
Threonine Facilitates Cd Excretion by Increasing the Abundance of Gut Escherichia coli in Cd-Exposed Mice
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Effects of AAs on Cd Levels and GM in Cd-Exposed Mice
2.2. Thr Restored the Proportion of Escherichia-Shigella in GM of Cd-Exposed Mice
2.3. Thr and E. coli Attenuated the Gut Damages Induced by Cd
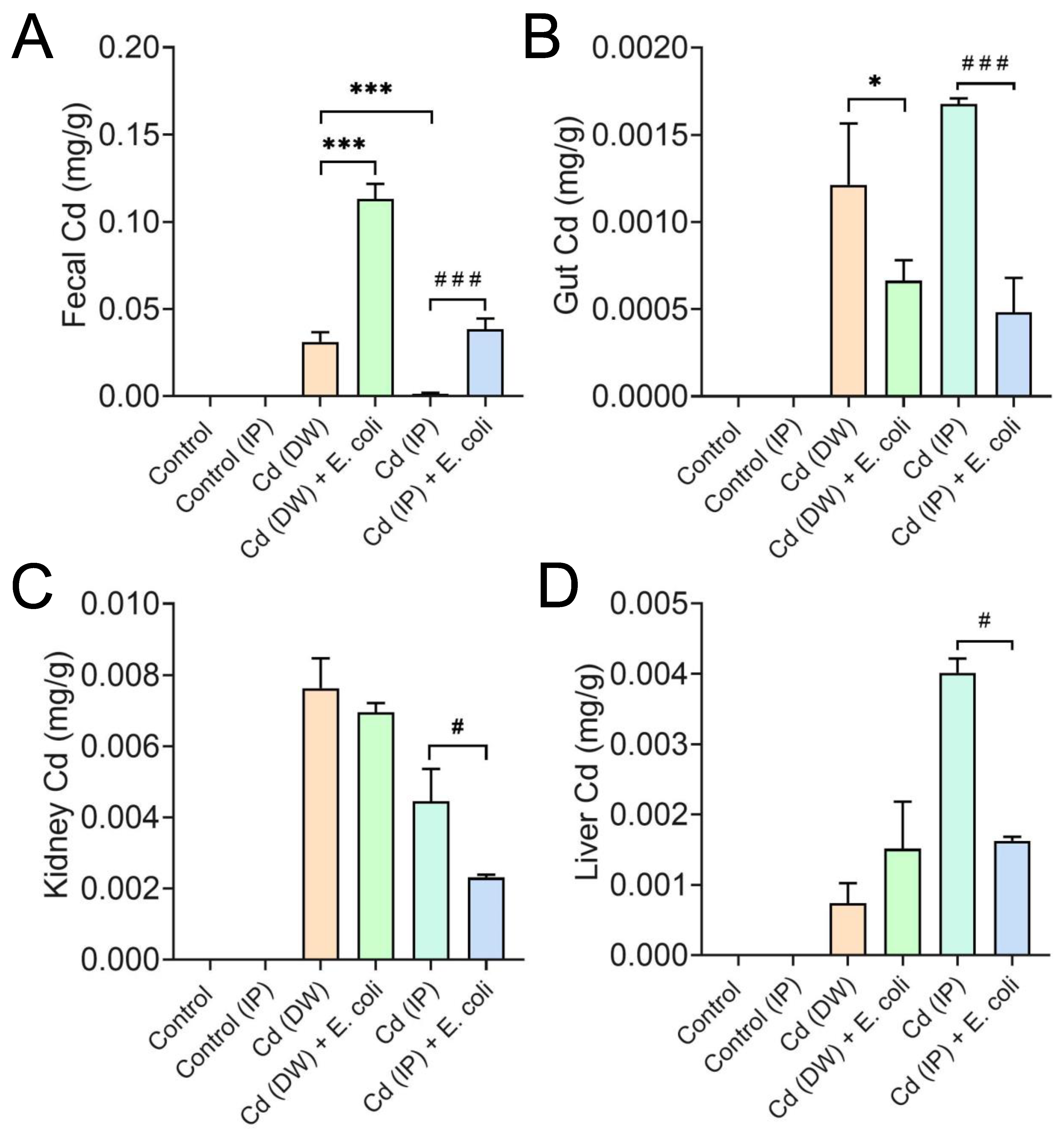
2.4. E. coli Facilitated the Excretion of Cd in Mice
3. Discussion
4. Materials and Methods
4.1. Reagents and Animals
4.2. Treatment of 20 AAs in Cd-Exposed Mice
4.3. Treatment of HLA-1-1 in Cd-Exposed Mice
4.4. Determination of Cd Contents in Feces and Tissues
4.5. Determination of Conductivity Difference
4.6. 16S rDNA Gene Sequencing of GM
4.7. Histopathological Examination
4.8. Determination of Cd-Chelating Ability
4.9. Cd-Sensitivity Assay for E. coli
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Shi, J.; Du, P.; Luo, H.; Wu, H.; Zhang, Y.; Chen, J.; Wu, M.; Xu, G.; Gao, H. Soil contamination with cadmium and potential risk around various mines in China during 2000–2020. J. Environ. Manag. 2022, 310, 114509. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Fei, J.; Cao, P.; Zhang, C.; Tang, M.; Cheng, J.; Zhao, H.; Fu, L. Serum cadmium positively correlates with inflammatory cytokines in patients with chronic obstructive pulmonary disease. Environ. Toxicol. 2021, 37, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Torres, J.; Zamudio-Cuevas, Y.; Martínez-Nava, G.A.; Aztatzi-Aguilar, O.G.; Sierra-Vargas, M.P.; Lozada-Pérez, C.A.; Suárez-Ahedo, C.; Landa-Solís, C.; Olivos-Meza, A.; Del Razo, L.M.; et al. Impact of Cadmium Mediated by Tobacco Use in Musculoskeletal Diseases. Biol. Trace Element Res. 2021, 200, 2008–2015. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, J.; Xu, C.; Da, M.; Xu, Y.; Chen, Y.; Mo, X. Increased serum levels of cadmium are associated with an elevated risk of cardiovascular disease in adults. Environ. Sci. Pollut. Res. 2021, 29, 1836–1844. [Google Scholar] [CrossRef]
- Satarug, S.; Đorđević, A.B.; Yimthiang, S.; Vesey, D.A.; Gobe, G.C. The NOAEL Equivalent of Environmental Cadmium Exposure Associated with GFR Reduction and Chronic Kidney Disease. Toxics 2022, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Plant, J.A.; Voulvoulis, N.; Oates, C.J.; Ihlenfeld, C. Cadmium levels in Europe: Implications for human health. Environ. Geochem. Health 2010, 32, 1–12. [Google Scholar] [CrossRef]
- Ngugi, M.M.; Gitari, H.I.; Muii, C.; Gweyi-Onyango, J.P. Cadmium mobility, uptake, and accumulation in spinach, kale, and amaranths vegetables as influenced by silicon fertilization. Bioremediation J. 2022, 26, 113–127. [Google Scholar] [CrossRef]
- Elhelaly, A.E.; Elbadry, S.; Eltanani, G.S.A.; Saad, M.F.; Darwish, W.S.; Tahoun, A.B.M.B.; Ellatif, S.S.A. Residual contents of the toxic metals (lead and cadmium), and the trace elements (copper and zinc) in the bovine meat and dairy products: Residues, dietary intakes, and their health risk assessment. Toxin Rev. 2022, 41, 968–975. [Google Scholar] [CrossRef]
