

Contribution of Insoluble Bound Antioxidants from Germinated Seeds of Wheat and Spelt to the Nutritional Value of White Bread

Abstract
:
1. Introduction
2. Results
2.1. Non-Germinated Seeds vs. Germinated Seeds
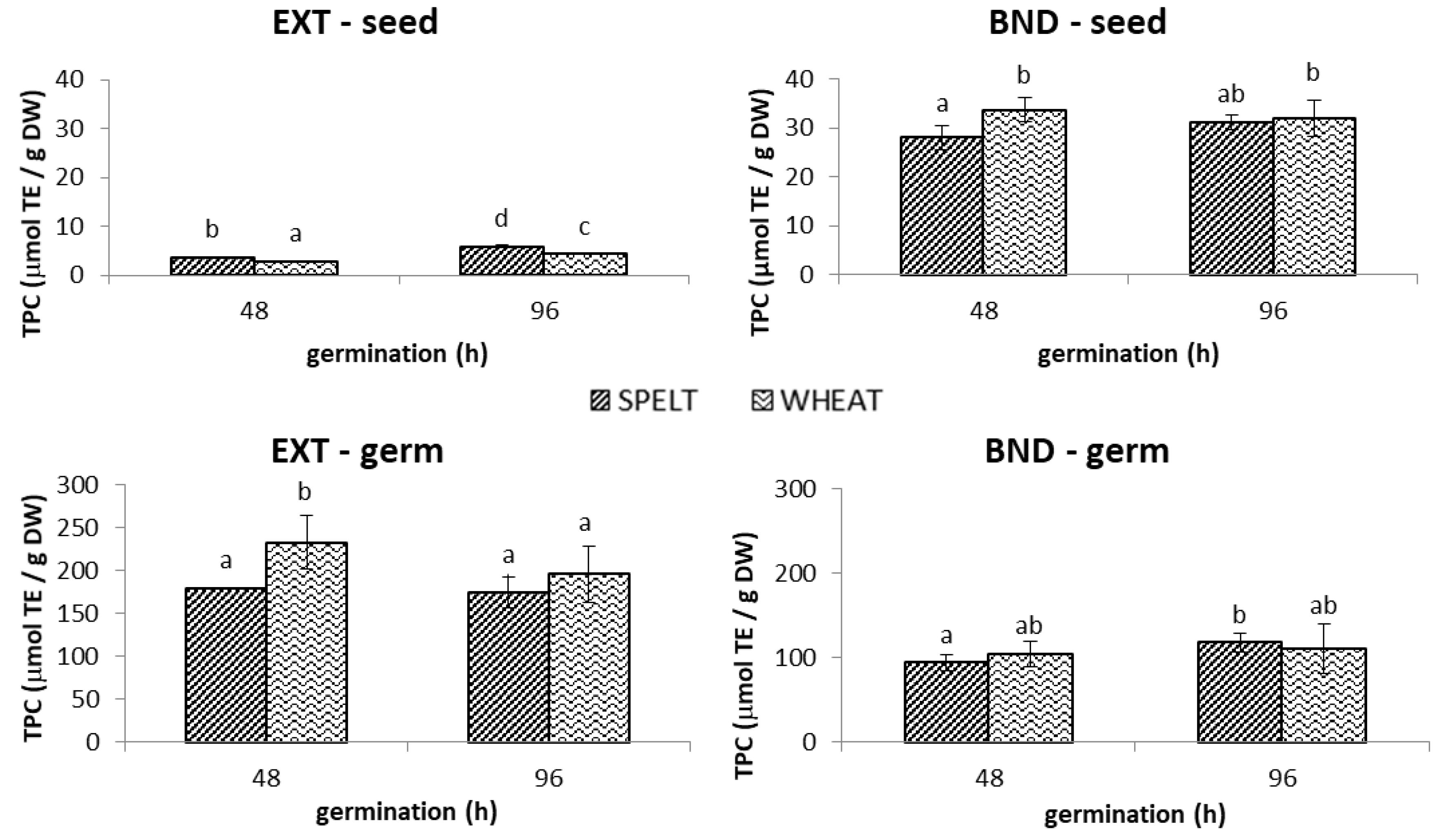
2.1.1. TPC
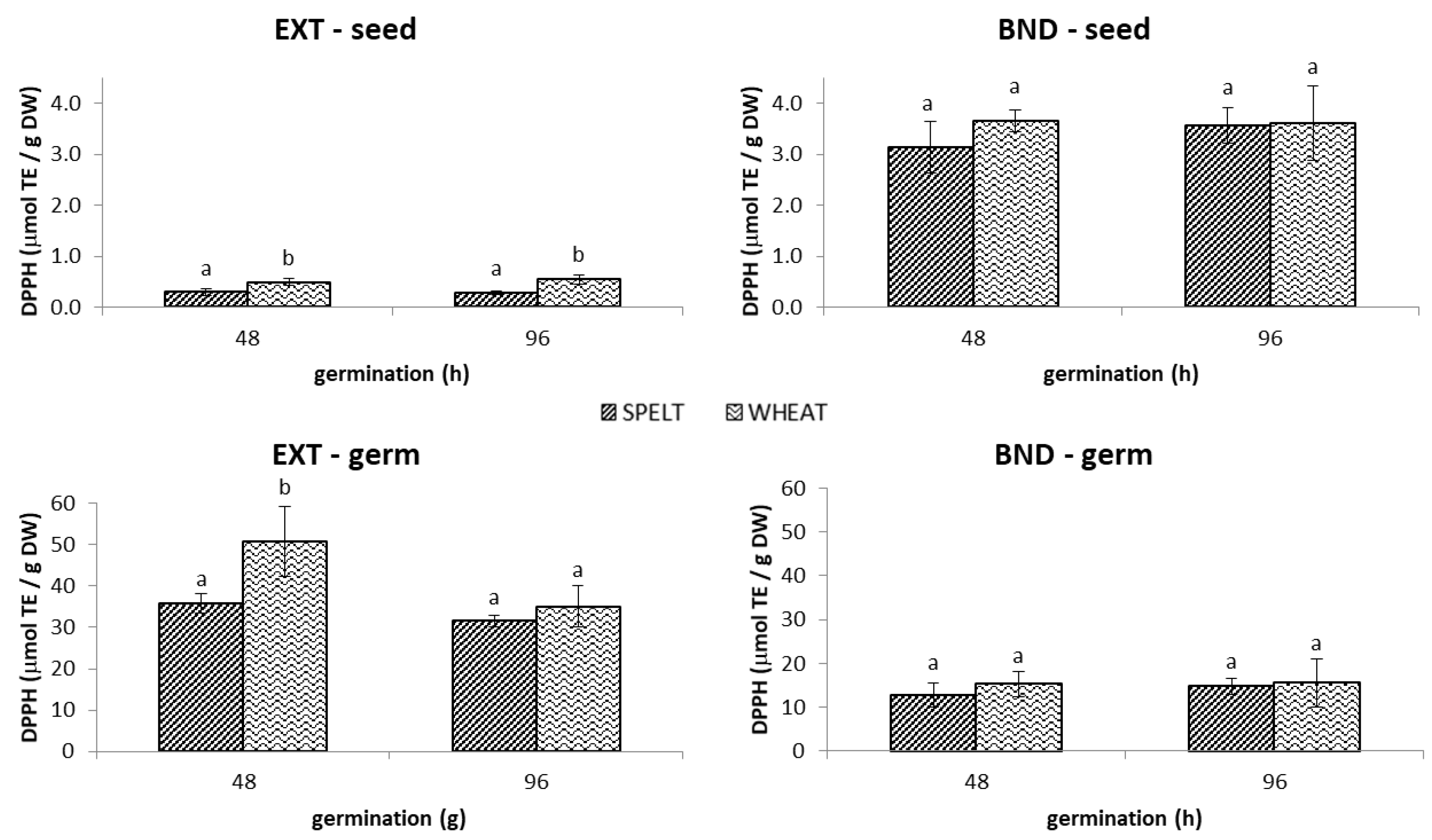
2.1.2. DPPH• Antioxidant Capacity
2.1.3. ABTS•+ Antioxidant Capacity
2.2. Content of Ferulic and p-Coumaric Acids
2.2.1. p-Coumaric Acid
2.2.2. trans-Ferulic Acid
2.2.3. cis-Ferulic Acid
2.3. Specific Volume
2.4. Color Measurement
2.5. Sensory Evaluation
2.6. Principal Component Analysis
3. Discussion
3.1. Germinated Seeds and Germs
3.2. Bread Enriched with Germinated Flour
4. Materials and Methods
4.1. Materials
4.2. Germination Performance
4.3. Spelt and Wheat Germinated Seeds Flour Preparation
4.4. Bread Making
4.5. Preparation of Crude Extracts of Extractable and Bound Phenolics
4.6. Purification of Crude Extracts
4.7. Determination of Total Phenolic Content (TPC)
4.8. Analysis of the Polyphenol Composition by LC-MS/MS
4.9. Antioxidant Activity Determined by Using DPPH Assay
4.10. Antioxidant Activity Determined by Using the ABTS Assay
4.11. Specific Volume
4.12. Color Measurement
4.13. Sensory Evaluation
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Silva, S.; Oliveira, A.L.S.; Ribeiro, T.; Madureira, A.R.; Pintado, M.M. Non-Extractable Polyphenols and Carotenoids: Importance in Human Nutrition and Health, 1st ed.; Pérez-Jiménez, J., Saura-Calixto, F., Eds.; Royal Society of Chemistry: London, UK, 2018; Chapter 17; ISBN 978-1-78801-106-8. [Google Scholar]
- Sevilla, M.T.E.; Martínez, C.J.; Cardador-Martínez, A. Phenolic compounds in cereals and legumes. In Phenolic Compounds in Food: Characterization and Analysis, 1st ed.; Nollet, L.M.L., Gutierrez-Uribe, J.A., Eds.; CRC Press: New York, NY, USA, 2018; pp. 319–332. ISBN 978-1-4987-2296-4. [Google Scholar]
