

The Protective Effect of (-)-Tetrahydroalstonine against OGD/R-Induced Neuronal Injury via Autophagy Regulation
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
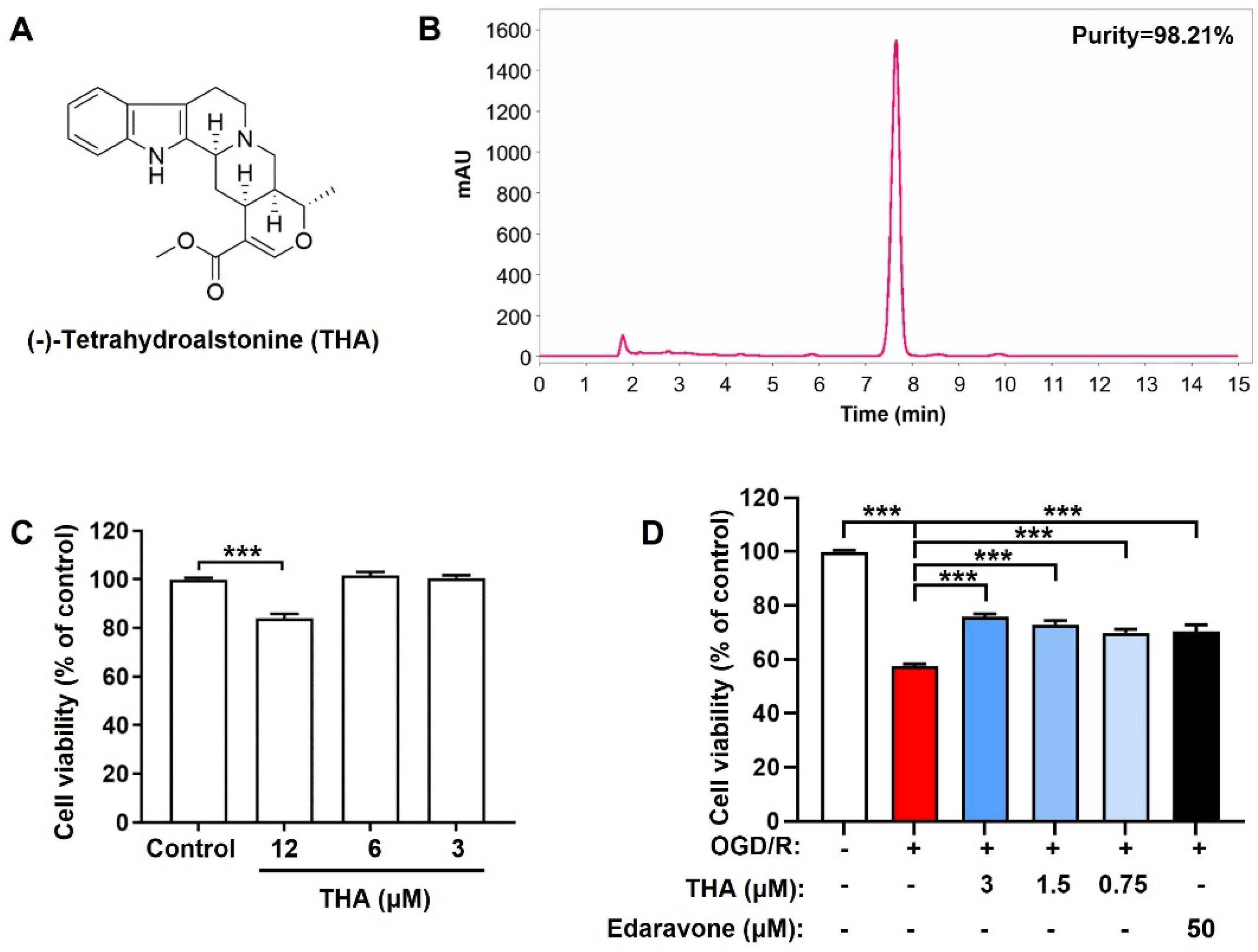
2.1. THA Alleviates OGD/R-Induced Neuronal Injury
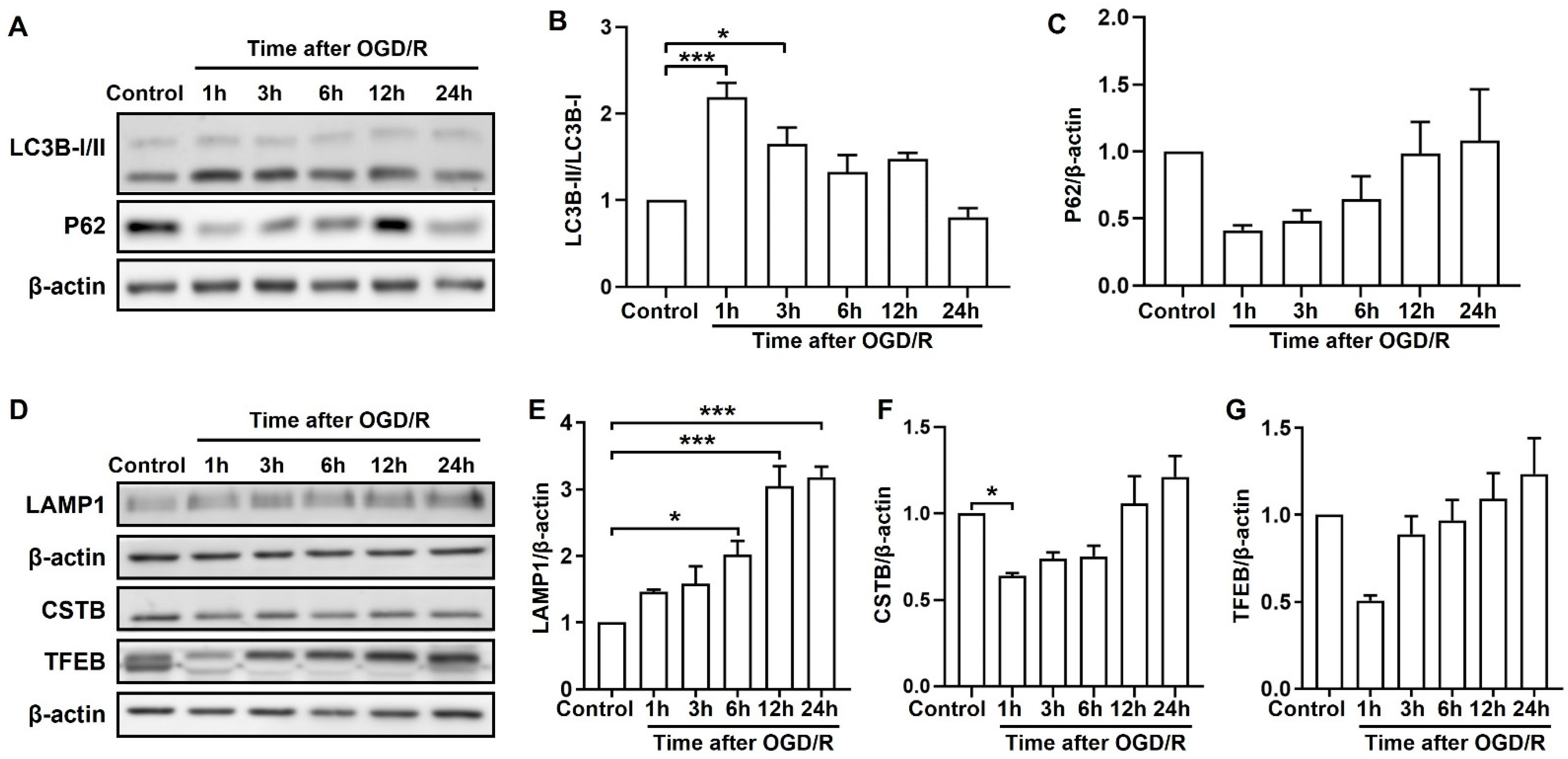
2.2. OGD/R Induces Autophagic and Lysosomal Dysfunction at Different Stages
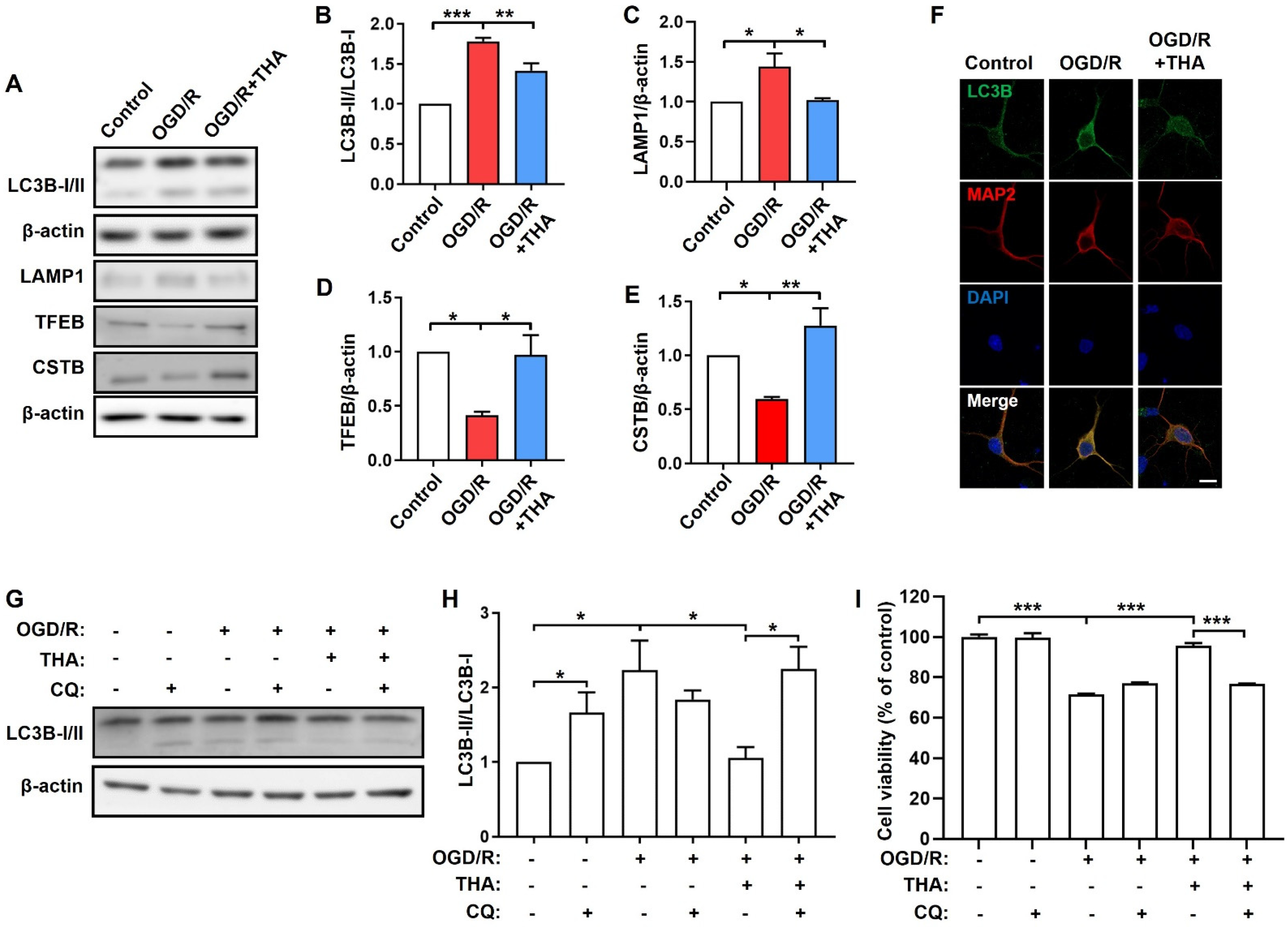
2.3. THA against OGD/R-Induced Neuronal Damage via Promoting Lysosomal Function
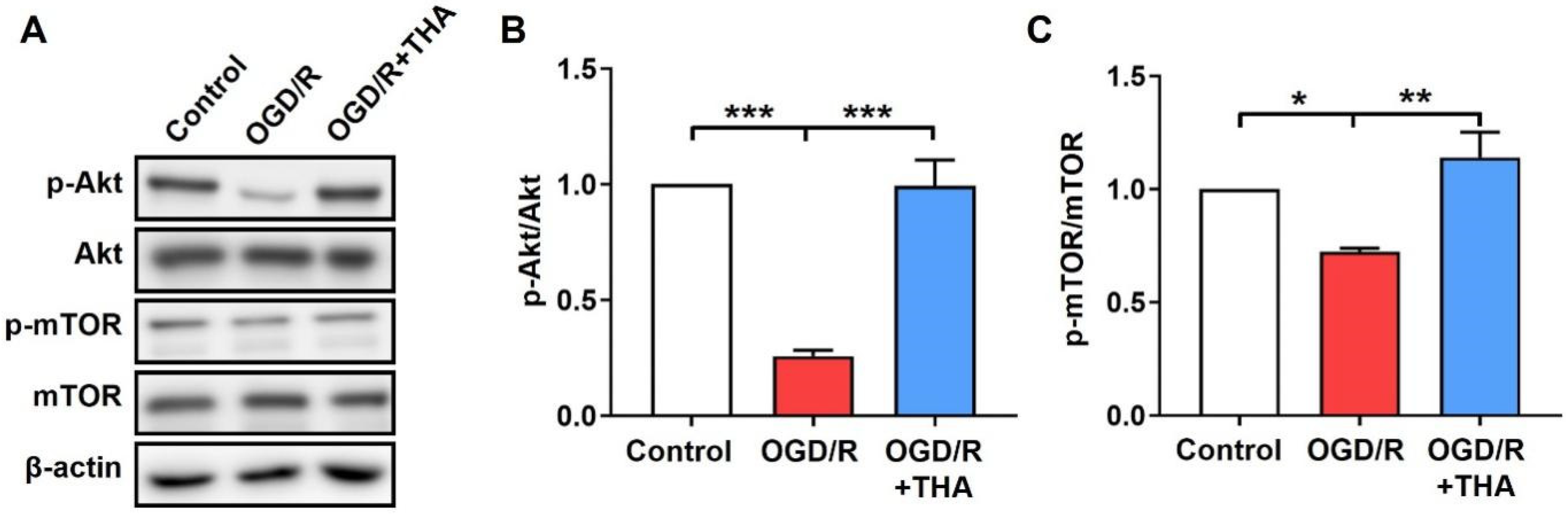
2.4. THA Ameliorates the OGD/R-Induced Inhibition of Akt/mTOR Pathway
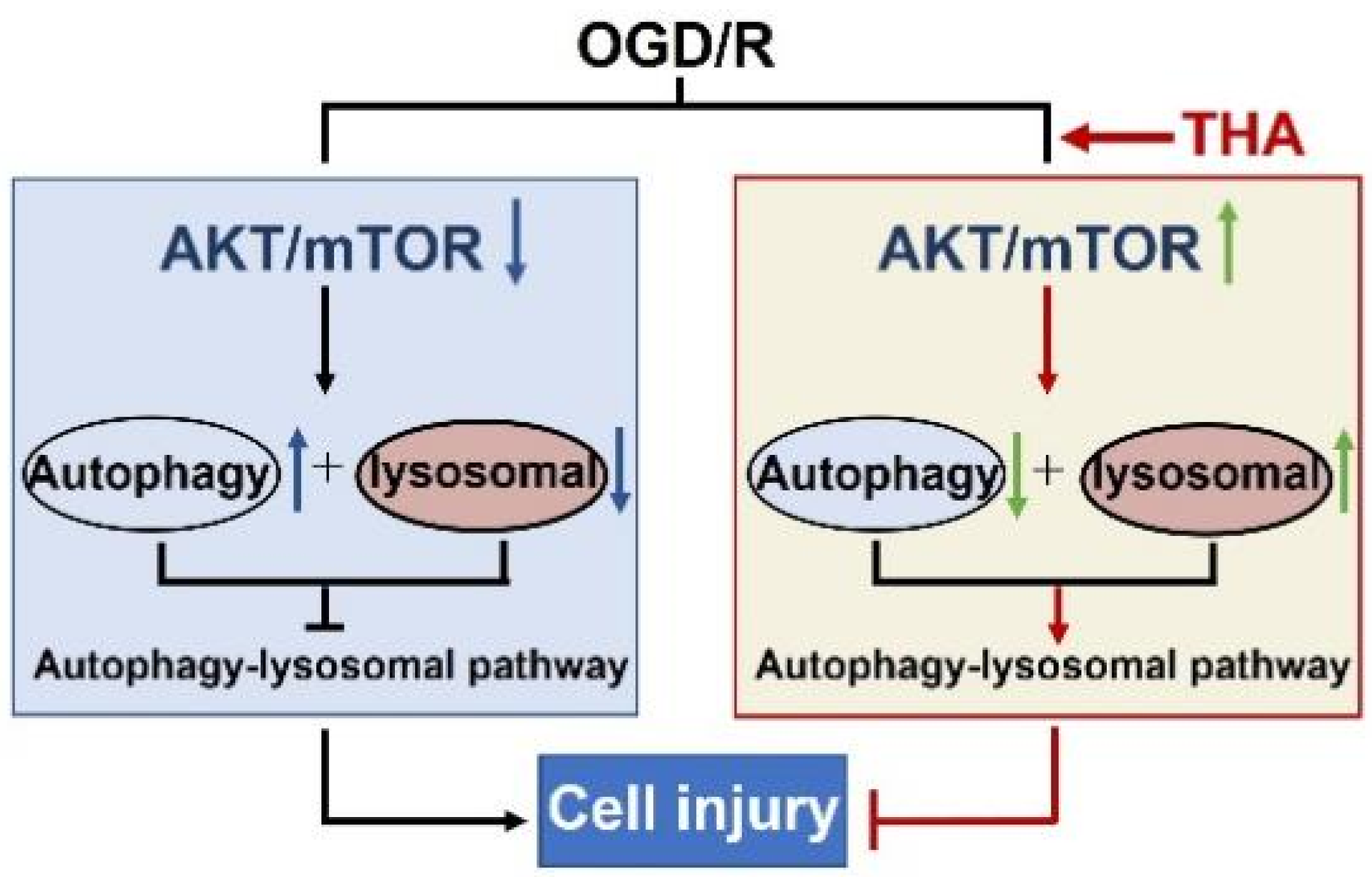
3. Discussion
4. Materials and Methods
4.1. Isolation and Purification of THA
4.2. Chemicals, Reagents and Antibodies
4.3. Primary Cortical Neuron Culturing
4.4. OGD/R and Drug Exposure
4.5. Cell Viability Assay
4.6. Western Blot Analysis
4.7. Immunofluorescence
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef]
- Han, G.J.; Min, X.Z.; Ma, S.S.; Ding, C.; Wang, X.Q. Xuesaitong combined with dexmedetomidine improves cerebral ischemia-reperfusion injury in rats by activating Keap1/Nrf2 signaling and mitophagy in hippocampal tissue. Oxid. Med. Cell Longev. 2022, 2022, 5126042. [Google Scholar] [CrossRef] [PubMed]
- Sekerdag, E.; Solaroglu, I.; Gursoy-Ozdemir, Y. Cell death mechanisms in stroke and novel molecular and cellular treatment options. Curr. Neuropharmacol. 2018, 16, 1396–1415. [Google Scholar] [CrossRef] [PubMed]
