

Bioactivities and Mechanisms of Action of Diphyllin and Its Derivatives: A Comprehensive Systematic Review
 ,
,
Abstract
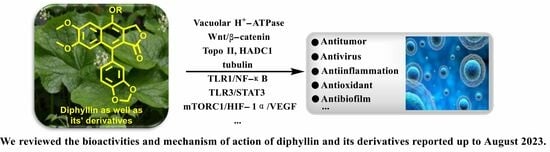
:
1. Introduction
2. Anti-Tumor Activity
3. Anti-Virus Activity
4. Other Biological Activities
5. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Mandal, A.K.; Kumar, M.; Dwivedi, K.; Singh, D. Prospective Challenges for Patenting and Clinical Trials of Anticancer Compounds from Natural Products: Coherent Review. Recent Pat. Anticancer Drug Discov. 2023, 18, 470–494. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Du, R.; Liu, M.; Rong, L. Lignans and Their Derivatives from Plants as Antivirals. Molecules 2020, 25, 183. [Google Scholar] [CrossRef] [PubMed]
- Štefánik, M.; Bhosale, D.S.; Haviernik, J.; Straková, P.; Fojtíková, M.; Dufková, L.; Huvarová, I.; Salát, J.; Bartáček, J.; Svoboda, J.; et al. Diphyllin Shows a Broad-Spectrum Antiviral Activity against Multiple Medically Important Enveloped RNA and DNA Viruses. Viruses 2022, 14, 354. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Mubarak, F.A.F.; Shehadi, I.A.; Mosallam, A.M.; Temairk, H.; Badr, M.; Abdelmonsef, A.H. Design and biological evaluation of 3-substituted quinazoline-2,4(1H,3H)-dione derivatives as dual c-Met/VEGFR-2-TK inhibitors. J. Enzyme Inhib. Med. Chem. 2023, 38, 2189578. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Xu, X.-L.; Lan, J.-X.; Huang, H.; Dai, W.; Peng, X.-P.; Liu, S.-L.; Chen, W.-M.; Huang, L.-J.; Liu, J.; Li, X.-J.; et al. Synthesis, biological activity and mechanism of action of novel allosecurinine derivatives as potential antitumor agents. Bioorgan. Med. Chem. 2023, 82, 117234. [Google Scholar] [CrossRef]
- Day, S.-H.; Chiu, N.-Y.; Won, S.-J.; Lin, C.-N. Cytotoxic Lignans of Justicia ciliata. J. Nat. Prod. 1999, 62, 1056–1058. [Google Scholar] [CrossRef]
- Innocenti, G.; Puricelli, L.; Piacente, S.; Caniato, R.; Filippini, R.; Cappelletti, E.M. Patavine, a new arylnaphthalene lignan glycoside from shoot cultures of Haplophyllum patavinum. Chem. Pharm. Bull. 2002, 50, 844–846. [Google Scholar] [CrossRef]
- Day, S.-H.; Lin, Y.-C.; Tsai, M.-L.; Tsao, L.-T.; Ko, H.-H.; Chung, M.-I.; Lee, J.-C.; Wang, J.-P.; Won, S.-J.; Lin, C.-N. Potent Cytotoxic Lignans from Justicia procumbens and Their Effects on Nitric Oxide and Tumor Necrosis Factor-α Production in Mouse Macrophages. J. Nat. Prod. 2002, 65, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, C.; Ravindranath, N.; Ram, T.S.; Das, B. Arylnaphthalide Lignans from Cleistanthus collinus. Chem. Pharm. Bull. 2003, 51, 1299–1300. [Google Scholar] [CrossRef]
- Susplugas, S.; Hung, N.V.; Bignon, J.; Thoison, O.; Kruczynski, A.; Sévenet, T.; Guéritte, F. Cytotoxic Arylnaphthalene Lignans from a Vietnamese Acanthaceae, Justicia patentiflora. J. Nat. Prod. 2005, 68, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.W.; Zhou, J.R.; Hon, P.M.; Li, S.L.; Zhou, Y.; Li, L.L.; Ye, W.C.; Xu, H.X.; Shaw, P.C.; But, P.P. Lignans from Dysosma versipellis with inhibitory effects on prostate cancer cell lines. J. Nat. Prod. 2007, 70, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hui, J.; Wang, D.; Zhu, L.; Fang, J.H.; Zhao, X.D. Synthesis, Cytotoxicity and Pro-apoptosis of Novel Benzoisoindolin Hydrazones as Anticancer Agents. Chem. Pharm. Bull. 2010, 58, 1324–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ni, C.; Zhang, Y.; Zhu, L. Synthesis and Bioevaluation of Diphyllin Glycosides as Novel Anticancer Agents. Arch. Pharm. 2012, 345, 622–628. [Google Scholar] [CrossRef]
