
Antiparasitic Ovalicin Derivatives from Pseudallescheria boydii, a Mutualistic Fungus of French Guiana Termites

 , , , and
, , , and
Abstract
:
1. Introduction
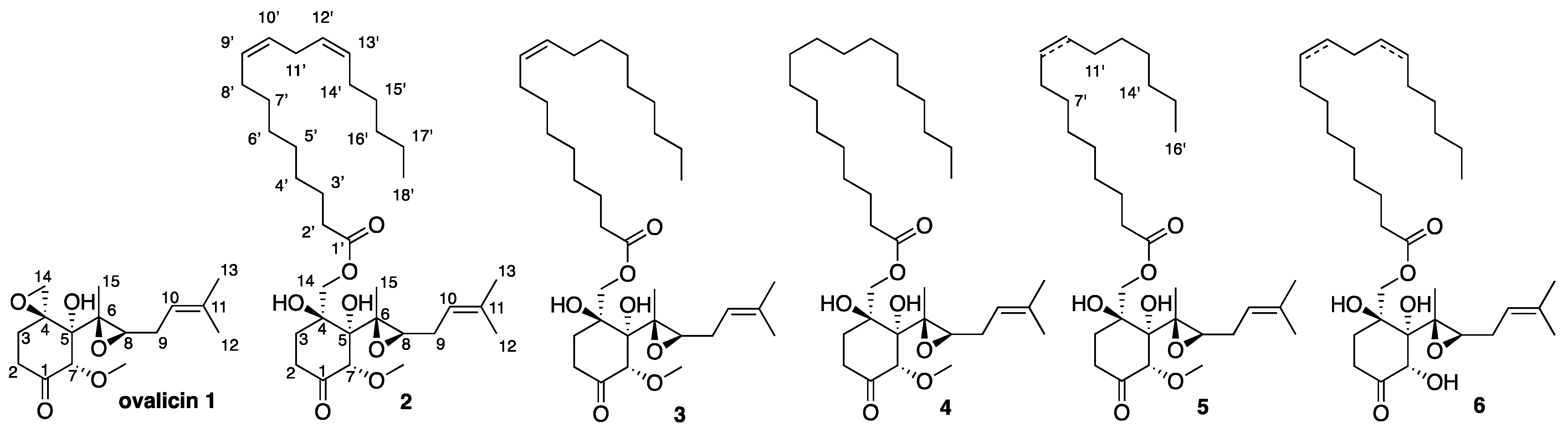
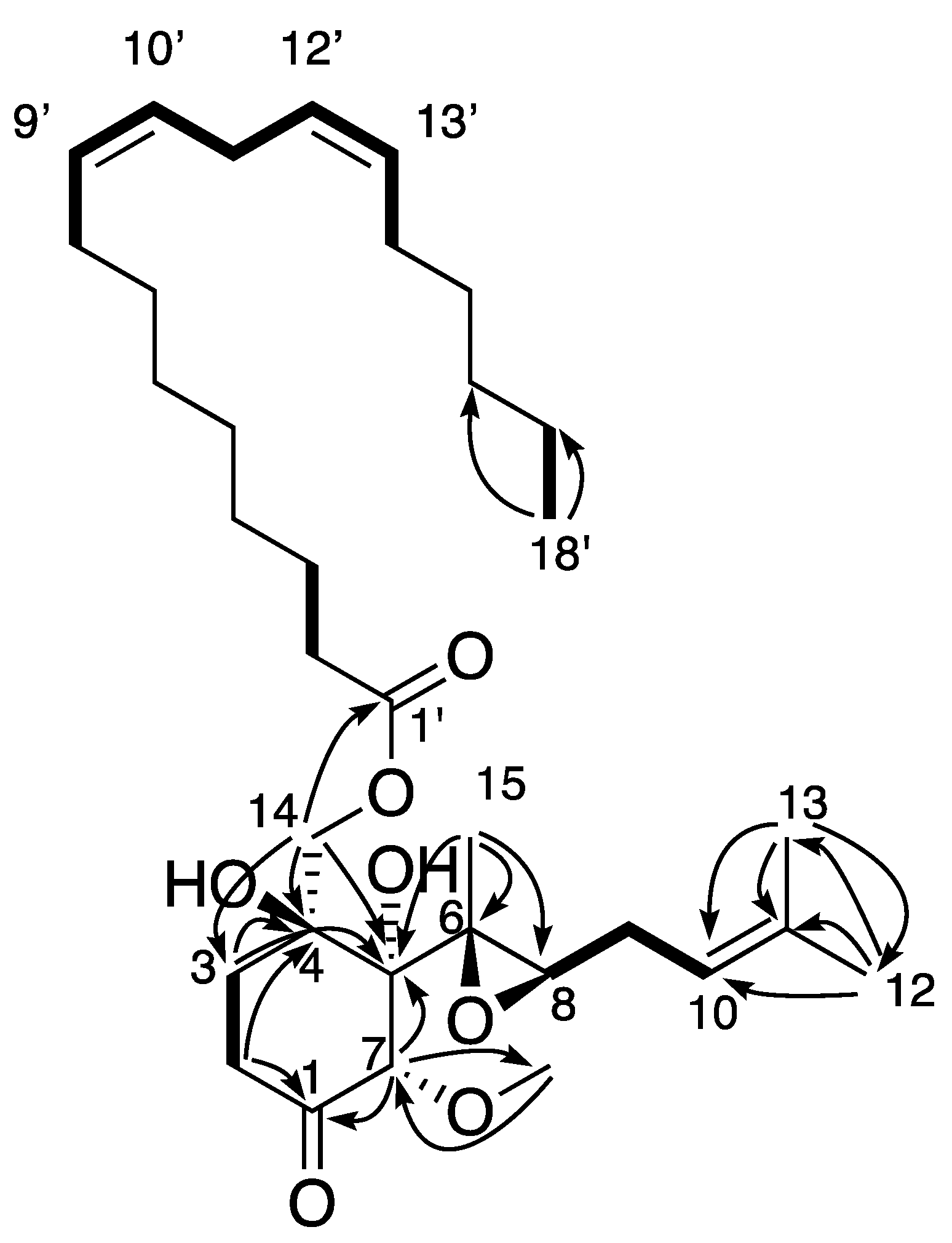
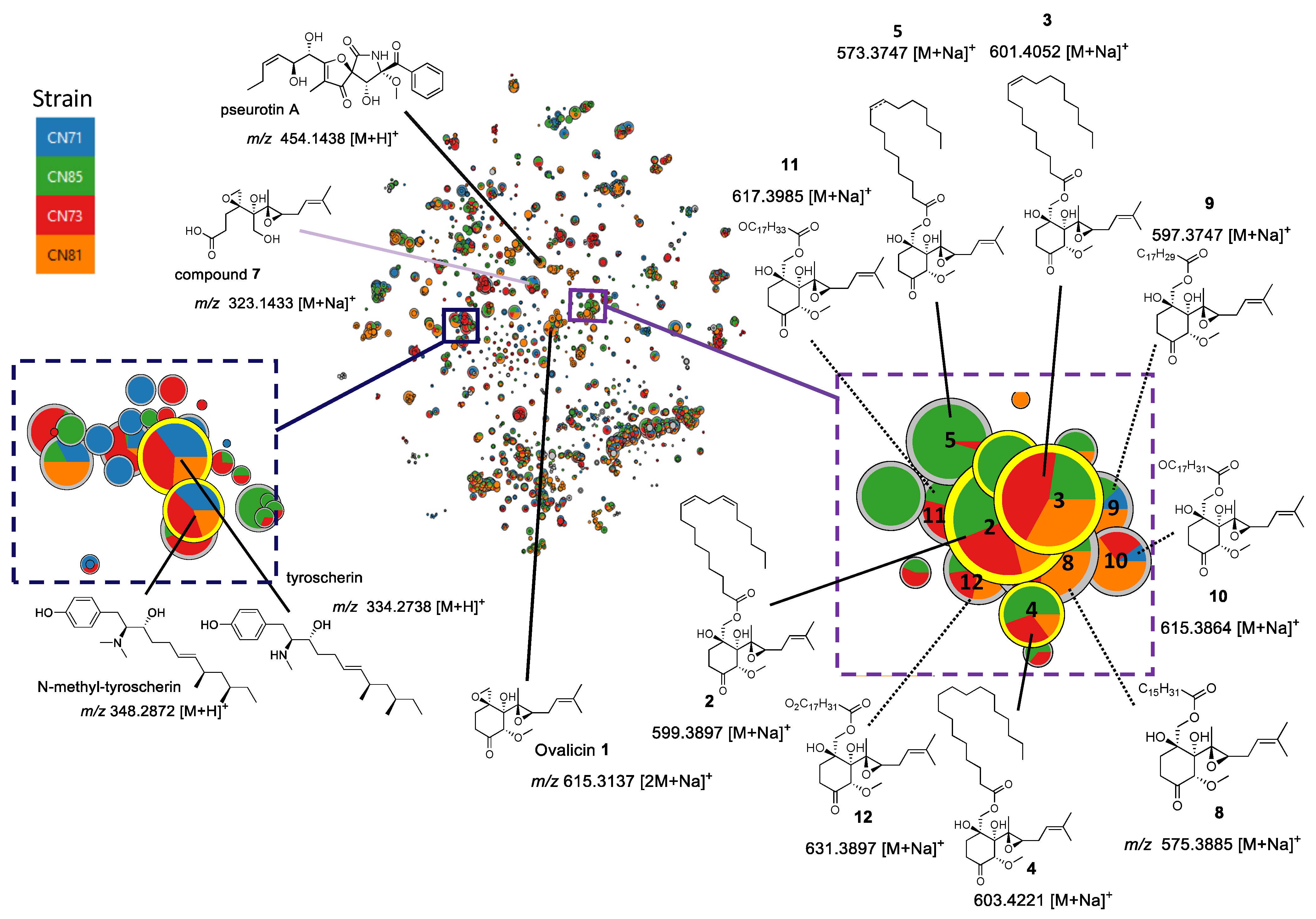
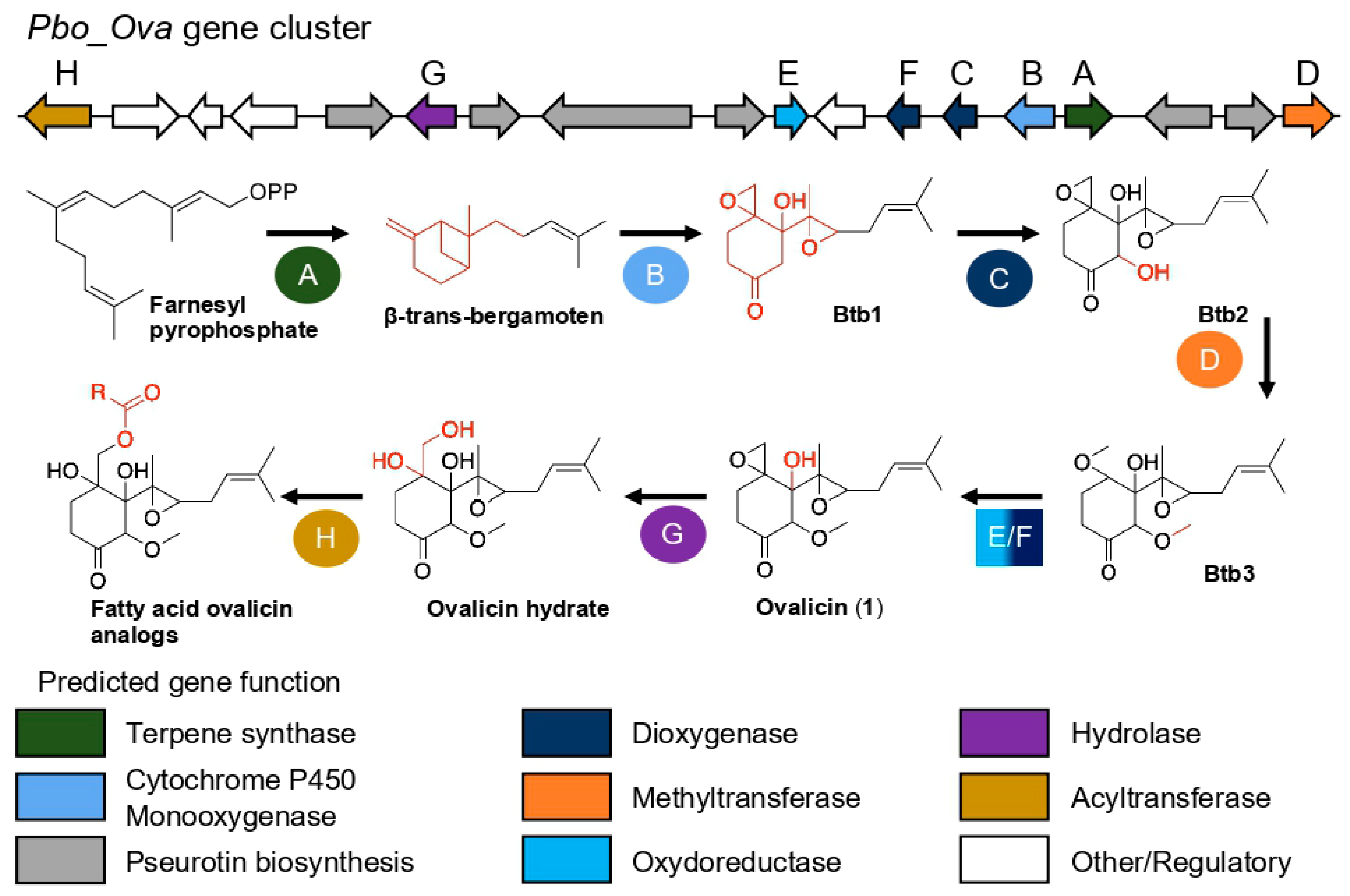
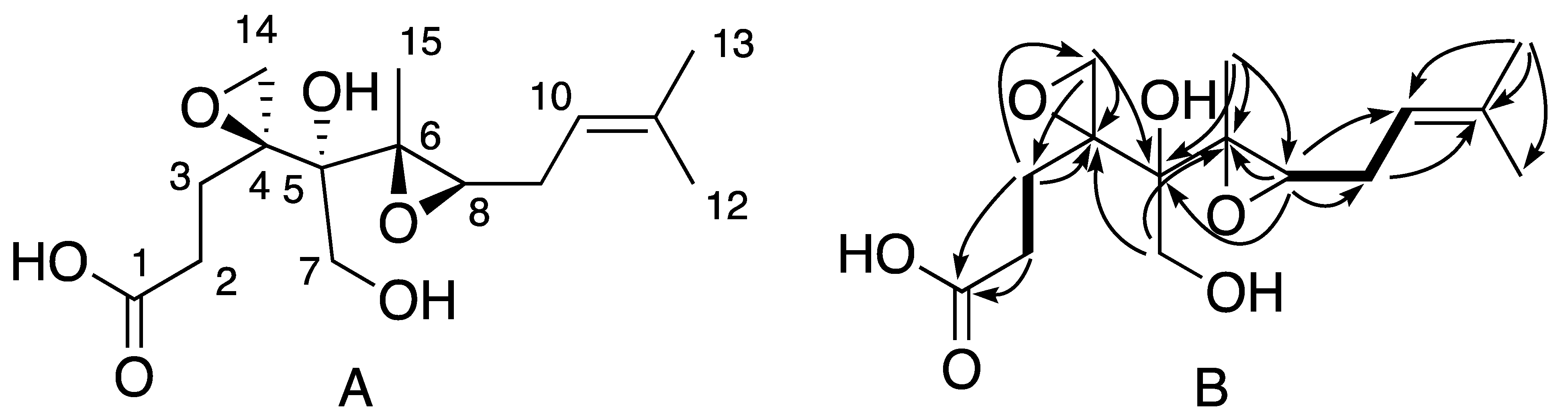
2. Results
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Collection and Identification of Pseudallescheria boydii
3.3. Culture, Extraction, and Isolation
3.4. Fatty Acid Methyl Ester Synthesis from Ovalicin Linoleate (2) and Ovalicin Oleate (3)
3.4.1. Hydrolysis
3.4.2. Esterification
3.5. Semisynthesis of Ovalicin Linoleate, Ovalicin Oleate, and Ovalicin Stearate
3.5.1. Synthesis of Ovalicin Linoleate (2b)
3.5.2. Synthesis of Ovalicin Oleate (3b)
3.5.3. Synthesis of Ovalicin Stearate (4b)
3.6. Identification of the Double-Bond Position in Fatty Acid Methyl Esters by Liquid Chromatography/Atmospheric Pressure Chemical Ionization Mass Spectrometry
3.7. Metabolomic Profiling and Molecular Network Analysis
3.7.1. LC–MS/MS and Data Analysis
3.7.2. Data Processing and Analysis
3.8. Genomic Analyses
3.8.1. gDNA Extraction of P. boydii Strains SNB-CN71, -CN73, -CN81, and -CN85
3.8.2. Whole Genome Sequencing and Hybrid Assembly
3.9. Biological Tests
3.9.1. Antimicrobial Assays
3.9.2. Antiparasitic Activities
Assay for In Vitro Inhibition of T. brucei brucei Growth
In Vitro Growth Inhibition of Chloroquine-Resistant P. falciparum
3.9.3. Cytotoxic Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Adnani, N.; Rajski, S.R.; Bugni, T.S. Symbiosis-inspired approaches to antibiotic discovery. Nat. Prod. Rep. 2017, 34, 784–814. [Google Scholar] [CrossRef] [PubMed]
- van Arnam, E.B.; Currie, C.R.; Clardy, J. Defense contracts: Molecular protection in insect-microbe symbioses. Chem. Soc. Rev. 2018, 47, 1638–1651. [Google Scholar] [CrossRef]
- Beemelmanns, C.; Guo, H.; Rischer, M.; Poulsen, M. Natural products from microbes associated with insects. Beilstein J. Org. Chem. 2016, 12, 314–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, G.; Poulsen, M.; Klassen, J.L.; Hou, Y.; Wyche, T.P.; Bugni, T.S.; Currie, C.R.; Clardy, J. Microtermolides A and B from Termite-Associated Streptomyces sp. and Structural Revision of Vinylamycin. Org. Lett. 2012, 14, 2822–2825. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A.; Tarnita, C.E.; Wilson, E.O. The evolution of eusociality. Nature 2010, 466, 1057–1062. [Google Scholar] [CrossRef]