- Wieser, H.; Koehler, P.; Scherf, K.A. Wheat—An Exceptional Crop: Botanical Features, Chemistry, Utilization, Nutritional and Health Aspects, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 61–102, 133–148. ISBN 978-0-12-821715-3. [Google Scholar]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient Wheat Species and Human Health: Biochemical and Clinical Implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef]
- Camire, M.E. Overview of grain components and changes occurring in grain constituents with different forms of processing. In Cereal Grain-Based Functional Foods Carbohydrate and Phytochemical Components, 1st ed.; Beta, T., Camire, M.E., Eds.; The Royal Society of Chemistry: London, UK, 2019; pp. 12–31. ISBN 978-1-78801-148-8. [Google Scholar]
- Mencin, M.; Abramovič, H.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Abiotic Stress Combinations Improve the Phenolics Profiles and Activities of Extractable and Bound Antioxidants from Germinated Spelt (Triticum spelta L.) Seeds. Food Chem. 2021, 344, 128704. [Google Scholar] [CrossRef]
- Mencin, M.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Enzymatic treatments of raw, germinated and fermented spelt (Triticum spelta L.) seeds improve the accessibility and antioxidant activity of their phenolics. LWT-Food Sci. Technol. 2022, 169, 114046. [Google Scholar] [CrossRef]
- Mencin, M.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Improving Accessibility and Bioactivity of Raw, Germinated and Enzymatic-Treated Spelt (Triticum Spelta L.) Seed Antioxidants by Fermentation. Food Chem. 2022, 394, 133483. [Google Scholar] [CrossRef]
- Mencin, M.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Simulated Gastrointestinal Digestion of Bioprocessed Spelt Seeds: Bioaccessibility and Bioactivity of Phenolics. Antioxidants 2022, 11, 1703. [Google Scholar] [CrossRef] [PubMed]
- Mencin, M.; Markanovič, N.; Mikulič-Petkovšek, M.; Veberič, R.; Terpinc, P. Changes in the Bioaccessibility of Antioxidants after Simulated In Vitro Digestion of Bioprocessed Spelt-Enhanced Wheat Bread. Antioxidants 2023, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Maqueo, A.; Escobedo-Avellaneda, Z.; Cano, M.P.; Welti-Chanes, J. Phenolic compounds in food. In Phenolic Compounds in Food: Characterization and Analysis, 1st ed.; Nollet, L.M.L., Gutierrez-Uribe, J.A., Eds.; CRC Press: New York, NY, USA, 2018; pp. 33–60. ISBN 978-1-4987-2296-4. [Google Scholar]