- Zhao, Y.; Ni, C.; Zhang, Y.; Zhu, L. Synthesis and Antiproliferatory Activity of Novel Diphyllin Hydroxamic Acid and Mercaptan Derivatives. Chin. J. Org. Chem. 2013, 33, 169–173. [Google Scholar] [CrossRef]
- Zhao, Y.; Hui, J.; Zhu, L. Synthesis and bioevaluation of novel arylnaphthalene lignans as anticancer agents. Med. Chem. Res. 2013, 22, 2505–2510. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, Y.; Ma, J.; Zhu, L. Synthesis and Evaluation of Cleistanthin A Derivatives as Potent Vacuolar H+-ATPase Inhibitors. Chem. Biol. Drug Des. 2015, 86, 691–696. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, R.; Lu, Y.; Ma, J.; Zhu, L. Synthesis and bioevaluation of heterocyclic derivatives of Cleistanthin-A. Bioorgan. Med. Chem. 2015, 23, 4884–4890. [Google Scholar] [CrossRef]
- Baskol, G.; Mazicioglu, M.M.; Özel, M. Therapeutic approaches on the interaction between SARS-CoV2 and ACE2: A biochemical perspective. Turk. J. Biochem. 2020, 45, 643–650. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, R.; Liu, S.; Zhao, Y.; Gao, J.; Zhu, L. ZT-25, a new vacuolar H+-ATPase inhibitor, induces apoptosis and protective autophagy through ROS generation in HepG2 cells. Eur. J. Pharmacol. 2016, 771, 130–138. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, C.; Zhang, W.; Zhao, Y. Synthesis and biological evaluation of novel lignan glycosides as anticancer agents. Chem. Biol. Drug Des. 2016, 88, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Zhu, L.; Shen, W.; Zhao, Y. Synthesis, cytotoxicity and anti-invasion activity of three natural diphyllin L-arabinopyranosides. Fitoterapia 2019, 137, 104198. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lu, Y.; Li, Y.; Ling, Y.; Zhao, Y. Synthesis and evaluation of diphyllin β-hydroxyl amino derivatives as novel V-ATPase inhibitors. Chem. Biol. Drug Des. 2021, 98, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Li, Y.; Zhu, L.; Wei, C.; Bao, X.; Zhao, Y. 2, 4, 5-Trideoxyhexopyranosides derivatives of diphyllin: Synthesis and anticancer activity. Chem. Biol. Drug Des. 2022, 100, 256–266. [Google Scholar] [CrossRef]
- Shen, W.; Zou, X.; Chen, M.; Liu, P.; Shen, Y.; Huang, S.; Guo, H.; Zhang, L. Effects of diphyllin as a novel V-ATPase inhibitor on gastric adenocarcinoma. Eur. J. Pharmacol. 2011, 667, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.-K.; Zhang, W.; Ding, N.; Li, M.; Li, Y.-X. Design, synthesis and biological evaluation of novel glycosylated diphyllin derivatives as topoisomerase II inhibitors. Eur. J. Med. Chem. 2012, 47, 424–431. [Google Scholar] [CrossRef]
- Pant, A.B.; Luo, J.; Hu, Y.; Kong, W.; Yang, M. Evaluation and Structure-Activity Relationship Analysis of a New Series of Arylnaphthalene lignans as Potential Anti-Tumor Agents. PLoS ONE 2014, 9, e93516. [Google Scholar]