- Matsui, T.; Tanaka, J.; Namihira, T.; Shinzato, N. Antibiotics production by an actinomycete isolated from the termite gut. J. Basic Microbiol. 2012, 52, 731–735. [Google Scholar] [CrossRef]
- Nirma, C.; Eparvier, V.; Stien, D. Antibacterial Ilicicolinic Acids C and D and Ilicicolinal from Neonectria discophora SNB-CN63 Isolated from a Termite Nest. J. Nat. Prod. 2015, 78, 159–162. [Google Scholar] [CrossRef]
- Nirma, C.; Eparvier, V.; Stien, D. Reactivation of antibiosis in the entomogenous fungus Chrysoporthe sp. SNB-CN74. J. Antibiot. 2015, 68, 586–590. [Google Scholar] [CrossRef]
- Sorres, J.; Sabri, A.; Brel, O.; Stien, D.; Eparvier, V. Ilicicolinic acids and ilicicolinal derivatives from the fungus Neonectria discophora SNB-CN63 isolated from the nest of the termite Nasutitermes corniger found in French Guiana show antimicrobial activity. Phytochemistry 2018, 151, 69–77. [Google Scholar] [CrossRef]
- Jouquet, P.; Chaudhary, E.; Kumar, A.R.V. Sustainable use of termite activity in agro-ecosystems with reference to earthworms. Agron. Sustain. Dev. 2018, 38, 3. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ahmad, W.; Paul, B. Ecological impacts of termites. In Termites and Sustainable Management. Sustainability in Plant and Crop Protection; Khan, M., Ahmad, W., Eds.; Springer: Cham, Switzerland, 2018; pp. 201–206. [Google Scholar] [CrossRef]
- Woon, J.S.; Boyle, M.J.W.; Ewers, R.M.; Chung, A.; Eggleton, P. Termite environmental tolerances are more linked to desiccation than temperature in modified tropical forests. Insectes Sociaux 2019, 66, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Bignell, D.; Eggleton, P. Termites in ecosystems. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academy: Amsterdam, The Netherlands, 2000; pp. 363–387. [Google Scholar]
- Roose-Amsaleg, C.; Brygoo, Y.; Harry, M. Ascomycete diversity in soil-feeding termite nests and soils from a tropical rainforest. Environ. Microbiol. 2004, 6, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Rosengaus, R.B.; Moustakas-Verho, J.E.; Calleri, D.V.; Traniello, J.F.A. Nesting ecology and cuticular microbial loads in dampwood (Zootermopsis angusticollis) and drywood termites (Incisitermes minor, I. schwarzi, Cryptotermes cavifrons). J. Insect Sci. 2003, 3, 1–6. [Google Scholar] [CrossRef]
