
The (Bio)Chemistry of Non-Transferrin-Bound Iron



1
LAQV-REQUIMTE, Departamento de Química e Bioquímica, Faculdade de Ciências, Universidade do Porto, Rua do Campo Alegre s/n, 4169-007 Porto, Portugal
2
LAQV-REQUIMTE, Instituto de Ciências Biomédicas Abel Salazar, Universidade to Porto, Rua Jorge Viterbo Ferreira N228, 4050-313 Porto, Portugal
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(6), 1784; https://doi.org/10.3390/molecules27061784
Submission received: 15 February 2022
/
Revised: 4 March 2022
/
Accepted: 7 March 2022
/
Published: 9 March 2022
(This article belongs to the Special Issue Molecular and Spectroscopic Insights into Metal Ions Speciation in Extracellular Fluids)
{kind=link}
{kind=link}
Abstract
:In healthy individuals, virtually all blood plasma iron is bound by transferrin. However, in several diseases and clinical conditions, hazardous non-transferrin-bound iron (NTBI) species occur. NTBI represents a potentially toxic iron form, being a direct cause of oxidative stress in the circulating compartment and tissue iron loading. The accumulation of these species can cause cellular damage in several organs, namely, the liver, spleen, and heart. Despite its pathophysiological relevance, the chemical nature of NTBI remains elusive. This has precluded its use as a clinical biochemical marker and the development of targeted therapies. Herein, we make a critical assessment of the current knowledge of NTBI speciation. The currently accepted hypotheses suggest that NTBI is mostly iron bound to citric acid and iron bound to serum albumin, but the chemistry of this system remains fuzzy. We explore the complex chemistry of iron complexation by citric acid and its implications towards NTBI reactivity. Further, the ability of albumin to bind iron is revised and the role of protein post-translational modifications on iron binding is discussed. The characterization of the NTBI species structure may be the starting point for the development of a standardized analytical assay, the better understanding of these species’ reactivity or the identification of NTBI uptake mechanisms by different cell types, and finally, to the development of new therapies.
Keywords:
iron overload; non-transferrin-bound iron; NTBI; labile plasma pool; LPI; citrate; albumin1. Introduction
Iron is an essential micronutrient for almost all living organisms, participating in a variety of metabolic processes. Perhaps the best-known function of iron is its presence in hemoglobin, where it is involved in the transport of oxygen. However, iron plays a crucial role in cellular processes such as DNA synthesis, epigenetic regulation, or cellular respiration [1].
The biological importance of iron results from its adaptable chemistry and natural abundance in the Earth’s crust [2]. Iron can accept or donate electrons, participating in oxidation–reduction reactions, occurring under various oxidation states (from −2 to +6). The most common oxidation states are Fe3+, a hard acid that prefers coordinating oxygen atoms, and Fe2+, an intermediate acid that favors bonding with ligands containing nitrogen or sulfur atoms [2]. However, these same chemical properties are responsible for iron toxicity. This harmful effect is related to the role played by iron in the Fenton reaction, which can amplify the deleterious effects of oxidative stress [3].
To avoid its hazardous effects, in the human body, iron homeostasis is tightly controlled. Humans do not actively excrete iron, and therefore, maintenance of iron balance involves the control of its absorption, storage, use and distribution. This is mainly achieved by the action of hepcidin, the hormone which controls cellular iron export and its release into circulation [4].
Hemoglobin-bound iron, present in circulating erythrocytes and developing erythroid precursors, represents about two-thirds of the total iron present in the body [5]. Most of the iron required for erythropoiesis comes from the recycling of senescent red cells promoted by macrophages of the reticuloendothelial system. This recycling mechanism is essential, as only 1 to 2 mg of iron are daily absorbed from the diet. The remaining iron in the body is stored in hepatocytes and macrophages, or it is being used for cellular metabolic processes.
The link between the places of iron absorption, utilization and storage is made by the blood circulation, where iron transport is carried out by transferrin (Tf). Tf is a glycoprotein, produced in the hepatocytes, with two globular domains, each with a binding site with high affinity for the Fe3+ ion [6]. Healthy individuals present typical Tf saturation levels around 30%, ensuring a large blood plasma unsaturated iron-binding capacity [7]. Iron-binding by Tf allows its solubilization at physiological pH values and hinders its participation in deleterious redox reactions. Furthermore, Tf functions as an iron addressing system, ensuring that cellular uptake only occurs in cells expressing the transferrin receptor (TfR) and avoiding undesired tissue iron deposition. However, under several pathological conditions, blood serum NTBI has been observed. NTBI has been proposed as a main toxicity mediator, responsible for increased oxidative stress and tissue iron deposition, leading to several comorbidities such as liver damage, endocrinopathies, cardiac disease, atherosclerosis and neurodegeneration [8,9,10,11,12,13,14]. Therefore, NTBI constitutes an important concept for the clinical management of patients [15,16], but lack of knowledge regarding the chemical nature of NTBI has precluded the development of standardized assays and its use as a biochemical marker in clinical practice [15,17].
2. NTBI: Non-Transferrin-Bound Iron
NTBI was first described by Hersko et al. as iron which was available for chelation by diethylenetriamine pentaacetic acid in the presence of fully saturated Tf in the serum of thalassemia patients [18]. Since, its presence has been confirmed in transfusional iron overload [19,20] and hereditary hemochromatosis (HH) [21,22,23]. However, the presence of NTBI is not restricted to iron overload diseases and these toxic iron species have been detected in diabetes mellitus, end-stage renal disease, cancer patients undergoing chemotherapy, or anemia patients under iron supplementation regimens [24,25,26,27].
At present, NTBI is defined as blood serum iron not bound to Tf, heme or ferritin [28]. As recently proposed by Cabantchik and Hershko [15], a further distinction should be made between iron associated with endogenous plasma ligands and iron associated with chelators used in therapeutic practices or ferric oxyhydroxy-carbohydrate structures derived from iron supplements. Along with NTBI, different terms are commonly used to refer to blood serum iron that is not bound to Tf. These indicate the chemical heterogeneity of NTBI species and probably reflect the existence of sub-pools showing different lability and redox reactivity [29]. Labile plasma iron (LPI) has been used to describe NTBI species which are readily available to participate in redox cycling and, similarly, directly chelatable iron (DCI) has been used to describe iron which is available to direct chelation by an exogenous ligand [29,30,31,32]. However, NTBI has mostly prevailed as the terminology used for extracellular iron species, while labile iron pool or labile cell iron are commonly used when referring to intracellular labile iron species [29,33].
The most common cause for the appearance of NTBI species will be high Tf saturation. Nevertheless, NTBI has been confirmed in the sera of HH patients with only partially saturated Tf [21,22,23,34]. Currently, it is commonly accepted that NTBI will appear when Tf saturation levels exceed 70% [12], but the presence of NTBI has been confirmed in diabetes patients with Tf saturation below 50% and even in healthy individuals [25,35]. Therefore, the presence of NTBI cannot be regarded as a simple Tf spillover phenomenon, but rather as an expression of the kinetics balance between iron egress into blood serum, binding by Tf, and removal/utilization from circulation [36].
NTBI is taken up by several tissues, but the main organ responsible for its clearance from circulation is the liver, as recently reviewed by Knutson [37], with hepatocytes being particularly effective in its uptake [38,39,40,41,42]. Despite the high efficiency of NTBI extraction by the liver, persistence of these iron species promotes iron deposition in other organs, such as the pancreas, kidney and heart [43,44,45]. Studies in hypotransferrinemic mice, with 59Fe, have shown that NTBI was removed from serum with a half-life of 30 s, as opposed to the 50 min required for Tf-bound iron [43]. This greater efficiency in the uptake of NTBI, when compared to Tf-bound iron, seems a common feature for various tissues. Furthermore, contrary to what is observed with Tf, NTBI internalization appears not to be inhibited by cellular iron loading [28,37].
Several proteins have been identified as NTBI transporters (reviewed by Knutson [37]). These proteins include divalent metal transporter 1 (DMT1), ZRT/IRT-like proteins 8 and 14 (ZIP8 and ZIP14), and calcium channels (LTCC, TTCC and VGCC). The relevance of these transporters seems to be cell-type-specific, with NTBI uptake in the hepatocytes and pancreas β-cells mainly occurring through ZIP14 [46,47], in neurons through ZIP8 and VGCC [48], while uptake in cardiomyocytes occurs through LTCC [49,50]. The referred proteins are divalent metal transporters, and an iron reduction step is required prior to NTBI uptake. This observation is supported by the inhibition of cellular Fe3+ species uptake in the presence of specific Fe2+ chelators [47,51]. The involvement of several cell surface ferrireductases in NTBI transport has been proposed, such as the cytochrome b561 (Cyt b561) family (including Dcytb), Steap proteins, prion protein (PrP), and α-synuclein [37]. Dcytb is expressed at the cell surface of astrocytes [52], and Steap2 colocalizes with Zip8 in neurons [48]. PrP is a ubiquitously expressed cell surface protein, and knock-out studies have indicated that it may play a role in NTBI uptake in the liver, kidney and pancreas [53,54].
