Oxidative Neurodegeneration Is Prevented by UCP0045037, an Allosteric Modulator for the Reduced Form of DJ-1, a Wild-Type of Familial Parkinson’s Disease-Linked PARK7

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
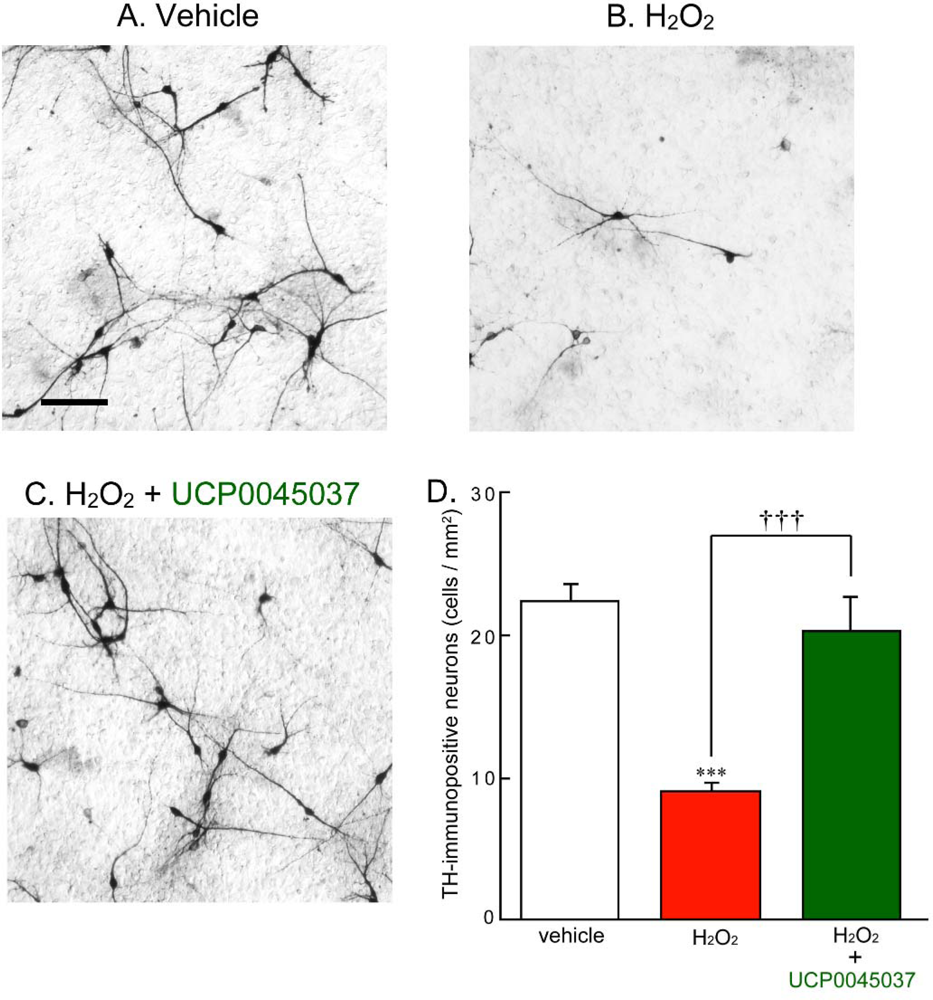
2.1. Effect of DJ-1 Ligand in the in Vivo Rat Brain
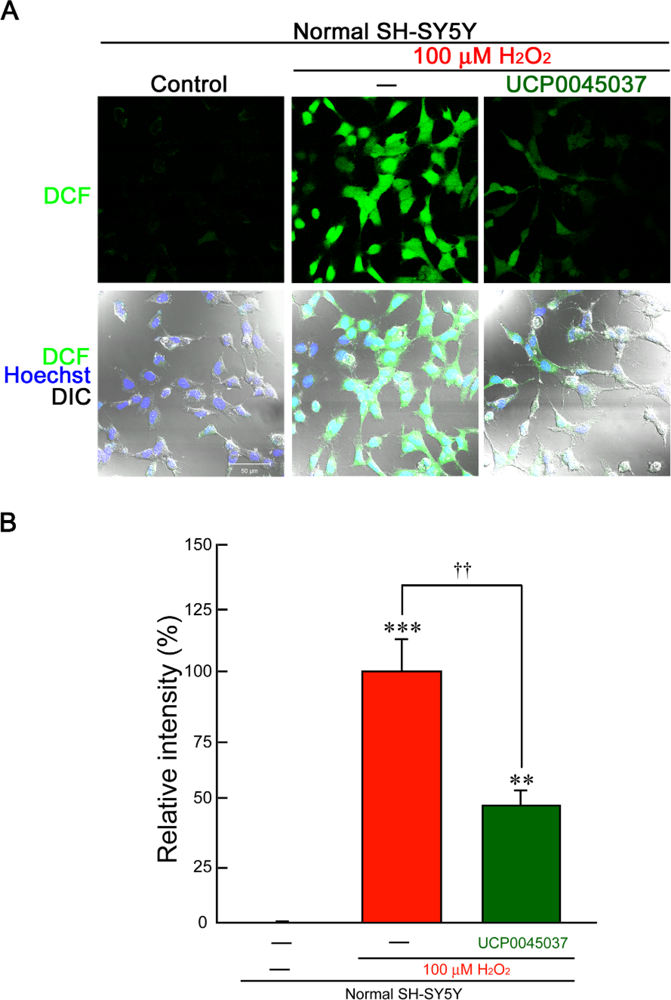
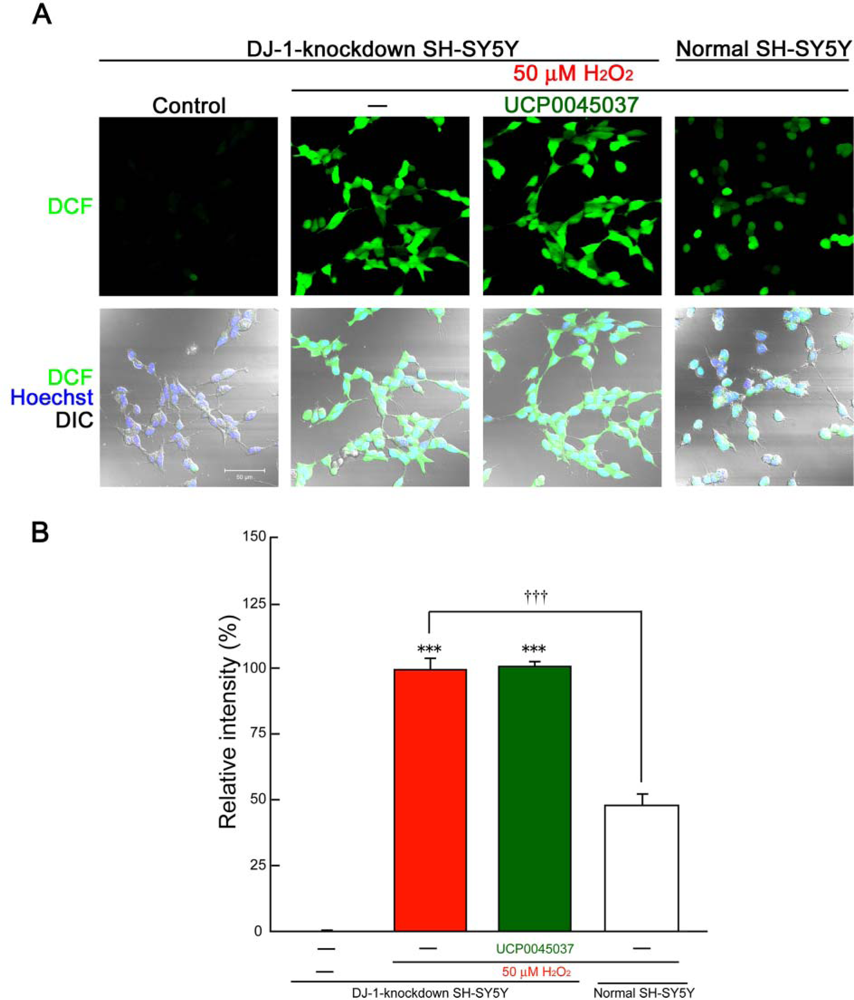
2.2. Effect of DJ-1 Ligand in the in Vitro Neuronal Cultures
3. Discussion
4. Experimental Section
4.1. In Vivo Model of Rat Focal Cerebral Ischemia
4.2. Intrastriatal Drug Administration to Ischemic Rats
4.3. Measurement of Infarct Volume in Rat Ischemic Brain
4.4. Primary Neuronal Culture of Rat Ventral Mesencephalon
4.5. In Vitro Cell Culture and Establishment of DJ-1-Knockdown Cells
4.6. Western Blotting Analysis
4.7. Assay of ROS Production
4.8. Measurement of Cell Viability
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
References and Notes
- Nagakubo, D; Taira, T; Kitaura, H; Ikeda, M; Tamai, K; Iguchi-Ariga, SMM; Ariga, H. DJ-1, a novel oncogene which transforms mouse NIH3T3 cells in cooperation with ras. Biochem. Biophys. Res. Commum 1997, 231, 509–513. [Google Scholar]
- Bonifati, V; Rizzu, P; van Baren, MJ; Schaap, O; Breedveld, GJ; Krieger, E; Dekker, MC; Squitieri, F; Ibanez, P; Josse, M; van Dongen, JW; Vanacore, N; van Swieten, JC; Brice, A; Meco, G; van Duijin, CM; Oostra, BA; Heutink, P. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar]
- Niki, T; Takahashi-Niki, K; Taira, T; Iguchi-Ariga, SMM; Ariga, H. DJBP: A novel DJ-1-binding protein, negatively regulates the androgen receptor by recruiting histone deacetylase complex, and DJ-1 antagonizes this inhibition by abrogation of this complex. Mol. Cancer Res 2003, 1, 247–261. [Google Scholar]
- Shendelman, S; Jonason, A; Martinat, C; Leete, T; Abeliovich, A. DJ-1 is a redox-dependent molecular chaperone that inhibits alpha-synuclein aggregate formation. PLoS. Biol 2004, 2, 1764–1773. [Google Scholar]
- Shinbo, Y; Taira, T; Niki, T; Iguchi-Ariga, SMM; Ariga, H. DJ-1 restores p53 transcription activity inhibited by Topors/p53BP3. Int. J. Oncol 2005, 26, 641–648. [Google Scholar]