- Korb, J.; Linsenmair, K.E. Thermoregulation of termite mounds: What role does ambient temperature and metabolism of the colony play? Insectes Sociaux 2000, 47, 357–363. [Google Scholar] [CrossRef]
- Stien, D. Marine Microbial Diversity as a Source of Bioactive Natural Products. Mar. Drugs 2020, 18, 215. [Google Scholar] [CrossRef]
- Ióca, L.P.; Allard, P.-M.; Berlinck, R.G.S. Thinking big about small beings–The (yet) underdeveloped microbial natural products chemistry in Brazil. Nat. Prod. Rep. 2014, 31, 646–675. [Google Scholar] [CrossRef]
- Nirma, C.; Eparvier, V.; Stien, D. Antifungal Agents from Pseudallescheria boydii SNB-CN73 Isolated from a Nasutitermes sp. Termite. J. Nat. Prod. 2013, 76, 988–991. [Google Scholar] [CrossRef]
- Sorres, J.; Nirma, C.; Barthélemy, M.; Eparvier, V.; Stien, D. Tyroscherin and tyroscherin analogs from Pseudallescheria boydii SNB-CN85 isolated from termite Termes cf. hispaniolae. Phytochem. Lett. 2017, 22, 142–144. [Google Scholar] [CrossRef]
- Sorres, J.; Nirma, C.; Eparvier, V.; Stien, D. Pseudallicins A–D: Four Complex Ovalicin Derivatives from Pseudallescheria boydii SNB-CN85. Org. Lett. 2017, 19, 3978–3981. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Jiang, N.; Han, W.B.; Mei, Y.N.; Ge, H.M.; Guo, Z.K.; Weng, N.S.; Tan, R.X. Antibacterial epipolythiodioxopiperazine and unprecedented sesquiterpene from Pseudallescheria boydii, a beetle (coleoptera)-associated fungus. Org. Biomol. Chem. 2014, 12, 9405–9412. [Google Scholar] [CrossRef] [PubMed]
- Pavlaskova, K.; Nedved, J.; Kuzma, M.; Zabka, M.; Sulc, M.; Sklenar, J.; Novak, P.; Benada, O.; Kofronova, O.; Hajduch, M.; et al. Characterization of Pseudacyclins A−E, a Suite of Cyclic Peptides Produced by Pseudallescheria boydii. J. Nat. Prod. 2010, 73, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kim, S.-K.; Nam, K.W.; Kang, J.S.; Choi, H.D.; Son, B.W. A New Antibacterial Dioxopiperazine Alkaloid Related to Gliotoxin from a Marine Isolate of the Fungus Pseudallescheria. J. Antibiot. 2006, 59, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Bibashi, E.; de Hoog, G.S.; Kostopoulou, E.; Tsivitanidou, M.; Sevastidou, J.; Geleris, P. Invasive infection caused by Pseudallescheria boydii in an immunocompetent patient. Hippokratia 2009, 13, 184–186. [Google Scholar]