Despite these recent advances in the identification of NTBI transporters, the full depiction of NTBI uptake routes will require the chemical characterization of NTBI speciation and the identification of pathophysiological NTBI species.
3. The Chemical Characterization of NTBI
Like most extracellular iron species, NTBI is assumed to be constituted by Fe3+. The release of iron into the bloodstream occurs through ferroportin, which requires an associated ferroxidase activity [55,56]. Furthermore, the abundance of ceruplasmin, a ferroxidase, in the serum should ensure that iron remains in the Fe3+ oxidation state [57]. Contrarily, in the intracellular environment, the predominant iron oxidation state is Fe2+ [58].
Given the poor solubility of the ferric ion, Fe3+, at the serum pH value, NTBI species must consist of iron associated with blood serum components responsible for its binding and solubilization. Low molecular weight (LMW) carboxylic acid compounds, with oxygen donor atoms, are the most likely candidates for NTBI ligands. In fact, early chemical models of iron speciation in the plasma predicted 99% of iron to be associated to citrate, as ferric citrate hydroxide [59]. The involvement of citrate in NTBI composition was later elegantly confirmed by nuclear magnetic resonance (NMR) experiments with sera from HH patients [60]. The comparison between the NMR spectra of plasma samples collected from healthy individuals and patients diagnosed with HH allowed the identification of differences in signal intensities from citrate and acetate. To validate if these differences could be ascertained to iron binding, plasma samples were also incubated with the iron chelator desferrioxamine or supplemented with exogenous Fe3+, indicating that spectral differences were indeed resulting from the presence of iron.
Early experiments applying size exclusion chromatography to the serum of hypotransferrinemic mice have indicated the presence of high molecular weight (HMW) NTBI species [61]. The existence of HMW-NTBI species has since been confirmed in the serum from thalassemia patients, with only approximately 10% of NTBI being able to penetrate 30 kDa exclusion ultrafiltration devices [62]. These HMW iron species could consist of polymeric iron species or iron non-specifically associated with serum proteins. Being the most abundant protein in the serum and recognized as a carrier both for organic compounds and metal ions [63,64], human serum albumin (HSA) has been proposed as the most likely protein candidate [65]. In fact, iron binding by albumin has been observed by several in vitro studies, but definitive proof for iron association with this protein in vivo has remained elusive [62,66].
4. Ferric Citrate Chemistry
Citrate is a ubiquitous iron chelator, acting as a bacterial or plant siderophore, as the main iron transporter in the plant xylem sap, or an NTBI ligand in the blood serum [67,68,69]. Citrate is a typical α-hydroxycarboxylate siderophore [67], capable to coordinate iron by its three carboxylic acid groups and its hydroxyl moiety. In fact, the hydroxyl group is fundamental for iron coordination and may determine the affinity of these iron chelators [70].
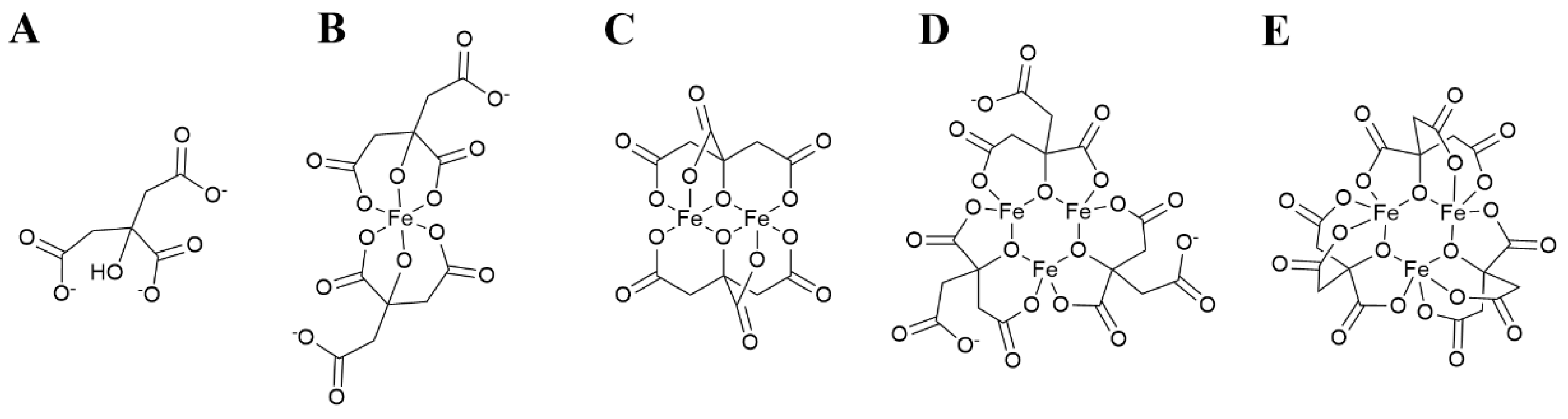
The structure of citrate (Figure 1A) determines a diversity of coordination modes, most commonly as a tridentate ligand, with coordination of Fe3+ occurring through two carboxylate and the hydroxyl moieties [71]. This coordination mode leaves a free carboxylate capable to bind another Fe3+ ion, which favors the formation of polymeric complexes. To date, monoferric dicitrate ([Fe(Cit)2]5−), diferric dicitrate ([Fe2(Cit)2]2−), diferric tricitrate ([Fe2(Cit)3]6−) and nonairon octacitrate ([Fe9O(Cit)8]7−) complexes have been isolated and their crystal structure has been determined [71,72,73,74] (schematic representation in Figure 1B,C). Species with 3 ferric ions ([Fe3(Cit)3]3− and [Fe3O(Cit)3]5−) have been identified by mass spectrometry [71,75,76] (Figure 1D,E).
Aqueous solution studies, probed by mass spectrometry, indicate that the different ferric citrate complexes co-exist in an intricate chemical equilibrium, with the relative abundance of individual species depending on the pH value and the iron:citrate molar ratio [71,76]. Low ratios (corresponding to high excess citrate) favor formation of [Fe(Cit)2]5− while increasing the iron abundance leads to the formation of polynuclear species [Fe2(Cit)2]2− and [Fe3(Cit)3]3−. This effect combines with medium acidity, with low pH values favoring polynuclear species and a basic environment favoring the mononuclear complex. Under the serum conditions of pH, physiological levels of citrate (80–120 µM) and typical NTBI concentrations (1–10 µM) [Fe(Cit)2]5− are the most abundant species with polynuclear [Fe2(Cit)2]2− and [Fe3(Cit)3]3− species only existing at relevant concentrations for the higher NTBI values.
The complex speciation of ferric citrate under physiological conditions provides valuable insights for the reactivity and bioavailability of NTBI. Bacteria seem to show a preference for polynuclear ferric citrate complexes, with FecA, the outer membrane ferric citrate transporter from E. coli, binding [Fe2(Cit)2]2− [77,78] and FecC, from B. cereus, binding [Fe3(Cit)3]3− [75]. A similar preference has been described for eukaryotic cells, with isolated T lymphocytes and hepatocytes showing a preference for the uptake of polynuclear species ([Fe3(Cit)3]3−) [39].
A required step for NTBI cellular uptake is Fe3+ reduction [39,79,80]. Reduction may be promoted by LMW reducing agents such as ascorbate or by a series of transmembrane ferrireductases [81,82]. The speciation of ferric citrate may be essential in determining NTBI availability to this reduction step. Furthermore, it may impact on the redox activity of NTBI and its ability to promote oxidative damage in the serum and extracellular environment. Although reduction potentials are difficult to determine for the ferric citrate system due to its complex speciation, Vukosava et al. have reported that at pH = 5.5 reduction of [Fe(Cit)2]5− occurs at −0.1 V, while reduction of polynuclear complexes is observed at lower reduction potentials (−0.28 V) [83]. Contrastingly, Adam and co-workers reported a reduction potential of ca. 0 V (−0.03 V < E°′ > +0.01 V) for Fe3+—citrate species at physiological pH, with only small variations resulting from different iron:citrate ratios [84]. Further, the authors have evaluated the ability of ascorbic acid to induce iron redox cycling in the presence of high citrate excess, where the [Fe(Cit)2]5− species will predominate. Their results supporting the occurrence of slow redox cycling in the presence of H2O2, and thus pinpointing Fe3+—citrate complexes as potential mediators of oxidative stress.
The speciation of ferric citrate species is also a determinant for the effective chelation of NTBI. Kinetics experiments conducted by Gautier-Luneau et al. have shown that the transfer of iron to external chelators (TRENCAMS or O-TRENSOX) at physiological pH occurs more than 20 times faster for the [Fe(Cit)2]5− species than for polynuclear complexes ([Fe2(Cit)2]2− or [Fe3(Cit)3]3−) [71].