- Beta, T. Introduction. In Cereal Grain-Based Functional Foods Carbohydrate and Phytochemical Components, 1st ed.; Beta, T., Camire, M.E., Eds.; The Royal Society of Chemistry: London, UK, 2019; pp. 1–11. ISBN 978-1-78801-148-8. [Google Scholar]
- Aziz, A.; Noreen, S.; Khalid, W.; Mubarik, F.; Niazi, M.K.; Koraqi, H.; Ali, A.; Lima, C.M.G.; Alansari, W.S.; Eskandrani, A.A.; et al. Extraction of Bioactive Compounds from Different Vegetable Sprouts and Their Potential Role in the Formulation of Functional Foods against Various Disorders: A Literature-Based Review. Molecules 2022, 27, 7320. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, P.; Weng, Y.; Ma, Y.; Gu, Z.; Yang, R. Comparison of phenolic profiles, antioxidant capacity and relevant enzyme activity of different Chinese wheat varieties during germination. Food Biosci. 2017, 20, 159–167. [Google Scholar] [CrossRef]
- Xu, M.; Rao, J.; Chen, B. Phenolic compounds in germinated cereal and pulse seeds: Classification, transformation, and metabolic process. Crit. Rev. Food Sci. Nutr. 2020, 60, 740–759. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food Agric. 2014, 95, 1795–1803. [Google Scholar] [CrossRef]
- Domínguez-Avila, J.A.; Ruiz-Canizales, J.; Pacheco-Ordaz, R.; Villegas-Ochoa, M.A.; González Aguilar, G.A. Antioxidant power. In Phenolic Compounds in Food: Characterization and Analysis, 1st ed.; Nollet, L.M.L., Gutierrez-Uribe, J.A., Eds.; CRC Press: New York, NY, USA, 2018; pp. 261–290. ISBN 978-1-4987-2296-4. [Google Scholar]
- Abdel-Aal, E.-S.M.; Rabalski, I. Bioactive Compounds and their Antioxidant Capacity in Selected Primitive and Modern Wheat Species. Open Agric. J. 2008, 2, 7–14. [Google Scholar] [CrossRef]
