
Isolation and Characterization of Bacteriocin-Producing Lacticaseibacillus rhamnosus XN2 from Yak Yoghurt and Its Bacteriocin

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Screening of Bacteriocin Production by LABs Isolated from Yak Yoghurt
2.2. Bacteriocin Characterization of XN2 Strain
2.2.1. Growth and Bacteriocin Production Curve
2.2.2. Bacteriocin Stability
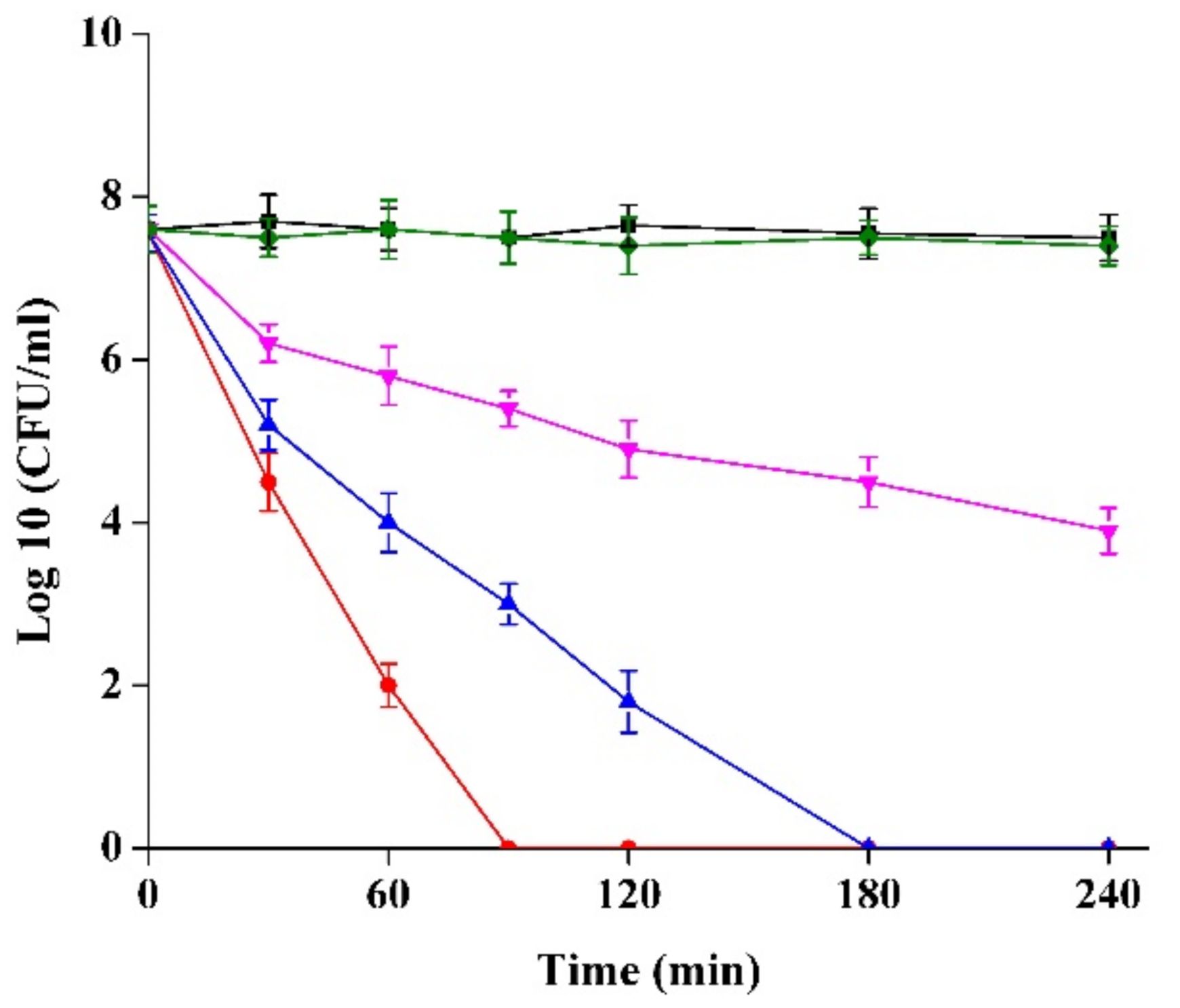
2.3. Kill Kinetics Curve of Semi-Purified CFS of Strain XN2 against S. aureus
2.4. Effect of Semi-Purified CFS of Strain XN2 on the Integrity of S. aureus
2.5. Effect of Semi-Purified CFS of Strain XN2 on α-Haemolysin Secreted by S. aureus
2.6. Effect of Semi-Purified CFS of Strain XN2 on the Biofilm Formation of S. aureus
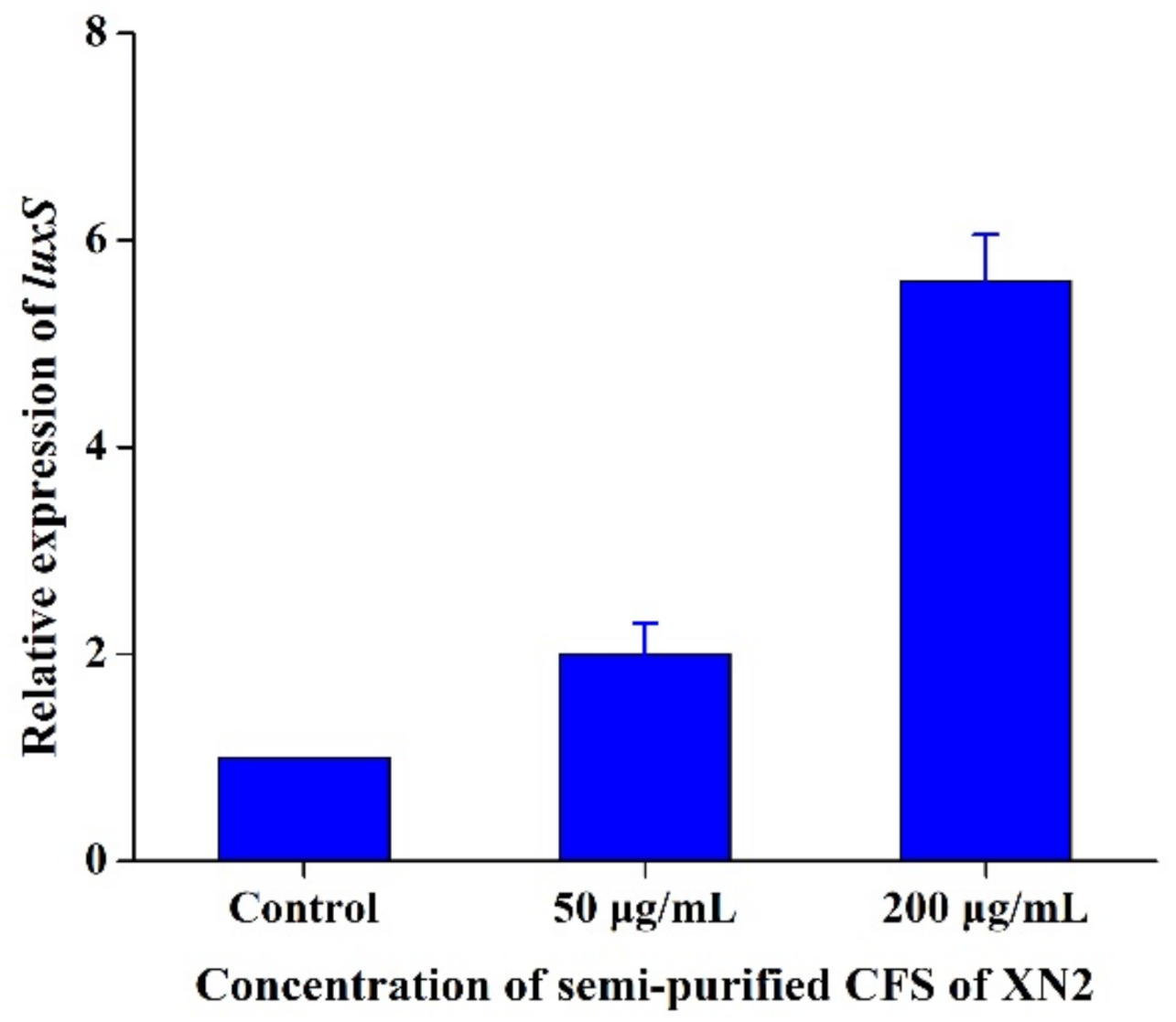
2.7. Effect of Semi-Purified CFS of Strain XN2 on the QS System of S. aureus
2.8. Purification of Bacteriocin
3. Materials and Methods
3.1. Isolation and Identification of Bacteriocin-Producing Stain
3.2. Bacteriocins Production by Strain XN2
3.3. Semi-Purification of CFS of Strain XN2
3.4. Antimicrobial Spectrum and Stability Testing
3.5. Time-Killing Kinetics
3.6. Flow Cytometry (FCM) Analysis
3.7. α-Haemolysin Secretion
3.8. Biofilm Formation
3.9. RT-PCR
3.10. Bacteriocin Purification
3.11. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ding, W.; Shi, C.; Chen, M.; Zhou, J.; Long, R.; Guo, X. Screening for lactic acid bacteria in traditional fermented Tibetan yak milk and evaluating their probiotic and cholesterol-lowering potentials in rats fed a high-cholesterol diet. J. Funct. Foods 2017, 32, 324–332. [Google Scholar] [CrossRef]
- Luo, F.; Feng, S.; Sun, Q.; Xiang, W.; Zhao, J.; Zhang, J.; Yang, Z. Screening for bacteriocin-producing lactic acid bacteria from kurut, atraditional naturally-fermented yak milk from Qinghai–Tibet plateau. Food Control 2011, 22, 50–53. [Google Scholar] [CrossRef]
- Ji, X.; Li, X.; Ma, Y.; Li, D. Differences in proteomic profiles of milk fat globule membrane in yak and cow milk. Food Chem. 2017, 221, 1822–1827. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Duan, S.; Wang, Q.; Ji, R.; Cao, Y.; Miao, J. Effects of the natural antimicrobial substance from Lactobacillus paracasei FX-6 on shelf life and microbial composition in chicken breast during refrigerated storage. Food Control 2020, 109, 106906. [Google Scholar] [CrossRef]
