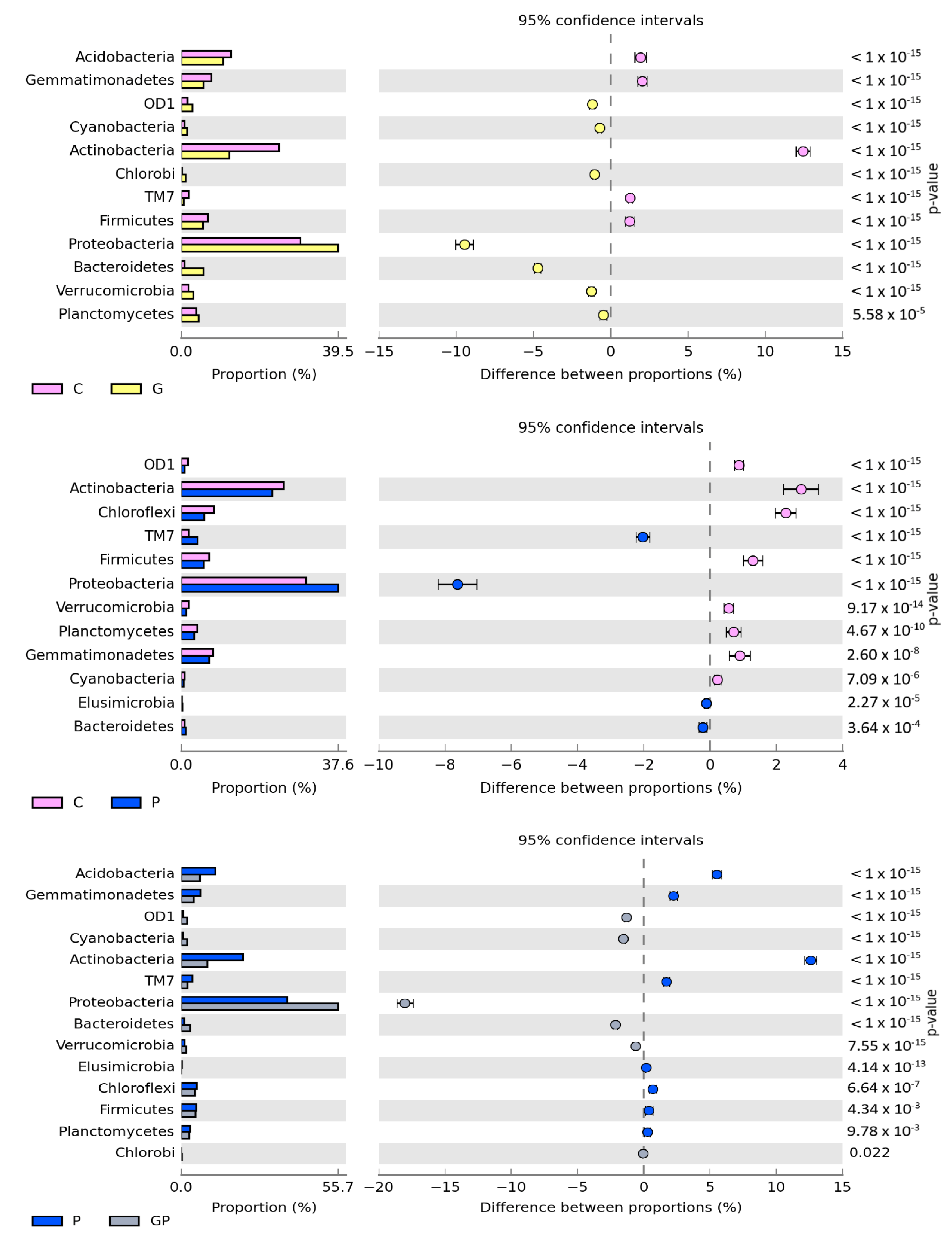
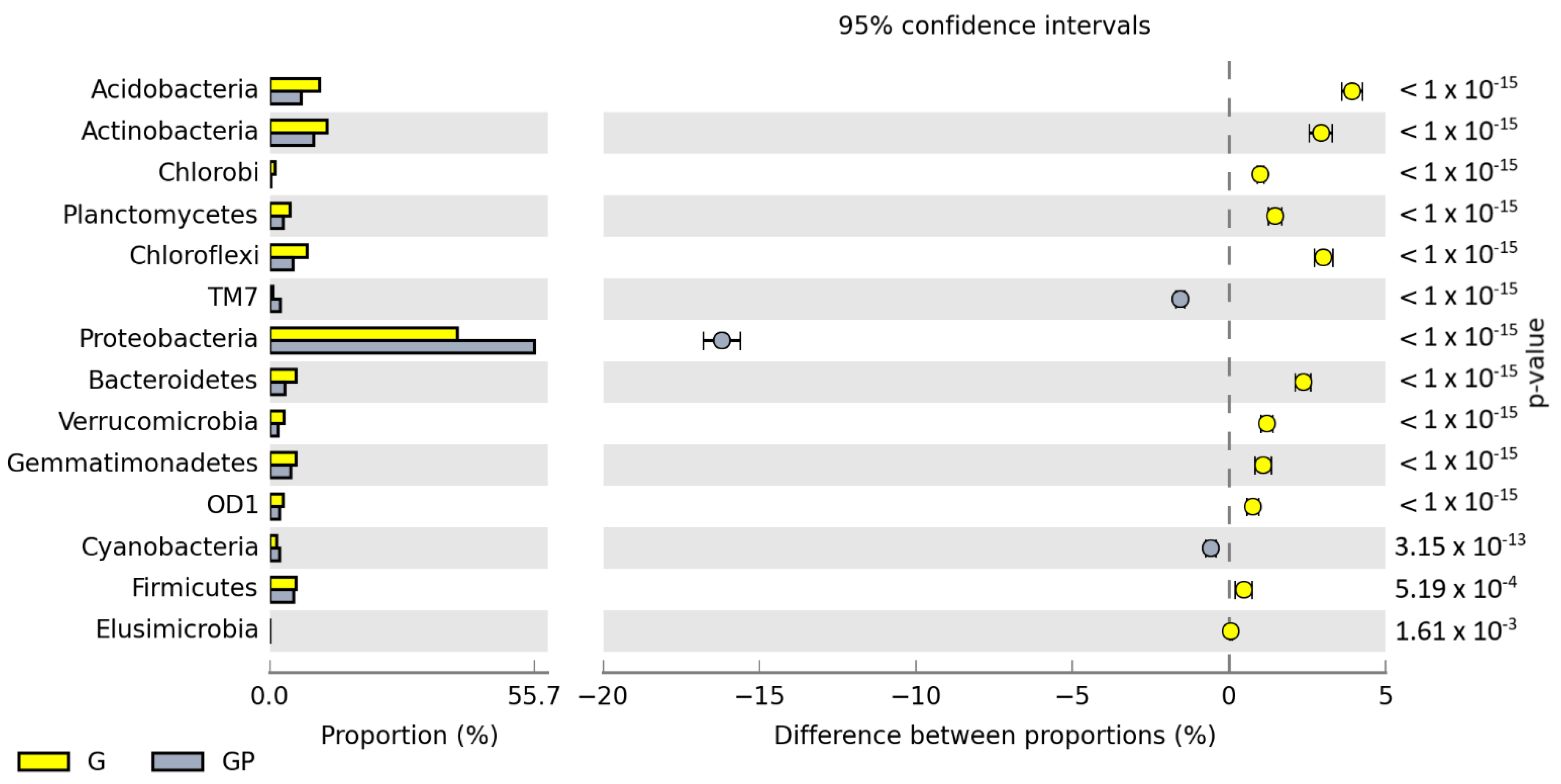
Figure 4.
Number of OTU ≥ 1% of 42 bacterial genera in the soil presented using the heat map with classification to the phylum. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
Figure 4.
Number of OTU ≥ 1% of 42 bacterial genera in the soil presented using the heat map with classification to the phylum. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.

Figure 5.
Effect of petrol (P) and cocksfoot (G) on (a) the number of unique and common bacterial genera, (b) unique and common genera of bacteria colonizing particular pots presented using the Veen diagram. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
Figure 5.
