
Elucidation of the Metabolite Profile of Yucca gigantea and Assessment of Its Cytotoxic, Antimicrobial, and Anti-Inflammatory Activities

 ,
,  ,
,  , , and
, , and 
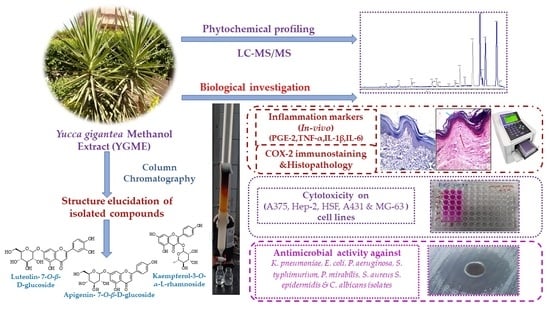
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Extraction, and Isolation of Pure Compounds
2.2. Animals
2.3. Materials, Drugs and Chemicals
2.4. LC-MS/MS for Metabolite Analysis
2.4.1. Sample Preparations
2.4.2. Acquisition Method and Analytical Parameters
2.4.3. Data Processing
2.5. Cell Lines
2.6. In Vitro Cytotoxicity
2.7. In Vitro Antimicrobial Activity
2.7.1. Agar Well Diffusion Method
2.7.2. Determination of MIC Values
2.8. In Vivo Anti-Inflammatory Activity
2.8.1. Induction and Assessment of Carrageenan-Induced Paw Edema
2.8.2. Sample Collection
2.8.3. Determination of Paw GSH Concentration
2.8.4. Determination of Paw NO Content
2.8.5. Determination of Paw MPO Activity
2.8.6. Determination of the Levels of Inflammatory Markers
2.8.7. Histopathologic Examination of the Paw Tissue
2.8.8. Immunohistochemical Determination of COX-2 Expression
2.9. Statistical Analysis
3. Results
3.1. LC-ESI-MS/MS Analysis of YGME
3.1.1. Characterization of Flavonols and Flavonols Glycosides
3.1.2. Characterization of Hydroxylated and/or Methoxylated Flavonols and Flavonols Glycosides
3.1.3. Characterization of Flavones and Flavones Glycosides
3.1.4. Characterization of Flavanones and Flavanone Glycosides
3.1.5. Characterization of Saponins
3.1.6. Characterization of Organic and Phenolic Acids
3.1.7. Characterization of Other Compounds
3.2. Structure Elucidation of Compounds Isolated from YGME
3.3. In Vitro Cytotoxic Activity of YGME
3.4. In Vitro Antimicrobial Activity of YGME
3.5. In Vivo Anti-Inflammatory Activity
3.5.1. Effect of YGME on the Average Edema Volume
3.5.2. Effect of YGME on the Average Paw Edema Weight
3.5.3. Effect of YGME on MPO Activity
3.5.4. Effect of YGME on the Oxidative Stress Markers
3.5.5. Effect of YGME on the Levels of the Inflammation Markers
3.5.6. Effect of YGME on Paw PGE-2 Levels
3.5.7. Effect of YGME on the Histopathology of Paw Tissues
3.5.8. Effects of YGME on the Immunohistochemical Staining of COX-2 in Paw Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chase, M.W.; Reveal, J.L.; Fay, M.F. A subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Bot. J. Linn. Soc. 2009, 161, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Smith, C. Yuccas: Giants among the Lilies; National Council for the Conservation of Plants & Gardens: Guildford, UK, 2004. [Google Scholar]
- Smith, G.F.; Figueiredo, E. Widely cultivated, large-growing yuccas: Notes on Yucca elephantipes Regel ex Trel., Yucca gigantea Lem., and Yucca guatemalensis Baker (Agavaceae). Bradleya 2016, 2016, 28–37. [Google Scholar] [CrossRef]
- Patel, S. Yucca: A medicinally significant genus with manifold therapeutic attributes. Nat. Prod. Bioprospect. 2012, 2, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Rasool, N.; Mansha, A.; Anjum, F.; Iqbal, M.; Mushtaq, M.; Shahid, M. Antioxidant, antibacterial, antifungal activities and phytochemical analysis of dagger (Yucca aloifolia) leaves extracts. J. Med. Plants Res. 2013, 7, 243–249. [Google Scholar]