- Cui, S.; Wang, Z.; Li, X.; Wang, H.; Wang, H.; Chen, W. A comprehensive assessment of heavy metal(loid) contamination in leafy vegetables grown in two mining areas in Yunnan, China—A focus on bioaccumulation of cadmium in Malabar spinach. Environ. Sci. Pollut. Res. 2022. [Google Scholar] [CrossRef]
- Spungen, J.H. Children’s exposures to lead and cadmium: FDA total diet study 2014–16. Food Addit. Contam. Part A 2019, 36, 893–903. [Google Scholar] [CrossRef]
- Johri, N.; Jacquillet, G.; Unwin, R. Heavy metal poisoning: The effects of cadmium on the kidney. Biometals 2010, 23, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Park, J.D.; Cherrington, N.J.; Klaassen, C.D. Intestinal Absorption of Cadmium Is Associated with Divalent Metal Transporter 1 in Rats. Toxicol. Sci. 2002, 68, 288–294. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Qi, Z.; Hou, H.; Qian, L.; Gao, J.; Zhang, X.-X. Structural and functional alterations of gut microbiome in mice induced by chronic cadmium exposure. Chemosphere 2020, 246, 125747. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Qi, Z.; Hou, H.; Gao, J.; Zhang, X.-X. Effects of chronic cadmium exposure at food limitation-relevant levels on energy metabolism in mice. J. Hazard. Mater. 2020, 388, 121791. [Google Scholar] [CrossRef]
- Zhu, G.; Cheng, D.; Wang, X.; Guo, Q.; Zhang, Q.; Zhang, J.; Tu, Q.; Li, W. Free amino acids, carbon and nitrogen isotopic compositions responses to cadmium stress in two castor (Ricinus communis L.) species. Plant Physiol. Biochem. 2022, 184, 40–46. [Google Scholar] [CrossRef]
- Su, X.; Gao, Y.; Yang, R. Gut microbiota-derived tryptophan metabolites maintain gut and systemic homeostasis. Cells 2022, 11, 2296. [Google Scholar] [CrossRef]
- Wozniak, H.; Beckmann, T.S.; Fröhlich, L.; Soccorsi, T.; Le Terrier, C.; de Watteville, A.; Schrenzel, J.; Heidegger, C.-P. The central and biodynamic role of gut microbiota in critically ill patients. Crit. Care 2022, 26, 250. [Google Scholar] [CrossRef]
- Zhang, X.; Gérard, P. Diet-gut microbiota interactions on cardiovascular disease. Comput. Struct. Biotechnol. J. 2022, 20, 1528–1540. [Google Scholar] [CrossRef]
- Schlechte, J.; Skalosky, I.; Geuking, M.B.; McDonald, B. Long-distance relationships—Regulation of systemic host defense against infections by the gut microbiota. Mucosal Immunol. 2022, 15, 809–818. [Google Scholar] [CrossRef]
- Kamioka, M.; Goto, Y.; Nakamura, K.; Yokoi, Y.; Sugimoto, R.; Ohira, S.; Kurashima, Y.; Umemoto, S.; Sato, S.; Kunisawa, J.; et al. Intestinal commensal microbiota and cytokines regulate Fut2 + Paneth cells for gut defense. Proc. Natl. Acad. Sci. USA 2022, 119, e2115230119. [Google Scholar] [CrossRef]
