
Enzymatic Carboxylation of Resorcinol in Aqueous Triethanolamine at Elevated CO2 Pressure

Institute of Technical Biocatalysis, Hamburg University of Technology, 21073 Hamburg, Germany
*
Authors to whom correspondence should be addressed.
Molecules 2024, 29(1), 25; https://doi.org/10.3390/molecules29010025
Submission received: 16 November 2023
/
Revised: 10 December 2023
/
Accepted: 14 December 2023
/
Published: 19 December 2023
(This article belongs to the Special Issue Contribution of Enzyme Catalysis to the Achievement of the UN Sustainable Development Goals)
Abstract
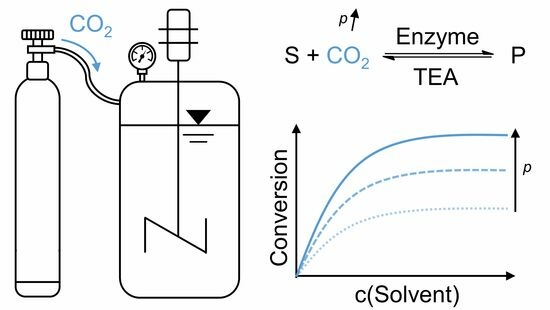
:The fixation of CO2 by enzymatic carboxylation for production of valuable carboxylic acids is one way to recycle carbon. Unfortunately, this type of reaction is limited by an unfavourable thermodynamic equilibrium. An excess of the C1 substrate is required to increase conversions. Solvents with a high CO2 solubility, such as amines, can provide the C1 substrate in excess. Here, we report on the effect of CO2 pressures up to 1100 kPa on the enzymatic carboxylation of resorcinol in aqueous triethanolamine. Equilibrium yields correlate to the bicarbonate concentration. However, inhibition is observed at elevated pressure, severely reducing the enzyme activity. The reaction yields were reduced at higher pressures, whereas at ambient pressure, higher yields were achieved. Overall, CO2 pressures above 100 kPa have been demonstrated to be counterproductive for improving the biotransformation, as productivity decreases rapidly for only a modest improvement in conversion. It is expected that CO2 carbamylation intensifies at elevated CO2 pressures, causing the inhibition of the enzyme. To further increase the reaction yield, the in situ product precipitation is tested by the addition of the quaternary ammonium salt tetrabutylammonium bromide.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Benzoic acid decarboxylases catalyse the decarboxylation of benzoic acids to their respective phenols, but they also work in the reverse direction to carboxylate phenols. Their ability to carboxylate various phenols selectively, such as resorcinol, resveratrol, and gnetol [1], via the utilization of CO2, attracted attention in the scientific community [2,3,4]. The major challenge of carboxylation reactions are the unfavourable thermodynamics, which prevent reaching high conversions under conventional conditions. CO2 has an especially high thermodynamic stability with a standard Gibbs free energy formation of −394 kJ mol−1 [5]. Carbon ligases (EC 6) overcome the energy barrier by performing substrate activation with energy-rich, but also expensive, cofactors such as ATP. The (de)carboxylases (EC 4) do not use cofactors and require different approaches to enhance the CO2 fixation. To make enzymatic carboxylations viable for industrial production of valuable carboxylated chemicals [4], research should focus on finding strategies to enhance the conversions. For example, the impact of the concentration of the C1 substrate, either by increasing the concentration of supplied bicarbonate salts, or increased CO2 partial pressures in aqueous systems, were studied [6]. However, the application of 2 M KHCO3 under ambient pressure and a CO2 partial pressure of 800 kPa only resulted in a conversion of around 25% for the carboxylation of catechol [6]. Substituting KHCO3 with other bicarbonate salts was found to have a strong impact on both the enzyme activity and the conversion. This dependency on bicarbonates was found to correlate to the Hofmeister series, but still, the conversion could not be sufficiently enhanced [7]. Higher conversions were reported when different organic solvents and ionic liquids were supplemented [7]. Using amines as solvents is particularly promising as amines capture gaseous CO2 and provide the biotransformation with a high C1 substrate concentration [8]. Recently, we showed that aqueous triethanolamine can be loaded very efficiently with gaseous CO2 by using microbubble sparging in a bubble column setup [9]. Moreover, we discovered that the equilibrium yield depends on the HCO3− species, which was the main species of the dissolved inorganic carbon (DIC) content [10]. Similarly, Fan et al. have recently shown that the carboxylation of catechol by a 2,3-dihydroxybenzoic acid decarboxylase from Aspergillus oryzae depends on the aqueous CO2 concentration, with a high concentration of HCO3− being required under alkaline conditions [11].
In contrast to approaches that increase the substrate concentration, the removal of the formed product can also increase the conversion. In the literature, several adsorber materials have been used [12,13]. However, none of these adsorbers are selective enough to remove the product without binding large amounts of substrate, too [12,13]. Another approach used enzyme cascades making use of the product from the carboxylation in subsequent reactions. This approach has been shown to be compatible for the production of amino acids [14]. However, this cascade needs to be optimized as the product titer achieved was very low, and the process is generally more expensive than traditional processes. Enzyme development and production costs were the main contributors to the overall process cost. A major improvement was achieved by Ren et al., who found that certain quaternary ammonium salts can selectively initiate crystallization of some benzoic acid derivatives, accomplishing an efficient in situ product removal enabling near-full conversions [15,16]. The main disadvantages of this technique are that a suitable pair of quaternary ammonium ions and benzoic acid derivatives must be found and that a large amount of bicarbonate salt is wasted. Dealing with unreacted bicarbonate salt is a concern that all such systems have in common. However, this can be mitigated by recycling the unused bicarbonate salt or even circumvented completely by using gaseous CO2 in combination with a reaction medium that has a high CO2 solubility. A promising system utilizes amines used for carbon capture in the industry to directly provide the C1 substrate for the biotransformation [10]. There, much higher conversions are already possible, and the combination with the previously mentioned quaternary ammonium precipitation method at low amine concentrations further increases conversions [10].