5. Iron Binding by Serum Albumin
HSA is the most abundant protein in the blood plasma, with a concentration around 40 g/L (~600 µM). HSA is a multicarrier protein, with the ability to bind numerous metabolites, fatty acids and also metal ions [63]. The interaction of HSA with divalent metal ions is well described and two binding sites have been characterized (reviewed by Bal et al. [85] and Al-Harthi et al. [63]): the protein N-terminal site (NTS) [86], which has shown a preference for Cu2+ and Ni2+ binding, and the metal binding site A (MBS-A) [87,88,89,90], at the interface of domains I and II, which shows greater selectivity towards Zn2+ and Cd2+ (Figure 2A). Additional metal ion binding sites have been proposed, with a putative MBS-B being involved in Zn2+ and Cd2+ binding [88,89,90]. Coordination at these binding sites involves both nitrogen and oxygen atoms, respectively, from histidine sidechains, the protein backbone or carboxylate residues. Binding of Hg2+, Au+ and Pt+ has been shown to occur at the reduced Cys34 residue [91,92,93].
Several reports of iron binding to HSA have also been published, but no specific binding site has been identified. Fe3+ present in the blood plasma should favor coordination to oxygen atoms in the carboxylate moieties of aspartate and glutamate sidechains. HSA is rich in ionic amino acids and has a net negative charge of –15 at physiological pH values, with a total of 98 carboxylate moieties (36 Asp and 62 Glu residues), most lying at the protein surface (Figure 2B). It is hypothesized that Fe3+ can bind non-specifically to some of these carboxylate clusters, similarly to what has been described for Ca2+ [95,96].
Anghileri has shown the possibility to prepare stable iron-HSA complexes from 59FeCl3 in acetate buffer, probed by paper electrophoresis and gel filtration chromatography [97]. Hershko and co-workers [18] noted that 59FeCl3 added to thalassemic sera migrated with the HSA fraction in electrophoretic separation. Using, bovine serum albumin (BSA) Loban et al. [98] predicted a total of 9 iron binding sites. However, more recently, employing a fluorescence titration, Xu et al. reported on the existence of only 1 specific binding site, with Kd = 3.46 × 10−8 M at 37 °C [99]. The described studies were performed in the absence of an iron ligand, and interference from the likely formation of iron hydroxide species cannot be excluded.
Due to the poor solubility of Fe3+ at physiological pH values, studies of iron binding to HSA have also been carried out in citrate media. Coddington and Perkins [100] have demonstrated binding at low pH value, with experiments with acetylated or esterified albumin suggesting the electrostatic association of negatively charged Fe3+-citrate complexes to protein positive charged sites. However, with native HSA, binding was suppressed at pH 7 and an iron:citrate molar ratio of 1:5 prevented binding above pH 5. Nevertheless, the authors estimated the existence of 13 iron binding sites. More recently, using gel filtration chromatography, Løvstad [101] has proposed the formation of ternary BSA-Fe3+-citrate complexes, with albumin being able to bind 2 to 3 ferric ions. This study was, however, carried out with iron and citrate present in a 1:1 molar ratio, which may promote the formation of iron-hydroxyde-citrate HMW structures [76,102] and may not reflect the presence of physiologically relevant Fe3+-citrate species.
More recently, Evans et al. [62] have assessed iron binding by HSA under conditions relevant for NTBI speciation, evaluating physiologically relevant molar ratios of iron:citrate, while maintaining physiologically relevant values for citrate concentration and pH, and iron levels typically found in β-thalassemia major patients. Under such conditions, it has been found that HSA was able to hinder the permeation of ferric citrate complexes through 30 kDa cutoff filtration units, suggesting that it plays a significant role in NTBI speciation. However, in the same study, it proved to be impossible to isolate HSA-bound iron from the sera of β-thalassemia patients and HSA immunoprecipitation did not reduce the detectable NTBI in these serum samples. Subsequently, we have evaluated the interaction of Fe3+ and HSA by gel permeation chromatography [66] under physiological concentrations of albumin (40 g/L) and citrate (100 µM). It was shown that with native HSA at low Fe3+ concentrations (1 µM), most of the Fe3+ available (~60%) associates with the protein fraction, this value decreasing logarithmically to less than 20% at higher iron values (>10 µM). 14C-labelled citrate was used to evaluate the formation of ternary complexes, with results suggesting that most of the HSA-bound iron is directly associated to the protein. Furthermore, our results demonstrate that non-enzymatic protein post-translational modifications, such as oxidation or glycation, have the potential significantly increase the ability of HSA to bind iron. These modifications were thought to introduce chelating functional groups to HSA, and even when present at low relative abundance, may have a profound impact in the distribution of NTBI species.
6. NTBI Detection and Quantification
Over the years, several methods have been developed to quantify NTBI [29]. The most widely used assay is based on NTBI mobilization by a chelating agent (commonly, nitrilotriacetic acid—NTA) under experimental conditions which limit the mobilization of transferrin-bound iron (TBI) [22,103]. The Fe-NTA complex is thus separated from TBI by molecular weight exclusion ultrafiltration and the iron quantified in the ultrafiltrate.
Alternative methods, such as the bleomycin and the labile iron pool (LPI) assays, take advantage of iron reactivity and depend on NTBI reduction. Bleomycin can act as a selective Fe2+ chelator [104]. In the LPI assay, which has been implemented in several laboratories, serum is supplemented with ascorbic acid to initiate the iron dependent Fenton catalyzed oxidation of a fluorescent probe (1,2,3-dihydrorhodamine) [31]. However, both assays tend to give rather low NTBI values, which is generally attributed to them only being able to assess a sub-pool of the heterogeneous NTBI group of species. Furthermore, both seem to being subject to interferences from heme and hemoglobin.
Recently, Ma and co-workers [105] have introduced a fluorescence bead assay which allows direct NTBI detection by flow cytometry. The main advantage of this bead assay would be the simplicity of use, taking advantage of common analytical equipment present in almost any modern hospital. The method has been extensively compared with the NTA-based assay, showing low agreement in their results [106]. Nevertheless, it has shown a lower dependence on Tf saturation, preventing both the existence of false negatives at the lower Tf saturation bound and false positives at high Tf saturation levels.
Two international round robins have been promoted, where samples have been circulated by several laboratories [17,107]. Results show that NTBI values for the same sample vary considerably between assays and even between laboratories reporting on the same method. Used in the most recent study, the bead assay failed to detect NTBI in most samples [17]. Furthermore, both the NTA-based assay and the LPI assay are laborious and seem to show analyst dependence.
The development of a consistent assay is a requirement for NTBI to become an important biochemical biomarker in the clinical management of iron overload and related diseases. It seems that a better understanding of the serum speciation of these elusive iron species is crucial to allow the development of new and more reliable analytical methods.
7. NTBI in Iron Related Disorders
NTBI was first identified in iron overload diseases, either HH or hematological iron overload disorders (β-thalassemia major and intermedia) and NTBI uptake is thought to be the major route for iron deposition in several tissues. Liver hepatocytes are particularly efficient in NTBI clearance [40,42,108], which justifies the high liver iron loading observed in HH. The heart and the pancreas are also important sites of iron accumulation in iron overload. Studies with mice have shown that these organs are loaded when the liver approaches its iron loading capacity [109,110]. Iron loading in the pancreas occurs mainly in acinar and β-cells [111,112], the latter being particularly susceptible to iron-promoted dysfunction [113]. Proportionally, cardiomyocytes seem to load less iron than the liver or the pancreas, but they show a preference for NTBI uptake over TBI [114] and their function is significantly hindered by iron loading [115].
NTBI has also been detected in diabetes mellitus (DM) [25,35,116,117] and type 2 DM patients show increased iron stores [118]. The presence of NTBI is thought to be a major contributor to increased oxidative stress both in intracellular and the circulating compartment. Furthermore, NTBI levels seem to correlate with both liver and pancreatic functions, with patients showing increased levels of insulin resistance and decreased insulin production [119,120]. NTBI-generated oxidative stress also seems to play a role in the development of comorbidities in the general population, with recent studies showing a correlation with increased cardiac disease [9] and a mechanistic link with the development of atherosclerosis [10].
These toxic iron species have also been detected in other pathologies such as end-stage renal disease, myelodysplastic syndrome, and during myeloablative therapy and stem cell transplantation [24,28]. Although the pathophysiological role of NTBI in these diseases is less explored, it is believed to promote tissue iron deposition and oxidative stress.