- Wollert, T. Autophagy. Curr Biol. 2019, 29, R671–R677. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, M.; Fan, J.; Yan, W.; Zha, X.; Song, H.; Wan, R.; Yin, Y.; Wang, W. Ischemia-induced upregulation of autophagy preludes dysfunctional lysosomal storage and associated synaptic impairments in neurons. Autophagy 2021, 17, 1519–1542. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Xu, D.; Li, F.; Chen, S.; Li, Y. The progress of neuronal autophagy in cerebral ischemia stroke: Mechanisms, roles and research methods. J. Neurol. Sci. 2019, 400, 72–82. [Google Scholar] [CrossRef]
- Ahsan, A.; Zheng, Y.; Ma, S.; Liu, M.; Cao, M.; Li, Y.; Zheng, W.; Zhou, X.; Xin, M.; Hu, W.W.; et al. Tomatidine protects against ischemic neuronal injury by improving lysosomal function. Eur. J. Pharmacol. 2020, 822, 173280. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Easson, M.E.A.M.; Simionescu, R.; Hajicek, J.; Thamm, A.M.K.; Salim, V.; De Luca, V. Solution of the multistep pathway for assembly of corynanthean, strychnos, iboga, and aspidosperma monoterpenoid indole alkaloids from 19E-geissoschizine. Proc. Natl. Acad. Sci. USA 2018, 115, 3180–3185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, M.; Ao, Y.L.; Li, Y.; Zou, X.; Xu, J.; Wang, Y.; Zhang, D.; Zhang, X.; Ye, W. Alstolarines A and B, two unusual monoterpenoid indole alkaloids with an acetal moiety from Alstonia scholaris. Org. Chem. Front. 2020, 7, 3468–3473. [Google Scholar] [CrossRef]