- Xu, C.; Fu, Y.; Liu, F.; Liu, Z.; Ma, J.; Jiang, R.; Song, C.; Jiang, Z.; Hou, J. Purification and antimicrobial mechanism of a novel bacteriocin produced by Lacticaseibacillus rhamnosus 1.0320—Sciencedirect. LWT 2021, 137, 110338. [Google Scholar] [CrossRef]
- Mirkovic, N.; Kulas, J.; Miloradovic, Z.; Miljkovic, M.; Tucovic, D.; Miocinovic, J.; Jovcic, B.; Mirkov, I.; Kojic, M. Lactolisterin BU-producer Lactococcus lactis subsp. lactis BGBU1-4: Bio-control of Listeria monocytogenes and Staphylocococcus aureus in fresh soft cheese and effect on immunological response of rats. Food Control 2020, 111, 107076. [Google Scholar] [CrossRef]
- Todorov, S.D.; Prévost, H.; Lebois, M.; Dousset, X.; LeBlanc, J.G.; Franco, B.D. Bacteriocinogenic Lacticaseibacillus rhamnosus ST16Pa isolated from papaya (Carica papaya)—From isolation to application: Characterization of a bacteriocin. Food Res. Int. 2011, 44, 1351–1363. [Google Scholar] [CrossRef]
- Srinivasan, R.; Kumawat, D.K.; Kumar, S.; Saxena, A.K. Purification and characterization of a bacteriocin from Lactobcillus rhamnosus L34. Ann. Microbiol. 2013, 3, 37–392. [Google Scholar]
- Winkelströter, L.K.; Tulini, F.L.; De Martinis, E.C. Identification of the bacteriocin produced by cheese isolate Lactiplantibacillus paraplantarum, FT259 and its potential influence on Listeria monocytogenes biofilm formation. LWT-Food Sci. Technol. 2015, 64, 586–592. [Google Scholar] [CrossRef]
- Chopra, L.; Singh, G.; Kumar, J.K.; Sahoo, D.K. Sonorensin: A new bacteriocin with potential of an anti-biofilm agent and a food biopreservative. Sci. Report. 2015, 5, 13412. [Google Scholar] [CrossRef] [Green Version]
- Al-Gburi, A.; Zehm, S.; Netrebov, V. Subtilosin Prevents Biofilm Formation by Inhibiting Bacterial Quorum Sensing. Probiotics Antimicrob. Proteins 2016, 9, 81–90. [Google Scholar] [CrossRef]
- Mhatre, E.; Monterrosa, R.G.; Kovács, Á.T. From environmental signals to regulators: Modulation of biofilm development in Grampositive bacteria. J. Basic Microbiol. 2014, 54, 616–632. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Li, X.; Han, H.; Tao, Y. Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 2018, 84, 111–117. [Google Scholar] [CrossRef]
- Yue, T.; Pei, J.; Yuan, Y. Purification and Characterization of Anti-Alicyclobacillus Bacteriocin Produced by Lacticaseibacillus rhamnosus. J. Food Prot. 2013, 76, 1575–1581. [Google Scholar] [CrossRef]