- Pascual-Mendoza, S.; Saynes-Vásquez, A.; Pérez-Herrera, A. Traditional knowledge of edible plants in an indigenous community in the Sierra Norte of Oaxaca, Mexico. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 1–13. [Google Scholar] [CrossRef]
- El Sayed, A.M.; Basam, S.M.; Marzouk, H.S.; El-Hawary, S. LC–MS/MS and GC–MS profiling as well as the antimicrobial effect of leaves of selected Yucca species introduced to Egypt. Sci. Rep. 2020, 10, 17778. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, C.R.; Zhang, Y.J. New Steroidal Saponins from the Leaves of Yucca elephantipes. Helv. Chim. Acta 2013, 96, 1807–1813. [Google Scholar] [CrossRef]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Ojha, S.; Patil, C.R. Animal models of inflammation for screening of anti-inflammatory drugs: Implications for the discovery and development of phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef] [Green Version]
- Karim, N.; Khan, I.; Khan, W.; Khan, I.; Khan, A.; Halim, S.A.; Khan, H.; Hussain, J.; Al-Harrasi, A. Anti-nociceptive and anti-inflammatory activities of asparacosin a involve selective cyclooxygenase 2 and inflammatory cytokines inhibition: An in-vitro, in-vivo, and in-silico approach. Front. Immunol. 2019, 10, 581. [Google Scholar] [CrossRef]
- Zhang, H.; Shang, C.; Tian, Z.; Amin, H.K.; Kassab, R.B.; Abdel Moneim, A.E.; Zhang, Y. Diallyl disulfide suppresses inflammatory and oxidative machineries following carrageenan injection-induced paw edema in mice. Mediat. Inflamm. 2020, 2020, 8508906. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.B.; Kwon, Y.-G.; Simu, S.Y.; Abrar Shahriyar, S.; Lee, S.H. Attenuation of Inflammatory Symptoms by Icariside B2 in Carrageenan and LPS-Induced Inflammation Models via Regulation of MAPK/NF-κB Signaling Cascades. Biomolecules 2020, 10, 1037. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Im, H.-W.; Karmacharya, M.B.; Kim, S.; Min, B.-H.; Park, S.R.; Choi, B.H. Low-intensity ultrasound attenuates paw edema formation and decreases vascular permeability induced by carrageenan injection in rats. J. Inflamm. 2020, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G. Cashew (Anacardium occidentale L.) nuts counteract oxidative stress and inflammation in an acute experimental model of Carrageenan-induced Paw edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkhel, S. Evaluation of the anti-inflammatory activities of Quillaja saponaria Mol. saponin extract in mice. Toxicol. Rep. 2016, 3, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Song, X.; Jia, R.; Yin, Z.; Zou, Y.; Li, L.; Yin, L.; He, C.; Liang, X.; Yue, G. Evaluation of analgesic and anti-inflammatory activities of water extract of Galla chinensis in vivo models. Evid.-Based Complement. Altern. Med. 2018, 2018, 6784032. [Google Scholar] [CrossRef] [Green Version]
- El Hawary, S.; El Sayed, A.; Helmy, M.W.; El Naggar, E.M.B.; Marzouk, H.S.; Bassam, S.M. DNA fingerprinting, biological and chemical investigation of certain Yucca species. Nat. Prod. Res. 2018, 32, 2617–2620. [Google Scholar] [CrossRef]