Here, we report on the combination of the aqueous amine system with increased CO2 pressure and quaternary ammonium precipitation and its effect on the investigated enzymatic carboxylation. As the model reaction system, the carboxylation of resorcinol by 2,6-dihydroxybenzoic acid (2,6-DHBA) decarboxylase (2,6-DHBD) from Rhizobium sp. in aqueous triethanolamine is investigated (Figure 1). An, up to now, unrecognized and critical inhibition is shown to limit the biotransformation at elevated CO2 pressures.
2. Results and Discussion
2.1. Carboxylation at Elevated CO2 Pressure
A usual strategy to increase conversion of the thermodynamic limited carboxylation is to perform the reaction under excess C1 substrate conditions. Compared to the conventional approach where the C1 substrate is supplied by addition of bicarbonate salts, the amine-mediated system utilizes gaseous CO2 to provide the C1 substrate. Bicarbonate salts are largely limited by their solubility in the particular medium, whereas the dissolved inorganic carbon (DIC) concentration is primarily dependent on the CO2 pressure.
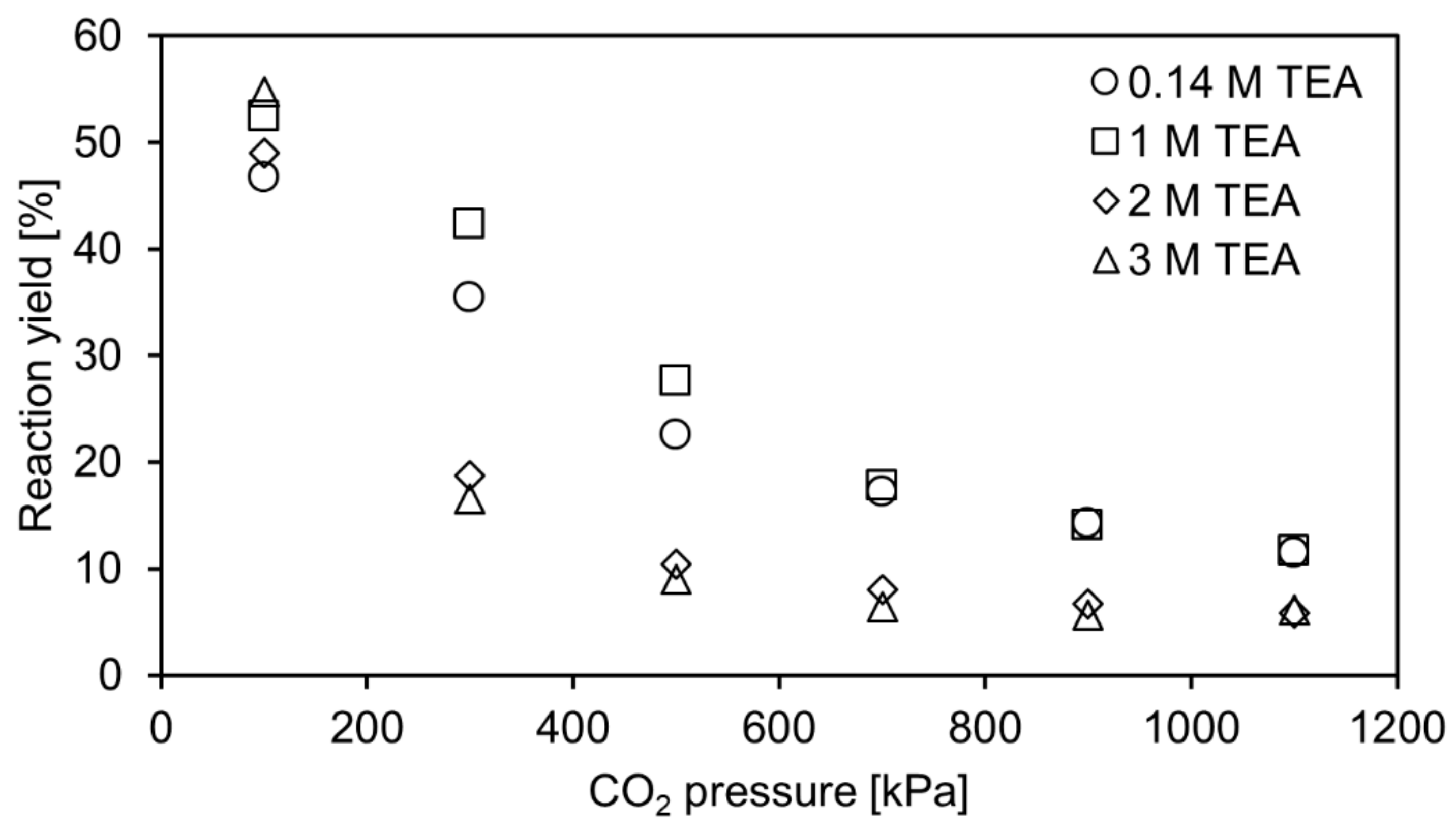
In this work, the carboxylation of resorcinol is investigated at pressures up to 1100 kPa of pure CO2 to determine the influence of the pressure on the reaction yield. For calculation of CO2 solubility in aqueous TEA, the correlation of Jou et al. (1985) is utilized [17]. Additionally, the pH of the reaction solution can be calculated with this correlation as it incorporates the method of Kent and Eisenberg (1976) using equilibrium constants to correlate solubility data [18]. This approach is applied to calculate theoretical concentrations of CO2/H2CO3, HCO3− and CO32− in aqueous TEA for the investigated pressure range. In combination with the equilibrium constant, determined in a previous work [10], theoretical equilibrium yields were calculated and compared with experimental reaction yields, as shown in Figure 2.