- Antunes, L.C.M.; McDonald, J.A.K.; Schroeter, K.; Carlucci, C.; Ferreira, R.B.R.; Wang, M.; Yurist-Doutsch, S.; Hira, G.; Jacobson, K.; Davies, J.; et al. Antivirulence Activity of the Human Gut Metabolome. MBio 2014, 5, e01183-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, H.; Yu, L.; Tian, F.; Zhai, Q.; Fan, L.; Chen, W. Gut microbiota: A target for heavy metal toxicity and a probiotic protective strategy. Sci. Total. Environ. 2020, 742, 140429. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Yang, J.; Zhao, S.; Ling, Z.; Han, R.; Wu, Y.; Salama, E.-S.; Kakade, A.; Khan, A.; Jin, W.; et al. Human supplementation with Pediococcus acidilactici GR-1 decreases heavy metals levels through modifying the gut microbiota and metabolome. NPJ Biofilms Microbiomes 2022, 8, 63. [Google Scholar] [CrossRef]
- Breton, J.Ô.; Daniel, C.; Dewulf, J.; Pothion, S.; Froux, N.; Sauty, M.; Thomas, P.; Pot, B.; Foligne, B. Gut microbiota limits heavy metals burden caused by chronic oral exposure. Toxicol. Lett. 2013, 222, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Narbad, A.; Chen, W. Oral Administration of Probiotics Inhibits Absorption of the Heavy Metal Cadmium by Protecting the Intestinal Barrier. Appl. Environ. Microbiol. 2016, 82, 4429–4440. [Google Scholar] [CrossRef] [Green Version]
- Fazeli, M.; Hassanzadeh, P.; Alaei, S. Cadmium chloride exhibits a profound toxic effect on bacterial microflora of the mice gastrointestinal tract. Hum. Exp. Toxicol. 2011, 30, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Chen, Y.; Li, Y.; Sun, L.; Deng, Q.; Wang, J.; Gooneratne, R. Oleic Acid Facilitates Cd Excretion by Increasing the Abundance of Burkholderia in Cd-Exposed Mice. Int. J. Mol. Sci. 2022, 23, 14718. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kumar, V.; Panwar, R.; Ram, C. Efficacy of indigenous probiotic Lactobacillus strains to reduce cadmium bioaccessibility—An in vitro digestion model. Environ. Sci. Pollut. Res. 2017, 24, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Narbad, A.; Chen, W. Dietary strategies for the treatment of cadmium and lead toxicity. Nutrients 2015, 7, 552–571. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Zhou, X.; Li, Y.; Li, Y.; Xia, H.; Li, Z.; Zhuang, P. Use of dietary components to reduce the bioaccessibility and bioavailability of cadmium in rice. J. Agric. Food Chem. 2020, 68, 4166–4175. [Google Scholar] [CrossRef]
- Rafieian-Naeini, H.R.; Zhandi, M.; Sadeghi, M.; Yousefi, A.R.; Marzban, H.; Benson, A.P. The effect of dietary coenzyme Q10 supplementation on egg quality and liver histopathology of layer quails under cadmium challenge. J. Anim. Physiol. Anim. Nutr. 2022. [Google Scholar] [CrossRef] [PubMed]
- Linru, H.; Zhijia, F.; Jian, G.; Jingwen, W.; Yongbin, L.; Lijun, S.; Yaling, W.; Jianmeng, L.; Ravi, G. Protective role of l -threonine against cadmium toxicity in Saccharomyces cerevisiae. J. Basic Microbiol. 2021, 61, 339–350. [Google Scholar] [CrossRef]