- Abramovič, H.; Grobin, B.; Poklar Ulrih, N.; Cigić, B. Relevance and Standardization of In Vitro Antioxidant Assays: ABTS, DPPH, and Folin–Ciocalteu. J. Chem. 2018, 2018, 4608405. [Google Scholar] [CrossRef]
- Ndolo, V.; Trust, B. Types and distribution of phenolic compounds in grains. In Cereal Grain-Based Functional Foods Carbohydrate and Phytochemical Components, 1st ed.; Beta, T., Camire, M.E., Eds.; The Royal Society of Chemistry: London, UK, 2019; pp. 235–277. ISBN 978-1-78801-148-8. [Google Scholar]
- Gawlik-Dziki, U.; Swieca, M.; Dziki, D. Comparison of Phenolic Acids Profile and Antioxidant Potential of Six Varieties of Spelt (Triticum spelta L.). J. Agric. Food Chem. 2012, 60, 4603–4612. [Google Scholar] [CrossRef] [PubMed]
- Atudorei, D.; Codină, G.G. Perspectives on the Use of Germinated Legumes in the Bread Making Process, a Review. Appl. Sci. 2020, 10, 6244. [Google Scholar] [CrossRef]
- Hallén, E.; İbanoğlu, Ş.; Ainsworth, P. Effect of Fermented/Germinated Cowpea Flour Addition on the Rheological and Baking Properties of Wheat Flour. J. Food Eng. 2004, 63, 177–184. [Google Scholar] [CrossRef]
- Andersen, G.; Koehler, P.; Somoza, V. Postprandial Glucose and Free Fatty Acid Response is Improved by Wheat Bread Fortified with Germinated Wheat Seedlings. Curr. Top. Nutraceutical Res. 2008, 6, 15–21. [Google Scholar]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.-A.; Van den Broeck, H.C.; Brouns, F.J.P.H.; De Brier, N.; et al. Impact of Cereal Seed Sprouting on Its Nutritional and Technological Properties: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2018, 18, 305–328. [Google Scholar] [CrossRef] [PubMed]
- Marti, A.; Cardone, G.; Nicolodi, A.; Quaglia, L.; Pagani, M.A. Sprouted Wheat as an Alternative to Conventional Flour Improvers in Bread-making. LWT-Food Sci. Technol. 2017, 80, 230–236. [Google Scholar] [CrossRef]
- de Ruiz, A.S.C.; Bressani, R. Effect of Germination on the Chemical Composition and Nutrition Value of Amaranth Grain. Cereal Chem. 1990, 67, 519–522. [Google Scholar]