At 100 kPa, predicted and experimental equilibrium yields fit well. In our previous work [10], lower yields were reported for the investigated aqueous TEA concentrations. For example, at 3 M aqueous TEA, an equilibrium yield of around 40% was reported, whereas around 50% are achieved in this work. However, these values cannot be directly compared as a crucial condition is changed. In the previous work, aqueous TEA was only presaturated, and then resorcinol and 2,6-DHBD were added, diluting the CO2 saturated TEA. Thereby, these experiments do not correspond to conditions at 100 kPa CO2 pressure, which was accounted for in the study. For example, in Figure 5 (right) and Figure 6 of Ohde et al. (2021) [10], it was predicted that reaching 50% equilibrium yield is possible at a dissolved inorganic content DIC of around 1700 mM and 3 M aqueous TEA. A DIC of around 1700 mM can be reached at 3 M aqueous TEA at 100 kPa. In turn, reaching 50% equilibrium yield at 3 M aqueous TEA is confirmed in the current study (see Figure 2). This is achieved by pressurization of the reactor or flushing with CO2 in the case of the control reactor after the initial CO2 presaturation and start of the reaction. In this way, the intended CO2 pressure is reached.
Comparing the predicted and the experimental yields for the whole pressure range tested in this study, it is observed that the yield prediction fits the experimental data only for the lower aqueous TEA concentrations (approximately up to 1 M). At high TEA concentration, the applied model apparently overestimates the yields, with the difference between prediction and experimentally obtained data increasing at higher applied CO2 pressure. No side product was formed that might explain the difference, as conversion and yield were identical in all experiments. Another explanation for the deviation might be that the equilibrium reaction yield was not yet reached in the reaction after 2 days of reaction time. In this case, the reaction yield cannot fit the equilibrium yield, which is predicted by the applied model. Therefore, additional experiments are performed, where the reaction time is extended to check if the reaction yield would still increase. The measurements confirmed that the reaction equilibrium was not achieved after 2 and even 9 days under elevated CO2 pressure. A further control experiment was performed. Instead of testing the carboxylation, the decarboxylation of 80 mM 2,6-DHBA up to 1200 kPa was performed. The decarboxylation is favoured in terms of thermodynamics, but even after 10 days, the equilibrium yield was not reached as observed in the previous experiments. The additional feature of this experiment is that resorcinol-dependent deactivation of the enzyme can be neglected due to resorcinol not being present at the start and only accumulating in the course of the reaction.
Considering a different type of enzyme deactivation, for example, due to a pH shift, it is unlikely that a pH-dependent deactivation has a major negative impact on the enzyme stability and activity in the investigated pressure range. At 100 kPa, the pH was measured to be in the neutral, up to slightly alkaline pH range for higher TEA concentrations. As the experimental setup made it impossible to measure the pH at elevated pressure, the pH values were instead calculated with the applied model as described above. In Figure 3, the pH-pressure profile up to 1200 kPa is shown for the investigated aqueous TEA concentrations. Initially, the pH decreases rapidly until being buffered by the TEA and the bicarbonate equilibrium. In this way, the slightly alkaline pH is lowered to a more neutral pH by loading the aqueous TEA with CO2. Only when using 0.14 M TEA, the pH gets slightly acidic, reaching a value around pH 6 above 800 kPa pressure. Instead of having a negative effect, a positive effect is expected as the enzyme is reported to be most active around the neutral pH region [19].
An additional effect of the lowered pH is that the mole fraction and concentration of CO2 increases. In turn, this is expected to increase the enzyme activity as demonstrated in a previously published kinetic investigation [10]. In Figure 4, the concentration profile of the different DIC species is shown for 1 M aqueous TEA. It can be seen that both CO2 and HCO3− concentrations increase, whereas the CO32− concentration has a maximum around 6 kPa, where the pH is calculated to be 8.5. The increase in CO2 and HCO3− concentrations cannot readily explain the reduced enzyme activity, as CO2 is reported to be the most likely active C1 substrate, whereas HCO3− largely determines the equilibrium yield at a CO2 pressure of 100 kPa [10]. An inhibition seems to occur at high CO2 pressures, which could be related to reversible or irreversible carbamylation of amino acids within the enzyme structure. Prominent examples found in nature are ribulose-1,5-bisphosphate carboxylase/oxygenase and haemoglobin, which are both CO2 regulated. In haemoglobin, the N-terminal valine is carbamylated forming a carbamate [20,21] that modulates the affinity to bind oxygen at physiological conditions [22]. In the case of ribulose-1,5-bisphosphate carboxylase/oxygenase, both Mg2+ and inorganic carbon act as cofactors [23] enabling the enzyme to fix atmospheric CO2. In the underlying mechanism to activate the enzyme, CO2 needs to bind to an alternative site on the enzyme to form a carbamate as demonstrated in several studies [24,25,26].