- Pei, J.; Feng, Z.; Ren, T.; Sun, H.; Han, H.; Jin, W.; Dang, J.; Tao, Y. Purification, characterization and application of a novel antimicrobial peptide from Andrias davidianus blood. Lett. Appl. Microbiol. 2018, 66, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.L.; Kato, N.; Matsumiya, Y.; Liu, C.X.; Kato, H.; Watanabe, K. Identification of Lactobacillus species of human origin by a commercial kit, API50CHL. J. Assoc. Rapid Method Autom. Microbiol. 1999, 10, 77–82. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Du, H.; Yang, J.; Lu, X.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lu, F. Purification, characterization, and mode of action of action of Plantaricin GZ1-27, a novel bactriocin against Bacillus cereus. J. Agric. Food Chem. 2018, 66, 4716–4724. [Google Scholar] [CrossRef]
- Pei, J.; Feng, Z.; Ren, T.; Jin, W.; Li, X.; Chen, D.; Dang, J. Selectivelyscreen the antibacterial peptide from the hydrolysates of highland barley. Eng. Life Sci. 2018, 18, 28–54. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, C.; Pan, J.; Deng, X.; Wang, J. Epigallocatechin gallate can attenuate human alveolar epithelial cell injury induced by alpha-haemolysin. Microb. Pathog. 2017, 115, 222–226. [Google Scholar] [CrossRef]
- Saporito, P.; Mouritzen, M.V.; Løbner-Olesen, A.; Jenssen, H. LL-37 fragments have antimicrobial activity against Staphylococcus epidermidis biofilms and wound healing potential in HaCaT cell line. J. Pept. Sci. 2018, 24, e3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Zhao, Y.; Tian, Y.; Zhang, W.; Li, X.; Jiang, X. The molecular mechanism of action of bactericidal gold nanoparticles on Escherichia coli. Biomaterials 2012, 33, 2327–2333. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, V.Q.; Camargo, A.C.; Todorov, S.D.; Nero, L.A. Novel bacteriocinogenic Enterococcus hirae and Pediococcus pentosaceus strains with antilisterial activity isolated from Brazilian artisanal cheese. J. Dairy Sci. 2017, 100, 2526–2535. [Google Scholar] [CrossRef] [PubMed]
- Rahmeh, R.; Akbar, A.; Kishk, M.; Onaizi, T.A.; Al-Shatti, A.; Shajan, A.; Akbar, B.; Al-Mutairi, S.; Yateem, A. Characterization of semipurified enterocins produced by Enterococcus faecium strains isolated from raw camel milk. J. Dairy Sci. 2018, 101, 4944–4952. [Google Scholar] [CrossRef]