Iron has also been long recognized as an important player in neurodegeneration [14]. Brain iron levels are associated with cognitive decline in Alzheimer’s disease [121] and iron has been shown to colocalize with amyloid β deposits [122,123,124]. Furthermore, iron deposits seem to be involved in the generation of oxidative stress and cell death through ferroptosis [125,126]. Several studies have also related iron accumulation in the substantia nigra of Parkinson patients with disease severity [127,128,129]. Dyshomeostasis of brain iron metabolism is also linked with Friedreich’s ataxia, multiple sclerosis, Huntington’s disease, aceruloplasminemia and neuroferritinopathy [14].
There is no clear evidence that high iron load and the persistent presence of circulating NTBI in HH and β-thalassemia major predispose to neurodegeneration, but recent studies have correlated the prevalence of HFE polymorphisms with brain iron load and homeostasis, suggesting that the brain may not be entirely protected in iron overload conditions [130,131]. Furthermore, brain iron accumulation has been reported in hypotransferrinemic mice [132] and β-thalassemia patients [133,134]. In fact, the transport into the brain of 59Fe-labelled NTBI (in the form of FeCl3 or ferric ammonium citrate) has been shown in a mouse model [135]. However, it should be noted that, given the high Tf saturation, NTBI is a physiological form of iron in the cerebrospinal fluid [136] and it is actively taken up by microglia, astrocytes and neurons [137].
8. Final Remarks
More than four decades have passed since the discovery of NTBI, but its biochemical characterization remains elusive. The term, NTBI, refers to a heterogenous pool of iron species, resulting from the binding Fe3+ to ligands in the blood serum. Currently, the best working chemical model assumes that the main serum ligands are citrate anions and HSA. The most recent results suggest that these compartments form a fine-tuned equilibrium dependent on Fe3+ concentration, iron:citrate molar and HSA modifications. At low NTBI levels, Fe3+ is expected to be mostly associated with HSA, with mononuclear [Fe(Cit)2]5− being the predominant LMW species. At the higher bound of pathophysiological NTBI concentrations (>10 uM), most of the NTBI will be associated with citrate, with mononuclear [Fe(Cit)2]5− being the predominant species and polynuclear complexes ([Fe2(Cit)2]2− or [Fe3(Cit)3]3−) representing 20% to 40% of the non-protein-bound iron. Recently, Dziuba et al. [138] reported that LMW iron in healthy plasma samples was not bound to citrate. Under these conditions, however, Fe3+-citrate is not expected to exist, as the predominant apo-Tf is predicted to out-compete citrate for the binding of such iron. These iron species, occur in the nM range and, most probably, are transient species rapidly cleared by Tf.
NTBI speciation will define the toxicity pathways of these iron species, by modulating their redox reactivity [84], or preferential cellular uptake routes [39]. In particular, the biological implications of iron binding by HSA have been less explored, but they are certain to play a decisive role in the biochemistry of NTBI. HSA may modulate the pro-oxidant character of NTBI in the plasma [84,139], and it facilitates iron donation to apo-Tf [140]. HSA also offers an alternative route for cellular uptake, through endocytosis [141]. Furthermore, HSA post-translational modifications offer a possible explanation for disease-specific modulation of NTBI reactivity and toxicity [66]. This is particularly relevant in a context where similar modifications may also impact the ability of Tf to bind iron [6]. Finally, it is relevant to note that HSA-bound iron is underestimated by the NTA-based assay [66].
Despite the progress achieved in the past years, a better understanding of NTBI speciation is a crucial step towards defining clinically relevant isoforms and the development of reliable analytical assay.
Author Contributions
Conceptualization, A.M.N.S. and M.R.; writing—original draft preparation, A.M.N.S. and M.R.; writing—review and editing, A.M.N.S. and M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work received financial support from PT national funds (FCT/MCTES, Fundação para a Ciência e Tecnologia and Ministério da Ciência, Tecnologia e Ensino Superior) through the project UIDB/50006/2020|UIDP/50006/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors have no conflict of interest to state.
References
- Crichton, R.R. The Importance of Iron for Biological Systems. In Inorganic Biochemistry of Iron Metabolism, 2nd ed.; Crichton, R.R., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 17–48. [Google Scholar]
- Sánchez, M.; Sabio, L.; Gálvez, N.; Capdevila, M.; Dominguez-Vera, J.M. Iron chemistry at the service of life. IUBMB Life 2017, 69, 382–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppenol, W.H.; Hider, R.H. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hentze, M.W.; Muckenthaler, M.U.; Andrews, N.C. Balancing acts: Molecular control of mammalian iron metabolism. Cell 2004, 117, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Andrews, N.C. Disorders of iron metabolism. N. Engl. J. Med. 1999, 341, 1986–1995. [Google Scholar] [CrossRef]
- Silva, A.M.N.; Moniz, T.; de Castro, B.; Rangel, M. Human transferrin: An inorganic biochemistry perspective. Coord. Chem. Rev. 2021, 449, 214186. [Google Scholar] [CrossRef]
- Cazzola, M.; Huebers, H.A.; Sayers, M.H.; MacPhail, A.P.; Eng, M.; Finch, C.A. Transferrin saturation, plasma iron turnover, and transferrin uptake in normal humans. Blood 1985, 66, 935–939. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Loréal, O. Role of non-transferrin-bound iron in the pathogenesis of iron overload and toxicity. Adv. Exp. Med. Biol. 2002, 509, 45–53. [Google Scholar] [CrossRef]
- Sugiura, T.; Dohi, Y.; Takase, H.; Fujii, S.; Seo, Y.; Ohte, N. Analytical evaluation of serum non-transferrin-bound iron and its relationships with oxidative stress and cardiac load in the general population. Medicine 2021, 100, e24722. [Google Scholar] [CrossRef]
- Vinchi, F.; Porto, G.; Simmelbauer, A.; Altamura, S.; Passos, S.T.; Garbowski, M.; Silva, A.M.N.; Spaich, S.; Seide, S.E.; Sparla, R.; et al. Atherosclerosis is aggravated by iron overload and ameliorated by dietary and pharmacological iron restriction. Eur. Heart J. 2020, 41, 2681–2695. [Google Scholar] [CrossRef]
- Roghi, A.; Poggiali, E.; Duca, L.; Mafrici, A.; Pedrotti, P.; Paccagnini, S.; Brenna, S.; Galli, A.; Consonni, D.; Cappellini, M.D. Role of Non-Transferrin-Bound Iron in the pathogenesis of cardiotoxicity in patients with ST-elevation myocardial infarction assessed by Cardiac Magnetic Resonance Imaging. Int. J. Cardiol. 2015, 199, 326–332. [Google Scholar] [CrossRef]
- Piga, A.; Longo, F.; Duca, L.; Roggero, S.; Vinciguerra, T.; Calabrese, R.; Hershko, C.; Cappellini, M.D. High nontransferrin bound iron levels and heart disease in thalassemia major. Am. J. Hematol. 2009, 84, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, S.A.; Brown, K.E. Iron-Induced Liver Injury: A Critical Reappraisal. Int. J. Mol. Sci. 2019, 20, 2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Cabantchik, I.Z.; Hershko, C. Plasma non transferrin bound iron—NTBI revisited. Implications for systemic iron overload and in iv iron supplementation. Am. J. Hematol. 2022, 97, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Silva, A.M.; Podinovskaia, M.; Ma, Y. Monitoring the efficiency of iron chelation therapy: The potential of nontransferrin-bound iron. Ann. N. Y. Acad. Sci. 2010, 1202, 94–99. [Google Scholar] [CrossRef] [PubMed]