- Maebayashi, Y.; Horie, Y.; Satoh, Y.; Yamazaki, M. Isolation of pseurotin A and a new pyrazine from Pseudallescheria boydii. JSM Mycotoxins 1985, 1985, 33–34. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.-H.; Tsou, Y.-J.; Ju, Y.-M.; Hsieh, H.-M.; Ann, P.-J. Production of a Fungistatic Substance by Pseudallescheria boydii Isolated from Soil Amended with Vegetable Tissues and Its Significance. Mycopathologia 2010, 169, 125–131. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Wolfender, J.-L.; Litaudon, M.; Touboul, D.; Queiroz, E.F. Innovative omics-based approaches for prioritisation and targeted isolation of natural products–New strategies for drug discovery. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching molecular structure databases with tandem mass spectra using CSI:FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Olivon, F.; Allard, P.-M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.-F.; Cachet, X.; et al. Bioactive Natural Products Prioritization Using Massive Multi-Informational Molecular Networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- Olivon, F.; Elie, N.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MetGem Software for the Generation of Molecular Networks Based on the t-SNE Algorithm. Anal. Chem. 2018, 90, 13900–13908. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Boutet-Mercey, S.; Cachet, X.; de La Torre, E.; Laboureur, L.; Gallard, J.-F.; Retailleau, P.; Brunelle, A.; Dorrestein, P.C.; Costa, J.; et al. Environmentally Friendly Procedure Based on Supercritical Fluid Chromatography and Tandem Mass Spectrometry Molecular Networking for the Discovery of Potent Antiviral Compounds from Euphorbia semiperfoliata. J. Nat. Prod. 2017, 80, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Vrkoslav, V.; Cvačka, J. Identification of the double-bond position in fatty acid methyl esters by liquid chromatography/atmospheric pressure chemical ionisation mass spectrometry. J. Chromatogr. A 2012, 1259, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Barthélemy, M.; Elie, N.; Pellissier, L.; Wolfender, J.-L.; Stien, D.; Touboul, D.; Eparvier, V. Structural Identification of Antibacterial Lipids from Amazonian Palm Tree Endophytes through the Molecular Network Approach. Int. J. Mol. Sci. 2019, 20, 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, D.J.; Gunstone, F.D. The PMR analysis of non-conjugated alkenoic and alkynoic acids and esters. Chem. Phys. Lipids 1975, 15, 53–85. [Google Scholar] [CrossRef]
- Gunstone, F.; Pollard, M.; Scrimgeour, C.; Vedanayagam, H. Fatty acids. Part 50. 13C nuclear magnetic resonance studies of olefinic fatty acids and esters. Chem. Phys. Lipids 1977, 18, 115–129. [Google Scholar] [CrossRef]