In general, carbamate formation can occur in all proteins at the N-terminal amino acid and amino acids with a free amino group such as lysine [27]. The process is usually reversible, and it was found that CO2-mediated posttranslational modification of proteins is relevant even at physiological conditions and impacts protein biochemistry [28]. However, it is known that the three-dimensional structure of enzymes may be significantly changed at higher CO2 pressures, which affects their stability and may cause denaturation [29]. Consequently, a loss of their activity occurs. Supercritical CO2 treatment for enzyme deactivation is, for example, commonly applied in food processing [30,31]. The inactivation by supercritical CO2 could be predominantly attributed to the pH-lowering effect during treatment as stated by Balaban et al. [32]. As previously discussed, it is unlikely that a pH-dependent deactivation occurred due to only slight changes in pH being calculated due to TEA acting as a buffer. Nonetheless, deactivation can even occur at subcritical CO2 conditions [33]. However, the chance for minor structural changes is higher, which may induce a reversible alternative active protein state with altered enzyme activity, specificity, and stability [34]. As only conditions up to 1100 kPa were investigated in this work, it remains open if reversible or irreversible changes occurred that might explain the decrease in 2,6-DHBD activity. In a future study, it will be investigated whether the enzyme regains activity after an incubation at high CO2 pressure without substrate (resorcinol or 2,6-DHBA).
Overall, this inhibition prevented reaching the equilibrium yield during the investigated reaction time. The presented data demonstrate that performing enzymatic carboxylation under elevated pressure up to 1100 kPa is an unfeasible strategy to enhance equilibrium yields compared to ambient CO2 pressure conditions.
2.2. Precipitation at Elevated CO2 Pressure
Under ambient conditions (30 °C, atmospheric pressure), quaternary ammonium salts such as tetrabutylammonium (TBA) salts are reported to enhance the conversion of the enzymatic carboxylation of hydroxylated benzenes [15,16]. This effect was also reported to work in the aqueous TEA system; however, at greatly reduced efficiency [10]. This approach has now been tested at elevated CO2 pressure to see if pressure has a positive effect on the reaction yield. Therefore, the enzymatic carboxylation of resorcinol to 2,6-DHBA was performed with TBA-bromide (TBA-Br) at elevated pressure. In Figure 5, the determined reaction yields are shown. Overall, the addition of TBA-Br improves the reaction yield at 100 kPa CO2 pressure, especially for 0.14 M and 1 M TEA, which is consistent with previous data [10]. However, the results demonstrate that the reaction yield decreases at higher pressures with reduced yields. For example, the yields are reduced by only around 12% for 1 M TEA at 1100 kPa instead of by 35% without the addition of TBA-Br, as shown in Figure 5.
The underlying mechanism by which CO2 pressure causes a strong effect on the reduction of reaction yield when adding TBA-Br is not fully understood. It is important to note that the mass balance is closed for all experiments and no side product formation is detected. To ensure that the quantification by HPLC is not distorted by formed precipitates, trifluoroacetic acid was added directly to the reaction vessels to stop the reaction and dissolve any formed precipitate. Therefore, side product formation and product contained in any solids can be excluded to cause the yield reduction. A possible reason is that the increased CO2 carbamylation causes an enzyme inhibition, which prevents reaching the equilibrium yield as discussed above. It is known that blends of amines with quaternary ammonium salts have a higher CO2 solubility [35], which would enhance carbamate formation. This explains part of the observed behaviour. As a consequence of carbamates forming on the enzyme backbone, TBA is expected to enter in electrostatic interactions with the negatively charged carbamate. In turn, the enzyme structure is disrupted, causing a loss in activity. A detailed investigation is required to gain a deeper understanding of the underlying mechanism for the observed behaviour. However, at this stage, it is already clear that it is not feasible to perform the enzymatic carboxylation at elevated pressure when using TBA-Br or most likely any other quaternary ammonium salts.
3. Materials and Methods
3.1. General
All chemicals, except triethanolamine, were sourced from Sigma-Aldrich (Darmstadt, Germany) with purities of ≥99%. Triethanolamine was purchased from Carl Roth (Karlsruhe, Germany). As for CO2, carbon dioxide 4.5 (≥99.995%), purchased from Linde (Pullach, Germany), was utilized.
3.2. Production of His-Tagged 2,6-Dihydroxybenzoic Acid Decarboxylase
The original plasmid vector (pET21a+) containing the gene for 2,6-dihydroxybenzoic acid decarboxylase from Rhizobium sp. strain MTP-10005 was provided by Prof. Kurt Faber and Dr. Silvia Glueck (Graz University, Graz, Austria). The preparation of the plasmid is described in Wuensch et al. (2013) [7]. For the transformation, E. coli BL21(DE3) cells were used. These transformed cells were provided by Dr. Lorenzo Pesci (Hamburg University of Technology, Hamburg, Germany). The detailed transformation procedure is described in Pesci et al. (2015) [6].
Overexpression of 2,6-DHBD was performed in a Luria Bertani [36] medium supplemented with 100 µg/mL of ampicillin in shake flasks according to Pesci et al. (2015) [6]. Cells were harvested by centrifugation at 8000 rpm (11,325 rcf) for 20 min at 4 °C. Cells were washed with 50 mM KPi pH 7.5, and the previously described centrifugation step was repeated. Several of these washing steps were performed, where the supernatant was discarded.
Cell-free extract was prepared by suspending wet cells in 50 mM KPi pH 7.0 to a concentration of 4 g/mL and performing a cell disruption by sonication. The sonication was performed with a 3 mm tip at 70% power using a BANDELIN Sonopuls from BANDELIN electronic (Berlin, Germany). Overall, three cycles were performed, each consisting of three minutes of sonication and a subsequent cooling of the sample on ice for three minutes. After sonication, the sample was centrifuged at 20,000 rpm (48,384 rcf) for 20 min at 4 °C. The obtained supernatant was filtrated using a 0.45 μm filter. Afterwards, the cell-free extract was cooled at 4 °C for subsequent enzyme purification.