- Ren, Y.; Lantvit, D.D.; Deng, Y.; Kanagasabai, R.; Gallucci, J.C.; Ninh, T.N.; Chai, H.-B.; Soejarto, D.D.; Fuchs, J.R.; Yalowich, J.C.; et al. Potent Cytotoxic Arylnaphthalene Lignan Lactones from Phyllanthus poilanei. J. Nat. Prod. 2014, 77, 1494–1504. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, J.; Zhu, L.; Zhao, Y. Synthesis and identification of cytotoxic diphyllin glycosides as vacuolar H+-ATPase inhibitors. Eur. J. Med. Chem. 2014, 82, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Lv, J.-J.; Zhao, J.-Q.; Li, Y.; Wang, D.; Yang, C.-R.; Xu, M.; Zhang, Y.-J. New cytotoxic lignan glycosides from Phyllanthus glaucus. Nat. Prod. Res. 2015, 30, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Al-Qathama, A.; Gibbons, S.; Prieto, J.M. Differential modulation of Bax/Bcl-2 ratio and onset of caspase-3/7 activation induced by derivatives of Justicidin B in human melanoma cells A375. Oncotarget 2017, 8, 95999–96012. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Cai, H.; Gu, L.; Cao, S. Cleistanthin A inhibits the invasion and metastasis of human melanoma cells by inhibiting the expression of matrix metallopeptidase-2 and -9. Oncol. Lett. 2017, 14, 6217–6223. [Google Scholar] [CrossRef]
- Chen, H.; Liu, P.; Zhang, T.; Gao, Y.; Zhang, Y.; Shen, X.; Li, X.; Shen, W. Effects of diphyllin as a novel V-ATPase inhibitor on TE-1 and ECA-109 cells. Oncol. Lett. 2018, 39, 921–928. [Google Scholar] [CrossRef]
- Shen, W.; Zhao, Y.; Chen, H.; Zhang, T.; Wu, S.; Liu, P. M3, a natural lignan xyloside, exhibits potent anticancer activity in HCT116 cells. Oncol. Lett. 2018, 17, 2117–2122. [Google Scholar] [CrossRef]
- Woodard, J.L.; Huntsman, A.C.; Patel, P.A.; Chai, H.-B.; Kanagasabai, R.; Karmahapatra, S.; Young, A.N.; Ren, Y.; Cole, M.S.; Herrera, D.; et al. Synthesis and antiproliferative activity of derivatives of the phyllanthusmin class of arylnaphthalene lignan lactones. Bioorgan. Med. Chem. 2018, 26, 2354–2364. [Google Scholar] [CrossRef]
- Yi, L.; Chen, L.; Guo, X.; Lu, T.; Wang, H.; Ji, X.; Zhang, J.; Ren, Y.; Pan, P.; Kinghorn, A.D.; et al. A Synthetic Disaccharide Derivative of Diphyllin, TAARD, Activates Human Natural Killer Cells to Secrete Interferon-Gamma via Toll-Like Receptor-Mediated NF-κB and STAT3 Signaling Pathways. Front. Immunol. 2018, 9, 1509. [Google Scholar] [CrossRef]
- Liu, S.; Wang, L.; Ding, W.; Wang, D.; Wang, X.; Luo, Q.; Lu, Y.; Zhu, L. Cleistanthin A inhibits the invasion of MDA-MB-231 human breast cancer cells: Involvement of the β-catenin pathway. Pharmacol. Rep. 2019, 72, 188–198. [Google Scholar] [CrossRef]
- Paha, J.; Kanjanasirirat, P.; Munyoo, B.; Tuchinda, P.; Suvannang, N.; Nantasenamat, C.; Boonyarattanakalin, K.; Kittakoop, P.; Srikor, S.; Kongklad, G.; et al. A novel potent autophagy inhibitor ECDD-S27 targets vacuolar ATPase and inhibits cancer cell survival. Sci. Rep. 2019, 9, 9177. [Google Scholar] [CrossRef]
