
Beneficial Effects of Dietary Fiber in Young Barley Leaf on Gut Microbiota and Immunity in Mice
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. YB Modifies cDC Function in the PP
2.2. YB Activates Bone Marrow-Derived DCs (BMDCs)
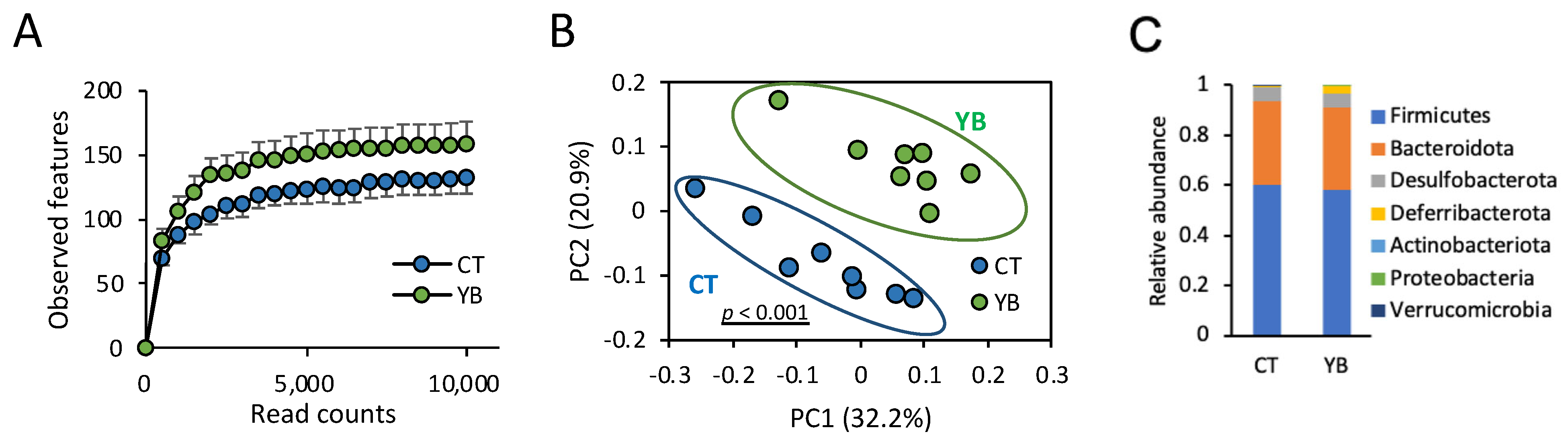
2.3. YB Modifies the Gut Microbiota
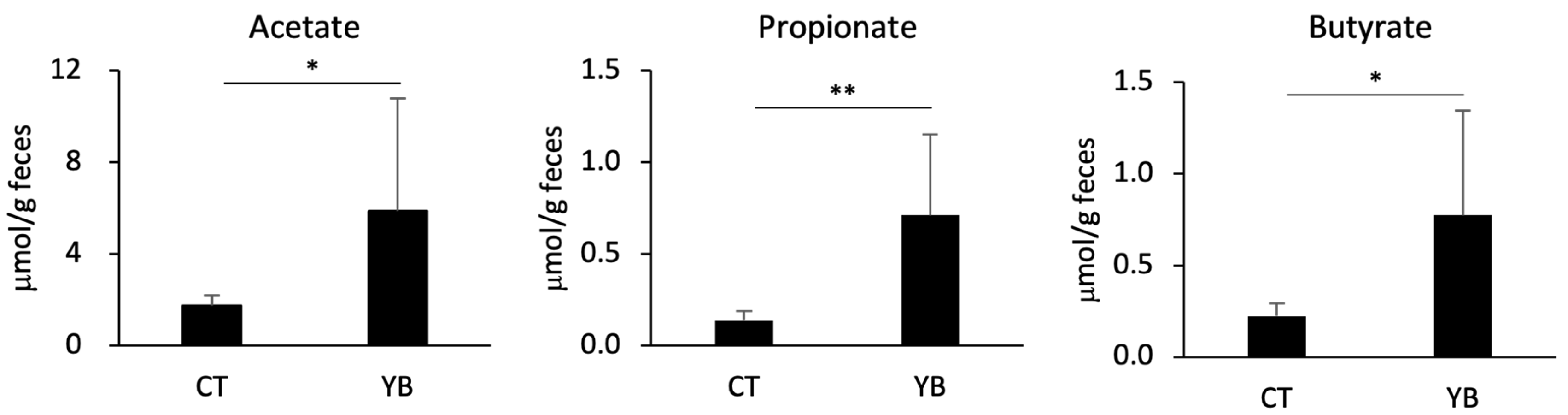
2.4. YB Augments Fecal SCFAs
2.5. YB Promotes RALDH Activity in Mesenteric Lymph Node (MLN) cDCs
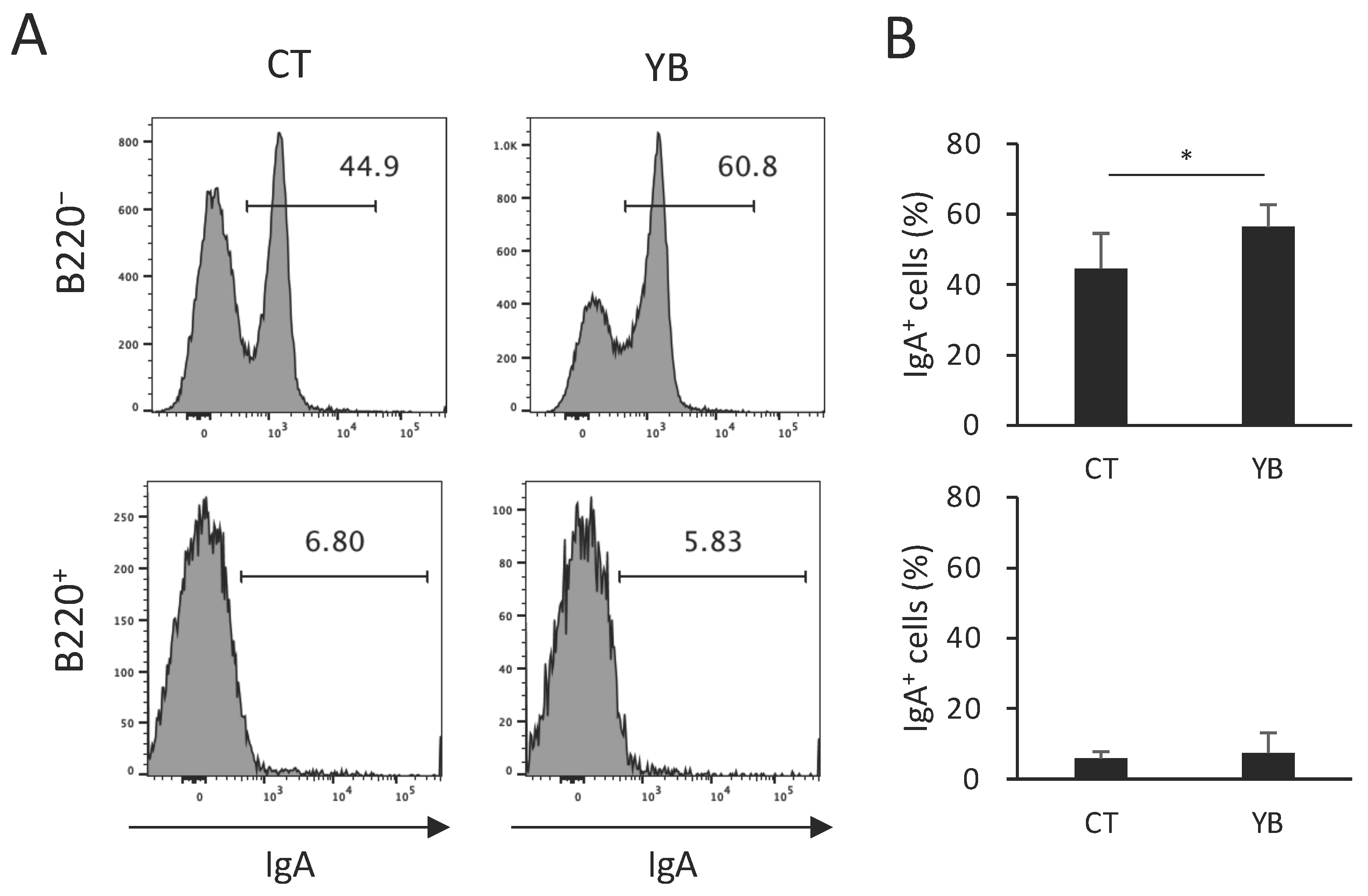
2.6. YB Augments IgA+ Plasma Cells in Colonic Lamina Propria
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of Cells from the PP, MLN, and Colonic Lamina Propria
4.3. Generation of BMDCs and Treatment
4.4. Flow Cytometry
4.5. Cytokine Measurement
4.6. Co-Culture of BMDCs with Naïve CD4+ T Cells
4.7. Isolation of Fecal DNA
4.8. 16S rRNA Sequencing
4.9. Gas Chromatography–Mass Spectrometry (GC/MS) Analysis of Fecal SCFAs
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14 (Suppl. S2), 49. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, H.; Ohteki, T. Regulation of IgA Production by Intestinal Dendritic Cells and Related Cells. Front. Immunol. 2019, 10, 1891. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Nguyen, A.; Gommerman, J.L. Dendritic Cell Subsets in Intestinal Immunity and Inflammation. J. Immunol. 2020, 204, 1075–1083. [Google Scholar] [CrossRef]
- Bos, A.; van Egmond, M.; Mebius, R. The role of retinoic acid in the production of immunoglobulin A. Mucosal Immunol. 2022, 15, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Chudan, S.; Ishibashi, R.; Nishikawa, M.; Tabuchi, Y.; Nagai, Y.; Ikushiro, S.; Furusawa, Y. Effect of Wheat-Derived Arabinoxylan on the Gut Microbiota Composition and Colonic Regulatory T Cells. Molecules 2023, 28, 3079. [Google Scholar] [CrossRef] [PubMed]
- Chudan, S.; Ishibashi, R.; Nishikawa, M.; Tabuchi, Y.; Nagai, Y.; Ikushiro, S.; Furusawa, Y. Effect of soluble oat fiber on intestinal microenvironment and TNBS-induced colitis. Food Funct. 2023, 14, 2188–2199. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.M.; Wu, C.H.; Tseng, Y.H.; Tsai, C.M.E.; Chang, W.C. Antioxidative and hypolipidemic effects of barley leaf essence in a rabbit model of atherosclerosis. Jpn. J. Pharmacol. 2002, 89, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Takano, A.; Kamiya, T.; Tomozawa, H.; Ueno, S.; Tsubata, M.; Ikeguchi, M.; Takagaki, K.; Okushima, A.; Miyata, Y.; Tamaru, S.; et al. Insoluble fiber in young barley leaf suppresses the increment of postprandial blood glucose level by increasing the digesta viscosity. Evid. Based Complement. Altern. Med. 2013, 2013, 137871. [Google Scholar] [CrossRef]