- Griffith, E.C.; Su, Z.; Niwayama, S.; Ramsay, C.A.; Chang, Y.-H.; Liu, J.O. Molecular recognition of angiogenesis inhibitors fumagillin and ovalicin by methionine aminopeptidase 2. Proc. Natl. Acad. Sci. USA 1998, 95, 15183–15188. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Moreno, G.; Cantizani, J.; Sánchez-Carrasco, P.; Ruiz-Perez, L.M.; Martín, J.; El Aouad, N.; Pérez-Victoria, I.; Tormo, J.; Gonzalez-Menendez, V.; González, I.; et al. Discovery of New Compounds Active against Plasmodium falciparum by High Throughput Screening of Microbial Natural Products. PLoS ONE 2016, 11, e0145812. [Google Scholar] [CrossRef]
- Ingber, D.; Fujita, T.; Kishimoto, S.; Sudo, K.; Kanamaru, T.; Brem, H.; Folkman, J. Synthetic analogues of fumagillin that inhibit angiogenesis and suppress tumour growth. Nature 1990, 348, 555–557. [Google Scholar] [CrossRef]
- Corey, E.J.; Guzman-Perez, A.; Noe, M.C. Short Enantioselective Synthesis of (−)−Ovalicin, a Potent Inhibitor of Angiogenesis, Using Substrate-Enhanced Catalytic Asymmetric Dihydroxylation. J. Am. Chem. Soc. 1994, 116, 12109–12110. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, S.; Tsunematsu, Y.; Sato, M.; Watanabe, K. Elucidation of Biosynthetic Pathways of Natural Products. Chem. Rec. 2017, 17, 1095–1108. [Google Scholar] [CrossRef]
- Cane, D.E.; McIlwaine, D.B. The biosynthesis of ovalicin from β-trans-bergamotene. Tetrahedron Lett. 1987, 28, 6545–6548. [Google Scholar] [CrossRef]
- Lin, H.-C.; Chooi, Y.-H.; Dhingra, S.; Xu, W.; Calvo, A.M.; Tang, Y. The Fumagillin Biosynthetic Gene Cluster in Aspergillus fumigatus Encodes a Cryptic Terpene Cyclase Involved in the Formation of β-trans-Bergamotene. J. Am. Chem. Soc. 2013, 135, 4616–4619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Shaw, S.; Kautsar, S.A.; Medema, M.H.; Weber, T. The antiSMASH database version 3: Increased taxonomic coverage and new query features for modular enzymes. Nucleic Acids Res. 2021, 49, D639–D643. [Google Scholar] [CrossRef]