- Takahashi, K; Taira, T; Niki, T; Seino, C; Iguchi-Ariga, SMM; Ariga, H. DJ-1 positively regulates the androgen receptor by impairing the binding of PIASx alpha to the receptor. J. Biol. Chem 2001, 276, 37556–37563. [Google Scholar]
- Honbou, K; Suzuki, NN; Horiuchi, M; Niki, T; Taira, T; Ariga, H; Inagaki, F. The crystal structure of DJ-1, a protein related to male fertility and Parkinson’s disease. J. Biol. Chem 2003, 278, 31380–31384. [Google Scholar]
- Huai, Q; Sun, Y; Wang, H; Chin, LS; Li, L; Robinson, H; Ke, H. Crystal structure of DJ-1/RS and implication on familial Parkinson’s disease. FEBS Lett 2003, 549, 171–175. [Google Scholar]
- Inden, M; Taira, T; Kitamura, Y; Yanagida, T; Tsuchiya, D; Takata, K; Yanagisawa, D; Nishimura, K; Taniguchi, T; Kiso, Y; Yoshimoto, K; Agatsuma, T; Koide-Yoshida, S; Iguchi-Ariga, SMM; Shimohama, S; Ariga, H. PARK7 DJ-1 protects against degeneration of nigral dopaminergic neurons in Parkinson’s disease rat model. Neurobiol. Dis 2006, 24, 144–158. [Google Scholar]
- Tao, X; Tong, L. Crystal structure of human DJ-1, a protein associated with early onset Parkinson’s disease. J. Biol. Chem 2003, 278, 31372–31379. [Google Scholar]
- Wilson, MA; Collins, JL; Hod, Y; Ringe, D; Petsko, GA. The 1.1-A resolution crystal structure of DJ-1, the protein mutated in autosomal recessive early onset Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2003, 100, 9256–9261. [Google Scholar]
- Kinumi, T; Kimata, J; Taira, T; Ariga, H; Niki, E. Cysteine-106 of DJ-1 is the most sensitive cysteine residue to hydrogen peroxide-mediated oxidation in vivo in human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun 2004, 317, 722–728. [Google Scholar]
- Taira, T; Saito, Y; Niki, T; Iguchi-Ariga, SMM; Takahashi, K; Ariga, H. DJ-1 has a role in antioxidative stress to prevent cell death. EMBO Rep 2004, 5, 213–218. [Google Scholar]
- Yanagida, T; Takata, K; Inden, M; Kitamura, Y; Taniguchi, T; Yoshimoto, K; Taira, T; Ariga, H. Distribution of DJ-1, Parkinson’s disease-related protein PARK7, and its alteration in 6-hydroxydopamine-treated hemiparkinsonian rat brain. J. Pharmacol. Sci 2006, 102, 243–247. [Google Scholar]