- Feng, H.; Chen, G.; Zhang, Y.; Guo, M. Exploring Multifunctional Bioactive Components from Podophyllum sinense Using Multi-Target Ultrafiltration. Front. Pharmacol. 2021, 12, 749189. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Q.; Xiao, R.; Ma, J.; Tang, Y.; Chen, W.; Zhang, R.; Jiang, L.; Chen, H.; Shen, B.; et al. Synthesis and anti-tumor activity of nitrogen-containing derivatives of the natural product diphyllin. Eur. J. Med. Chem. 2022, 243, 114708. [Google Scholar] [CrossRef] [PubMed]
- Young, A.N.; Herrera, D.; Huntsman, A.C.; Korkmaz, M.A.; Lantvit, D.D.; Mazumder, S.; Kolli, S.; Coss, C.C.; King, S.; Wang, H.; et al. Phyllanthusmin Derivatives Induce Apoptosis and Reduce Tumor Burden in High-Grade Serous Ovarian Cancer by Late-Stage Autophagy Inhibition. Mol. Cancer Ther. 2018, 17, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Salvi, A.; Young, A.N.; Huntsman, A.C.; Pergande, M.R.; Korkmaz, M.A.; Rathnayake, R.A.; Mize, B.K.; Kinghorn, A.D.; Zhang, X.; Ratia, K.; et al. PHY34 inhibits autophagy through V-ATPase V0A2 subunit inhibition and CAS/CSE1L nuclear cargo trafficking in high grade serous ovarian cancer. Cell Death Dis. 2022, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, H.; Wu, M.; Zhang, T.; Zhu, L.; Zhao, Y. Synthesis, Cytotoxicity, Anti-Migration and Anti-Invasion Activity of Diphyllin Heterocyclic Derivatives. Med. Chem. 2022, 18, 122–129. [Google Scholar] [CrossRef]
- Puli, S.; Raveendran, R. Anti-proliferative and Anti-migratory Activities of Diphyllin on Human Colorectal Cancer Cells. J. Young Pharm. 2022, 14, 198–202. [Google Scholar] [CrossRef]
- Luo, J.; Kong, W.; Yang, M. HJC, a New Arylnaphthalene Lignan Isolated from Justicia procumbens, Causes Apoptosis and Caspase Activation in K562 Leukemia Cells. J. Pharmacol. Sci. 2014, 125, 355–363. [Google Scholar] [CrossRef]
- Rosa, V.; Ho, D.; Sabino-Silva, R.; Siqueira, W.L.; Silikas, N. Fighting viruses with materials science: Prospects for antivirus surfaces, drug delivery systems and artificial intelligence. Dent. Mater. 2021, 37, 496–507. [Google Scholar] [CrossRef]
- Asano, J.; Chiba, K.; Tada, M.; Yoshii, T. Antiviral activity of lignans and their glycosides from Justicia procumbens. Phytochemistry 1996, 42, 713–717. [Google Scholar] [CrossRef]
- Chen, H.-W.; Cheng, J.X.; Liu, M.-T.; King, K.; Peng, J.-Y.; Zhang, X.-Q.; Wang, C.-H.; Shresta, S.; Schooley, R.T.; Liu, Y.-T. Inhibitory and combinatorial effect of diphyllin, a v-ATPase blocker, on influenza viruses. Antiviral Res. 2013, 99, 371–382. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Chang, W.-S.; Fang, Z.-S.; Chen, Y.-T.; Wang, W.-L.; Tsai, H.-H.; Chueh, L.-L.; Takano, T.; Hohdatsu, T.; Chen, H.-W. Nanoparticulate vacuolar ATPase blocker exhibits potent host-targeted antiviral activity against feline coronavirus. Sci. Rep. 2017, 7, 13043. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Chen, Y.-T.; Fang, Z.-S.; Chang, W.-S.; Chen, H.-W. Antiviral efficacy of nanoparticulate vacuolar ATPase inhibitors against influenza virus infection. Int. J. Nanomed. 2018, 13, 8579–8593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-J.; Rumschlag-Booms, E.; Guan, Y.-F.; Liu, K.-L.; Wang, D.-Y.; Li, W.