- Wiemann, P.; Guo, C.-J.; Palmer, J.M.; Sekonyela, R.; Wang, C.C.C.; Keller, N.P. Prototype of an intertwined secondary-metabolite supercluster. Proc. Natl. Acad. Sci. USA 2013, 110, 17065–17070. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Vrkoslav, V.; Háková, M.; Pecková, K.; Urbanová, K.; Cvačka, J. Localization of double bonds in wax esters by high-performance liquid chromatography/atmospheric pressure chemical ionization mass utilizing the fragmentation of acetonitrile-related adducts. Anal. Chem. 2011, 83, 2978–2986. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Fauchery, L.; Uroz, S.; Buee, M.; Kohler, A. Purification of Fungal High Molecular Weight Genomic DNA from Environmental Samples. In Fungal Genomics; de Vries, R.P., Tsang, A., Grigoriev, I.V., Eds.; Springer: New York, NY, USA, 2018; Volume 1775, pp. 21–35. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeast, 3rd ed.; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Thévenin, M.; Mouray, E.; Grellier, P.; Dubois, J. Facile Formation of Methylenebis(chalcone)s through Unprecedented Methylenation Reaction. Application to Antiparasitic and Natural Product Synthesis. Eur. J. Org. Chem. 2014, 2014, 2986–2992. [Google Scholar] [CrossRef]
- Kavvadias, D.; Sand, P.; Youdim, K.A.; Qaiser, M.Z.; Rice-Evans, C.; Baur, R.; Sigel, E.; Rausch, W.-D.; Riederer, P.; Schreier, P. The flavone hispidulin, a benzodiazepine receptor ligand with positive allosteric properties, traverses the blood-brain barrier and exhibits anticonvulsive effects. Br. J. Pharmacol. 2004, 142, 811–820. [Google Scholar] [CrossRef]
- Bosc, D.; Lethu, S.; Mouray, E.; Grellier, P.; Dubois, J. Improvement of the trypanocidal activity of 3-arylthiophene farnesyltransferase inhibitors by modulation of their 3-aryl group. Med. Chem. Commun. 2012, 3, 1512–1517. [Google Scholar] [CrossRef]
- Takamatsu, S.; Kim, Y.P.; Komiya, T.; Sunazuka, T.; Hayashi, M.; Tanaka, H.; Komiyama, K.; Omura, S. Chlovalicin, a new cytocidal antibiotic produced by Sporothrix sp. FO-4649. II. Physicochemical properties and structural elucidation. J. Antibiot. 1996, 49, 635–638. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | 2 | 3 | 4 | 5 | 6 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Pos. | δc, Type | δH (J in Hz) | δc | δH | δc | δH | δH | δc | δH |
| 1 | 211.1, C | 211.1, C | 211.2, C | 211.6, C | |||||
| 2 | 36.5, CH2 | 2.74, m | 36.5, CH2 | 2.74, bsex (7.3) | 36.5, CH2 | 2.74, td (13.7; 7.6) | 2.74, m | 36.5, CH2 | 2.76, m |
| 2.22, m | 2.22, m | 2.23, m | 2.22, m | 2.29, m | |||||
| 3 | 32.8, CH2 | 2.10, dd (13.6, 5.1) | 32.7, CH2 | 2.09, m | 32.8, CH2 | 2.09, td (13.6; 7.1) | 2.09, m | 32.8, CH2 | 2.12, m |
| 2.01, ddd (13.6, 7.1, 2.0) | 2.01, m | 2.01, ddd (13.6; 7.1; 1.9) | 2.01, m | 2.04, m | |||||