- de Swart, L.; Hendriks, J.C.; van der Vorm, L.N.; Cabantchik, Z.I.; Evans, P.J.; Hod, E.A.; Brittenham, G.M.; Furman, Y.; Wojczyk, B.; Janssen, M.C.; et al. Second international round robin for the quantification of serum non-transferrin-bound iron and labile plasma iron in patients with iron-overload disorders. Haematologica 2016, 101, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershko, C.; Graham, G.; Bates, G.W.; Rachmilewitz, E.A. Non-specific serum iron in thalassaemia: An abnormal serum iron fraction of potential toxicity. Br. J. Haematol. 1978, 40, 255–263. [Google Scholar] [CrossRef]
- Anuwatanakulchai, M.; Pootrakul, P.; Thuvasethakul, P.; Wasi, P. Non-transferrin plasma iron in beta-thalassaemia/Hb E and haemoglobin H diseases. Scand. J. Haematol. 1984, 32, 153–158. [Google Scholar] [CrossRef]
- Wang, W.C.; Ahmed, N.; Hanna, M. Non-transferrin-bound iron in long-term transfusion in children with congenital anemias. J. Pediatr. 1986, 108, 552–557. [Google Scholar] [CrossRef]
- Aruoma, O.I.; Bomford, A.; Polson, R.J.; Halliwell, B. Nontransferrin-bound iron in plasma from hemochromatosis patients: Effect of phlebotomy therapy. Blood 1988, 72, 1416–1419. [Google Scholar] [CrossRef] [Green Version]
- Gosriwatana, I.; Loreal, O.; Lu, S.; Brissot, P.; Porter, J.; Hider, R.C. Quantification of non-transferrin-bound iron in the presence of unsaturated transferrin. Anal. Biochem. 1999, 273, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Loréal, O.; Gosriwatana, I.; Guyader, D.; Porter, J.; Brissot, P.; Hider, R.C. Determination of non-transferrin-bound iron in genetic hemochromatosis using a new HPLC-based method. J. Hepatol. 2000, 32, 727–733. [Google Scholar] [CrossRef]
- Breuer, W.; Hershko, C.; Cabantchik, Z.I. The importance of non-transferrin bound iron in disorders of iron metabolism. Transfus. Sci. 2000, 23, 185–192. [Google Scholar] [CrossRef]
- Lee, D.H.; Liu, D.Y.; Jacobs, D.R., Jr.; Shin, H.R.; Song, K.; Lee, I.K.; Kim, B.; Hider, R.C. Common presence of non-transferrin-bound iron among patients with type 2 diabetes. Diabetes Care 2006, 29, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Dresow, B.; Petersen, D.; Fischer, R.; Nielsen, P. Non-transferrin-bound iron in plasma following administration of oral iron drugs. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2008, 21, 273–276. [Google Scholar] [CrossRef]
- Halliwell, B.; Aruoma, O.I.; Mufti, G.; Bomford, A. Bleomycin-detectable iron in serum from leukaemic patients before and after chemotherapy. Therapeutic implications for treatment with oxidant-generating drugs. FEBS Lett. 1988, 241, 202–204. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Ropert, M.; Le Lan, C.; Loréal, O. Non-transferrin bound iron: A key role in iron overload and iron toxicity. Biochim. Biophys. Acta 2012, 1820, 403–410. [Google Scholar] [CrossRef]
- Cabantchik, Z.I. Labile iron in cells and body fluids: Physiology, pathology, and pharmacology. Front. Pharmacol. 2014, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Breuer, W.; Ermers, M.J.; Pootrakul, P.; Abramov, A.; Hershko, C.; Cabantchik, Z.I. Desferrioxamine-chelatable iron, a component of serum non-transferrin-bound iron, used for assessing chelation therapy. Blood 2001, 97, 792–798. [Google Scholar] [CrossRef] [Green Version]
- Esposito, B.P.; Breuer, W.; Sirankapracha, P.; Pootrakul, P.; Hershko, C.; Cabantchik, Z.I. Labile plasma iron in iron overload: Redox activity and susceptibility to chelation. Blood 2003, 102, 2670–2677. [Google Scholar] [CrossRef] [Green Version]
- Pootrakul, P.; Breuer, W.; Sametband, M.; Sirankapracha, P.; Hershko, C.; Cabantchik, Z.I. Labile plasma iron (LPI) as an indicator of chelatable plasma redox activity in iron-overloaded β-thalassemia/HbE patients treated with an oral chelator. Blood 2004, 104, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Cabantchik, Z.I. The labile iron pool: Characterization, measurement, and participation in cellular processes(1). Free Radic. Biol. Med. 2002, 33, 1037–1046. [Google Scholar] [CrossRef]
- de Valk, B.; Addicks, M.A.; Gosriwatana, I.; Lu, S.; Hider, R.C.; Marx, J.J. Non-transferrin-bound iron is present in serum of hereditary haemochromatosis heterozygotes. Eur. J. Clin. Investig. 2000, 30, 248–251. [Google Scholar] [CrossRef]
- Van Campenhout, A.; Van Campenhout, C.; Lagrou, A.R.; Moorkens, G.; De Block, C.; Manuel-y-Keenoy, B. Iron-binding antioxidant capacity is impaired in diabetes mellitus. Free Radic. Biol. Med. 2006, 40, 1749–1755. [Google Scholar] [CrossRef]
- Porter, J.B.; Walter, P.B.; Neumayr, L.D.; Evans, P.; Bansal, S.; Garbowski, M.; Weyhmiller, M.G.; Harmatz, P.R.; Wood, J.C.; Miller, J.L.; et al. Mechanisms of plasma non-transferrin bound iron generation: Insights from comparing transfused diamond blackfan anaemia with sickle cell and thalassaemia patients. Br. J. Haematol. 2014, 167, 692–696. [Google Scholar] [CrossRef]
- Knutson, M.D. Non-transferrin-bound iron transporters. Free Radic. Biol. Med. 2019, 133, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.; Baker, S.M.; Morgan, E.H. Characterisation of non-transferrin-bound iron (ferric citrate) uptake by rat hepatocytes in culture. Biochim. Biophys. Acta 1998, 1380, 21–30. [Google Scholar] [CrossRef]
- Arezes, J.; Costa, M.; Vieira, I.; Dias, V.; Kong, X.L.; Fernandes, R.; Vos, M.; Carlsson, A.; Rikers, Y.; Porto, G.; et al. Non-transferrin-bound iron (NTBI) uptake by T lymphocytes: Evidence for the selective acquisition of oligomeric ferric citrate species. PLoS ONE 2013, 8, e79870. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Wright, T.L.; Ma, W.L.; Weisiger, R.A. Efficient clearance of non-transferrin-bound iron by rat liver. Implications for hepatic iron loading in iron overload states. J. Clin. Investig. 1985, 76, 1463–1470. [Google Scholar] [CrossRef] [Green Version]
- Graham, R.M.; Morgan, E.H.; Baker, E. Characterisation of citrate and iron citrate uptake by cultured rat hepatocytes. J. Hepatol. 1998, 29, 603–613. [Google Scholar] [CrossRef]
- Wright, T.L.; Brissot, P.; Ma, W.L.; Weisiger, R.A. Characterization of non-transferrin-bound iron clearance by rat liver. J. Biol. Chem. 1986, 261, 10909–10914. [Google Scholar] [CrossRef]
- Craven, C.M.; Alexander, J.; Eldridge, M.; Kushner, J.P.; Bernstein, S.; Kaplan, J. Tissue distribution and clearance kinetics of non-transferrin-bound iron in the hypotransferrinemic mouse: A rodent model for hemochromatosis. Proc. Natl. Acad. Sci. USA 1987, 84, 3457–3461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkitkasemwong, S.; Wang, C.-Y.; Coffey, R.; Zhang, W.; Chan, A.; Biel, T.; Kim, J.-S.; Hojyo, S.; Fukada, T.; Knutson, M.D. SLC39A14 Is Required for the Development of Hepatocellular Iron Overload in Murine Models of Hereditary Hemochromatosis. Cell Metab. 2015, 22, 138–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-Y.; Knutson, M.D. Hepatocyte divalent metal-ion transporter-1 is dispensable for hepatic iron accumulation and non-transferrin-bound iron uptake in mice. Hepatology 2013, 58, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Coffey, R.; Knutson, M.D. The plasma membrane metal-ion transporter ZIP14 contributes to nontransferrin-bound iron uptake by human β-cells. Am. J. Physiol. Cell Physiol. 2017, 312, C169–C175. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Aydemir, F.; Nam, H.; Knutson, M.D.; Cousins, R.J. Zip14 (Slc39a14) mediates non-transferrin-bound iron uptake into cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13612–13617. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Kosman, D.J. Molecular mechanisms of non-transferrin-bound and transferring-bound iron uptake in primary hippocampal neurons. J. Neurochem. 2015, 133, 668–683. [Google Scholar] [CrossRef] [Green Version]
- Oudit, G.Y.; Sun, H.; Trivieri, M.G.; Koch, S.E.; Dawood, F.; Ackerley, C.; Yazdanpanah, M.; Wilson, G.J.; Schwartz, A.; Liu, P.P.; et al. L-type Ca2+ channels provide a major pathway for iron entry into cardiomyocytes in iron-overload cardiomyopathy. Nat. Med. 2003, 9, 1187–1194. [Google Scholar] [CrossRef]