- Jones, J.M. CODEX-aligned dietary fiber definitions help to bridge the ‘fiber gap’. Nutr. J. 2014, 13, 34. [Google Scholar] [CrossRef]
- Li, M.Z.; Huang, X.J.; Wen, J.J.; Chen, S.K.; Wu, X.C.; Ma, W.N.; Cui, S.W.; Xie, M.Y.; Nie, S.P. Innate immune receptors co-recognition of polysaccharides initiates multi-pathway synergistic immune response. Carbohydr. Polym. 2023, 305, 120533. [Google Scholar] [CrossRef] [PubMed]
- Mata-Martinez, P.; Bergon-Gutierrez, M.; Del Fresno, C. Dectin-1 Signaling Update: New Perspectives for Trained Immunity. Front. Immunol. 2022, 13, 812148. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, N.M.; Schols, H.A.; Faas, M.M.; de Vos, P. Arabinoxylan activates Dectin-1 and modulates particulate β-glucan-induced Dectin-1 activation. Mol. Nutr. Food Res. 2016, 60, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Sinkovic, L.; Rakszegi, M.; Pipan, B.; Meglic, V. Compositional Traits of Grains and Groats of Barley, Oat and Spelt Grown at Organic and Conventional Fields. Foods 2023, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Bravo Nunez, A.; Sahin, A.W.; Arendt, E.K. Arabinoxylans as Functional Food Ingredients: A Review. Foods 2022, 11, 1026. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Govers, C.; Wichers, H.J.; Mes, J.J. Macrophages treated with non-digestible polysaccharides reveal a transcriptionally unique phenotype. J. Funct. Foods 2017, 36, 280–289. [Google Scholar] [CrossRef]
- Requena, T.; Martinez-Cuesta, M.C.; Pelaez, C. Diet and microbiota linked in health and disease. Food Funct. 2018, 9, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Isobe, J.; Maeda, S.; Obata, Y.; Iizuka, K.; Nakamura, Y.; Fujimura, Y.; Kimizuka, T.; Hattori, K.; Kim, Y.G.; Morita, T.; et al. Commensal-bacteria-derived butyrate promotes the T-cell-independent IgA response in the colon. Int. Immunol. 2020, 32, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, D.; Sasaki, K.; Kadowaki, Y.; Aotsuka, Y.; Kondo, A. Bifidogenic and butyrogenic effects of young barely leaf extract in an in vitro human colonic microbiota model. AMB Express 2019, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Hase, K.; Kawano, K.; Nochi, T.; Pontes, G.S.; Fukuda, S.; Ebisawa, M.; Kadokura, K.; Tobe, T.; Fujimura, Y.; Kawano, S.; et al. Uptake through glycoprotein 2 of FimH+ bacteria by M cells initiates mucosal immune response. Nature 2009, 462, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef]