- Liu, Z.W.; Tang, B.Q.; Zhang, Q.H.; Wang, W.J.; Huang, X.J.; Zhang, J.; Shi, L.; Zhang, X.Q.; Ye, W.C. Ervaoffines E-G, three iboga-type alkaloids featuring ring C cleavage and rearrangement from Ervatamia officinalis. RSC Adv. 2017, 7, 21883–21889. [Google Scholar] [CrossRef] [Green Version]
- Ge, D.; Tao, H.R.; Fang, L.; Kong, X.Q.; Han, L.N.; Li, N.; Xu, Y.X.; Li, L.Y.; Yu, M.; Zhang, H. 11-methoxytabersonine induces necroptosis with autophagy through AMPK/mTOR and JNK pathways in human lung cancer cells. Chem. Pharm. Bull. 2020, 68, 244–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, R.; You, F.M.; Zhao, Q.; Xie, X.; Peng, C.; Zhan, G.; Han, B. Naturally derived indole alkaloids targeting regulated cell death (RCD) for cancer therapy: From molecular mechanisms to potential therapeutic targets. J. Hematol. Oncol. 2022, 15, 133. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Gou, Z.P.; Shang, J.H.; Li, W.Y.; Kuang, Y.; Li, M.Y.; Luo, X.D. Anti-microbial Effects In Vitro and In Vivo of Alstonia scholaris. Nat. Prod. Bioprospect. 2021, 11, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Jash, R.; Chowdary, K.A. Ethanolic extracts of Alstonia Scholaris and Bacopa Monniera possess neuroleptic activity due to anti-dopaminergic effect. Pharmacogn. Res. 2014, 6, 46–51. [Google Scholar]
