Expression of Mipu1 in Response to Myocardial Infarction in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Acute Myocardial Infarction
2.2. RNA Extraction and cDNA Synthesis
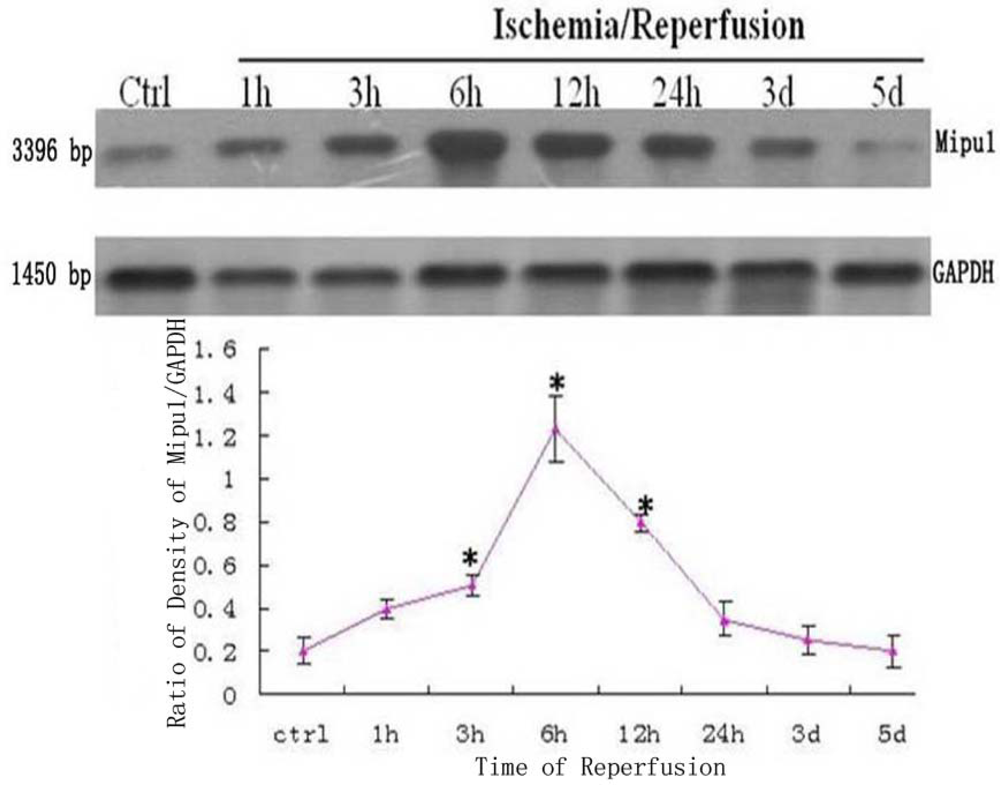
2.3. Northern Blotting
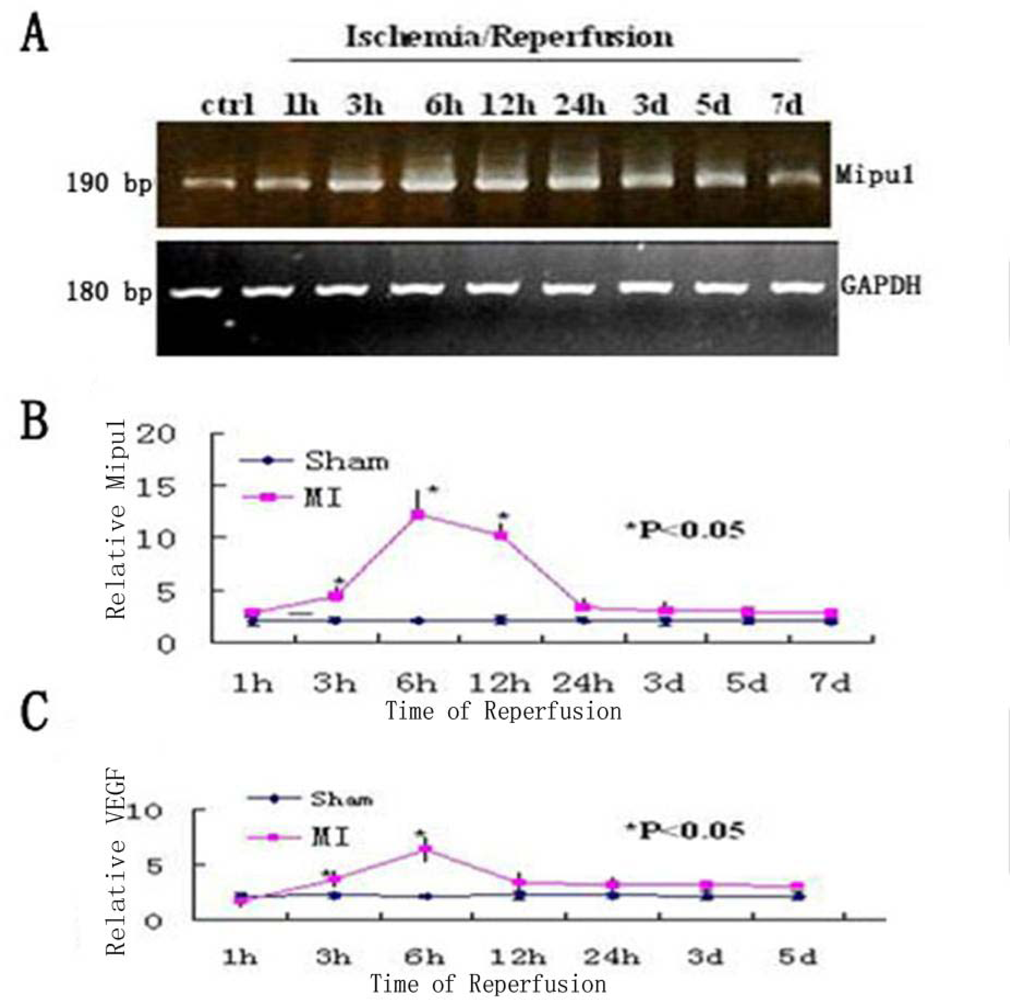
2.4. Quantitative Real-Time RT-PCR
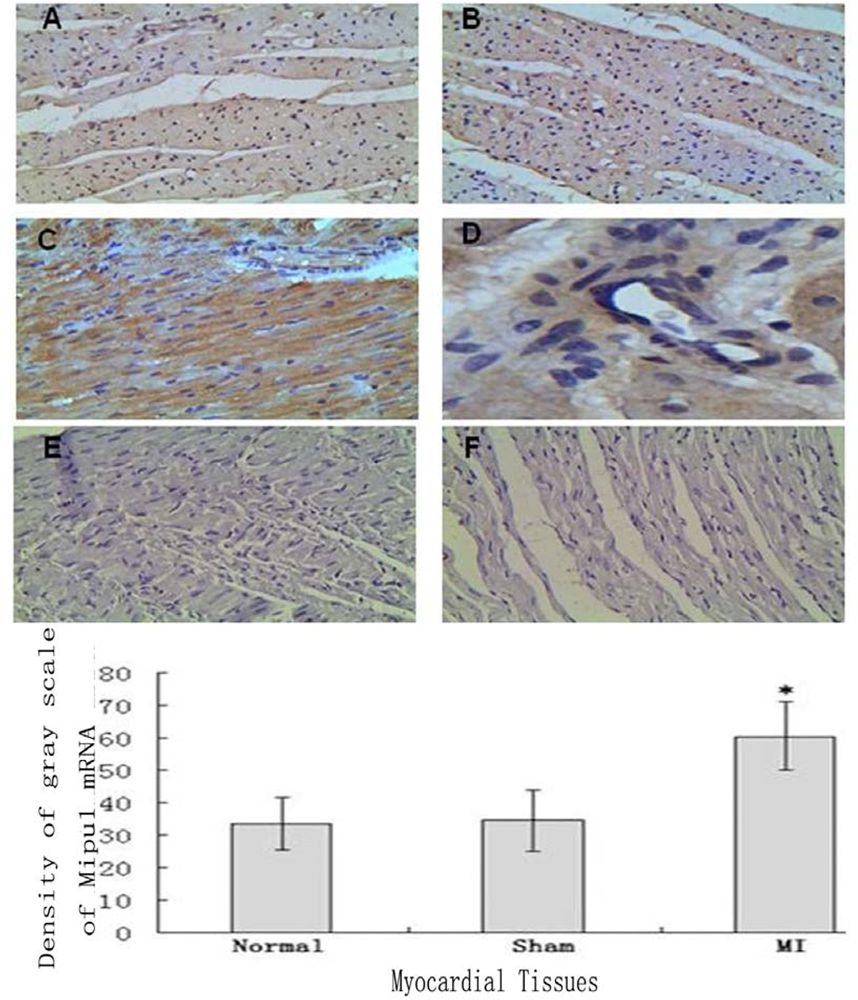
2.5. In Situ Hybridization
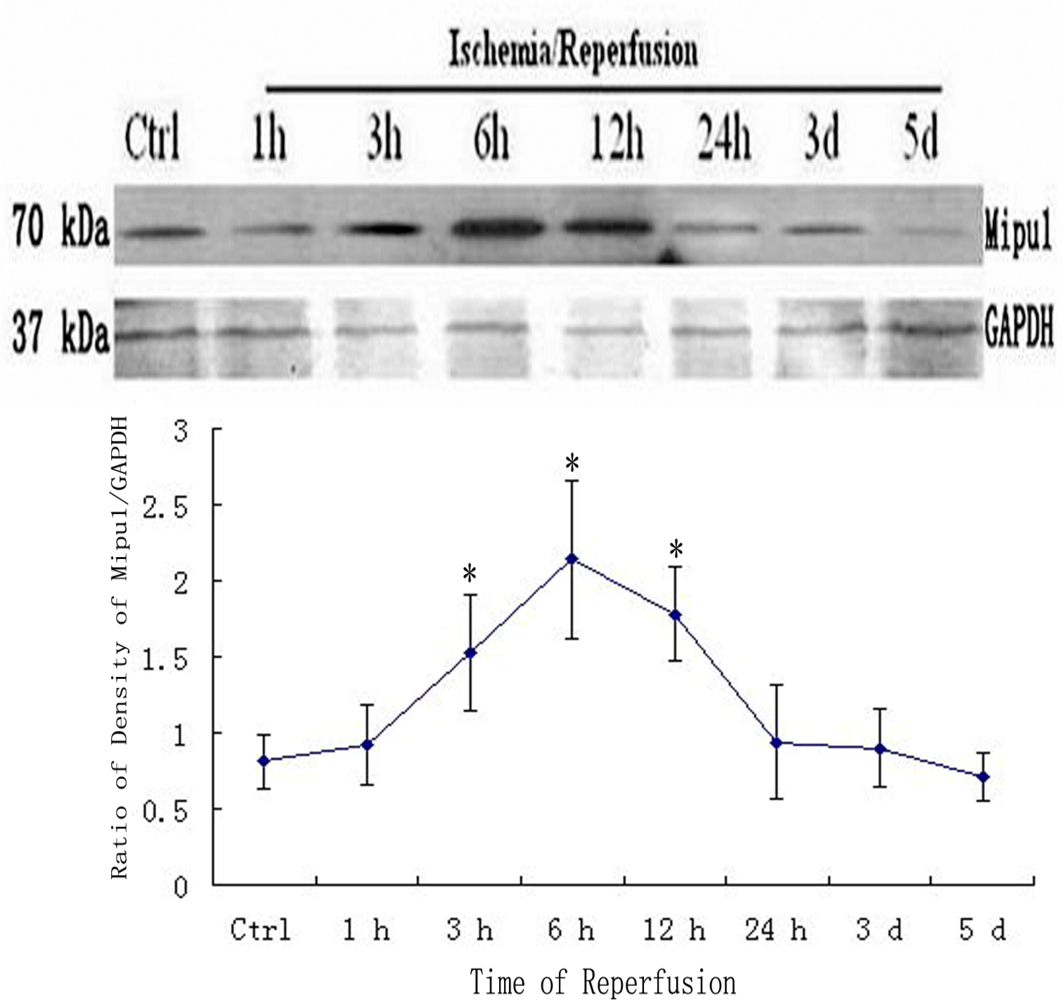
2.6. Western Blotting
2.7. Immunohistochemical Staining
2.8. Fluorescent Immunocytochemistry Study of the Sub-cellular Localization of the Mipu1 Protein in H9c2 Myogenic Cells and Expression after the Cell Treatment with H2O2
2.9. Statistical Analysis
3. Results
3.1. Northern Blotting
3.2. Quantitative Real-Time RT-PCR Analysis
3.3. In Situ Hybridization
3.4. Western blotting
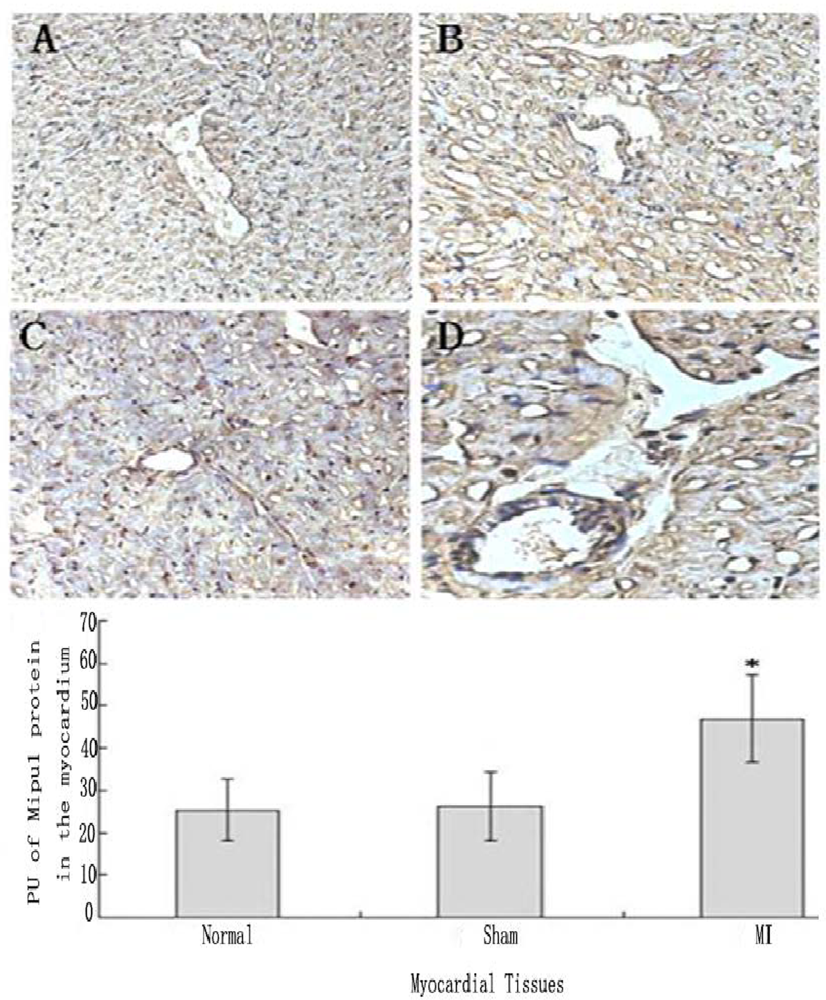