| 4 | 76.5, C | 76.5, C | 76.5 *, C | 76.3 *, C | |||||
| 5 | 82.8, C | 82.7, C | 82.8, C | 81.8 *, C | |||||
| 6 | 62.8, C | 62.7 *, C | 62.7 *, C | 62.3 *, C | |||||
| 7 | 86.2, CH | 4.69, s | 86.2, CH | 4.69, s | 86.2, CH | 4.69, s | 4.69, s | 77.5, CH | 5.02, s |
| 8 | 58.4, CH | 2.97, t (6.5) | 58.3, CH | 2.97, t (6.5) | 58.4, CH | 2.97, t (6.5) | 2.97, t (6.5) | 57.9 *, CH | 2.96, t (6.8) |
| 9 | 28.4, CH2 | 2.38, m | 28.4, CH2 | 2.37, m | 28.4, CH2 | 2.38, m | 2.37, m | 28.4, CH2 | 2.38, m |
| 2.26, m | 2.27, m | 2.25, m | 2.26, m | 2.26, m | |||||
| 10 | 119.8, CH | 5.28, m | 119.8, CH | 5.27, m | 119.8, CH | 5.27, m | 5.27, m | 119.9, CH | 5.28, m |
| 11 | 136.1, C | 136.1, C | 136.1, C | 136.0, C | |||||
| 12 | 26.1, CH3 | 1.72, brd (1.2) | 26.1, CH3 | 1.73, bd (1.1) | 26.0, CH3 | 1.73, bs | 1.73, bd (1.2) | 26.1, CH3 | 1.72, bs |
| 13 | 18.2, CH3 | 1.68, brd (0.9) | 18.2, CH3 | 1.68, bd (0.8) | 18.2, CH3 | 1.68, bs | 1.68, bd (0.8) | 18.2, CH3 | 1.68, bs |
| 14 | 69.4, CH2 | 4.17, d (11.2) | 69.4, CH2 | 4.17, d (11.1) | 69.4, CH2 | 4.17, d (11.1) | 4.17, d (11.1) | 69.5, CH2 | 4.18, d (11.1) |
| 4.11, d (11.2) | 4.11, d (11.1) | 4.11, d (11.1) | 4.11, d (11.1) | 4.12, d (11.1) | |||||
| 15 | 16.3, CH3 | 1.54, s | 16.3, CH3 | 1.55, s | 16.3, CH3 | 1.54, s | 1.55, s | 16.3, CH3 | 1.61, s |
| OMe | 59.7, CH3 | 3.49, s | 59.7, CH3 | 3.49, s | 59.7, CH3 | 3.48, s | 3.49, s | 175.2, C | |
| 1′ | 175.5, C | 175.4, C | 175.5, C | 35.1, CH2 | 2.38, m | ||||
| 2′ | 35.1, CH2 | 2.38, t (7.4) | 35.1, CH2 | 2.38, m | 35.1, CH2 | 2.38, m | 2.38, m | 26.2, CH2 | 1.64, m |
| 3′ | 26.2, CH2 | 1.64, m | 26.1, CH2 | 1.64, m | 26.4, CH2 | 1.25–1.37, m | 1.64, m | 30.2–30.8, CH2 | 1.27–1.40, m |
| 4′–7′ | 30.3−30.8, CH2 | 1.28, m–1.40, m | 30.2–31.0, CH2 | 1.26–1.40, m | 30.3–31.0, CH2 | 1.25–1.37, m | 1.25–1.40, m | 30.4, CH2 | 1.37, m |
| 8′ | 28.3, CH2 | 2.07, m | 28.3, CH2 | 2.03, m | 33.2, CH2 | 1.25–1.37, m | 2.03, m | 28.4, CH2 | 2.07, m |
| 9′ | 131.0, CH | 5.36, m | 130.9, CH | 5.35, m | 23.9, CH2 | 1.25–1.37, m | 5.35, m | 131.0, CH | 5.36, m |
| 10′ | 129.3, CH | 5.33, m | 131.1, CH | 5.35, m | 14.6, CH3 | 1.25–1.37, m | 5.35, m | 129.3, CH | 5.33, m |
| 11′ | 26.7, CH2 | 2.78, m | 28.3, CH2 | 2.03, m | 211.2, C | 1.25–1.37, m | 2.03, m | 26.7, CH2 | 2.77, m |
| 12′ | 129.2, CH | 5.33, m | 30.2–31.0, CH2 | 1.26–1.40, m | 36.5, CH2 | 1.25–1.37, m | 1.26–1.40, m | 129.2, CH | 5.33, m |