- Hwanhlem, N.; Ivanova, T.; Biscola, V.; Choiset, Y.; Haertlé, T. Bacteriocin producing Enterococcus faecalis isolated from chicken gastrointestinal tract originating from Phitsanulok, Thailand: Isolation, screening, safety evaluation and probiotic properties. Food Control 2017, 78, 187–195. [Google Scholar] [CrossRef]
- Sabo, S.S.; Converti, A.; Ichiwaki, S.; Oliveira, R.P. Bacteriocin production by Lactobacillus plantarum ST16Pa in supplemented whey powder formulations. J. Dairy Sci. 2019, 102, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Ahn, H.; Kim, J.; Kim, W.J. Isolation and characterization of bacteriocin-producing Pediococcus acidilactici HW01 from malt and its potential to control beer spoilage lactic acid bacteria. Food Control 2017, 80, 59–66. [Google Scholar] [CrossRef]
- Goh, H.F.; Philip, K. Isolation and mode of action of bacteriocin BacC1 produced by nonpathogenic Enterococcus faecium C1. J. Dairy Sci. 2015, 98, 5080–5090. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.K.; Noll, S.K.; Cavera, V.L.; Chikindas, M.L. Improved antimicrobial activities of synthetic-hybrid bacteriocins designed from enterocin E50-52 and pediocin PA-1. Appl. Environ. Microbiol. 2015, 81, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Singh, T.; Malik, R. Antibacterial efficacy of Nisin, Pediocin 34 and Enterocin FH99 against Listeria monocytogenes and cross resistance of its bacteriocin resistant variants to common food preservatives. Braz. J. Microbiol. 2013, 14, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Heredia-Castro, P.Y.; Méndez-Romero, J.I.; Hernández-Mendoza, A.; Acedo-Félix, E.; González-Córdova, A.F.; Vallejo-Cordoba, B. Antimicrobial activity and partial characterization of bacteriocin-like inhibitory substances produced by Lactobacillus spp. isolated from artisanal Mexican cheese. J. Dairy Sci. 2015, 98, 8285–8293. [Google Scholar] [CrossRef] [Green Version]
- Panagiota, K.; Kyriakou, E.B.; Kristiansen, P.E.; Kaznessis, Y.N. Interactions of a class IIb bacteriocin with a model lipid bilayer, investigated through molecular dynamics simulations. Biochim. Biophys. Acta (BBA)—Biomembr. 2016, 1858, 824–835. [Google Scholar]