- Singh, H.; Arora, R.; Arora, S.; Singh, B. Ameliorative potential of Alstonia scholaris (Linn.) R. Br. against chronic constriction injury-induced neuropathic pain in rats. BMC Complement Altern Med. 2017, 17, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Zhou, Z.; Chen, J.; Wang, Q.; Tang, P.; Ding, Q.; Yin, G.; Gu, J.; Fan, J. Edaravone inhibits autophagy after neuronal oxygen-glucose deprivation/recovery injury. Int. J. Neurosci. 2019, 129, 501–510. [Google Scholar] [CrossRef]
- Fu, X.; Liu, Y.; Zhang, H.; Yu, X.; Wang, X.; Wu, C.; Yang, J. Pseudoginsenoside F11 ameliorates the dysfunction of the autophagy-lysosomal pathway by activating calcineurin-mediated TFEB nuclear translocation in neuron during permanent cerebral ischemia. Exp. Neurol. 2021, 338, 113598. [Google Scholar] [CrossRef]
- Maiese, K. Targeting molecules to medicine with mTOR, autophagy and neuro-degenerative disorders. Br. J. Clin. Pharmacol. 2016, 82, 1245–1266. [Google Scholar] [CrossRef] [Green Version]
- Villa-González, M.; Martín-López, G.; Pérez-Álvarez, M.J. Dysregulation of mTOR signaling after brain ischemia. Int. J. Mol. Sci. 2022, 23, 2814. [Google Scholar] [CrossRef]
- Soltani, N.; Nazarian-Firouzabadi, F.; Shafeinia, A.; Sadr, A.S.; Shirali, M. The expression of Terpenoid Indole Alkaloid (TIAs) pathway genes in Catharanthus roseusin response to salicylic acid treatment. Mol. Biol. Rep. 2020, 47, 7009–7016. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Chen, Q.; Yang, G.F. A review on recent developments of indole-containing antiviral agents. Eur. J. Med. Chem. 2015, 89, 421–441. [Google Scholar] [CrossRef]