- Dziki, D.; Różyło, R.; Gawlik-Dziki, U.; Świeca, M. Current Trends in the Enhancement of Antioxidant Activity of Wheat Bread by the Addition of Plant Materials Rich in Phenolic Compounds. Trends Food Sci. Technol. 2014, 40, 48–61. [Google Scholar] [CrossRef]
- Millar, K.A.; Barry-Ryan, C.; Burke, R.; McCarthy, S.; Gallagher, E. Dough Properties and Baking Characteristics of White Bread, as Affected by Addition of Raw, Germinated and Toasted Pea Flour. Innov. Food Sci. Emerg. Technol. 2019, 56, 102189. [Google Scholar] [CrossRef]
- Levent, H.; Bilgiçli, N.; Ertaş, N. The Assessment of Leavened and Unleavened Flat Breads Properties Enriched with Wheat Germ. Qual. Assur. Saf. Crops Foods. 2014, 7, 321–326. [Google Scholar] [CrossRef]
- Diowksz, A.; Kordialik-Bogacka, E.; Ambroziak, W. Se-enriched Sprouted Seeds as Functional Additives in Sourdough Fermentation. LWT-Food Sci. Technol. 2014, 56, 524–528. [Google Scholar] [CrossRef]
- Gutfinger, T. Polyphenols in Olive Oils. J. Am. Oil Chem. Soc. 1981, 58, 966–968. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- AACC Approved Methods of Analysis 11th Ed. Method 10-05.01. Guidelines for Measurement of Volume by Rapeseed Displacement. Available online: http://methods.aaccnet.org/summaries/10-05-01.aspx (accessed on 25 August 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination | TPC | DPPH | ABTS | ||||
|---|---|---|---|---|---|---|---|
| Time | Seeds | Extractable | Bound | Extractable | Bound | Extractable | Bound |
| 0 h | wheat | 1.84 a | 31.67 b | 0.46 a | 3.01 b | 1.83 a | 29.71 b |
| spelt | 2.47 a | 27.07 a | 0.34 a | 2.59 a | 3.39 b | 26.53 a | |
| 48 h | wheat | 10.42 b | 36.01 c | 2.15 c | 4.05 d | 10.19 c | 42.64 d |
| spelt | 10.75 b | 30.71 b | 1.75 b | 3.54 c | 10.69 c | 36.45 c | |
| 96 h | wheat | 26.03 c | 40.79 d | 4.45 d | 4.96 e | 22.39 d | 49.51 e |
| spelt | 30.64 d | 43.93 d | 4.87 d | 4.83 e | 26.91 e | 52.72 e | |
| p-Coumaric Acid | trans-Ferulic Acid | cis-Ferulic Acid | ||||
|---|---|---|---|---|---|---|
| Samples | Extractable | Bound | Extractable | Bound | Extractable | Bound |
| wheat non-germinated | 1.43 ± 0.17 a | 11.47 ± 0.11 A | 11.29 ± 0.56 a | 345.67 ± 15.24 A | 11.09 ± 0.44 a | 155.78 ± 1.97 A |