- Tsushima, R.G.; Wickenden, A.D.; Bouchard, R.A.; Oudit, G.Y.; Liu, P.P.; Backx, P.H. Modulation of iron uptake in heart by L-type Ca2+ channel modifiers: Possible implications in iron overload. Circ. Res. 1999, 84, 1302–1309. [Google Scholar] [CrossRef] [Green Version]
- Parkes, J.G.; Olivieri, N.F.; Templeton, D.M. Characterization of Fe2+ and Fe3+ transport by iron-loaded cardiac myocytes. Toxicology 1997, 117, 141–151. [Google Scholar] [CrossRef]
- Tulpule, K.; Robinson, S.R.; Bishop, G.M.; Dringen, R. Uptake of ferrous iron by cultured rat astrocytes. J. Neurosci. Res. 2010, 88, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Tripathi, A.; Qian, J.; Beserra, A.; Suda, S.; McElwee, M.; Turner, J.; Hopfer, U.; Singh, N. Prion protein promotes kidney iron uptake via its ferrireductase activity. J. Biol. Chem. 2015, 290, 5512–5522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.K.; Haldar, S.; Qian, J.; Beserra, A.; Suda, S.; Singh, A.; Hopfer, U.; Chen, S.G.; Garrick, M.D.; Turner, J.R.; et al. Prion protein functions as a ferrireductase partner for ZIP14 and DMT1. Free. Radic. Biol. Med. 2015, 84, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, Z.L.; Durley, A.P.; Man, T.K.; Gitlin, J.D. Targeted gene disruption reveals an essential role for ceruloplasmin in cellular iron efflux. Proc. Natl. Acad. Sci. USA 1999, 96, 10812–10817. [Google Scholar] [CrossRef] [Green Version]
- Vulpe, C.D.; Kuo, Y.M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar] [CrossRef]
- Healy, J.; Tipton, K. Ceruloplasmin and what it might do. J. Neural Transm. 2007, 114, 777–781. [Google Scholar] [CrossRef]
- Crichton, R.R.; Pierre, J.L. Old iron, young copper: From Mars to Venus. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2001, 14, 99–112. [Google Scholar] [CrossRef] [Green Version]
- May, P.M.; Linder, P.W.; Williams, D.R. Computer simulation of metal-ion equilibria in biofluids: Models for the low-molecular-weight complex distribution of calcium(II), magnesium(II), manganese(II), iron(III), copper(II), zinc(II), and lead(II) ions in human blood plasma. J. Chem. Soc. Dalton Trans. 1977, 6, 588–595. [Google Scholar] [CrossRef]
- Grootveld, M.; Bell, J.D.; Halliwell, B.; Aruoma, O.I.; Bomford, A.; Sadler, P.J. Non-transferrin-bound iron in plasma or serum from patients with idiopathic hemochromatosis. Characterization by high performance liquid chromatography and nuclear magnetic resonance spectroscopy. J. Biol. Chem. 1989, 264, 4417–4422. [Google Scholar] [CrossRef]
- Simpson, R.J.; Cooper, C.E.; Raja, K.B.; Halliwell, B.; Evans, P.J.; Aruoma, O.I.; Singh, S.; Konijn, A.M. Non-transferrin-bound iron species in the serum of hypotransferrinaemic mice. Biochim. Biophys. Acta 1992, 1156, 19–26. [Google Scholar] [CrossRef]
- Evans, R.W.; Rafique, R.; Zarea, A.; Rapisarda, C.; Cammack, R.; Evans, P.J.; Porter, J.B.; Hider, R.C. Nature of non-transferrin-bound iron: Studies on iron citrate complexes and thalassemic sera. J. Biol. Inorg. Chem. 2008, 13, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Al-Harthi, S.; Lachowicz, J.I.; Nowakowski, M.E.; Jaremko, M.; Jaremko, Ł. Towards the functional high-resolution coordination chemistry of blood plasma human serum albumin. J. Inorg. Biochem. 2019, 198, 110716. [Google Scholar] [CrossRef] [PubMed]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C. Nature of nontransferrin-bound iron. Eur. J. Clin. Investig. 2002, 32, 50–54. [Google Scholar] [CrossRef]
- Silva, A.M.N.; Hider, R.C. Influence of non-enzymatic post-translation modifications on the ability of human serum albumin to bind iron. Implications for non-transferrin-bound iron speciation. Biochim. Biophys. Acta 2009, 1794, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Ariga, T.; Hazama, K.; Yanagisawa, S.; Yoneyama, T. Chemical forms of iron in xylem sap from graminaceous and non-graminaceous plants. Soil Sci. Plant Nutr. 2014, 60, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Rellán-Alvarez, R.; Giner-Martínez-Sierra, J.; Orduna, J.; Orera, I.; Rodríguez-Castrillón, J.A.; García-Alonso, J.I.; Abadía, J.; Alvarez-Fernández, A. Identification of a tri-iron(III), tri-citrate complex in the xylem sap of iron-deficient tomato resupplied with iron: New insights into plant iron long-distance transport. Plant Cell Physiol. 2010, 51, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.N.; Kong, X.; Hider, R.C. Determination of the pKa value of the hydroxyl group in the alpha-hydroxycarboxylates citrate, malate and lactate by 13C NMR: Implications for metal coordination in biological systems. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2009, 22, 771–778. [Google Scholar] [CrossRef]
- Gautier-Luneau, I.; Merle, C.; Phanon, D.; Lebrun, C.; Biaso, F.; Serratrice, G.; Pierre, J.-L. New Trends in the Chemistry of Iron(III) Citrate Complexes: Correlations between X-ray Structures and Solution Species Probed by Electrospray Mass Spectrometry and Kinetics of Iron Uptake from Citrate by Iron Chelators. Chem.—Eur. J. 2005, 11, 2207–2219. [Google Scholar] [CrossRef]
- Bino, A.; Shweky, I.; Cohen, S.; Bauminger, E.R.; Lippard, S.J. A Novel Nonairon(III) Citrate Complex: A “Ferric Triple-Decker”. Inorg. Chem. 1998, 37, 5168–5172. [Google Scholar] [CrossRef]
- Matzapetakis, M.; Raptopoulou, C.P.; Tsohos, A.; Papaefthymiou, V.; Moon, N.; Salifoglou, A. Synthesis, Spectroscopic and Structural Characterization of the First Mononuclear, Water Soluble Iron−Citrate Complex, (NH4)5Fe(C6H4O7)2·2H2O. J. Am. Chem. Soc. 1998, 120, 13266–13267. [Google Scholar] [CrossRef]
- Shweky, I.; Bino, A.; Goldberg, D.P.; Lippard, S.J. Syntheses, Structures, and Magnetic Properties of Two Dinuclear Iron(III) Citrate Complexes. Inorg. Chem. 1994, 33, 5161–5162. [Google Scholar] [CrossRef]
- Fukushima, T.; Sia, A.K.; Allred, B.E.; Nichiporuk, R.; Zhou, Z.; Andersen, U.N.; Raymond, K.N. Bacillus cereus iron uptake protein fishes out an unstable ferric citrate trimer. Proc. Natl. Acad. Sci. USA 2012, 109, 16829–16834. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.N.; Kong, X.; Parkin, M.C.; Cammack, R.; Hider, R.C. Iron(III) citrate speciation in aqueous solution. Dalton Trans. 2009, 40, 8616–8625. [Google Scholar] [CrossRef]
- Yue, W.W.; Grizot, S.; Buchanan, S.K. Structural Evidence for Iron-free Citrate and Ferric Citrate Binding to the TonB-dependent Outer Membrane Transporter FecA. J. Mol. Biol. 2003, 332, 353–368. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Chakraborty, R.; Smith, B.S.; Esser, L.; van der Helm, D.; Deisenhofer, J. Structural Basis of Gating by the Outer Membrane Transporter FecA. Science 2002, 295, 1715–1719. [Google Scholar] [CrossRef]
- Thorstensen, K.; Romslo, I. Uptake of iron from transferrin by isolated hepatocytes. Biochim. Biophys. Acta 1984, 804, 200–208. [Google Scholar] [CrossRef]
- Trinder, D.; Morgan, E. Mechanisms of ferric citrate uptake by human hepatoma cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 1998, 275, G279–G286. [Google Scholar] [CrossRef]
- Kosman, D.J. A holistic view of mammalian (vertebrate) cellular iron uptake. Metallomics 2020, 12, 1323–1334. [Google Scholar] [CrossRef]
- Sherman, H.G.; Jovanovic, C.; Stolnik, S.; Baronian, K.; Downard, A.J.; Rawson, F.J. New Perspectives on Iron Uptake in Eukaryotes. Front. Mol. Biosci. 2018, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Vukosav, P.; Mlakar, M.; Tomišić, V. Revision of iron(III)–citrate speciation in aqueous solution. Voltammetric and spectrophotometric studies. Anal. Chim. Acta 2012, 745, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Adam, F.I.; Bounds, P.L.; Kissner, R.; Koppenol, W.H. Redox properties and activity of iron-citrate complexes: Evidence for redox cycling. Chem. Res. Toxicol. 2015, 28, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Bal, W.; Sokołowska, M.; Kurowska, E.; Faller, P. Binding of transition metal ions to albumin: Sites, affinities and rates. Biochim. Biophys. Acta 2013, 1830, 5444–5455. [Google Scholar] [CrossRef]