- Moens, F.; De Vuyst, L. Inulin-type fructan degradation capacity of Clostridium cluster IV and XIVa butyrate-producing colon bacteria and their associated metabolic outcomes. Benef. Microbes 2017, 8, 473–490. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.S.; Wanapat, M.; Hou, F.J. Rumen bacteria influence milk protein yield of yak grazing on the Qinghai-Tibet plateau. Anim. Biosci. 2021, 34, 1466–1478. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Russler-Germain, E.V.; Yi, J.; Young, S.; Nutsch, K.; Wong, H.S.; Ai, T.L.; Chai, J.N.; Durai, V.; Kaplan, D.H.; Germain, R.N.; et al. Gut Helicobacter presentation by multiple dendritic cell subsets enables context-specific regulatory T cell generation. Elife 2021, 10, e54792. [Google Scholar] [CrossRef] [PubMed]
- Ruane, D.T.; Lavelle, E.C. The role of CD103(+) dendritic cells in the intestinal mucosal immune system. Front. Immunol. 2011, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Evaldson, G.; Heimdahl, A.; Kager, L.; Nord, C.E. The normal human anaerobic microflora. Scand. J. Infect. Dis. Suppl. 1982, 35, 9–15. [Google Scholar] [PubMed]
- Bures, J.; Cyrany, J.; Kohoutova, D.; Forstl, M.; Rejchrt, S.; Kvetina, J.; Vorisek, V.; Kopacova, M. Small intestinal bacterial overgrowth syndrome. World J. Gastroenterol. 2010, 16, 2978–2990. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Fujii, G.; Sakurai, H.; Yoshitome, H.; Mutoh, M.; Wada, M. Intestinal Peyer’s patches prevent tumorigenesis in mice. J. Clin. Biochem. Nutr. 2015, 56, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Hussein, S.M.; Yokhana, J.S.; Frankel, T.L. Supplementing the feeds of layer pullets, at different ages with two different fiber sources improves immune function. Poultry Sci. 2017, 96, 2718–2727. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, X.; Wu, Y.; Zhou, L.; Wang, Z.; Xiao, W. Effect of Lentinan on Peyer’s patch structure and function in an immunosuppressed mouse model. Int. J. Biol. Macromol. 2019, 137, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, M.H.; Matsumoto, T.; Kiyohara, H.; Yamada, H. Detection and tissue distribution of anti-ulcer pectic polysaccharides from Bupleurum falcatum by polyclonal antibody. Planta Med. 1996, 62, 341–346. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, M.; Ostroff, G.R.; Levitz, S.M.; Bartling, T.R.; Mantis, N.J. A population of Langerin-positive dendritic cells in murine Peyer’s patches involved in sampling beta-glucan microparticles. PLoS ONE 2014, 9, e91002. [Google Scholar] [CrossRef] [PubMed]