- Snyder, A.B.; Worobo, R.W. Chemical and genetic characterization of bacteriocins: Antimicrobial peptides for food safety. J. Sci. Food Agric. 2014, 94, 28–44. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Testing Condition | Antibacterial Activities of Semi-Purified CFS of Stain XN2 | |
|---|---|---|
| Lipase, α-Amylase, | + | |
| Proteinase K, Papain, α-Chymotrypsin, Trypsin, Pepsin | − | |
| Catalase | + | |
| Effect of temperature and storage on the activity of semi-purified CFS of stain XN2 | 60 °C, 80 °C, and 100 °C | + |
| 37 °C for 14 d | + | |
| 2 months at 4 °C | + | |
| Effect of pH on the activity of semi-purified CFS of stain XN2 | pH 2–8 | + |
| pH 9–10 | − | |
| The activity of semi-purified CFS of stain XN2 against gram-positive bacteria | Bacillus subtilis CICC 10034 | + |
| B. cereus CICC 2155 | + | |
| Micrococcus luteus CICC 10209 | + | |
| Brochothrix thermosphacta CICC 10509 | + | |
| Clostridium butyricum CICC 10350 | + | |
| Staphylococcus aureus CICC 10384 | + | |
| S. aureus CICC 10201 | + | |
| Methicillin-resistant S. aureus * | + | |
| Listeria innocua CICC 10416 | + | |
| L. monocytogenes CICC 21529 | + | |
| The activity of semi-purified CFS of stain XN2 against gram-negative bacteria | Escherichia coli CICC 10302 | + |
| E. coli CGMCC 3373 | + | |
| E. coli CICC 10300 | + | |
| Pseudomonas aeruginosa CICC 21636 | − | |
| Enterobacter cloacae CICC 21539 | − | |
| Salmonella paratyphi β CICC 10437 | − | |
| The activity of semi-purified CFS of stain XN2 against fungi | Aspergillus niger CICC 2124 | − |
| Candida albicans CICC 1965 | − | |
| Saccharomyces cerevisiae CICC 1002 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Wang, J.; Liu, Z.; Pei, J.; Brennan, C.; Abd El-Aty, A.M. Isolation and Characterization of Bacteriocin-Producing Lacticaseibacillus rhamnosus XN2 from Yak Yoghurt and Its Bacteriocin. Molecules 2022, 27, 2066. https://doi.org/10.3390/molecules27072066
Wei Y, Wang J, Liu Z, Pei J, Brennan C, Abd El-Aty AM. Isolation and Characterization of Bacteriocin-Producing Lacticaseibacillus rhamnosus XN2 from Yak Yoghurt and Its Bacteriocin. Molecules. 2022; 27(7):2066. https://doi.org/10.3390/molecules27072066
Chicago/Turabian StyleWei, Yonghua, Jinze Wang, Zhe Liu, Jinjin Pei, Charles Brennan, and A.M. Abd El-Aty. 2022. "Isolation and Characterization of Bacteriocin-Producing Lacticaseibacillus rhamnosus XN2 from Yak Yoghurt and Its Bacteriocin" Molecules 27, no. 7: 2066. https://doi.org/10.3390/molecules27072066