-F.; Nguyen, V.H.; Cuong, N.M.; Soejarto, D.D.; Fong, H.H.S.; et al. Anti-HIV diphyllin glycosides from Justicia gendarussa. Phytochemistry 2017, 136, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, A.; Anantpadma, M.; Baker, L.; Raghavendra, N.M.; Davey, R.; Davisson, V.J. Phenotypic Prioritization of Diphyllin Derivatives That Block Filoviral Cell Entry by Vacuolar (H+)-ATPase Inhibition. ChemMedChem 2018, 13, 2664–2676. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Kielian, M. Flaviviruses: Braking the entering. Curr. Opin. Virol. 2013, 3, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Persaud, M.; Martinez-Lopez, A.; Buffone, C.; Porcelli, S.A.; Diaz-Griffero, F. Infection by Zika viruses requires the transmembrane protein AXL, endocytosis and low pH. Virology 2018, 518, 301–312. [Google Scholar] [CrossRef]
- Smit, J.; Moesker, B.; Rodenhuis-Zybert, I.; Wilschut, J. Flavivirus Cell Entry and Membrane Fusion. Viruses 2011, 3, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lopez, A.; Persaud, M.; Chavez, M.P.; Zhang, H.; Rong, L.; Liu, S.; Wang, T.T.; Sarafianos, S.G.; Diaz-Griffero, F. Glycosylated diphyllin as a broad-spectrum antiviral agent against Zika virus. EBioMedicine 2019, 47, 269–283. [Google Scholar] [CrossRef]
- Stefanik, M.; Strakova, P.; Haviernik, J.; Miller, A.D.; Ruzek, D.; Eyer, L. Antiviral Activity of Vacuolar ATPase Blocker Diphyllin against SARS-CoV-2. Microorganisms 2021, 9, 471. [Google Scholar] [CrossRef]
- Plescia, C.B.; Lindstrom, A.R.; Quintero, M.V.; Keiser, P.; Anantpadma, M.; Davey, R.; Stahelin, R.V.; Davisson, V.J. Evaluation of Phenol-Substituted Diphyllin Derivatives as Selective Antagonists for Ebola Virus Entry. ACS Infect. Dis. 2022, 8, 942–957. [Google Scholar] [CrossRef]
- Toth, G.; Horvati, K.; Kraszni, M.; Ausbuttel, T.; Palyi, B.; Kis, Z.; Mucsi, Z.; Kovacs, G.M.; Bosze, S.; Boldizsar, I. Arylnaphthalene Lignans with Anti-SARS-CoV-2 and Antiproliferative Activities from the Underground Organs of Linum austriacum and Linum perenne. J. Nat. Prod. 2023, 86, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Thamburaj, S.; Ramaraj, E.; Sethupathy, S.; Kamalanathan, C.; Raji, A.; Rajasekharan, S.K. Antibacterial and antibiofilm activities of diphyllin against fish pathogens. Microb. Pathog. 2020, 145, 104232. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.F.; Wang, Z.Q.; Liu, J.; Yu, H.T.; Lin, Q.W.; Chen, W.M.; Lin, J. Design and Synthesis of Novel c-di-GMP G-Quadruplex Inducers as Bacterial Biofilm Inhibitors. J. Med. Chem. 2021, 64, 11074–11089. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Xu, X.F.; Hou, W.; Meng, Y.; Huang, M.Y.; Lin, J.; Chen, W.M. Synthetic cajaninstilbene acid derivatives eradicate methicillin-resistant Staphylococcus aureus persisters and biofilms. Eur. J. Med. Chem. 2021, 224, 113691. [Google Scholar] [CrossRef]
- Day, S.H.; Chiu, N.Y.; Tsao, L.T.; Wang, J.P.; Lin, C.N. New Lignan Glycosides with Potent Antiinflammatory Effect, Isolated from Justicia ciliata. J. Nat. Prod. 2000, 63, 1560–1562. [Google Scholar] [CrossRef]
- Duan, Y.-N.; Ge, X.; Jiang, H.-W.; Zhang, H.-J.; Zhao, Y.; Li, J.-L.; Zhang, W.; Li, J.-Y. Diphyllin Improves High-Fat Diet-Induced Obesity in Mice through Brown and Beige Adipocytes. Front. Endocrinol. 2020, 11, 592818. [Google Scholar] [CrossRef]