- Sadler, P.J.; Tucker, A.; Viles, J.H. Involvement of a lysine residue in the N-terminal Ni2+ and Cu2+ binding site of serum albumins. Eur. J. Biochem. 1994, 220, 193–200. [Google Scholar] [CrossRef]
- Lu, J.; Stewart, A.J.; Sadler, P.J.; Pinheiro, T.J.T.; Blindauer, C.A. Albumin as a zinc carrier: Properties of its high-affinity zinc-binding site. Biochem. Soc. Trans. 2008, 36, 1317–1321. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.J.; Blindauer, C.A.; Berezenko, S.; Sleep, D.; Sadler, P.J. Interdomain zinc site on human albumin. Proc. Natl. Acad. Sci. USA 2003, 100, 3701. [Google Scholar] [CrossRef] [Green Version]
- Sadler, P.J.; Viles, J.H. 1H and (113)Cd NMR Investigations of Cd(2+) and Zn(2+) Binding Sites on Serum Albumin: Competition with Ca(2+), Ni(2+), Cu(2+), and Zn(2+). Inorg. Chem. 1996, 35, 4490–4496. [Google Scholar] [CrossRef]
- Goumakos, W.; Laussac, J.P.; Sarkar, B. Binding of cadmium(II) and zinc(II) to human and dog serum albumins. An equilibrium dialysis and 113Cd-NMR study. Biochem. Cell Biol. 1991, 69, 809–820. [Google Scholar] [CrossRef]
- Ivanov, A.I.; Christodoulou, J.; Parkinson, J.A.; Barnham, K.J.; Tucker, A.; Woodrow, J.; Sadler, P.J. Cisplatin binding sites on human albumin. J. Biol. Chem. 1998, 273, 14721–14730. [Google Scholar] [CrossRef] [Green Version]
- Talib, J.; Beck, J.L.; Ralph, S.F. A mass spectrometric investigation of the binding of gold antiarthritic agents and the metabolite [Au(CN)2]− to human serum albumin. JBIC J. Biol. Inorg. Chem. 2006, 11, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, X.-P.; Chen; Xia, Y.-L.; Jiang, Y. Human Serum Albumin−Mercurial Species Interactions. J. Proteome Res. 2007, 6, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal structure of human serum albumin at 2.5 Å resolution. Protein Eng. Des. Sel. 1999, 12, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Kragh-Hansen, U.; Vorum, H. Quantitative analyses of the interaction between calcium ions and human serum albumin. Clin. Chem. 1993, 39, 202–208. [Google Scholar] [CrossRef]
- Vorum, H.; Fisker, K.; Otagiri, M.; Pedersen, A.O.; Kragh-Hansen, U. Calcium ion binding to clinically relevant chemical modifications of human serum albumin. Clin. Chem. 1995, 41, 1654–1661. [Google Scholar] [CrossRef]
- Anghileri, L.J. Fate of intravenously injected iron compounds: Ferric-fructose complex, iron-EDTA, ferric hydroxide and iron-albumin labeled with 59Fe. Biochem. Pharmacol. 1967, 16, 2033–2036. [Google Scholar] [CrossRef]
- Loban, A.; Kime, R.; Powers, H. Iron-Binding Antioxidant Potential of Plasma Albumin. Clin. Sci. 1997, 93, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, L.; Shen, D.; Wu, H.; Liu, Q. Oxygen-dependent Oxidation of Fe(II) to Fe(III) and Interaction of Fe(III) with Bovine Serum Albumin, Leading to a Hysteretic Effect on the Fluorescence of Bovine Serum Albumin. J. Fluoresc. 2008, 18, 193–201. [Google Scholar] [CrossRef]
- Coddington, A.; Perkins, D.J. The binding of Fe+++ to native and chemically modified human serum albumin in the presence of sodium citrate. Biochim. Biophys. Acta 1960, 44, 361–363. [Google Scholar] [CrossRef]
- Løvstad, R.A. Interaction of serum albumin with the Fe(III)-citrate complex. Int. J. Biochem. 1993, 25, 1015–1017. [Google Scholar] [CrossRef]
- Spiro, T.G.; Pape, L.; Saltman, P. Hydrolytic polymerization of ferric citrate. I. Chemistry of the polymer. J. Am. Chem. Soc. 1967, 89, 5555–5559. [Google Scholar] [CrossRef]
- Singh, S.; Hider, R.C.; Porter, J.B. A direct method for quantification of non-transferrin-bound iron. Anal. Biochem. 1990, 186, 320–323. [Google Scholar] [CrossRef]
- Evans, P.J.; Halliwell, B. Measurement of iron and copper in biological systems: Bleomycin and copper-phenanthroline assays. Methods Enzymol. 1994, 233, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Podinovskaia, M.; Evans, P.J.; Emma, G.; Schaible, U.E.; Porter, J.; Hider, R.C. A novel method for non-transferrin-bound iron quantification by chelatable fluorescent beads based on flow cytometry. Biochem. J. 2014, 463, 351–362. [Google Scholar] [CrossRef]
- Garbowski, M.W.; Ma, Y.; Fucharoen, S.; Srichairatanakool, S.; Hider, R.; Porter, J.B. Clinical and methodological factors affecting non-transferrin-bound iron values using a novel fluorescent bead assay. Transl. Res. 2016, 177, 19–30.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, E.M.; Hendriks, J.C.; van Tits, B.L.; Evans, P.J.; Breuer, W.; Liu, D.Y.; Jansen, E.H.; Jauhiainen, K.; Sturm, B.; Porter, J.B.; et al. Results of an international round robin for the quantification of serum non-transferrin-bound iron: Need for defining standardization and a clinically relevant isoform. Anal. Biochem. 2005, 341, 241–250. [Google Scholar] [CrossRef]
- Fawwaz, R.A.; Winchell, H.S.; Pollycove, M.; Sargent, T. Hepatic Iron Deposition in Humans: I. First-Pass Hepatic Deposition of Intestinally Absorbed Iron in Patients with Low Plasma Latent Iron-Binding Capacity. Blood 1967, 30, 417–424. [Google Scholar] [CrossRef]
- Noetzli, L.J.; Papudesi, J.; Coates, T.D.; Wood, J.C. Pancreatic iron loading predicts cardiac iron loading in thalassemia major. Blood 2009, 114, 4021–4026. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, V.N.; McDonald, C.J.; Ostini, L.; Lusby, P.E.; Wockner, L.F.; Ramm, G.A.; Wallace, D.F. Hepatic iron deposition does not predict extrahepatic iron loading in mouse models of hereditary hemochromatosis. Am. J. Pathol. 2012, 181, 1173–1179. [Google Scholar] [CrossRef]
- Kishimoto, M.; Endo, H.; Hagiwara, S.; Miwa, A.; Noda, M. Immunohistochemical findings in the pancreatic islets of a patient with transfusional iron overload and diabetes: Case report. J. Med. Investig. 2010, 57, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.-P.; Hayashi, K. Selective iron deposition in pancreatic islet B cells of transfusional iron-overloaded autopsy cases. Pathol. Int. 1994, 44, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Marku, A.; Galli, A.; Marciani, P.; Dule, N.; Perego, C.; Castagna, M. Iron Metabolism in Pancreatic Beta-Cell Function and Dysfunction. Cells 2021, 10, 2841. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Parkes, J.G.; Templeton, D.M. Differential accumulation of non-transferrin-bound iron by cardiac myocytes and fibroblasts. J. Mol. Cell. Cardiol. 2003, 35, 505–514. [Google Scholar] [CrossRef]
- Link, G.; Athias, P.; Grynberg, A.; Pinson, A.; Hershko, C. Effect of iron loading on transmembrane potential, contraction, and automaticity of rat ventricular muscle cells in culture. J. Lab. Clin. Med. 1989, 113, 103–111. [Google Scholar]
- Leoncini, S.; Rossi, V.; Signorini, C.; Tanganelli, I.; Comporti, M.; Ciccoli, L. Oxidative stress, erythrocyte ageing and plasma non-protein-bound iron in diabetic patients. Free Radic. Res. 2008, 42, 716–724. [Google Scholar] [CrossRef]
- Sulieman, M.; Asleh, R.; Cabantchik, Z.I.; Breuer, W.; Aronson, D.; Suleiman, A.; Miller-Lotan, R.; Hammerman, H.; Levy, A.P. Serum chelatable redox-active iron is an independent predictor of mortality after myocardial infarction in individuals with diabetes. Diabetes Care 2004, 27, 2730–2732. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Real, J.M.; Lopez-Bermejo, A.; Ricart, W. Cross-talk between iron metabolism and diabetes. Diabetes 2002, 51, 2348–2354. [Google Scholar] [CrossRef] [Green Version]