- GARCÍA-BORES, A.M.; Bello, C.; Campos, Y.; del Carmen Benitez, J.; Flores, S.; Canales, M.; Hernandez, T.; ACEVEDO, J.G.A. Photoprotective activity of Yucca periculosa polyphenols. Bol. Latinoam. Caribe Plantas Med. Aromát. 2010, 9, 100–108. [Google Scholar]
- Attallah, N.G.M.; Negm, W.A.; Elekhnawy, E.; Elmongy, E.I.; Altwaijry, N.; El-Haroun, H.; El-Masry, T.A.; El-Sherbeni, S.A. Elucidation of Phytochemical Content of Cupressus macrocarpa Leaves: In Vitro and In Vivo Antibacterial Effect against Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Antibiotics 2021, 10, 890. [Google Scholar] [CrossRef]
- Negm, W.A.; El-Kadem, A.H.; Elekhnawy, E.; Attallah, N.G.; Al-Hamoud, G.A.; El-Masry, T.A.; Zayed, A. Wound-Healing Potential of Rhoifolin-Rich Fraction Isolated from Sanguisorba officinalis Roots Supported by Enhancing Re-Epithelization, Angiogenesis, Anti-Inflammatory, and Antimicrobial Effects. Pharmaceuticals 2022, 15, 178. [Google Scholar] [CrossRef]
- Alotaibi, B.; Mokhtar, F.A.; El-Masry, T.A.; Elekhnawy, E.; Mostafa, S.A.; Abdelkader, D.H.; Elharty, M.E.; Saleh, A.; Negm, W.A. Antimicrobial Activity of Brassica rapa L. Flowers Extract on Gastrointestinal Tract Infections and Antiulcer Potential Against Indomethacin-Induced Gastric Ulcer in Rats Supported by Metabolomics Profiling. J. Inflamm. Res. 2021, 14, 7411. [Google Scholar] [CrossRef]
- Attallah, N.G.; Negm, W.A.; Elekhnawy, E.; Altwaijry, N.; Elmongy, E.I.; El-Masry, T.A.; Alturki, E.A.; Yousef, D.A.; Y Shoukheba, M. Antibacterial Activity of Boswellia sacra Flueck. Oleoresin Extract against Porphyromonas gingivalis Periodontal Pathogen. Antibiotics 2021, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; 20th Informational Supplement; CLSI Document M100; CLSI: Wayne, PA, USA, 2017; pp. 1–50. [Google Scholar]
- Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Elekhnawy, E. Investigation of the Antibacterial Activity and Efflux Pump Inhibitory Effect of Cycas thouarsii R. Br. Extract against Klebsiella pneumoniae Clinical Isolates. Pharmaceuticals 2021, 14, 756. [Google Scholar] [CrossRef] [PubMed]
- Khadka, S.; Sherchand, J.B.; Pokhrel, B.M.; Parajuli, K.; Mishra, S.K.; Sharma, S.; Shah, N.; Kattel, H.P.; Dhital, S.; Khatiwada, S. Isolation, speciation and antifungal susceptibility testing of Candida isolates from various clinical specimens at a tertiary care hospital, Nepal. BMC Res. Notes 2017, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, M.P. Performance Standards for Antimicrobial Disk Susceptibility Tests. National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2018. [Google Scholar]
- Ince, S.; Kucukkurt, I.; Turkmen, R.; Demirel, H.H.; Sever, E. Dietary Yucca schidigera supplementation reduces arsenic-induced oxidative stress in Swiss albino mice. Toxicol. Ind. Health 2013, 29, 904–914. [Google Scholar] [CrossRef]
- Coura, C.O.; Souza, R.B.; Rodrigues, J.A.G.; Vanderlei, E.d.S.O.; de Araújo, I.W.F.; Ribeiro, N.A.; Frota, A.F.; Ribeiro, K.A.; Chaves, H.V.; Pereira, K.M.A. Mechanisms involved in the anti-inflammatory action of a polysulfated fraction from Gracilaria cornea in rats. PLoS ONE 2015, 10, e0119319. [Google Scholar]
- Amdekar, S.; Roy, P.; Singh, V.; Kumar, A.; Singh, R.; Sharma, P. Anti-inflammatory activity of lactobacillus on carrageenan-induced paw edema in male wistar rats. Int. J. Inflamm. 2012, 2012, 752015. [Google Scholar] [CrossRef] [Green Version]