- Rafiq, S.; Huma, N.; Pasha, I.; Sameen, A.; Mukhtar, O.; Khan, M.I. Chemical Composition, Nitrogen Fractions and Amino Acids Profile of Milk from Different Animal Species. Asian-Australas. J. Anim. Sci. 2015, 29, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Ni, F.; Yu, W.-M.; Li, Z.; Graham, D.K.; Jin, L.; Kang, S.; Rossi, M.R.; Li, S.; Broxmeyer, H.E.; Qu, C.-K. Critical role of ASCT2-mediated amino acid metabolism in promoting leukaemia development and progression. Nat. Metab. 2019, 1, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Zemanová, V.; Pavlík, M.; Pavlíková, D.; Tlustoš, P. The significance of methionine, histidine and tryptophan in plant responses and adaptation to cadmium stress. Plant Soil Environ. 2014, 60, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Paniagua-Castro, N.; Escalona-Cardoso, G.; Cevallos, G.C. Glycine reduces cadmium-induced teratogenic damage in mice. Reprod. Toxicol. 2007, 23, 92–97. [Google Scholar] [CrossRef]
- Hwang, D.F.; Wang, L.C. Effect of taurine on toxicity of cadmium in rats. Toxicology 2001, 167, 173–180. [Google Scholar] [CrossRef]
- Bifari, F.; Ruocco, C.; Decimo, I.; Fumagalli, G.; Valerio, A.; Nisoli, E. Amino acid supplements and metabolic health: A potential interplay between intestinal microbiota and systems control. Genes Nutr. 2017, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Kaminaka, K.; Kiyozumi, M.; Honda, T. Comparative effects of three chelating agents on distribution and excretion of cadmium in rats. Toxicol. Appl. Pharmacol. 1986, 83, 516–524. [Google Scholar] [CrossRef]
- Firouzian, F.; Pourshoja, P.; Nili-Ahmadabadi, A.; Ranjbar, A. Hepatoprotective effect of N-acetylcystein loaded niosomes on liver function in paraquat-induced acute poisoning. Pestic. Biochem. Physiol. 2019, 160, 146–153. [Google Scholar] [CrossRef]
- Nabi, F.; Arain, M.A.; Bhutto, Z.A.; Shah, Q.A.; Bangulzai, N.; Ujjan, N.A.; Fazlani, S.A. Effect of early feeding of L-arginine and L-threonine on hatchability and post-hatch performance of broiler chicken. Trop. Anim. Health Prod. 2022, 54, 380. [Google Scholar] [CrossRef] [PubMed]
- Worlanyo, H.G.; Jiang, S.; Yu, Y.; Liu, B.; Zhou, Q.; Sun, C.; Miao, L.; Lin, Y.; Zheng, X.; Saidyleigh, M.; et al. Effects of dietary threonine on growth and immune response of oriental river prawn (Macrobrachium nipponense). Fish Shellfish Immunol. 2022, 128, 288–299. [Google Scholar] [CrossRef]
- Harper, A.E.; Monson, W.J.; Benton, D.A.; Elvehjem, C.A. The Influence of Protein and Certain Amino Acids, Particularly Threonine, on the Deposition of Fat in the Liver of the Rat. J. Nutr. 1953, 50, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jo, Y.; Cho, D.; Ryu, D. L-threonine promotes healthspan by expediting ferritin-dependent ferroptosis inhibition in C. elegans. Nat. Commun. 2022, 13, 6554. [Google Scholar] [CrossRef] [PubMed]
- Daabees, A.Y. Portubations of amino acid levels in plasma and liver of rat by cadmium chloride. Bull. Fac. Sci. Univ. Alex. 1987, 27, 99–113. [Google Scholar]
- Fang, Z.; Li, Y.; Wang, J.; Wang, X.; Huang, L.; Sun, L.; Deng, Q. Alleviative effect of threonine on cadmium-induced liver injury in mice. Biol. Trace Element Res. 2022. [Google Scholar] [CrossRef]