For enzyme purification, immobilized metal affinity chromatography in a packed bed column using 90 μm beads of Nickel Sepharose® 6 Fast Flow from Cytiva, Marlborough, MA, USA was performed according to Pesci et al. (2015) [6]. For the final buffer exchange, 50 mM KPi pH 7.0 was used. The purified enzyme was aliquoted and stored at −20 °C. Several batches of purified 2,6-DHBD were produced and combined.
3.3. Enzymatic Reactions at Elevated CO2 Pressure
In all experiments, purified 2,6-DHBD from one combined batch was used. Reactions were carried out in open 1.5 mL HPLC screw cap vials placed in high-pressure reactors consisting of stainless steel cylinders capped with bronze lids. Several vials can be placed in these reactors, which have a volume of 160 mL and can be operated at pressures of up to 400 bar.
First, reaction mixtures without purified 2,6-DHBD were presaturated with CO2 by bubbling through a cannula acting as a sparger until the pH was constant. Then, the enzyme was added to start the reaction. Afterwards, 80 mM of resorcinol and 16.4 U/mg enzyme were used as final concentrations. The enzyme activity was determined according to Ohde et al. (2021) [10]. Immediately after starting the reaction, the vials were placed in the high-pressure reactor, which was flushed three times with CO2 to remove air. Then, the reactors were pressurized with CO2. Each batch was incubated at room temperature without shaking or stirring.
Control experiments were performed in aluminium reactors. Similar to the high-pressure reactors, these reactors have a volume of 160 mL and were flushed with CO2, as well. In this way, the condition for ~100 kPa CO2 pressure was ensured.
3.4. Enzymatic Reactions at Elevated CO2 Pressure with TBA-Br
In situ product precipitation experiments were performed similar to the procedure described in Section 3.3. The only difference was that resorcinol and tetrabutylammonium bromide were added after CO2 saturation of aqueous TEA solutions. This was done to prevent precipitates already forming during CO2 gassing, as was the case when resorcinol and TBA-Br were added beforehand. The final concentration of resorcinol and TBA-Br in the reactions were 80 mM and 350 mM, respectively. The reaction was started by adding 2,6-DHBD (16.4 U/mg).
3.5. Analytics
An RP-HPLC method was used for quantification of resorcinol and 2,6-DHBA. The sample preparation was created by mixing a sample in a ratio of 1:1 with trifluoroacetic acid (TFA). This stopped the reaction and removed amine-bound CO2. For samples containing TBA-Br, water and acetonitrile were added in a volume ratio of 1:1:1:1 (sample:TFA:water:ACN). This was done to dissolve formed precipitates. After mixing and complete degassing, which was indicated by no more (CO2) bubbles being formed, a centrifugation was performed for 5 min at 13,000 rpm. The supernatant was analysed in an Agilent (Waldbronn, Germany) LC-1100 HPLC system. For separation, a LichroCART 250-4 Lichrospher 100 RP column (5 μm) was used. For analyte detection, the HPLC was equipped with a diode array detector. A gradient method was applied using solutions of acetonitrile with 0.1% (v/v) TFA and water with 0.1% (v/v) TFA. The applied gradient is described in a previously reported protocol [37]. Typical retention times were 7.0 min and 8.3 min for resorcinol and 2,6-DHBA, respectively.
4. Conclusions
The enzymatic carboxylation of resorcinol to 2,6-dihydroxybenzoic acid can be enhanced by the application of elevated CO2 pressure. However, when applying pressures above 100 kPa CO2, the increase in conversion is not sufficient to justify the application of higher pressure. This is especially supported by two reasons. First, there would be the need for a more expensive setup that supports biotransformations at elevated pressure. Second, it was found that elevated pressure up to 1100 kPa has a counterproductive effect on the enzyme activity, which results in a strong reduction of the carboxylation reaction rate. Due to this, this strategy is unfeasible, and not even the equilibrium yield is reached without adding enzymes in excess. Attempting to enhance conversion via the addition of TBA-Br for in situ product precipitation reduced the conversion at elevated pressure to values below the ones obtained when not using the quaternary ammonium salt. It is expected that CO2 carbamylation of amino acids on the enzyme backbone changes its conformation, reducing its activity. When adding TBA-Br, the formed carbamates could form electrostatic interactions with it, reducing the activity further. The underlying mechanism of inhibition needs further investigation. It is concluded that the application of elevated pressure on the enzymatic carboxylation of resorcinol is not an expedient solution at this stage.
Author Contributions
Conceptualization, D.O., B.T., P.B. and A.L.; Methodology, D.O. and B.T.; Validation, B.T.; Formal analysis, D.O.; Investigation, D.O.; Resources, A.L.; Writing–original draft, D.O.; Writing–review & editing, D.O., B.T., P.B. and A.L.; Visualization, D.O.; Supervision, P.B. and A.L.; Project administration, P.B. and A.L.; Funding acquisition, A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the German Research Foundation (DFG, grant number 322785574). Publishing fees were supported by Funding Programme Open Access Publishing of Hamburg University of Technology (TUHH).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and raw data is available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
2,6-DHBD, 2,6-dihydroxybenzoic acid decarboxylase from Rhizobium sp.; 2,6-DHBA, 2,6-dihydroxybenzoic acid; TEA, triethanolamine; DIC, dissolved inorganic carbon concentration; TBA-Br, tetrabutylammonium bromide; ‘CO2’, CO2 and H2CO3.