3.5. Tissue Immunohistochemistry
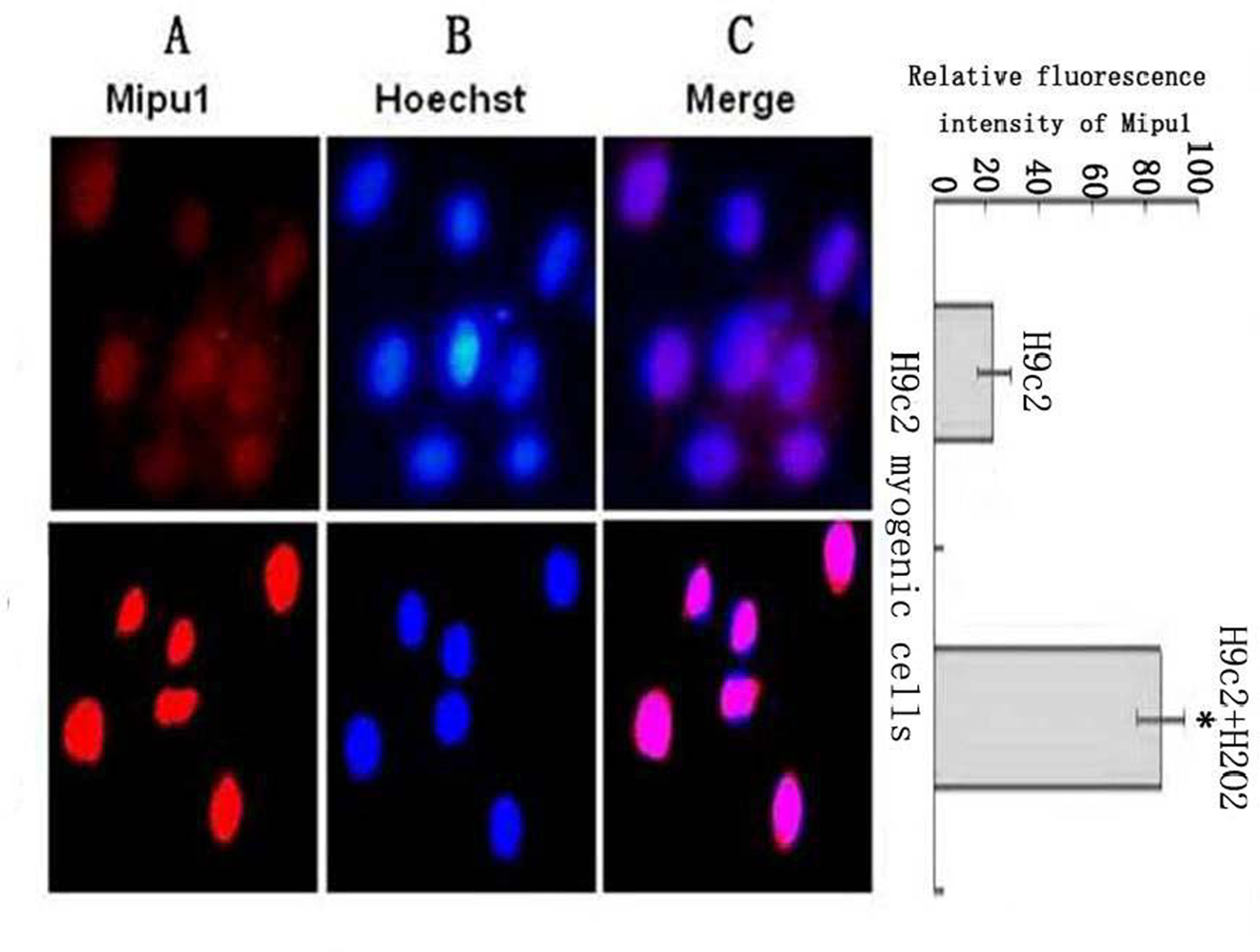
3.6. Fluorescent Immunocytochemistry Shows the Sub-cellular Localization of the Mipu1 Protein in H9c2 Myogenic Cells and Expression after Cell Treatment with H2O2
4. Discussion
5. Conclusions
Acknowledgments
References
- Plumier, JC; Robertson, HA; Currie, RW. Differential accumulation of mRNA for immediate early genes and heat shock genes in heart after ischaemic injury. J. Mol. Cell. Cardiol 1996, 28, 1251–1260. [Google Scholar]
- Nelson, DP; Wechsler, SB; Miura, T; Staggc, A; Newburger, JW; Mayer, JE, Jr; Neufeld, EJ. Myocardial immediate early gene activation after cardiopulmonary bypass with cardiac ischemia-reperfusion. Ann. Thorac. Surg 2002, 73, 156–162. [Google Scholar]