- Jin, Q.; Zhao, Y.L.; Liu, Y.P.; Zhang, R.S.; Zhu, P.F.; Zhao, L.Q.; Qin, X.J.; Luo, X.D. Anti-inflammatory and analgesic monoterpenoid indole alkaloids of Kopsia officinalis. J. Ethnopharmacol. 2022, 285, 114848. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.; Bhatt, H.; Judd, E.K.; Oparil, S.; Calhoun, D.A. Reserpine substantially lowers blood pressure in patients with refractory hypertension: A proof-of-concept study. Am. J. Hypertens. 2020, 33, 741–747. [Google Scholar] [CrossRef]
- Xie, Q.; Li, H.; Lu, D.; Yuan, J.; Ma, R.; Li, J.; Ren, M.; Li, Y.; Chen, H.; Wang, J.; et al. Neuroprotective effect for cerebral ischemia by natural products: A review. Front. Pharmacol. 2021, 12, 607412. [Google Scholar] [CrossRef]
- Li, L.; Su, Y.; Liu, J.; Chen, C. Efficacy of Vincamine treatment in a rat model of anterior ischemic optic neuropathy. Eur. J. Ophthalmol. 2021, 31, 3442–3449. [Google Scholar] [CrossRef]
- Sun, S.F.; Zhong, H.J.; Zhao, Y.L.; Ma, X.Y.; Luo, J.B.; Zhu, L.; Zhang, Y.T.; Wang, W.X.; Luo, X.D.; Geng, J.W. Indole alkaloids of Alstonia scholaris (L.) R. Br. alleviated nonalcoholic fatty liver disease in mice fed with high-fat diet. Nat. Prod. Bioprospect. 