Effect of petrol (P) and cocksfoot (G) on (a) the number of unique and common bacterial genera, (b) unique and common genera of bacteria colonizing particular pots presented using the Veen diagram. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
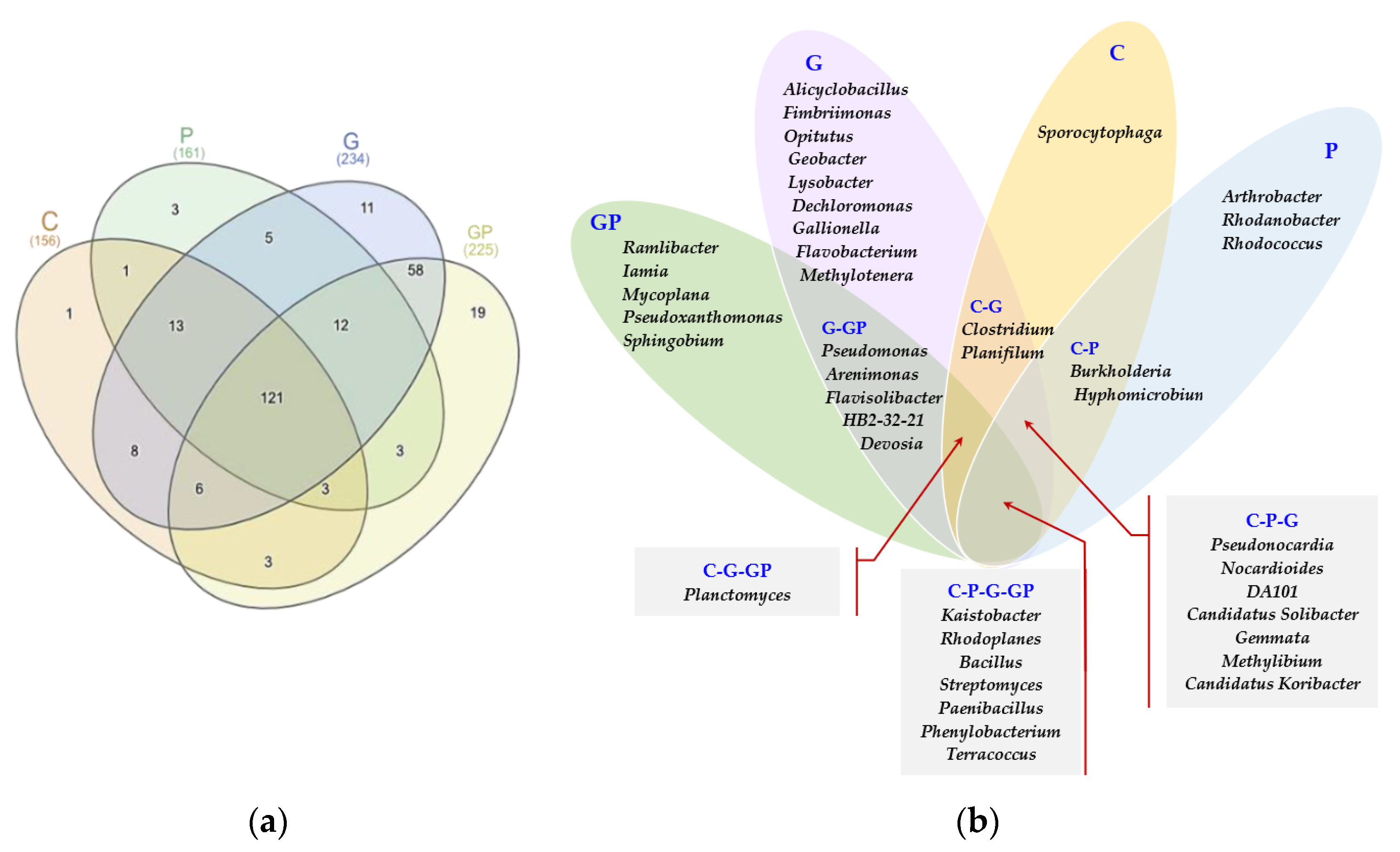
Figure 6.