References
- Sato, M.; Sakurai, N.; Suzuki, H.; Shibata, D.; Kino, K. Enzymatic carboxylation of hydroxystilbenes by the γ-resorcylic acid decarboxylase from Rhizobium radiobacter WU-0108 under reverse reaction conditions. J. Mol. Catal. B Enzym. 2015, 122, 348–352. [Google Scholar] [CrossRef]
- Kirimura, K.; Ishii, Y. Enzymatic Kolbe–Schmitt Reaction for the Syntheses of Value-Added Compounds. In Future Directions in Biocatalysis, 2nd ed.; Matsuda, T., Ed.; Elsevier: Amsterdam, The Netherlands; Cambridge, MA, USA, 2017; pp. 135–147. ISBN 9780444637437. [Google Scholar]
- Payer, S.E.; Faber, K.; Glueck, S.M. Non-Oxidative Enzymatic (De)Carboxylation of (Hetero)Aromatics and Acrylic Acid Derivatives. Adv. Synth. Catal. 2019, 361, 2402–2420. [Google Scholar] [CrossRef] [PubMed]
- Aleku, G.A.; Roberts, G.W.; Titchiner, G.R.; Leys, D. Synthetic Enzyme-Catalyzed CO2 Fixation Reactions. ChemSusChem 2021, 14, 1781–1804. [Google Scholar] [CrossRef] [PubMed]
- Lide, D.R. CRC Handbook of Chemistry and Physics: A Ready-Reference Book of Chemical and Physical Data, 88th ed.; CRC: Boca Raton, FL, USA; London, UK, 2008; ISBN 0849304881. [Google Scholar]
- Pesci, L.; Glueck, S.M.; Gurikov, P.; Smirnova, I.; Faber, K.; Liese, A. Biocatalytic carboxylation of phenol derivatives: Kinetics and thermodynamics of the biological Kolbe-Schmitt synthesis: Kinetics and thermodynamics of the biological Kolbe-Schmitt synthesis. FEBS J. 2015, 282, 1334–1345. [Google Scholar] [CrossRef]
- Wuensch, C.; Schmidt, N.; Gross, J.; Grischek, B.; Glueck, S.M.; Faber, K. Pushing the equilibrium of regio-complementary carboxylation of phenols and hydroxystyrene derivatives. J. Biotechnol. 2013, 168, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Pesci, L.; Gurikov, P.; Liese, A.; Kara, S. Amine-Mediated Enzymatic Carboxylation of Phenols Using CO2 as Substrate Increases Equilibrium Conversions and Reaction Rates. Biotechnol. J. 2017, 12, 1700332. [Google Scholar] [CrossRef] [PubMed]
- Ohde, D.; Thomas, B.; Matthes, S.; Tanaka, S.; Bubenheim, P.; Terasaka, K.; Schlüter, M.; Liese, A. Microbubble enhanced mass transfer efficiency of CO2 capture utilizing aqueous triethanolamine for enzymatic resorcinol carboxylation. RSC Adv. 2021, 11, 4087–4096. [Google Scholar] [CrossRef]
- Ohde, D.; Thomas, B.; Bubenheim, P.; Liese, A. Enhanced CO2 fixation in the biocatalytic carboxylation of resorcinol: Utilization of amines for amine scrubbing and in situ product precipitation. Biochem. Eng. J. 2021, 166, 107825. [Google Scholar] [CrossRef]
- Fan, Y.; Feng, J.; Yang, M.; Tan, X.; Fan, H.; Guo, M.; Wang, B.; Xue, S. CO2 (aq) concentration–dependent CO2 fixation via carboxylation by decarboxylase. Green Chem. 2021, 23, 4403–4409. [Google Scholar] [CrossRef]
- Meyer, L.-E.; Plasch, K.; Kragl, U.; von Langermann, J. Adsorbent-Based Downstream-Processing of the Decarboxylase-Based Synthesis of 2,6-Dihydroxy-4-methylbenzoic Acid. Org. Process Res. Dev. 2018, 22, 963–970. [Google Scholar] [CrossRef]
- Pesci, L. Biocatalytic (De)carboxylation of Phenolic Compounds: Fundamentals and Applications. Ph.D. Dissertation, TUHH Universitätsbibliothek, Hamburg, Germany, 2017. [Google Scholar]
- Martin, J.; Eisoldt, L.; Skerra, A. Fixation of gaseous CO2 by reversing a decarboxylase for the biocatalytic synthesis of the essential amino acid l-methionine. Nat. Catal. 2018, 1, 555–561. [Google Scholar] [CrossRef]
- Ren, J.; Yao, P.; Yu, S.; Dong, W.; Chen, Q.; Feng, J.; Wu, Q.; Zhu, D. An Unprecedented Effective Enzymatic Carboxylation of Phenols. ACS Catal. 2016, 6, 564–567. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, J.; Yao, P.; Gong, R.; Wang, M.; Wu, Q.; Zhu, D. Biochemical characterization and substrate profiling of a reversible 2,3-dihydroxybenzoic acid decarboxylase for biocatalytic Kolbe-Schmitt reaction. Enzym. Microb. Technol. 2018, 113, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Jou, F.-Y.; Otto, F.D.; Mather, A.E. Equilibria of H2S and CO2 in triethanolamine solutions. Can. J. Chem. Eng. 1985, 63, 122–125. [Google Scholar] [CrossRef]
- Kent, R.L.; Elsenberg, B. Better Data for Amine Treating. Hydrocarb. Process. 1976, 55, 87–90. [Google Scholar]