2022, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Shang, J.H.; Pu, S.B.; Wang, H.S.; Wang, B.; Liu, L.; Liu, Y.P.; Shen, H.M.; Luo, X.D. Effect of total alkaloids from Alstonia scholaris on airway inflammation in rats. J. Ethnopharmacol. 2016, 178, 258–265. [Google Scholar] [CrossRef]
- Mo, Y.; Sun, Y.Y.; Liu, K.Y. Autophagy and inflammation in ischemic stroke. Neural. Regen. Res. 2020, 15, 1388–1396. [Google Scholar] [PubMed]
- Papadakis, M.; Hadley, G.; Xilouri, M.; Hoyte, L.C.; Nagel, S.; McMenamin, M.M.; Tsaknakis, G.; Watt, S.M.; Drakesmith, C.W.; Chen, R.; et al. Tsc1 (hamartin) confers neuroprotection against ischemia by inducing autophagy. Nat. Med. 2013, 19, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Wang, W.; Wang, X.; Wang, Y.; Xu, X.; Ping, F.; Du, Y.; Jiang, W.; Cui, D. BFGF plays a neuroprotective role by suppressing excessive autophagy and apoptosis after transient global cerebral ischemia in rats. Cell Death Dis. 2018, 9, 172. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xue, X.; Zhang, H.; Che, X.; Luo, J.; Wang, P.; Xu, J.; Xing, Z.; Yuan, L.; Liu, Y.; et al. Neuronal-targeted TFEB rescues dysfunction of the autophagy-lysosomal pathway and alleviates ischemic injury in permanent cerebral ischemia. Autophagy 2019, 15, 493–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Tang, Y.; Hu, Z.; Xu, W.; Ma, Y.; Xu, P.; Xing, H.; Niu, Q. The inhibition of TRPML1/TFEB leads to lysosomal biogenesis disorder, contributes to developmental fluoride neurotoxicity. Ecotoxicol. Environ. Saf. 2023, 250, 114511. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Zhang, M.; Feng, Y.S.; Xing, Y.; Tan, Z.X.; Li, W.B.; Dong, F.; Zhang, F. Electroacupuncture inhibits neuronal autophagy and apoptosis via the PI3K/AKT pathway following ischemic stroke. Front. Cell Neurosci. 2020, 14, 134. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Qu, T.; Luo, J.; An, P.; Ren, F.; Luo, Y.; Li, Y. mTORC2: A multifaceted regulator of autophagy. Cell Commun. Signal. 2023, 21, 4. [Google Scholar] [CrossRef]
- Shi, Q.; Zhang, Q.; Peng, Y.; Zhang, X.; Wang, Y.; Shi, L. A natural diarylheptanoid protects cortical neurons against oxygen-glucose deprivation-induced autophagy and apoptosis. J. Pharm. Pharmacol. 2019, 71, 1110–1118. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Shi, Q.; Li, L.; Zhang, Q.; Wu, Z.L.; Huang, X.J.; Zhang, Q.W.; Ye, W.C.; Wang, Y.; et al. Dimeric Diarylheptanoids with Neuroprotective Activities from Rhizomes of Alpinia officinarum. ACS Omega 2020, 5, 10167–10175. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.W.; Wu, D.P.; Liang, H.F.; Tang, G.Y.; Fan, C.L.; Shi, L.; Ye, W.C.; Li, M.M. Total Saponins of Panax notoginseng Activate Akt/mTOR Pathway and Exhibit Neuroprotection in vitro and in vivo against Ischemic Damage. Chin. J. Integr. Med. 2022, 28, 410–418. [Google Scholar] [CrossRef]