Phylogenetic tree of unique and common bacterial species. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
Figure 6.
Phylogenetic tree of unique and common bacterial species. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot.
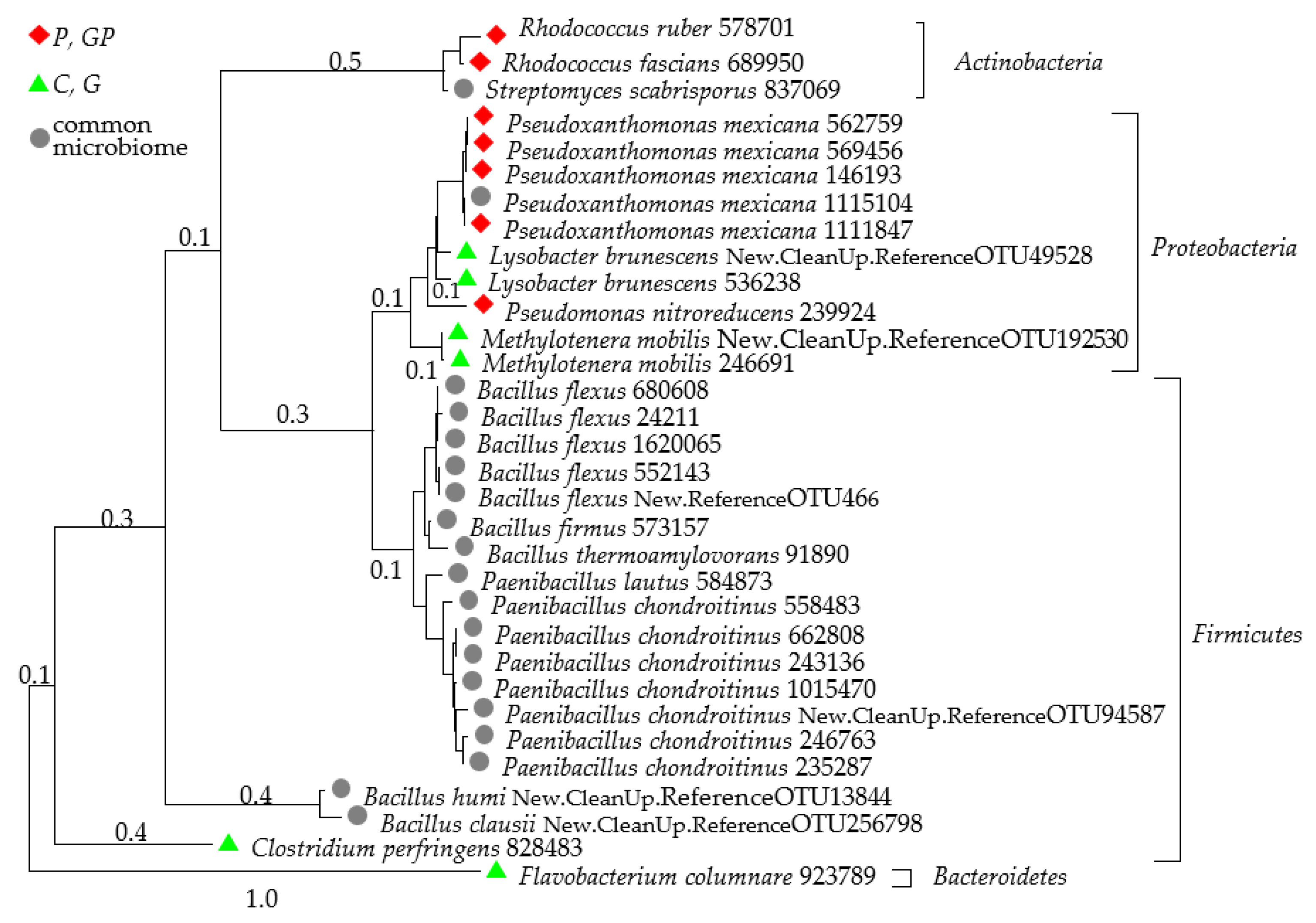
Figure 7.