- Daisley, B.A.; Monachese, M.; Trinder, M.; Bisanz, J.E.; Chmiel, J.A.; Burton, J.P.; Reid, G. Immobilization of cadmium and lead by Lactobacillus rhamnosus GR-1 mitigates apical-to-basolateral heavy metal translocation in a Caco-2 model of the intestinal epithelium. Gut Microbes 2019, 10, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Yin, R.; Yu, L.; Wang, G.; Tian, F.; Yu, R.; Zhao, J.; Liu, X.; Chen, Y.Q.; Zhang, H.; et al. Screening of lactic acid bacteria with potential protective effects against cadmium toxicity. Food Control. 2015, 54, 23–30. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodríguez-Piñeiro, A.M.; Schütte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E.V. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Breton, J.; Massart, S.; Vandamme, P.; De Brandt, E.; Pot, B.; Foligné, B. Ecotoxicology inside the gut: Impact of heavy metals on the mouse microbiome. BMC Pharmacol. Toxicol. 2013, 14, 62. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, Y.; Xia, Y.; Liu, K.; Ren, L.; Ji, Y. The dysbiosis of gut microbiota caused by low-dose cadmium aggravate the injury of mice liver through increasing intestinal permeability. Microorganisms 2020, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.J.; Berrios, R.; Stelzhammer, S.; Hohmann, M.; Verri, W., Jr.; Bracarense, A.P.F.R.L. Ingestion of organic acids and cinnamaldehyde improves tissue homeostasis of piglets exposed to enterotoxic Escherichia coli (ETEC). J. Anim. Sci. 2020, 98, skaa012. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.-B.; Shi, J.-J.; Wang, C.-H.; Chang, J.-S. Biosorption of lead, copper and cadmium by an indigenous isolate Enterobacter sp. J1 possessing high heavy-metal resistance. J. Hazard. Mater. 2006, 134, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Pramanik, K.; Sarkar, A.; Ghosh, P.K.; Soren, T.; Maiti, T.K. Bioaccumulation of cadmium by Enterobacter sp. and enhancement of rice seedling growth under cadmium stress. Ecotoxicol. Environ. Saf. 2018, 156, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Badawy, I.H.; Hmed, A.A.; Sofy, M.R.; Al-Mokadem, A.Z. Alleviation of cadmium and nickel toxicity and phyto-stimulation of tomato plant l. by endophytic micrococcus luteus and enterobacter cloacae. Plants 2022, 11, 2018. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Narbad, A.; Chen, Y.Q.; Zhang, H.; Tian, F.; Chen, W. Protective Effects of Lactobacillus plantarum CCFM8610 against Chronic Cadmium Toxicity in Mice Indicate Routes of Protection besides Intestinal Sequestration. Appl. Environ. Microbiol. 2014, 80, 4063–4071. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Liu, Y.; Wang, C.; Zhao, J.; Zhang, H.; Tian, F.; Lee, Y.-k.; Chen, W. Increased cadmium excretion due to oral administration of lactobacillus plantarum strains by regulating enterohepatic circulation in mice. J. Agric. Food Chem. 2019, 67, 3956–3965. [Google Scholar] [CrossRef]