- Jowett, G.M.; Read, E.; Roberts, L.B.; Coman, D.; Vila Gonzalez, M.; Zabinski, T.; Niazi, U.; Reis, R.; Trieu, T.J.; Danovi, D.; et al. Organoids capture tissue-specific innate lymphoid cell development in mice and humans. Cell Rep. 2022, 40, 111281. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Upchurch, K.; Zhang, W.; Ni, L.; Li, D.; Xue, Y.; Li, X.H.; Hori, T.; Zurawski, S.; Liu, Y.J.; et al. Opposing Roles of Dectin-1 Expressed on Human Plasmacytoid Dendritic Cells and Myeloid Dendritic Cells in Th2 Polarization. J. Immunol. 2015, 195, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Pupovac, A.; Evtimov, V.; Boyd, N.; Shu, R.; Boyd, R.; Trounson, A. Enhancing a Natural Killer: Modification of NK Cells for Cancer Immunotherapy. Cells 2021, 10, 1058. [Google Scholar] [CrossRef]
- Molino, S.; Lerma-Aguilera, A.; Jimenez-Hernandez, N.; Rufian Henares, J.A.; Francino, M.P. Evaluation of the Effects of a Short Supplementation With Tannins on the Gut Microbiota of Healthy Subjects. Front. Microbiol. 2022, 13, 848611. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Yu, K.W.; Hong, H.D.; Shin, K.S. Effect of arabinoxylan- and rhamnogalacturonan I-rich polysaccharides isolated from young barley leaf on intestinal immunostimulatory activity. J. Funct. Foods 2017, 35, 384–390. [Google Scholar] [CrossRef]
- Duncan, S.H.; Russell, W.R.; Quartieri, A.; Rossi, M.; Parkhill, J.; Walker, A.W.; Flint, H.J. Wheat bran promotes enrichment within the human colonic microbiota of butyrate-producing bacteria that release ferulic acid. Environ. Microbiol. 2016, 18, 2214–2225. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef] [PubMed]
- Vital, M.; Howe, A.C.; Tiedje, J.M. Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. mBio 2014, 5, e00889. [Google Scholar] [CrossRef] [PubMed]
- Whibley, N.; Tucci, A.; Powrie, F. Regulatory T cell adaptation in the intestine and skin. Nat. Immunol. 2019, 20, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Furusawa, Y.; Endo, T.A.; Sharif, J.; Takahashi, D.; Atarashi, K.; Nakayama, M.; Onawa, S.; Fujimura, Y.; Takahashi, M.; et al. The epigenetic regulator Uhrf1 facilitates the proliferation and maturation of colonic regulatory T cells. Nat. Immunol. 2014, 15, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Yamanouchi, Y.; Chudan, S.; Ishibashi, R.; Ohue-Kitano, R.; Nishikawa, M.; Tabuchi, Y.; Kimura, I.; Nagai, Y.; Ikushiro, S.; Furusawa, Y. The Impact of Low Viscosity Soluble Dietary Fibers on Intestinal Microenvironment and Experimental Colitis: A Possible Preventive Application of Alpha-Cyclodextrin in Intestinal Inflammation. Mol. Nutr. Food Res. 2022, 66, e2200063. [Google Scholar] [CrossRef] [PubMed]