- El-Shitany, N.A.; El-Bastawissy, E.A.; El-desoky, K. Ellagic acid protects against carrageenan-induced acute inflammation through inhibition of nuclear factor kappa B, inducible cyclooxygenase and proinflammatory cytokines and enhancement of interleukin-10 via an antioxidant mechanism. Int. Immunopharmacol. 2014, 19, 290–299. [Google Scholar] [CrossRef]
- El-Mahdy, N.A.; El-Sayad, M.E.S.; El-Kadem, A.H.; Abu-Risha, S.E.S. Targeting IL-10, ZO-1 gene expression and IL-6/STAT-3 trans-signaling by a combination of atorvastatin and mesalazine to enhance anti-inflammatory effects and attenuates progression of oxazolone-induced colitis. Fundam. Clin. Pharmacol. 2021, 35, 143–155. [Google Scholar] [CrossRef]
- El-Mahdy, N.A.; El-Sayad, M.E.-S.; El-Kadem, A.H. Combination of telmisartan with sildenafil ameliorate progression of diabetic nephropathy in streptozotocin-induced diabetic model. Biomed. Pharmacother. 2016, 81, 136–144. [Google Scholar] [CrossRef]
- El-Mahdy, N.A.; El-Sayad, M.E.-S.; El-Kadem, A.H.; Abu-Risha, S.E.-S. Metformin alleviates inflammation in oxazolone induced ulcerative colitis in rats: Plausible role of sphingosine kinase 1/sphingosine 1 phosphate signaling pathway. Immunopharmacol. Immunotoxicol. 2021, 43, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wang, J.; Ruan, J.; Yao, X.; Huang, P.; Wang, Y.; Yu, H.; Han, L.; Zhang, Y.; Wang, T. Spirostane-type saponins obtained from Yucca schidigera. Molecules 2018, 23, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usia, T.; Iwata, H.; Hiratsuka, A.; Watabe, T.; Kadota, S.; Tezuka, Y. Sesquiterpenes and Flavonol Glycosides from Zingiber a romaticum and Their CYP3A4 and CYP2D6 Inhibitory Activities. J. Nat. Prod. 2004, 67, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Mabry, T.; Markham, K.R.; Thomas, M.B. The Systematic Identification of Flavonoids; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Chiruvella, K.K.; Mohammed, A.; Dampuri, G.; Ghanta, R.G.; Raghavan, S.C. Phytochemical and antimicrobial studies of methyl angolensate and luteolin-7-O-glucoside isolated from callus cultures of Soymida febrifuga. Int. J. Biomed. Sci. IJBS 2007, 3, 269. [Google Scholar]
- SHEN, C.-C.; CHANG, Y.-S.; HO, L.-K. Nuclear-magnetic-resonance studies of 5,7-dihydroxyflavonoids. Phytochemistry 1993, 34, 843–845. [Google Scholar] [CrossRef]
- Utari, F.; Itam, A.; Syafrizayanti, S.; Putri, W.H.; Ninomiya, M.; Koketsu, M.; Tanaka, K.; Efdi, M. Isolation of flavonol rhamnosides from Pometia pinnata leaves and investigation of α-glucosidase inhibitory activity of flavonol derivatives. J. Appl. Pharm. Sci. 2019, 9, 53–65. [Google Scholar]
- Mahjbeen, S.; Amjad, Y.; Iqbal, K.; Hamid, I. Antibacterial and anti-oxidant potential of Yucca elephantipes and comparative safety profile of its various extracts. J. Toxicol. Pharm. Sci. 2018, 2, 28–36. [Google Scholar]
- Boominathan, R.; Parimaladevi, B.; Mandal, S.; Ghoshal, S. Anti-inflammatory evaluation of Ionidium suffruticosam Ging. in rats. J. Ethnopharmacol. 2004, 91, 367–370. [Google Scholar] [CrossRef]