- Wlazlo, N.; van Greevenbroek, M.M.; Ferreira, I.; Jansen, E.H.; Feskens, E.J.; van der Kallen, C.J.; Schalkwijk, C.G.; Bravenboer, B.; Stehouwer, C.D. Iron metabolism is associated with adipocyte insulin resistance and plasma adiponectin: The Cohort on Diabetes and Atherosclerosis Maastricht (CODAM) study. Diabetes Care 2013, 36, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Wlazlo, N.; van Greevenbroek, M.M.; Ferreira, I.; Jansen, E.H.; Feskens, E.J.; van der Kallen, C.J.; Schalkwijk, C.G.; Bravenboer, B.; Stehouwer, C.D. Iron metabolism is prospectively associated with insulin resistance and glucose intolerance over a 7-year follow-up period: The CODAM study. Acta Diabetol. 2015, 52, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Ayton, S.; Wang, Y.; Diouf, I.; Schneider, J.A.; Brockman, J.; Morris, M.C.; Bush, A.I. Brain iron is associated with accelerated cognitive decline in people with Alzheimer pathology. Mol. Psychiatry 2020, 25, 2932–2941. [Google Scholar] [CrossRef]
- van Bergen, J.M.; Li, X.; Hua, J.; Schreiner, S.J.; Steininger, S.C.; Quevenco, F.C.; Wyss, M.; Gietl, A.F.; Treyer, V.; Leh, S.E.; et al. Colocalization of cerebral iron with Amyloid beta in Mild Cognitive Impairment. Sci. Rep. 2016, 6, 35514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, J.; Collingwood, J.F.; Tjendana-Tjhin, V.; Brooks, J.; Lermyte, F.; Plascencia-Villa, G.; Hands-Portman, I.; Dobson, J.; Perry, G.; Telling, N.D. Nanoscale synchrotron X-ray speciation of iron and calcium compounds in amyloid plaque cores from Alzheimer’s disease subjects. Nanoscale 2018, 10, 11782–11796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plascencia-Villa, G.; Ponce, A.; Collingwood, J.F.; Arellano-Jiménez, M.J.; Zhu, X.; Rogers, J.T.; Betancourt, I.; José-Yacamán, M.; Perry, G. High-resolution analytical imaging and electron holography of magnetite particles in amyloid cores of Alzheimer’s disease. Sci. Rep. 2016, 6, 24873. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.; Jeandriens, J.; Parkes, H.G.; So, P.-W. Iron dyshomeostasis, lipid peroxidation and perturbed expression of cystine/glutamate antiporter in Alzheimer’s disease: Evidence of ferroptosis. Redox Biol. 2020, 32, 101494. [Google Scholar] [CrossRef]
- Jakaria, M.; Belaidi, A.A.; Bush, A.I.; Ayton, S. Ferroptosis as a mechanism of neurodegeneration in Alzheimer’s disease. J. Neurochem. 2021, 159, 804–825. [Google Scholar] [CrossRef]
- Dexter, D.T.; Wells, F.R.; Agid, F.; Agid, Y.; Lees, A.J.; Jenner, P.; Marsden, C.D. Increased nigral iron content in postmortem parkinsonian brain. Lancet 1987, 330, 1219–1220. [Google Scholar] [CrossRef]
- Dexter, D.T.; Wells, F.R.; Lees, A.J.; Agid, F.; Agid, Y.; Jenner, P.; Marsden, C.D. Increased nigral iron content and alterations in other metal ions occurring in brain in Parkinson’s disease. J. Neurochem. 1989, 52, 1830–1836. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Brandel, J.P.; Galle, P.; Javoy-Agid, F.; Agid, Y. Iron and aluminum increase in the substantia nigra of patients with Parkinson’s disease: An X-ray microanalysis. J. Neurochem. 1991, 56, 446–451. [Google Scholar] [CrossRef]
- Kalpouzos, G.; Mangialasche, F.; Falahati, F.; Laukka, E.J.; Papenberg, G. Contributions of HFE polymorphisms to brain and blood iron load, and their links to cognitive and motor function in healthy adults. Neuropsychopharmacol. Rep. 2021, 41, 393–404. [Google Scholar] [CrossRef]
- Nandar, W.; Connor, J.R. HFE gene variants affect iron in the brain. J. Nutr. 2011, 141, 729s–739s. [Google Scholar] [CrossRef]
- Kaplan, J.; Craven, C.; Alexander, J.; Kushner, J.; Lamb, J.; Bernstein, S. Regulation of the Distribution of Tissue Iron. Ann. N. Y. Acad. Sci. 1988, 526, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Metafratzi, Z.; Argyropoulou, M.I.; Kiortsis, D.N.; Tsampoulas, C.; Chaliassos, N.; Efremidis, S.C. T2 relaxation rate of basal ganglia and cortex in patients with β-thalassaemia major. Br. J. Radiol. 2001, 74, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Chan, G.C.F.; Chu, J.; Chan, Q.; Ha, S.Y.; Moseley, M.E.; Khong, P.L. MR Quantitative susceptibility imaging for the evaluation of iron loading in the brains of patients with β-thalassemia major. Am. J. Neuroradiol. 2014, 35, 1085–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.K.; Karmakar, S.; Asthana, A.; Ashok, A.; Desai, V.; Baksi, S.; Singh, N. Transport of Non-Transferrin Bound Iron to the Brain: Implications for Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 58, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, M.W.B. Transport of iron in the blood-brain-cerebrospinal fluid system. J. Neurochem. 1997, 69, 443–454. [Google Scholar] [CrossRef]
- Bishop, G.M.; Dang, T.N.; Dringen, R.; Robinson, S.R. Accumulation of Non-Transferrin-Bound Iron by Neurons, Astrocytes, and Microglia. Neurotox. Res. 2011, 19, 443–451. [Google Scholar] [CrossRef]
- Dziuba, N.; Hardy, J.; Lindahl, P.A. Low-molecular-mass iron in healthy blood plasma is not predominately ferric citrate. Metallomics 2018, 10, 802–817. [Google Scholar] [CrossRef]
- Løvstad, R.A. A kinetic study of the coupled iron-ceruloplasmin catalyzed oxidation of ascorbate in the presence of albumin. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 1995, 8, 328–331. [Google Scholar] [CrossRef]
- Matias, C.; Belnap, D.W.; Smith, M.T.; Stewart, M.G.; Torres, I.F.; Gross, A.J.; Watt, R.K. Citrate and albumin facilitate transferrin iron loading in the presence of phosphate. J. Inorg. Biochem. 2017, 168, 107–113. [Google Scholar] [CrossRef]
- Sohn, Y.S.; Ghoti, H.; Breuer, W.; Rachmilewitz, E.; Attar, S.; Weiss, G.; Cabantchik, Z.I. The role of endocytic pathways in cellular uptake of plasma non-transferrin iron. Haematologica 2012, 97, 670–678. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of the structure of citrate and ferric citrate complexes in aqueous solution. (A) Citrate anion. (B) [FeCit2]5−. (C) [Fe2Cit2]2−. (D) [Fe3Cit3]3−. (E) alternative structure for [Fe3Cit3]3−. Structures C, D and E are adapted from Gautier-Luneau et al. [71], structure E has been proposed by Fukushima et al. [75].
Figure 1.
Schematic representation of the structure of citrate and ferric citrate complexes in aqueous solution. (A) Citrate anion. (B) [FeCit2]5−. (C) [Fe2Cit2]2−. (D) [Fe3Cit3]3−. (E) alternative structure for [Fe3Cit3]3−. Structures C, D and E are adapted from Gautier-Luneau et al. [71], structure E has been proposed by Fukushima et al. [75].

Figure 2.
Human serum albumin structure (PDB: 1BM0 [94]). (A) Known metal ion binding sites in HSA: NTS (N-terminal site), MBS (Metal Binding Site A) and Cys34 (reduced cysteine at position 34). (B) Surface map from HSA, highlighting sidechain oxygen atoms from aspartate and glutamate residues (red) and sidechain nitrogen atoms from arginine and lysine residues (blue). The 4 N-terminal amino acid residues in the protein sequence were absent from the crystallographic structure and were manually modelled for this representation.
Figure 2.
Human serum albumin structure (PDB: 1BM0 [94]). (A) Known metal ion binding sites in HSA: NTS (N-terminal site), MBS (Metal Binding Site A) and Cys34 (reduced cysteine at position 34). (B) Surface map from HSA, highlighting sidechain oxygen atoms from aspartate and glutamate residues (red) and sidechain nitrogen atoms from arginine and lysine residues (blue). The 4 N-terminal amino acid residues in the protein sequence were absent from the crystallographic structure and were manually modelled for this representation.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Silva, A.M.N.; Rangel, M. The (Bio)Chemistry of Non-Transferrin-Bound Iron. Molecules 2022, 27, 1784. https://doi.org/10.3390/molecules27061784
AMA Style
Silva AMN, Rangel M. The (Bio)Chemistry of Non-Transferrin-Bound Iron. Molecules. 2022; 27(6):1784. https://doi.org/10.3390/molecules27061784
Chicago/Turabian StyleSilva, André M. N., and Maria Rangel. 2022. "The (Bio)Chemistry of Non-Transferrin-Bound Iron" Molecules 27, no. 6: 1784. https://doi.org/10.3390/molecules27061784