- Liu, T.; Liang, X.; Lei, C.; Huang, Q.; Song, W.; Fang, R.; Li, C.; Li, X.; Mo, H.; Sun, N.; et al. High-Fat Diet Affects Heavy Metal Accumulation and Toxicity to Mice Liver and Kidney Probably via Gut Microbiota. Front. Microbiol. 2020, 11, 1604. [Google Scholar] [CrossRef]
- Laddaga, R.A.; Silver, S. Cadmium uptake in Escherichia coli K-12. J. Bacteriol. 1985, 162, 1100–1105. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Deng, Y.; Deng, Q.; Sun, D.; Fang, Z.; Sun, L.; Wang, Y.; Gooneratne, R. Vibrio parahaemolyticus infection in mice reduces protective gut microbiota, Augmenting Disease Pathways. Front. Microbiol. 2020, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Shang, Q.; Song, G.; Zhang, M.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jiang, W. Application of high-throughput sequencing in understanding human oral microbiome related with health and disease. Front. Microbiol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zheng, Y.; Zhou, Y.; Guo, W.; Tang, Q.; Rong, G.; Hu, W.; Tang, J.; Luo, H. Gut Dysbiosis with Minimal Enteritis Induced by High Temperature and Humidity. Sci. Rep. 2019, 9, 18686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Yeoh, B.S.; Saha, P.; Tian, Y.; Singh, V.; Patterson, A.D.; Vijay-Kumar, M. Modulation of urinary siderophores by the diet, gut microbiota and inflammation in mice. J. Nutr. Biochem. 2016, 41, 25–33. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | n | Concentration of Cd in Drinking Water (μM) a | Oral Administration (mmol/g/d bw) b |
|---|---|---|---|
| Control | 6 | - | - |
| Cd (DW) c | 6 | 100 | - |
| Cd (DW) + AA | 6 × 20 | 100 | 40 (AA) |
| Groups | n | Concentration of Cd in Drinking Water (μM) | Intraperitoneal Injection of Cd (μg/d) | Oral Administration of E. coli (CFU/d) |
|---|---|---|---|---|
| Control (DW) | 6 | - | - | - |
| Control (IP) | 6 | - | - | - |
| Cd (DW) | 6 | 100 | - | - |
| Cd (IP) | 6 | - | 9 | - |
| Cd (DW) + E. coli | 6 | 100 | - | 1 × 109 |
| Cd (IP) + E. coli | 6 | - | 9 | 1 × 109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Fang, Z.; Zhou, X.; Gao, J.; Wang, J.; Huang, L.; Chen, Y.; Sun, L.; Deng, Q.; Gooneratne, R. Threonine Facilitates Cd Excretion by Increasing the Abundance of Gut Escherichia coli in Cd-Exposed Mice. Molecules 2023, 28, 177. https://doi.org/10.3390/molecules28010177
Li Y, Fang Z, Zhou X, Gao J, Wang J, Huang L, Chen Y, Sun L, Deng Q, Gooneratne R. Threonine Facilitates Cd Excretion by Increasing the Abundance of Gut Escherichia coli in Cd-Exposed Mice. Molecules. 2023; 28(1):177. https://doi.org/10.3390/molecules28010177
Chicago/Turabian StyleLi, Yongbin, Zhijia Fang, Xuewei Zhou, Jian Gao, Jingwen Wang, Linru Huang, Yinyan Chen, Lijun Sun, Qi Deng, and Ravi Gooneratne. 2023. "Threonine Facilitates Cd Excretion by Increasing the Abundance of Gut Escherichia coli in Cd-Exposed Mice" Molecules 28, no. 1: 177. https://doi.org/10.3390/molecules28010177