- Ikeguchi, M.; Tsubata, M.; Takano, A.; Kamiya, T.; Takagaki, K.; Ito, H.; Sugawa-Katayama, Y.; Tsuji, H. Effects of young barley leaf powder on gastrointestinal functions in rats and its efficacy-related physicochemical properties. Evid. Based Complement. Altern. Med. 2014, 2014, 974840. [Google Scholar] [CrossRef] [PubMed]
- Gorzelak, M.A.; Gill, S.K.; Tasnim, N.; Ahmadi-Vand, Z.; Jay, M.; Gibson, D.L. Methods for Improving Human Gut Microbiome Data by Reducing Variability through Sample Processing and Storage of Stool. PLoS ONE 2015, 10, e0134802. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, R.; Furusawa, Y.; Honda, H.; Watanabe, Y.; Fujisaka, S.; Nishikawa, M.; Ikushiro, S.; Kurihara, S.; Tabuchi, Y.; Tobe, K.; et al. Isoliquiritigenin Attenuates Adipose Tissue Inflammation and Metabolic Syndrome by Modifying Gut Bacteria Composition in Mice. Mol. Nutr. Food Res. 2022, 66, e2101119. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, R.; Matsuhisa, R.; Nomoto, M.; Chudan, S.; Nishikawa, M.; Tabuchi, Y.; Ikushiro, S.; Nagai, Y.; Furusawa, Y. Effect of Oral Administration of Polyethylene Glycol 400 on Gut Microbiota Composition and Diet-Induced Obesity in Mice. Microorganisms 2023, 11, 1882. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Nanjo, F. Dietary cycloinulooligosaccharides enhance intestinal immunoglobulin A production in mice. Biosci. Biotechnol. Biochem. 2009, 73, 677–682. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chudan, S.; Kurakawa, T.; Nishikawa, M.; Nagai, Y.; Tabuchi, Y.; Ikushiro, S.; Furusawa, Y. Beneficial Effects of Dietary Fiber in Young Barley Leaf on Gut Microbiota and Immunity in Mice. Molecules 2024, 29, 1897. https://doi.org/10.3390/molecules29081897
Chudan S, Kurakawa T, Nishikawa M, Nagai Y, Tabuchi Y, Ikushiro S, Furusawa Y. Beneficial Effects of Dietary Fiber in Young Barley Leaf on Gut Microbiota and Immunity in Mice. Molecules. 2024; 29(8):1897. https://doi.org/10.3390/molecules29081897
Chicago/Turabian StyleChudan, Seita, Takuto Kurakawa, Miyu Nishikawa, Yoshinori Nagai, Yoshiaki Tabuchi, Shinichi Ikushiro, and Yukihiro Furusawa. 2024. "Beneficial Effects of Dietary Fiber in Young Barley Leaf on Gut Microbiota and Immunity in Mice" Molecules 29, no. 8: 1897. https://doi.org/10.3390/molecules29081897