- Long, P.; Li, Y.; Wen, Q.; Huang, M.; Li, S.; Lin, Y.; Huang, X.; Chen, M.; Ouyang, J.; Ao, Y.; et al. 3′-Oxo-tabernaelegantine A (OTNA) selectively relaxes pulmonary arteries by inhibiting AhR. Phytomedicine 2021, 92, 153751. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, L.; Li, Z.; Huang, Z.; Yang, Y.; Liu, P.; Guo, D.; Li, Y.; Wu, T.; Luo, R.; et al. eEF2 in the prefrontal cortex promotes excitatory synaptic transmission and social novelty behavior. EMBO Rep. 2022, 23, e54543. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Wang, J.-Y.; Pan, Y.; Zou, X.; Wang, C.; Peng, Y.; Ao, Y.-L.; Lam, M.F.; Zhang, X.; Zhang, X.-Q.; et al. The Protective Effect of (-)-Tetrahydroalstonine against OGD/R-Induced Neuronal Injury via Autophagy Regulation. Molecules 2023, 28, 2370. https://doi.org/10.3390/molecules28052370
Liao Y, Wang J-Y, Pan Y, Zou X, Wang C, Peng Y, Ao Y-L, Lam MF, Zhang X, Zhang X-Q, et al. The Protective Effect of (-)-Tetrahydroalstonine against OGD/R-Induced Neuronal Injury via Autophagy Regulation. Molecules. 2023; 28(5):2370. https://doi.org/10.3390/molecules28052370
Chicago/Turabian StyleLiao, Yumei, Jun-Ya Wang, Yan Pan, Xueyi Zou, Chaoqun Wang, Yinghui Peng, Yun-Lin Ao, Mei Fong Lam, Xiaoshen Zhang, Xiao-Qi Zhang, and et al. 2023. "The Protective Effect of (-)-Tetrahydroalstonine against OGD/R-Induced Neuronal Injury via Autophagy Regulation" Molecules 28, no. 5: 2370. https://doi.org/10.3390/molecules28052370