- Suzuki, K; Sawa, Y; Kagisaki, K; Taketani, S; Ichikawa, H; Kaneda, Y; Matsuda, H. Reduction in myocardial apoptosis associated with overexpression of heat shock protein 70. Basic Res. Cardiol 2000, 95, 397–403. [Google Scholar]
- Nidorf, SM; Siu, SC; Galambos, G; Weyman, AE; Picard, MH. Benefit of late coronary reperfusion on ventricular morphology and function after myocardial infarction. J. Am. Coll. Cardiol 1993, 21, 683–691. [Google Scholar]
- Yanagisawa-Miwa, A; Uchida, Y; Nakamura, F; Tomaru, T; Kido, H; Kamijo, T; Sugimoto, T; Kaji, K; Utsuyama, M; Kurashima, C; et al. Salvage of infarcted myocardiumby angiogenic action of basic fibroblast growth factor. Science 1992, 257, 1401–1403. [Google Scholar]
- Battler, A; Scheinowitz, M; Bor, A; Hasdai, D; Vered, Z; DiSegni, E; Varda-Bloom, N; Nass, D; Engelberg, S; Eldar, M; et al. Intracoronary injection of basic fibroblast growth factor enhances angiogenesis in infarcted swine myocardium. J. Am. Coll. Cardiol 1993, 22, 2001–2006. [Google Scholar]