Effect of petrol (P) and cocksfoot (G) on the bacteria diversity, at particular taxonomic levels, estimated using the Shannon–Wiener index. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot. Homogeneous groups denoted with letters (a–d) were calculated separately for each taxon.
Figure 7.
Effect of petrol (P) and cocksfoot (G) on the bacteria diversity, at particular taxonomic levels, estimated using the Shannon–Wiener index. C—non-polluted and non-sown soil, G—non-polluted soil sown with cocksfoot, P—non-sown soil polluted with petrol, and GP—soil polluted with petrol and sown with cocksfoot. Homogeneous groups denoted with letters (a–d) were calculated separately for each taxon.

Figure 8.
Indices of: (a) cocksfoot effect (IFG) on the activity of soil enzymes, (b) petrol effect (IFP) on the activity of soil enzymes, and (c) resistance (RS) of soil enzymes to the effects of petrol (P). Homogeneous groups denoted with letters (a,b) were calculated separately for each enzyme.
Figure 8.
Indices of: (a) cocksfoot effect (IFG) on the activity of soil enzymes, (b) petrol effect (IFP) on the activity of soil enzymes, and (c) resistance (RS) of soil enzymes to the effects of petrol (P). Homogeneous groups denoted with letters (a,b) were calculated separately for each enzyme.
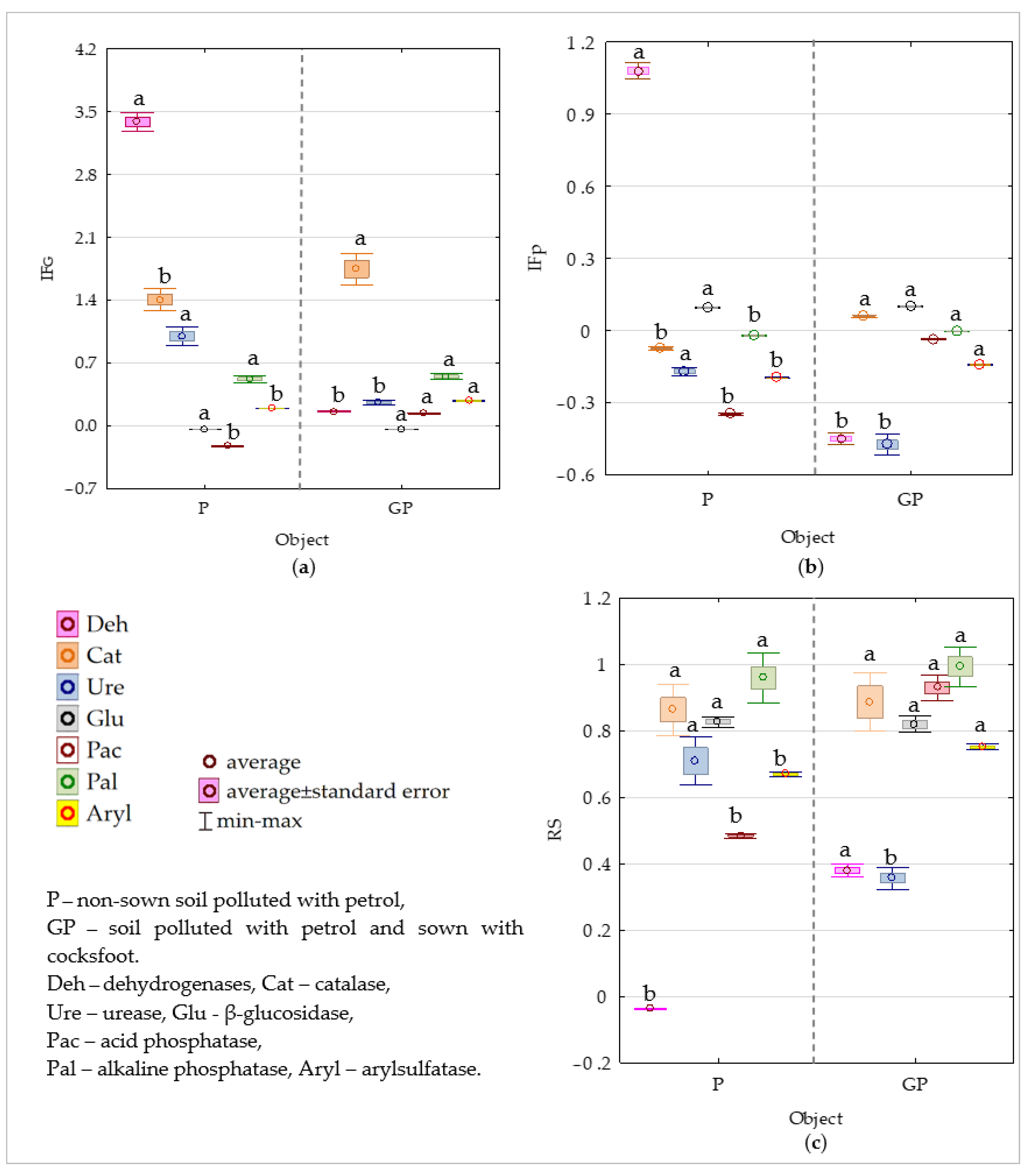
Figure 9.