- Yanagisawa, D; Kitamura, Y; Inden, M; Takata, K; Taniguchi, T; Morikawa, S; Morita, M; Inubushi, T; Tooyama, I; Taira, T; Iguchi-Ariga, SMM; Akaike, A; Ariga, H. DJ-1 protects against neurodegeneration caused by focal cerebral ischemia and reperfusion in rats. J. Cereb. Blood Flow Metab 2008, 28, 563–578. [Google Scholar]
- Miyazaki, S; Yanagida, T; Nunome, K; Ishikawa, S; Inden, M; Kitamura, Y; Nakagawa, S; Taira, T; Hirota, K; Niwa, M; Iguchi-Ariga, SMM; Ariga, H. DJ-1-binding compounds prevent oxidative stress-induced cell death and movement defect in Parkinson’s disease model rats. J. Neurochem 2008, 105, 2418–2434. [Google Scholar]
- Hata, R; Maeda, K; Hermann, D; Mies, G; Hossmann, KA. Dynamics of regional brain metabolism and gene expression after middle cerebral artery occlusion in mice. J. Cereb. Blood Flow Metab 2000, 20, 306–315. [Google Scholar]
- Peters, O; Back, T; Lindauer, U; Busch, C; Megow, D; Dreier, J; Dirnagi, U. Increased formation of reactive oxygen species after permanent and reversible middle cerebral artery occlusion in the rat. J. Cereb. Blood Flow Metab 1998, 18, 196–205. [Google Scholar]
- Andersen, JK. Oxidative stress in neurodegeneration: cause or consequence? Nat. Med 2004, 10, S18–S25. [Google Scholar]
- Margaill, I; Plotkine, M; Lerouet, D. Antioxidant strategies in the treatment of stroke. Free Radic. Biol. Med 2005, 39, 429–443. [Google Scholar]
- Yanagisawa, D; Qi, M; Kim, D-H; Kitamura, Y; Inden, M; Tsuchiya, D; Takata, K; Taniguchi, T; Yoshimoto, K; Shimohama, S; Akaike, A; Sumi, S; Inoue, K. Improvement of focal ischemia-induced rat dopaminergic dysfunction by striatal transplantation of mouse embryonic stem cells. Neurosci. Lett 2006, 407, 74–79. [Google Scholar]
- Kitamura, Y; Kosaka, T; Kakimura, J-I; Matsuoka, Y; Kohno, Y; Nomura, Y; Taniguchi, T. Protective effects of the antiparkinsonian drugs talipexole and pramipexole against 1-methyl-4-phenylpyridinium-induced apoptotic death in human neuroblastoma SH-SY5Y cells. Mol. Pharmacol 1998, 54, 1046–1054. [Google Scholar]