- Yoshida, T.; Hayakawa, Y.; Matsui, T.; Nagasawa, T. Purification and characterization of 2,6-dihydroxybenzoate decarboxylase reversibly catalyzing nonoxidative decarboxylation. Arch. Microbiol. 2004, 181, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Kilmartin, J.V.; Rossi-Bernardi, L. Inhibition of CO2 combination and reduction of the Bohr effect in haemoglobin chemically modified at its alpha-amino groups. Nature 1969, 222, 1243–1246. [Google Scholar] [CrossRef] [PubMed]
- Kilmartin, J.V.; Rossi-Bernardi, L. The binding of carbon dioxide by horse haemoglobin. Biochem. J. 1971, 124, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Kilmartin, J.V.; Rossi-Bernardi, L. Interaction of hemoglobin with hydrogen ions, carbon dioxide, and organic phosphates. Physiol. Rev. 1973, 53, 836–890. [Google Scholar] [CrossRef]
- Andrews, T.J.; Badger, M.R.; Lorimer, G.H. Factors affecting interconversion between kinetic forms of ribulose diphosphate carboxylase-oxygenase from spinach. Arch. Biochem. Biophys. 1975, 171, 93–103. [Google Scholar] [CrossRef]
- Cleland, W.W.; Andrews, T.J.; Gutteridge, S.; Hartman, F.C.; Lorimer, G.H. Mechanism of Rubisco: The Carbamate as General Base. Chem. Rev. 1998, 98, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Lorimer, G.H.; Badger, M.R.; Andrews, T.J. The activation of ribulose-1,5-bisphosphate carboxylase by carbon dioxide and magnesium ions. Equilibria, kinetics, a suggested mechanism, and physiological implications. Biochemistry 1976, 15, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Lorimer, G.H.; Miziorko, H.M. Carbamate formation on the epsilon-amino group of a lysyl residue as the basis for the activation of ribulosebisphosphate carboxylase by CO2 and Mg2+. Biochemistry 1980, 19, 5321–5328. [Google Scholar] [CrossRef] [PubMed]
- Blake, L.I.; Cann, M.J. Carbon Dioxide and the Carbamate Post-Translational Modification. Front. Mol. Biosci. 2022, 9, 825706. [Google Scholar] [CrossRef]
- Linthwaite, V.L.; Janus, J.M.; Brown, A.P.; Wong-Pascua, D.; O’Donoghue, A.C.; Porter, A.; Treumann, A.; Hodgson, D.R.W.; Cann, M.J. The identification of carbon dioxide mediated protein post-translational modifications. Nat. Commun. 2018, 9, 3092. [Google Scholar] [CrossRef]
- Wimmer, Z.; Zarevúcka, M. A review on the effects of supercritical carbon dioxide on enzyme activity. Int. J. Mol. Sci. 2010, 11, 233–253. [Google Scholar] [CrossRef]
- Wang, W.; Rao, L.; Wu, X.; Wang, Y.; Zhao, L.; Liao, X. Supercritical Carbon Dioxide Applications in Food Processing. Food Eng. Rev. 2021, 13, 570–591. [Google Scholar] [CrossRef]
- Silva, E.K.; Meireles, M.A.A.; Saldaña, M.D. Supercritical carbon dioxide technology: A promising technique for the non-thermal processing of freshly fruit and vegetable juices. Trends Food Sci. Technol. 2020, 97, 381–390. [Google Scholar] [CrossRef]
- Balaban, M.O.; Arreola, A.G.; Marshall, M.; Peplow, A.; Wei, C.I.; Cornell, J. Inactivation of Pectinesterase in Orange Juice by Supercritical Carbon Dioxide. J. Food Sci. 1991, 56, 743–746. [Google Scholar] [CrossRef]
- Park, S.-J.; Lee, J.-I.; Park, J. Effects of a Combined Process of High-Pressure Carbon Dioxide and High Hydrostatic Pressure on the Quality of Carrot Juice. J. Food Sci. 2002, 67, 1827–1834. [Google Scholar] [CrossRef]
- Habulin, M.; Knez, Ž. Activity and stability of lipases from different sources in supercritical carbon dioxide and near-critical propane. J. Chem. Technol. Biotechnol. 2001, 76, 1260–1266. [Google Scholar] [CrossRef]
- Perumal, M.; Jayaraman, D.; Balraj, A. Experimental studies on CO2 absorption and solvent recovery in aqueous blends of monoethanolamine and tetrabutylammonium hydroxide. Chemosphere 2021, 276, 130159. [Google Scholar] [CrossRef] [PubMed]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ohde, D.; Thomas, B.; Matthes, S.; Percin, Z.; Engelmann, C.; Bubenheim, P.; Terasaka, K.; Schlüter, M.; Liese, A. Fine Bubble-based CO2 Capture Mediated by Triethanolamine Coupled to Whole Cell Biotransformation. Chem. Ing. Tech. 2019, 91, 1822–1826. [Google Scholar] [CrossRef]
Figure 1.
Enzymatic carboxylation of resorcinol to 2,6-dihydroxybenzoic acid (2,6-DHBA) by 2,6-DHBA decarboxylase (2,6-DHBD) in aqueous triethanolamine (TEA) saturated with CO2.
Figure 1.
Enzymatic carboxylation of resorcinol to 2,6-dihydroxybenzoic acid (2,6-DHBA) by 2,6-DHBA decarboxylase (2,6-DHBD) in aqueous triethanolamine (TEA) saturated with CO2.