- Panthong, A.; Norkaew, P.; Kanjanapothi, D.; Taesotikul, T.; Anantachoke, N.; Reutrakul, V. Anti-inflammatory, analgesic and antipyretic activities of the extract of gamboge from Garcinia hanburyi Hook f. J. Ethnopharmacol. 2007, 111, 335–340. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; El-Kholy, M.S. Productive performance, egg quality, blood constituents, immune functions, and antioxidant parameters in laying hens fed diets with different levels of Yucca schidigera extract. Environ. Sci. Pollut. Res. 2016, 23, 6774–6782. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Abd El-Hack, M.E.; El-Sayed, S.A.; Ahmed, S.Y.; Samak, D.H. Yucca schidigera extract modulates the lead-induced oxidative damage, nephropathy and altered inflammatory response and glucose homeostasis in Japanese quails. Ecotoxicol. Environ. Saf. 2018, 156, 311–321. [Google Scholar] [CrossRef]
- Cheeke, P.; Piacente, S.; Oleszek, W. Anti-inflammatory and anti-arthritic effects of Yucca schidigera: A review. J. Inflamm. 2006, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzocco, S.; Piacente, S.; Pizza, C.; Oleszek, W.; Stochmal, A.; Pinto, A.; Sorrentino, R.; Autore, G. Inhibition of inducible nitric oxide synthase expression by yuccaol C from Yucca schidigera roezl. Life Sci. 2004, 75, 1491–1501. [Google Scholar] [CrossRef]
- Abdelwahab, S.I.; Hassan, L.E.A.; Sirat, H.M.; Yagi, S.M.A.; Koko, W.S.; Mohan, S.; Taha, M.M.E.; Ahmad, S.; Chuen, C.S.; Narrima, P. Anti-inflammatory activities of cucurbitacin E isolated from Citrullus lanatus var. citroides: Role of reactive nitrogen species and cyclooxygenase enzyme inhibition. Fitoterapia 2011, 82, 1190–1197. [Google Scholar] [CrossRef]
- Kiraly, A.J.; Soliman, E.; Jenkins, A.; Van Dross, R.T. Apigenin inhibits COX-2, PGE2, and EP1 and also initiates terminal differentiation in the epidermis of tumor bearing mice. Prostaglandins Leukot. Essent. Fat. Acids 2016, 104, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Okla, M.; Al Madani, J.O.; Chung, S.; Alfayez, M. Apigenin Reverses Interleukin-1β-Induced Suppression of Adipocyte Browning via COX2/PGE2 Signaling Pathway in Human Adipocytes. Mol. Nutr. Food Res. 2020, 64, 1900925. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.-Q.; Zhang, Q.; Zhao, X.-H.; Shi, J. The In Vitro Anti-Inflammatory Activities of Galangin and Quercetin towards the LPS-Injured Rat Intestinal Epithelial (IEC-6) Cells as Affected by Heat Treatment. Molecules 2021, 26, 7495. [Google Scholar] [CrossRef]
- Ko, H.-H.; Weng, J.-R.; Tsao, L.-T.; Yen, M.-H.; Wang, J.-P.; Lin, C.-N. Anti-inflammatory flavonoids and pterocarpanoid from Crotalaria pallida and C. assamica. Bioorg. Med. Chem. Lett. 2004, 14, 1011–1014. [Google Scholar] [CrossRef]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Wölfle, U. Luteolin as a modulator of skin aging and inflammation. Biofactors 2021, 47, 170–180. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef]
- Sato, S.; Mukai, Y. Modulation of chronic inflammation by quercetin: The beneficial effects on obesity. J. Inflamm. Res. 2020, 13, 421. [Google Scholar] [CrossRef] [PubMed]