| spelt non-germinated | 1.37 ± 0.02 a | 12.77 ± 0.83 B | 10.83 ± 0.15 a | 349.60 ± 59.08 A | 10.92 ± 0.22 a | 122.01 ± 32.31 A |
| Seed | ||||||
| wheat 48 h | 2.15 ± 0.10 a | 16.51 ± 1.92 A | 16.38 ± 0.84 a | 430.75 ± 98.45 A | 15.62 ± 0.20 a | 136.53 ± 20.36 A |
| spelt 48 h | 2.21 ± 0.07 a | 15.93 ± 1.30 A | 15.73 ± 0.47 a | 387.59 ± 32.43 A | 15.29 ± 0.24 a | 156.44 ± 36.01 AB |
| wheat 96 h | 2.15 ± 0.13 a | 17.23 ± 1.47 A | 16.60 ± 0.21 a | 469.29 ± 55.95 AB | 16.25 ± 0.75 b | 155.73 ± 21.38 AB |
| spelt 96 h | 2.18 ± 0.06 a | 20.64 ± 1.40 B | 16.43 ± 0.36 a | 549.74 ± 43.79 B | 16.30 ± 0.52 b | 186.90 ± 31.50 B |
| Germ | ||||||
| wheat 48 h | 53.24 ± 6.08 a | 177.79 ± 14.66 A | 391.14 ± 30.25 b | 2352.27 ± 171.06 A | 392.33 ± 32.60 b | 1693.67 ± 47.02 B |
| spelt 48 h | 48.58 ± 3.11 a | 187.28 ± 8.96 A | 319.57 ± 25.35 a | 2342.07 ± 388.21 A | 344.36 ± 22.35 a | 1317.26 ± 193.42 A |
| wheat 96 h | 54.52 ± 5.52 a | 488.45 ± 56.44 C | 334.07 ± 47.35 ab | 2658.34 ± 231.34 A | 307.81 ± 4.94 a | 1323.36 ± 167.00 A |
| spelt 96 h | 58.35 ± 9.40 a | 384.70 ± 56.79 B | 369.40 ± 23.29 b | 2624.71 ± 161.35 A | 361.43 ± 20.81 b | 1829.02 ± 143.22 B |
| p-Coumaric Acid | trans-Ferulic Acid | cis-Ferulic Acid | ||||
|---|---|---|---|---|---|---|
| Bread | Extractable | Bound | Extractable | Bound | Extractable | Bound |
| S0 | 10.48 ± 0.74 a | 15.53 ± 1.2 a | 0.66 ± 0.18 a | 58.17 ± 3.06 a | 1.17 ± 0.71 a | 59.50 ± 12.63 ab |
| S5 | 19.89 ± 0.16 bc | 20.40 ± 3.95 a | 4.83 ± 0.69 c | 95.28 ± 10.30 a | 3.97 ± 0.78 c | 55.69 ± 0.14 ab |
| S15 | 10.57 ± 0.67 a | 53.41 ± 6.08 b | 4.05 ± 0.33 c | 213.63 ± 42.41 b | 2.50 ± 0.64 abc | 111.83 ± 12.73 c |
| S30 | 10.55 ± 0.99 a | 79.36 ± 2.56 c | 2.98 ± 0.28 b | 428.16 ± 16.8 c | 3.44 ± 0.68 abc | 154.49 ± 1.03 d |
| W0 | 16.47 ± 0.03 b | 15.90 ± 0.03 a | 0.65 ± 0.14 a | 42.85 ± 8.41 a | 1.48 ± 0.82 ab | 33.63 ± 7.36 a |
| W5 | 21.1 ± 3.31 bc | 21.70 ± 3.96 a | 2.16 ± 0.41 b | 65.23 ± 9.41 a | 3.72 ± 0.67 bc | 87.91 ± 18.91 bc |