- Li, J; Brown, LF; Hibberd, MG; Grossman, JD; Morgan, JP; Simons, M. VEGF, flk-1, and flt-1 expression in a rat myocardial infarction model of angiogenesis. Amer. J. Physiol 1996, 270, H1803–H1811. [Google Scholar]
- Yuan, C; Zhang, HL; Liu, Y; Wang, QP; Xiao, XZ. Cloning and characterization of a new gene Mip1 up-regulated during myocardial ishemia-reperfusion. Prog. Biochem. Biophys 2004, 31, 231–236. [Google Scholar]
- Jiang, L; Tang, DL; Wang, KK; Zhang, HL; Yuan, C; Duan, DY; Xiao, XZ. Functional analysis of a novel KRAB/C2H2 zinc finger protein Mipu1. Biochem. Biophy. Res. Commun 2007, 356, 829–835. [Google Scholar]
- Shen, H. Quantitative method studies of immunohistochemistry staining. Chin. J. Histochem. Cytochem 1995, 4, 89–92. [Google Scholar]
- Dion, S; Demattéi, MV; Renault, S. Zinc finger proteins: tools for site-specific correction or modification of the genome. Med. Sci. (Paris) 2007, 23, 834–839. [Google Scholar]
- Drummond, IA; Madden, SL; Rohwer-Nutter, P; Bell, GI; Sukhatme, VP; Rauscher, FJ. Repression of the insulin-like growth factor II gene by the Wilms tumor suppressor WT1. Science 1992, 257, 674–678. [Google Scholar]
- Möröy, T; Zeng, H; Jin, J; Schmid, KW; Carpinteiro, A; Gulbins, E. The zinc finger protein and transcriptional repressor Gfi1 as a regulator of the innate immune response. Immunobiology 2008, 213, 341–352. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | |
|---|---|---|
| Mipu1 | Sence | 5’-ATGCCTGCAGCCCGAGGGAAATC-3’ |
| Antisence | 5’- CGATGATATTTGGCCTCCGGCAGGC-3’ | |
| VEGF | Sence | 5’-TCTTCAAGCCATCCTGTGT-3’ |
| Antisence | 5’-CTTTCTTTGGTCTGCATTC-3’ | |
| GAPDH | Sence | 5’-AACACAGTCCATGCCATCAC-3’ |
| Antisence | 5’-TCCACCACCCTGTTGCTGTA-3’ |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, G.; Zuo, X.; Liu, J.; Jiang, L.; Liu, Y.; Zheng, Y.; Zhang, B.; Xiao, X. Expression of Mipu1 in Response to Myocardial Infarction in Rats. Int. J. Mol. Sci. 2009, 10, 492-506. https://doi.org/10.3390/ijms10020492
Wang G, Zuo X, Liu J, Jiang L, Liu Y, Zheng Y, Zhang B, Xiao X. Expression of Mipu1 in Response to Myocardial Infarction in Rats. International Journal of Molecular Sciences. 2009; 10(2):492-506. https://doi.org/10.3390/ijms10020492
Chicago/Turabian StyleWang, Guiliang, Xiaoxia Zuo, Junwen Liu, Lei Jiang, Ying Liu, Yazhu Zheng, Bin Zhang, and Xianzhong Xiao. 2009. "Expression of Mipu1 in Response to Myocardial Infarction in Rats" International Journal of Molecular Sciences 10, no. 2: 492-506. https://doi.org/10.3390/ijms10020492