- Saeedi-Boroujeni, A.; Mahmoudian-Sani, M.-R. Anti-inflammatory potential of Quercetin in COVID-19 treatment. J. Inflamm. 2021, 18, 3. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, A.; Caporali, S.; Daniele, N.D.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S. Anti-inflammatory and proliferative properties of luteolin-7-O-glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef] [PubMed]
- Ruangnoo, S.; Jaiaree, N.; Makchuchit, S.; Panthong, S.; Thongdeeying, P.; Itharat, A. An in vitro inhibitory effect on RAW 264.7 cells by anti-inflammatory compounds from Smilax corbularia Kunth. Asian Pac. J. Allergy Immunol. 2012, 30, 268. [Google Scholar]
- Scuro, L.S.; Simioni, P.; Grabriel, D.; Saviani, E.E.; Modolo, L.V.; Tamashiro, W.M.; Salgado, I. Suppression of nitric oxide production in mouse macrophages by soybean flavonoids accumulated in response to nitroprusside and fungal elicitation. BMC Biochem. 2004, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Xagorari, A.; Papapetropoulos, A.; Mauromatis, A.; Economou, M.; Fotsis, T.; Roussos, C. Luteolin inhibits an endotoxin-stimulated phosphorylation cascade and proinflammatory cytokine production in macrophages. J. Pharmacol. Exp. Ther. 2001, 296, 181–187. [Google Scholar]
- Li, Y.-C.; Yeh, C.-H.; Yang, M.-L.; Kuan, Y.-H. Luteolin suppresses inflammatory mediator expression by blocking the Akt/NFκB pathway in acute lung injury induced by lipopolysaccharide in mice. Evid.-Based Complement. Altern. Med. 2012, 2012, 383608. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-J.; Chen, Y.-M.; Wang, T.-M.; Lee, C.-K.; Chen, K.-J.; Lee, T.-H. Flavonoids with iNOS inhibitory activity from Pogonatherum crinitum. J. Ethnopharmacol. 2008, 118, 71–78. [Google Scholar] [CrossRef]
- Nwanodi, O. Nutraceuticals: Curative integrative cancer treatment. Altern. Integr. Med. 2017, 6, 1000240. [Google Scholar] [CrossRef]
- Yi, L.; Ma, S.; Ren, D. Phytochemistry and bioactivity of Citrus flavonoids: A focus on antioxidant, anti-inflammatory, anticancer and cardiovascular protection activities. Phytochem. Rev. 2017, 16, 479–511. [Google Scholar] [CrossRef]
- Rais, J.; Jafri, A.; Siddiqui, S.; Tripathi, M.; Arshad, M. Phytochemicals in the treatment of ovarian cancer. Front. Biosci. (Elite Ed.) 2017, 9, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential therapeutic targets of quercetin, a plant flavonol, and its role in the therapy of various types of cancer through the modulation of various cell signaling pathways. Molecules 2021, 26, 1315. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, P.A.; Walle, T. Inhibition of benzo[a]pyrene-activating enzymes and DNA binding in human bronchial epithelial BEAS-2B cells by methoxylated flavonoids. Carcinogenesis 2006, 27, 1579–1585. [Google Scholar] [CrossRef]
- Wen, X.; Walle, U.K.; Walle, T. 5,7-Dimethoxyflavone downregulates CYP1A1 expression and benzo[a]pyrene-induced DNA binding in Hep G2 cells. Carcinogenesis 2005, 26, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Ohkusa, T.; Nomura, T.; Sato, N. The role of bacterial infection in the pathogenesis of inflammatory bowel disease. Intern. Med. 2004, 43, 534–539. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [Green Version]