- Löfvall, H.; Newbould, H.; Karsdal, M.A.; Dziegiel, M.H.; Richter, J.; Henriksen, K.; Thudium, C.S. Osteoclasts degrade bone and cartilage knee joint compartments through different resorption processes. Arthritis Res. Ther. 2018, 20, 67. [Google Scholar] [CrossRef]
- Sorensen, M.G.; Henriksen, K.; Neutzsky-Wulff, A.V.; Dziegiel, M.H.; Karsdal, M.A. Diphyllin, a novel and naturally potent V-ATPase inhibitor, abrogates acidification of the osteoclastic resorption lacunae and bone resorption. J. Bone Miner. Res. 2007, 22, 1640–1648. [Google Scholar] [CrossRef]
- Indrawinata, K.; Argiropoulos, P.; Sugita, S. Structural and functional understanding of disease-associated mutations in V-ATPase subunit a1 and other isoforms. Front. Mol. Neurosci. 2023, 16, 1135015. [Google Scholar] [CrossRef]
- Wang, H.; Rubinstein, J.L. CryoEM of V-ATPases: Assembly, disassembly, and inhibition. Curr. Opin. Struct. Biol. 2023, 80, 102592. [Google Scholar] [CrossRef]
- Collins, M.P.; Forgac, M. Regulation and function of V-ATPases in physiology and disease. BBA-Biomembranes 2020, 1862, 183341. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Parshad, B.; Prasad, A.K.; Bhatia, S. Design, Synthesis, and Biomedical Applications of Glycotripods for Targeting Trimeric Lectins. Eur. J. Org. Chem. 2023, 26, e202201360. [Google Scholar] [CrossRef]
- Hou, W.; Dai, W.; Huang, H.; Liu, S.-L.; Liu, J.; Huang, L.-J.; Huang, X.-H.; Zeng, J.-L.; Gan, Z.-W.; Zhang, Z.-Y.; et al. Pharmacological activity and mechanism of pyrazines. Eur. J. Med. Chem. 2023, 258, 115544. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Huang, H.; Wu, X.-Q.; Lan, J.-X. Bioactivities and mechanism of action of securinega alkaloids derivatives reported prior to 2022. Biomed. Pharmacother. 2023, 158, 114190. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Number in Original References | Biological Tests and Assays | Anti-Tumor Mechanisms | References |
|---|---|---|---|
| 1, 5, and 7 | MTT | unreported | [9] |
| diphyllin (3) | MTT | unreported | [10] |
| justicidin A (5), diphyllin (3), and tuberculatin (4) | MTT, ELISA kit | tumor necrosis factor-α augment | [11] |
| cleistanone (1), 7 | MTT | unreported | [12] |
| 2-6 | MTT, flow cytometry, P388 leukemia model | G0/G1 phase blockade | [13] |
| 2-5 | MTS | unreported | [14] |
| 1, 3, and 4 | MTT, PI and EB staining, Western blot | mitochondrial-mediated apoptosis induction, Bcl-2 decrease, Bax increase | [15] |
| 5 and 6 | MTT, kDNA decatenation assay, tubulin polymerization assay | Topo II and tubulin inhibition | [16] |
| 3 | MTT, Western blot | Topo II and HADC1 inhibition | [17] |
| 5d | MTT, Western blot | mitochondrial-mediated apoptosis induction, Bcl-2 decrease, Bax increase | [18] |
| 3a and 3e | MTT, colorimetric assay | vacuolar H+-ATPase inhibition | [19] |
| 3a | MTT, colorimetric assay | vacuolar H+-ATPase inhibition | [20] |
| ZT-25 | MTT, Western blot | G1/G0 phase blockade, mitochondrial membrane potential dissipation, ATP depletion, Bcl-2 downregulation, Bax and cleaved caspase-3 upregulation. Autophagy induction, LC3 I to LC3 II, Beclin-1 increase, and p-mTOR decrease | [22] |