- Sawada, H; Kohno, R; Kihara, T; Izumi, Y; Sakka, N; Ibi, M; Nakanishi, M; Nakamizo, T; Yamakawa, K; Shibasaki, H; Yamamoto, N; Akaike, A; Inden, M; Kitamura, Y; Taniguchi, T; Shimohama, S. Proteasome mediates dopaminergic neuronal degeneration, and its inhibition causes alpha-synuclein inclusions. J. Biol. Chem 2004, 279, 10710–10719. [Google Scholar]
- Zhou, W; Zhu, M; Wilson, MA; Petsko, GA; Fink, AL. The oxidation state of DJ-1 regulates its chaperone activity toward alpha-synuclein. J. Mol. Biol 2006, 356, 1036–1048. [Google Scholar]
- Allard, L; Burkhard, PR; Lescuyer, P; Burgess, JA; Walter, N; Hochstrasser, DF; Sanchez, JC. PARK7 and nucleoside diphosphate kinase A as plasma markers for the early diagnosis of stroke. Clin. Chem 2005, 51, 2043–2051. [Google Scholar]
- Waragai, M; Wei, J; Fujita, M; Nakai, M; Ho, GJ; Masliah, E; Akatsu, H; Yamada, T; Hashimoto, M. Increased level of DJ-1 in the cerebrospinal fluids of sporadic Parkinson’s disease. Biochem. Biophys. Res. Commun 2006, 345, 967–972. [Google Scholar]
- Maita, C; Tsuji, S; Yabe, I; Hamada, S; Ogata, A; Maita, H; Iguchi-Ariga, SMM; Sasaki, H; Ariga, H. Secretion of DJ-1 into the serum of patients with Parkinson’s disease. Neurosci. Lett 2008, 431, 86–89. [Google Scholar]
- Hirotani, M; Maita, C; Niino, M; Iguchi-Ariga, SMM; Hamada, S; Ariga, H; Sasaki, H. Correlation between DJ-1 levels in the cerebrospainal fluid and the progression of disabilities in multiple sclerosis patients. Multiple Sclerosis 2008, 14, 1056–1060. [Google Scholar]
- Clements, CM; McNally, RS; Conti, BJ; Mak, TW; Ting, JP. DJ-1, a cancer-and Parkinson’s disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc. Natl. Acad. Sci. USA 2006, 103, 15091–15096. [Google Scholar]
- Xu, J; Zhong, N; Wang, H; Elias, JE; Kim, CY; Woldman, I; Pifl, C; Gygi, SP; Geula, C; Yankner, BA. The Parkinson’s disease-associated DJ-1 protein is a transcriptional co-activator that protects against neuronal apoptosis. Hum. Mol. Genet 2005, 14, 1231–1241. [Google Scholar]