| 13′ | 131.1, CH | 5.36, m | 30.2–1.0, CH2 | 1.26–1.40, m | 1.25–1.37, m | 1.30, m | 131.1, CH | 5.36, m | |
| 14′ | 28.3, CH2 | 2.07, m | 30.2–31.0, CH2 | 1.26–1.40, m | 32.8, CH2 | 1.25–1.37, m | 1.29, m | 28.4, CH2 | 2.07, m |
| 15′ | 30.3–30.8, CH2 | 1.28, m–1.40, m | 30.2–31.0, CH2 | 1.26–1.40, m | 1.25–1.37, m | 0.90, t (7.0) | 30.4, CH2 | 1.37, m | |
| 16′ | 32.8, CH2 | 1.31, m | 33.2, CH2 | 1.30, m | 76.5 *, C | 1.29, m | 32.8, CH2 | 1.31, m | |
| 17′ | 23.8, CH2 | 1.34, m | 23.9, CH2 | 1.29, m | 82.8, C | 1.29, m | 23.8, CH2 | 1.33, m | |
| 18′ | 14.6, CH3 | 0.91, t (7.0) | 14.6, CH3 | 0.90, t (7.0) | 62.7 *, C | 0.90, t (7.0) | 14.6 | 0.91, t (7.2) | |
| Cmpd. | P. falciparum (IC50 µM) | T. brucei brucei (IC50 µM) | HUVECs (IC50 µM) |
|---|---|---|---|
| 1 | 13.6 ± 6.4 | 0.41 ± 0.17 | 13 ± 2 |
| 2 | 19.8 ± 6.7 | 1.1 ± 0.5 | 37 ± 3 |
| 3 | >50 | 4.8 ± 2.7 | 45 ± 2 |
| 4 | 20.6 ± 4.6 | 4.1 ± 2.1 | 70 ± 1 |
| Position | δC, Type | δH (J in Hz) |
|---|---|---|
| 1 | 178.0, C | |
| 2 | 29.3, CH2 | 2.22 |
| 3 | 25.6, CH2 | 2.27, m |
| 2.05, m | ||
| 4 | 61.5, C | |
| 5 | 76.0, C | |
| 6 | 63.4, C | |
| 7 | 63.6, CH2 | 3.64, d (11.6) |
| 3.60, d (11.6) | ||
| 8 | 58.3, CH | 3.26, t (6.5) |
| 9 | 28.0, CH2 | 2.36, m |
| 2.22, m | ||
| 10 | 119.8, C | 5.24, m |
| 11 | 135.4, C | |
| 12 | 25.8, CH3 | 1.67, s |
| 13 | 17.9, CH3 | 1.73, s |
| 14 | 48.8 | 3.00, d (3.9) |
| 2.68, d (3.9) | ||
| 15 | 17.6, CH3 | 1.37, s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorres, J.; Hebra, T.; Elie, N.; Leman-Loubière, C.; Grayfer, T.; Grellier, P.; Touboul, D.; Stien, D.; Eparvier, V. Antiparasitic Ovalicin Derivatives from Pseudallescheria boydii, a Mutualistic Fungus of French Guiana Termites. Molecules 2022, 27, 1182. https://doi.org/10.3390/molecules27041182
Sorres J, Hebra T, Elie N, Leman-Loubière C, Grayfer T, Grellier P, Touboul D, Stien D, Eparvier V. Antiparasitic Ovalicin Derivatives from Pseudallescheria boydii, a Mutualistic Fungus of French Guiana Termites. Molecules. 2022; 27(4):1182. https://doi.org/10.3390/molecules27041182
Chicago/Turabian StyleSorres, Jonathan, Téo Hebra, Nicolas Elie, Charlotte Leman-Loubière, Tatyana Grayfer, Philippe Grellier, David Touboul, Didier Stien, and Véronique Eparvier. 2022. "Antiparasitic Ovalicin Derivatives from Pseudallescheria boydii, a Mutualistic Fungus of French Guiana Termites" Molecules 27, no. 4: 1182. https://doi.org/10.3390/molecules27041182