| 1e | MTT, lysosome acidity assay | vacuolar H+-ATPase inhibition | [23] |
| 1, 4, and 7 | MTT, transwell invasion assay | unreported | [24] |
| 2I | MTT, vacuolar H+-ATPase activity assay kit | vacuolar H+-ATPase inhibition | [25] |
| diphyllin, 5c3, and 5c4 | MTT, vacuolar H+-ATPase activity assay kit, molecular docking | vacuolar H+-ATPase inhibition | |
| diphyllin | MTT, vacuolar H+-ATPase activity assay kit, Western blot, real-time PCR | Wnt/β-catenin signaling inhibition (phospho-LRP6 and β-catenin expression inhibition, c-Myc and cyclin D1 gene downregulation) | [27] |
| 15 | Sulforhodamine B (SRB), MTT, DNA fragmentation electrophoresis, flow cytometry | topoisomerase II inhibition, G0/G phase arrest | [28] |
| arylnaphthalene lignan (HJC) | MTT, Western blot, real-time PCR | activation of caspase 3 cascade, SOD inhibition, ROS elevation | [47] |
| HJB, HJA, JB, and CME | MTT, SOD activity assay kit, flow cytometry, Western blot | SOD inhibition, ROS elevation, caspase-dependent intrinsic and/or extrinsic apoptosis pathways | [29] |
| 1 and 7 | MTT, Western blot, Topoisomerase II assay, hollow fiber assay | caspase 3-dependent apoptosis | [30] |
| Cleistanthin-A and Cleistanthoside A tetraacetate | MTT, vacuolar H+-ATPase activity assay kit | vacuolar H+-ATPase inhibition | [31] |
| 5 | MTT | unreported | [32] |
| Justicidin B | MTT, Western blot | Bax/Bcl-2 ratio augment and caspase-3/7 activation | [33] |
| Cleistanthin A | MTT, Western blot, vacuolar H+-ATPase activity assay kit | MMP-2 and MMP-9 inhibition, vacuolar H+-ATPase inhibition | [34] |
| diphyllin | MTT, real-time PCR | decreased the mRNA expressions of mTORC1, HIF-1α, and VEGF | [35] |
| 7c | MTT | unreported | [37] |
| M3 | MTT, Western blot, CytoDYNAMIX Screen 03 Tubulin Polymerization assay | caspase 3-dependent apoptosis, promotion of microtubule depolymerization | [36] |
| TAARD | MTT, Western blot | TLR1/NF-κB and TLR3/STAT3 pathway inhibition | [38] |
| Cleistanthin A | MTT, Western blot, real-time PCR | β-catenin pathway inhibition | [39] |
| ECDD-S27 | MTT, immunoblot analysis | restrain of autophagic flux and inhibition of vacuolar H+-ATPase activity | [40] |
| diphyllin derivatives | MTT, UF-LC/MS screening assay, molecular docking | COX-2 inhibition | [41] |
| 15 | MTT, Western blot, flow cytometry, mouse pancreatic cancer model | G0/G1 phase blockade, CDK4, and cyclin D1 decrease | [42] |
| PHY34 | MTT, mass spectrometry-based chemoproteomics | targeting ATP6V0A2 subunit to induce autophagy inhibition and interacting with CAS to alter the nuclear localization of proteins | [44] |
| 3 | MTT, transwell invasion and scratch wound assay, vacuolar H+-ATPase activity assay kit | vacuolar H+-ATPase inhibition | [45] |
| diphyllin | MTT, AO/EB dual staining assay, flow cytometry, wound-healing assay | apoptosis induction | [46] |
| Compound Number in Original References | Biological Tests and Assays | Anti-Virus Mechanisms | References |
|---|---|---|---|
| diphyllin (5), diphyllin apioside (6, and diphyllin pioside-5-acetate (7) | MIC test | unreported | [49] |