- El-Zayat, M.M.; Eraqi, M.M.; Alrefai, H.; El-Khateeb, A.Y.; Ibrahim, M.A.; Aljohani, H.M.; Aljohani, M.M.; Elshaer, M.M. The Antimicrobial, Antioxidant, and Anticancer Activity of Greenly Synthesized Selenium and Zinc Composite Nanoparticles Using Ephedra aphylla Extract. Biomolecules 2021, 11, 470. [Google Scholar] [CrossRef]
- Serwecińska, L. Antimicrobials and antibiotic-resistant bacteria: A risk to the environment and to public health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Donninelli, G.; Del Cornò, M.; Pierdominici, M.; Scazzocchio, B.; Varì, R.; Varano, B.; Pacella, I.; Piconese, S.; Barnaba, V.; D’Archivio, M. Distinct blood and visceral adipose tissue regulatory T cell and innate lymphocyte profiles characterize obesity and colorectal cancer. Front. Immunol. 2017, 8, 643. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min.) | [M − H]− m/z | MS2 m/z or Fragments | Formula | Idenification |
|---|---|---|---|---|---|
| 1 | 1.128 | 115.000 | 115.001, 89.023, 71.012 | C4H4O4 | Maleic acid |
| 2 | 1.348 | 449.085 | 449.088, 269.015, 113.019 | C21H22O11 | Okanin-4′-O-glucoside (Marein) |
| 3 | 1.398 | 129.019 | 129.019 | C5H6O4 | Citraconic acid |
| 4 | 1.411 | 141.018 | 141.020, 97.028 | C6H6O4 | Muconic acid |
| 5 | 4.37 | 739.790 | 739.792, 577.415 | C39H61O13 | Spirostan-3-ol-glucoside-galactoside |
| 6 | 4.77 | 429.171 | 429.171 | C27H42O4 | Hecogenin |
| 7 | 4.901 | 593.152 | 593.156, 473.105, 341.111, 285,251 | C27H30O15 | Kaempferol-7-O-neohesperidoside |
| 8 | 5.64 | 431.192 | 431.163, 392.918, 385.182, 341.164, 324.942 | C21H20O10 | Kaempferol-3-O-α-l-rhamnoside |
| 9 | 6.22 | 739.790 | 739.792, 577.415 | C39H61O13 | Spirostan-3-ol-diglucoside |
| 10 | 6.239 | 447.09 | 447.090, 285.214, 248.960 | C27H30O16 | Luteolin-7-O-β-d-glucoside |
| 11 | 6.276 | 577.156 | 577.160, 413.081, 311.048, 293.046 | C27H30O14 | Vitexin-2″-O-rhamnoside |
| 12 | 6.495 | 609.147 | 609.143, 301.023 | C28H34O15 | Hesperetin-7-O-neohesperidoside |
| 13 | 6.659 | 315.135 | 315.197, 287.198 | C16H12O7 | 3′-Methoxy-4′,5,7-trihydroxyflavonol (Isorhamnetin) |
| 14 | 6.817 | 463.085 | 463.089, 354.916, 326.930, 301.041, 286.936 | C21H20O12 | Quercetin-4′-O-glucoside |
| 15 | 6.978 | 623.197 | 623.159, 577.163, 315.049 | C28H32O16 | Isorhamnetin-3-O-rutinoside |
| 16 | 7.093 | 593.519 | 593.149, 285.030, 241.042 | C30H26O13 | Kaempferol-3-O-(6-p-coumaroyl)- glucopyranoside |
| 17 | 7.263 | 593.265 | 593.149, 570.246, 547.242, 285.038 | C28H34O14 | Isosakuranetin-7-O-neohesperidoside (Poncirin) |
| 18 | 8.182 | 285.077 | 285.071, 179.037, 165.016, 119.049 | C15H10O6 | 4′,5,7-Trihydroxyflavonol |
| 19 | 9.458 | 301.069 | 301.073, 273.072, 139.048 | C16H14O6 | Hesperetin |
| 20 | 10.199 | 271.060 | 271.064, 196.049, 165.024, 151.005 | C15H12O5 | Naringenin |
| 21 | 10.488 | 431.17 | 431.170, 269.040, 253.054 | C15H10O5 | Apigenin-7-O-β-d-glucoside |
| 22 | 10.876 | 285.039 | 285.065, 256.039, 179.033, 145.030 | C15H10O6 | Luteolin |
| 23 | 10.992 | 179.033 | 179.037, 135.042 | C9H8O4 | Caffeic acid |
| 24 | 11.313 | 283.060 | 283.064, 211.070, 189.021, 177.014 | C15H12O6 | Acacetin |
| 25 | 11.531 | 315.092 | 315.086, 297.075, 193.009, 179.033, 152.010 | C16H12O7 | 3,3′,4′,5-tetrahydroxy-7-methoxyflavone |