- Junn, E; Taniguchi, H; Jeong, BS; Zhao, X; Ichijo, H; Mouradian, MM. Interaction of DJ-1 with Daxx inhibits apoptosis signal-regulating kinase 1 activity and cell death. Proc. Natl. Acad. Sci. USA 2005, 102, 9691–9696. [Google Scholar]
- Sekito, A; Koide-Yoshida, S; Niki, T; Taira, T; Iguchi-Ariga, SM; Ariga, H. DJ-1 interacts with HIPK1 and affects H2O2-induced cell death. Free Radic. Res 2006, 40, 155–165. [Google Scholar]
- Kitamura, Y; Yanagisawa, D; Inden, M; Takata, K; Tsuchiya, D; Kawasaki, T; Taniguchi, T; Shimohama, S. Recovery of focal brain ischemia-induced behavioral dysfunction by intracerebroventricular injection of microglia. J. Pharmacol. Sci 2005, 97, 289–293. [Google Scholar]
- Takahashi-Niki, K; Niki, T; Taira, T; Iguchi-Ariga, SMM; Ariga, H. Reduced anti-oxidative stress activities of DJ-1 mutants found in Parkinson’s disease patients. Biochem. Biophys. Res. Commun 2004, 320, 389–397. [Google Scholar]






© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yamane, K.; Kitamura, Y.; Yanagida, T.; Takata, K.; Yanagisawa, D.; Taniguchi, T.; Taira, T.; Ariga, H. Oxidative Neurodegeneration Is Prevented by UCP0045037, an Allosteric Modulator for the Reduced Form of DJ-1, a Wild-Type of Familial Parkinson’s Disease-Linked PARK7. Int. J. Mol. Sci. 2009, 10, 4789-4804. https://doi.org/10.3390/ijms10114789
Yamane K, Kitamura Y, Yanagida T, Takata K, Yanagisawa D, Taniguchi T, Taira T, Ariga H. Oxidative Neurodegeneration Is Prevented by UCP0045037, an Allosteric Modulator for the Reduced Form of DJ-1, a Wild-Type of Familial Parkinson’s Disease-Linked PARK7. International Journal of Molecular Sciences. 2009; 10(11):4789-4804. https://doi.org/10.3390/ijms10114789
Chicago/Turabian StyleYamane, Koichiro, Yoshihisa Kitamura, Takashi Yanagida, Kazuyuki Takata, Daijiro Yanagisawa, Takashi Taniguchi, Takahiro Taira, and Hiroyoshi Ariga. 2009. "Oxidative Neurodegeneration Is Prevented by UCP0045037, an Allosteric Modulator for the Reduced Form of DJ-1, a Wild-Type of Familial Parkinson’s Disease-Linked PARK7" International Journal of Molecular Sciences 10, no. 11: 4789-4804. https://doi.org/10.3390/ijms10114789