| W15 | 22.00 ± 2.24 c | 49.32 ± 1.73 b | 3.05 ± 0.43 b | 197.88 ± 27.66 b | 4.15 ± 0.36 c | 183.45 ± 23.57 d |
| W30 | 17.38 ± 0.63 bc | 70.10 ± 12.64 c | 2.73 ± 0.21 b | 212.98 ± 44.43 b | 2.97 ± 0.27 abc | 180.84 ± 17.84 d |
| Bread | Specific Volume (mL/g) | Luminosity (L*) | Browning (100-L*) | Redness (a*) | Yellowness (b*) | Total Color Change (ΔE) |
|---|---|---|---|---|---|---|
| S0 | 2.56 ± 0.07 c | 67.67 ± 3.83 e | 32.33 ± 3.83 b | 9.03 ± 2.24 b | 36.71 ± 1.67 d | / |
| S5 | 2.85 ± 0.10 d | 59.72 ± 2.64 d | 40.28 ± 2.64 c | 11.88 ± 1.47 d | 32.78 ± 1.79 c | 9.30 |
| S15 | 2.63 ± 0.04 c | 56.52 ± 3.22 c | 43.48 ± 3.22 d | 8.61 ± 0.54 b | 26.91 ± 1.50 b | 14.85 |
| S30 | 1.75 ± 0.04 a | 43.91 ± 1.79 a | 56.09 ± 1.79 f | 11.41 ± 2.14 cd | 21.07 ± 4.36 a | 28.54 |
| W0 | 3.10 ± 0.02 e | 69.71 ± 1.91 f | 30.29 ± 1.91 a | 6.64 ± 0.81 a | 26.89 ± 1.14 b | / |
| W5 | 3.83 ± 0.13 f | 59.79 ± 3.14 d | 40.21 ± 3.14 c | 8.41 ± 0.52 b | 28.19 ± 1.17 b | 10.16 |
| W15 | 3.15 ± 0.07 e | 55.64 ± 2.30 c | 44.36 ± 2.30 d | 8.22 ± 0.69 b | 26.33 ± 1.27 b | 14.17 |
| W30 | 2.20 ± 0.02 b | 46.06 ± 2.69 b | 53.94 ± 2.69 e | 10.34 ± 1.90 c | 23.17 ± 1.63 a | 24.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mencin, M.; Golob, K.; Krek, M.; Polak, T.; Požrl, T.; Terpinc, P. Contribution of Insoluble Bound Antioxidants from Germinated Seeds of Wheat and Spelt to the Nutritional Value of White Bread. Molecules 2023, 28, 6311. https://doi.org/10.3390/molecules28176311
Mencin M, Golob K, Krek M, Polak T, Požrl T, Terpinc P. Contribution of Insoluble Bound Antioxidants from Germinated Seeds of Wheat and Spelt to the Nutritional Value of White Bread. Molecules. 2023; 28(17):6311. https://doi.org/10.3390/molecules28176311
Chicago/Turabian StyleMencin, Marjeta, Katja Golob, Maja Krek, Tomaž Polak, Tomaž Požrl, and Petra Terpinc. 2023. "Contribution of Insoluble Bound Antioxidants from Germinated Seeds of Wheat and Spelt to the Nutritional Value of White Bread" Molecules 28, no. 17: 6311. https://doi.org/10.3390/molecules28176311