| 26 | 12.594 | 299.093 | 299.090, 193.049, 149.058 | C16H12O6 | 3,5,7-Trihydroxy-4′-methoxyflavone |
| 27 | 13.760 | 461.260 | 461.262 | C27H44O3 | 25 R or S-Spirostanol-3-ol |
| 28 | 14.034 | 223.171 | 223.163, 113.991 | C11H12O5 | 3-(4-Hydroxy-3,5-dimethoxyphenyl)-2-propenoic acid |
| 29 | 19.75 | 277.197 | 277.218, 276.367, 259.204, 233.224, 205.203 | C18H30O2 | gamma-Linolenic acid |
| Pathogenic Bacterial Isolate | Inhibition Zone Diameter (mm) | MIC Values (µg/mL) | |
|---|---|---|---|
| YGME | Chlorhexidine | ||
| Gram-negative bacteria | |||
| Klebsiella pneumoniae | 12.5 ± 0.41 | 27.5 ± 1.35 | 106.67 ± 30.1 |
| Escherichia coli | 13.3 ± 0.65 | 28.5 ± 1.50 | 85.3 ± 30.0 |
| Pseudomonas aeruginosa | 10.2 ± 0.69 | 23.4 ± 0.77 | 42.6 ± 15.0 |
| Proteus mirabilis | 14.8 ± 0.33 | 26.4 ± 0.89 | 74.67 ± 39.9 |
| Salmonella typhimurium | 10.5 ± 0.42 | 23.3 ± 0.85 | 53.3 ± 15.08 |
| Pseudomonas aeruginosa | 10.2 ± 0.69 | 23.4 ± 0.77 | 42.6 ± 15.0 |
| Gram-positive bacteria | |||
| Staphylococcus aureus | 14.46 ± 1.14 | 24.9 ± 1.40 | 85.3 ± 30.0 |
| Staphylococcus epidermidis | 16.5 ±0.75 | 26.6 ± 0.98 | 106.67 ± 30.1 |
| Fungi | |||
| Candida albicans | 12 ± 1.3 | 17.8 ± 2.1 | 21.3 ± 7.5 |
| Time (h) | The Average Change in Edema Volume (mm) * | |||
|---|---|---|---|---|
| Group II | Group III | Group IV | Group V | |
| 1 | 0.36 ± 0.05 | 0.3508 ± 0.031 | 0.32 ± 0.08 | 0.3703 ± 0.012 |
| 2 | 0.62 ± 0.12 | 0.208 ± 0.061 | 0.24 ± 0.031 | 0.2307 ± 0.02 |
| 3 | 0.81 ± 0.04 | 0.1108 ± 0.011 | 0.18 ± 0.04 | 0.1303 ± 0.03 |
| 4 | 1.20 ± 0.11 | 0.01 ± 0.001 | 0.12 ± 0.013 | 0.01 ± 0.001 |
| Average Paw Weight (g) | Paw NO Content (nmol/g Tissue) | Paw GSH Content (µmol/g Tissue) | Paw MPO Activity (µM/min/g Tissue) | |
|---|---|---|---|---|
| Group I | 0.03 ± 0.001 | 10.6 ± 0.89 | 13.94 ± 1.1 | 2.73 ± 0.39 |
| Group II | 0.37 ± 0.01 a | 20.2 ± 1.30 a | 9.74 ± 0.81 a | 13.36 ± 0.77 a |
| Group III | 0.15 ± 0.012 b | 10.8 ± 0.83 b | 15 ± 1.1 b | 4.48 ± 0.94 b |
| Group IV | 0.17 ± 0.014 b | 13.6 ± 1.1 b | 16.8 ± 1.3 b | 5.97 ± 0.0.39 b |
| Group V | 0.07 ± 0.002 bc | 10.5 ± 0. 5 bc | 23.2 ± 1.48 bc | 6.21 ± 0.16 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attallah, N.G.M.; El-Sherbeni, S.A.; El-Kadem, A.H.; Elekhnawy, E.; El-Masry, T.A.; Elmongy, E.I.; Altwaijry, N.; Negm, W.A. Elucidation of the Metabolite Profile of Yucca gigantea and Assessment of Its Cytotoxic, Antimicrobial, and Anti-Inflammatory Activities. Molecules 2022, 27, 1329. https://doi.org/10.3390/molecules27041329
Attallah NGM, El-Sherbeni SA, El-Kadem AH, Elekhnawy E, El-Masry TA, Elmongy EI, Altwaijry N, Negm WA. Elucidation of the Metabolite Profile of Yucca gigantea and Assessment of Its Cytotoxic, Antimicrobial, and Anti-Inflammatory Activities. Molecules. 2022; 27(4):1329. https://doi.org/10.3390/molecules27041329
Chicago/Turabian StyleAttallah, Nashwah G. M., Suzy A. El-Sherbeni, Aya H. El-Kadem, Engy Elekhnawy, Thanaa A. El-Masry, Elshaymaa I. Elmongy, Najla Altwaijry, and Walaa A. Negm. 2022. "Elucidation of the Metabolite Profile of Yucca gigantea and Assessment of Its Cytotoxic, Antimicrobial, and Anti-Inflammatory Activities" Molecules 27, no. 4: 1329. https://doi.org/10.3390/molecules27041329