| diphyllin | MTT, acridine orange labeling assay, fluorescence microscopy assay, RT-PCR, Western blot, TCID50 assay, hemagglutination test, plaque assay, CPE inhibition assay | vacuolar H+-ATPase inhibition | [50] |
| diphyllin | MTT, ADE model of FIPV infection, blood chemistry analysis | vacuolar H+-ATPase inhibition | [51] |
| diphyllin | transmission electron microscopy and dynamic light scattering assay, cell cytopathic effect inhibition assay, mouse model of the sublethal influenza challenge | vacuolar H+-ATPase inhibition | [52] |
| justiprocumins A and B | luciferase gene reporter assay | unreported | [53] |
| 2e, 2g, 2h | pseudovirus assays, MTT, vacuolar H+-ATPase inhibition | vacuolar H+-ATPase inhibition | [54] |
| 6-deoxyglucose-diphyllin (DGP) | acridine orange staining assay, MTT, qRT-PCR | acidification of endosomal/lysosomal compartments prevention | [58] |
| diphyllin, diphyllinoside cleistanthin B, helioxanthin 8-1, and helioxanthin 5-4-2 | viral titer reduction assay, MTT, plaque assays, immunofluorescence staining | unreported | [59] |
| diphyllin, diphyllinosides (Cleistanthin B 8) | acridine orange staining assay, GFP-EBOV virus infection assay, GFP-VLP uptake and trafficking assay, immunofluorescence staining, confocal imaging | vacuolar H+-ATPase inhibition | [60] |
| diphyllin, justicidin B (2), linadiacin A (3), linadiacin B (4) | MTT, TCID50 value determination | unreported | [61] |
| Compound Number in Original References | Activity | Biological Tests and Assays | Mechanisms | References |
|---|---|---|---|---|
| diphyllin | antibacterial and antibiofilm | MIC, MBC, biofilm disruption assay, ROS and PI staining | membrane damage and intracellular DNA degradation, ROS generation | [62] |
| ciliatoside A and ciliatoside B | anti-inflammation | LPS-treated RAW 264.7 cell model | unreported | [65] |
| diphyllin | anti-obesity | SRB | differentiation and thermogenesis induction | [66] |
| diphyllin | anti-rheumatic diseases | colorimetric calcium assay, TRAP activity measurement, von Kossa staining | vacuolar-H+-ATPase inhibition, osteochondral CTX-I release inhibition, Ca2+ concentration and TRAP activity decrease, and cell viability increase | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, W.; Huang, L.-J.; Huang, H.; Liu, S.-L.; Dai, W.; Li, Z.-M.; Zhang, Z.-Y.; Xin, S.-Y.; Wang, J.-Y.; Zhang, Z.-Y.; et al. Bioactivities and Mechanisms of Action of Diphyllin and Its Derivatives: A Comprehensive Systematic Review. Molecules 2023, 28, 7874. https://doi.org/10.3390/molecules28237874
Hou W, Huang L-J, Huang H, Liu S-L, Dai W, Li Z-M, Zhang Z-Y, Xin S-Y, Wang J-Y, Zhang Z-Y, et al. Bioactivities and Mechanisms of Action of Diphyllin and Its Derivatives: A Comprehensive Systematic Review. Molecules. 2023; 28(23):7874. https://doi.org/10.3390/molecules28237874
Chicago/Turabian StyleHou, Wen, Le-Jun Huang, Hao Huang, Sheng-Lan Liu, Wei Dai, Zeng-Min Li, Zhen-Yu Zhang, Su-Ya Xin, Jin-Yang Wang, Zi-Yun Zhang, and et al. 2023. "Bioactivities and Mechanisms of Action of Diphyllin and Its Derivatives: A Comprehensive Systematic Review" Molecules 28, no. 23: 7874. https://doi.org/10.3390/molecules28237874