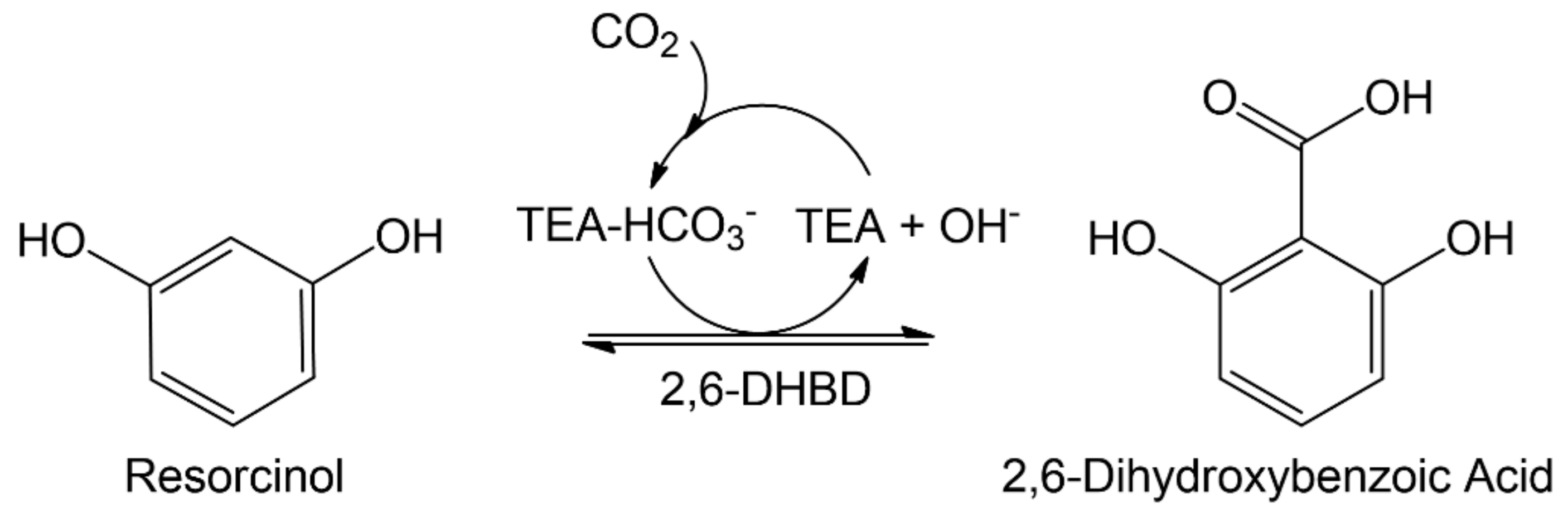
Figure 2.
Theoretical equilibrium yield (lines) and experimentally determined reaction yield (symbols) for the enzymatic carboxylation of 80 mM resorcinol to 2,6-dihydroxybenzoic acid up to 1200 kPa CO2 in 0.14 M to 3 M aqueous triethanolamine. Samples were incubated for 2 days and 9 days. Only the 9-day samples of 2 and 3 M aqueous TEA are shown as their yield still increased.
Figure 2.
Theoretical equilibrium yield (lines) and experimentally determined reaction yield (symbols) for the enzymatic carboxylation of 80 mM resorcinol to 2,6-dihydroxybenzoic acid up to 1200 kPa CO2 in 0.14 M to 3 M aqueous triethanolamine. Samples were incubated for 2 days and 9 days. Only the 9-day samples of 2 and 3 M aqueous TEA are shown as their yield still increased.

Figure 3.
Calculated pH profiles for different concentrations of aqueous triethanolamine (TEA) solutions at 25 °C and CO2 pressures up to 1200 kPa. The calculation was performed according to Jou et al. (1985) [17].
Figure 3.
Calculated pH profiles for different concentrations of aqueous triethanolamine (TEA) solutions at 25 °C and CO2 pressures up to 1200 kPa. The calculation was performed according to Jou et al. (1985) [17].

Figure 4.
Dissolved concentration of CO2/H2CO3, HCO3−, and CO32− in 1 M aqueous triethanolamine up to 1200 kPa at 25 °C.
Figure 4.
Dissolved concentration of CO2/H2CO3, HCO3−, and CO32− in 1 M aqueous triethanolamine up to 1200 kPa at 25 °C.

Figure 5.
Reaction yield for the enzymatic carboxylation of 80 mM resorcinol to 2,6-dihydroxybenzoic acid with 350 mM tetrabutylammonium bromide under elevated CO2 pressure in aqueous triethanolamine (TEA) after 2 days.
Figure 5.
Reaction yield for the enzymatic carboxylation of 80 mM resorcinol to 2,6-dihydroxybenzoic acid with 350 mM tetrabutylammonium bromide under elevated CO2 pressure in aqueous triethanolamine (TEA) after 2 days.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ohde, D.; Thomas, B.; Bubenheim, P.; Liese, A. Enzymatic Carboxylation of Resorcinol in Aqueous Triethanolamine at Elevated CO2 Pressure. Molecules 2024, 29, 25. https://doi.org/10.3390/molecules29010025
AMA Style
Ohde D, Thomas B, Bubenheim P, Liese A. Enzymatic Carboxylation of Resorcinol in Aqueous Triethanolamine at Elevated CO2 Pressure. Molecules. 2024; 29(1):25. https://doi.org/10.3390/molecules29010025
Chicago/Turabian StyleOhde, Daniel, Benjamin Thomas, Paul Bubenheim, and Andreas Liese. 2024. "Enzymatic Carboxylation of Resorcinol in Aqueous Triethanolamine at Elevated CO2 Pressure" Molecules 29, no. 1: 25. https://doi.org/10.3390/molecules29010025