Index of resistance (RS) of cocksfoot to the effects of petrol. Homogeneous groups denoted with letters (a,b,c) were calculated separately for each of hydrocarbons.
Figure 9.
Index of resistance (RS) of cocksfoot to the effects of petrol. Homogeneous groups denoted with letters (a,b,c) were calculated separately for each of hydrocarbons.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of cocksfoot on degradation of petrol hydrocarbons in the soil, in %.
| Object | C6–C12 | C12–C35 | Ben | EtB | Tol | Xyl | Nap |
| P | 99.53 ± 1.06 a | 71.41 ± 0.70 b | 95.81 ± 0.09 b | 99.83 ± 0.09 a | 99.86 ± 0.15 a | 99.87 ± 0.10 a | 90.63 ± 0.09 b |
| GP | 99.76 ± 0.90 a | 76.56 ± 0.69 a | 98.11 ± 0.19 a | 99.95 ± 0.13 a | 99.96 ± 0.20 a | 99.96 ± 0.13 a | 99.84 ± 0.09 a |
| Object | Ant | Chr | BaA | BaP | BbF | BkF | IP |
| P | 59.09 ± 0.14 b | 18.33 ± 0.10 b | 40.91 ± 0.09 b | 35.00 ± 0.07 b | 16.92 ± 0.04 b | 13.33 ± 0.07 b | 40.00 ± 0.08 a |
| GP | 84.85 ± 0.11a | 41.67 ± 0.09 a | 54.55 ± 0.11 a | 50.00 ± 0.10 a | 30.77 ± 0.06 a | 33.33 ± 0.07 a | 40.00 ± 0.08 a |
C6–C12—gasoline fractions; C12–C35—mineral oil; Ben—benzene; EtB—ethylbenzene; Tol—toluene; X—xylene; Nap—naphthalene; Ant—anthracene; Chr—chrysene; BaA—benzo[a]anthracene; BaP—benzo(a)pyrene; BbF—benzo[b]fluoranthene; BkF—benzo(k)fluoranthene; IP—indeno(1.2.3-cd)pyrene. P—non-sown soil polluted with petrol. GP—soil polluted with petrol and sown with cocksfoot. Homogeneous groups denoted with letters (a,b) were calculated separately for each of hydrocarbons.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Borowik, A.; Wyszkowska, J.; Kucharski, J. Microbiological Study in Petrol-Spiked Soil. Molecules 2021, 26, 2664. https://doi.org/10.3390/molecules26092664
AMA Style
Borowik A, Wyszkowska J, Kucharski J. Microbiological Study in Petrol-Spiked Soil. Molecules. 2021; 26(9):2664. https://doi.org/10.3390/molecules26092664
Chicago/Turabian StyleBorowik, Agata, Jadwiga Wyszkowska, and Jan Kucharski. 2021. "Microbiological Study in Petrol-Spiked Soil" Molecules 26, no. 9: 2664. https://doi.org/10.3390/molecules26092664