Relationships among the A Genomes of Triticum L. Species as Evidenced by SSR Markers, in Iran

Abstract
:1. Introduction
2. Results and Discussion
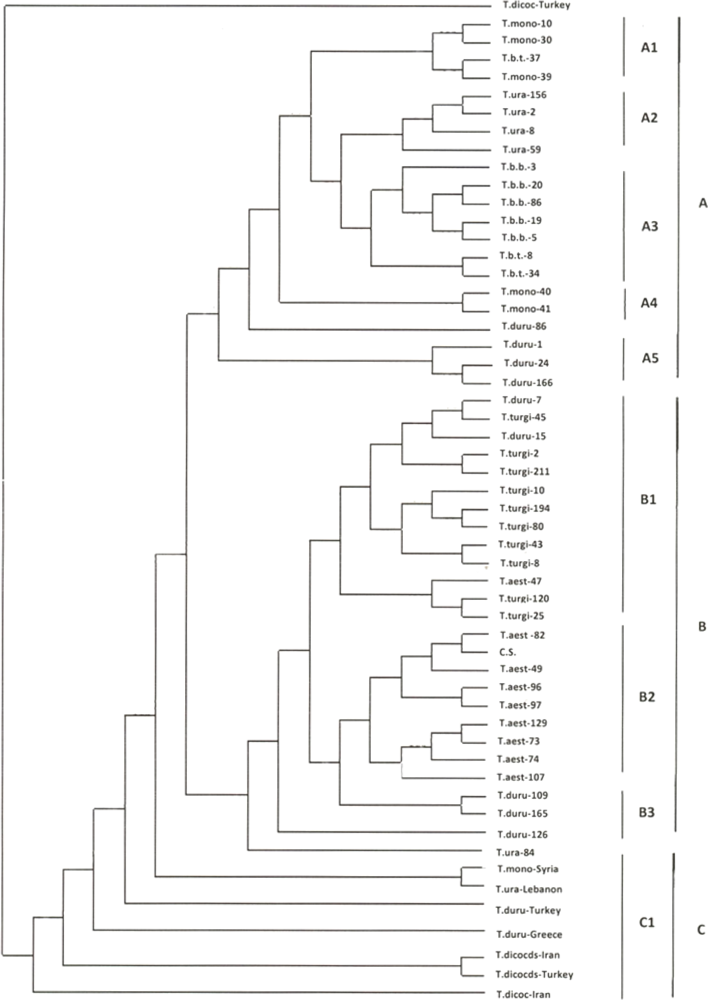
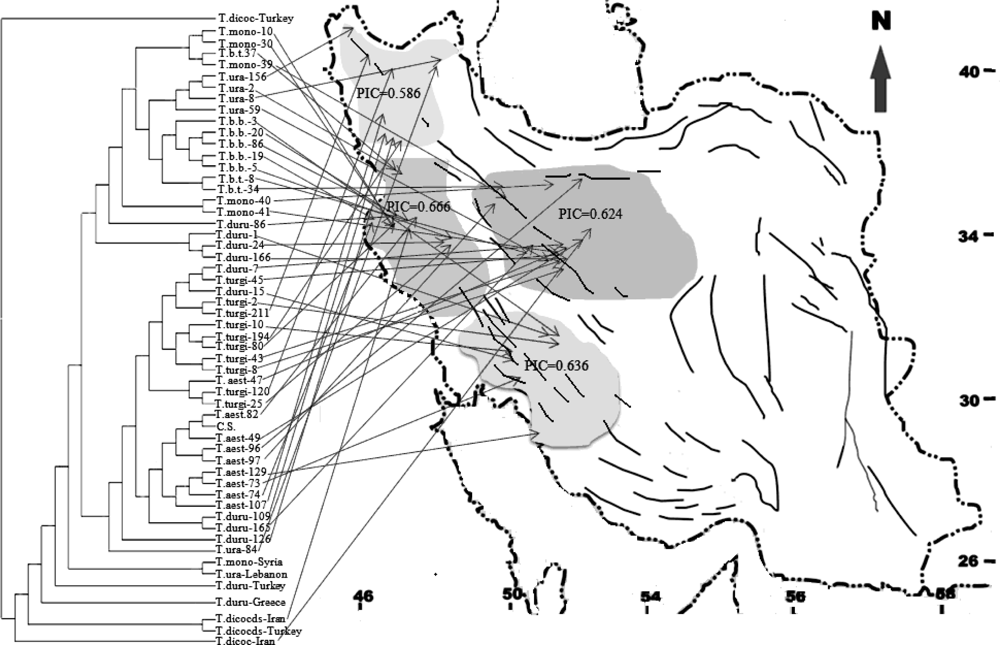
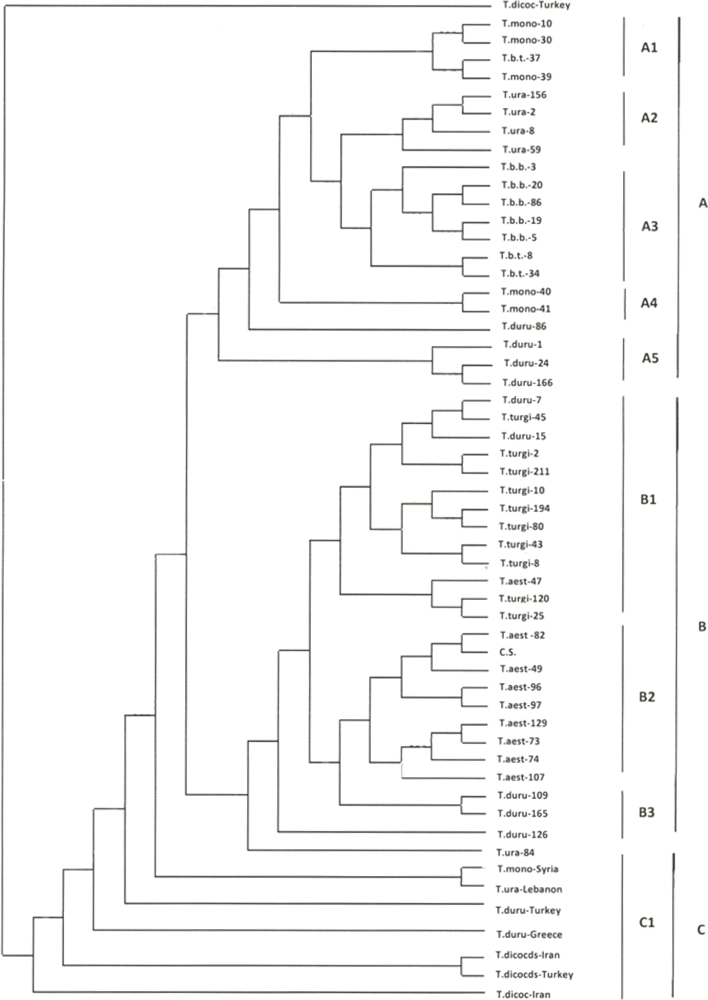
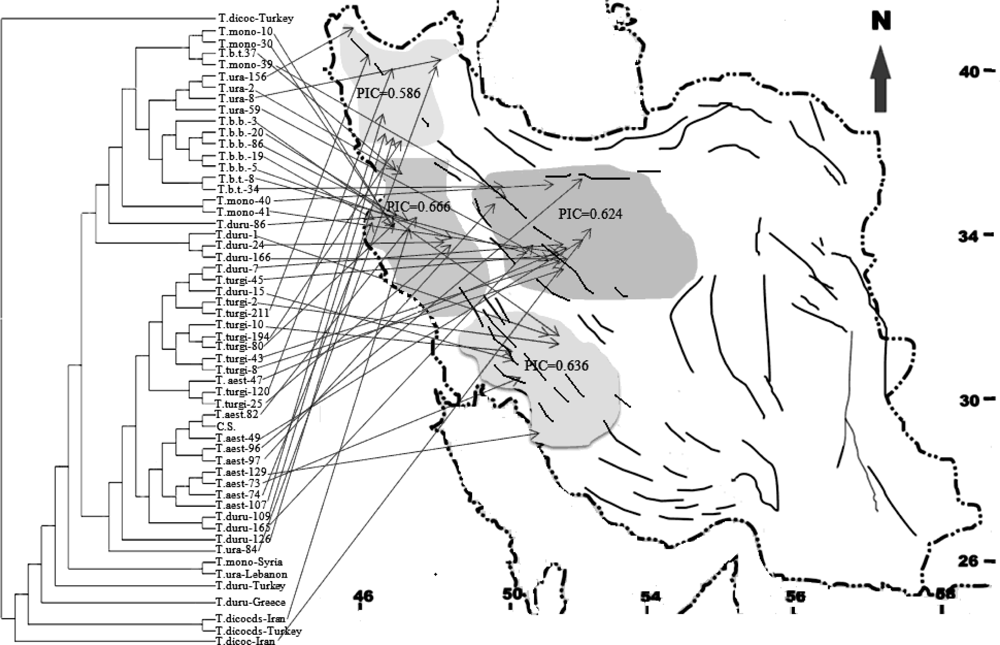
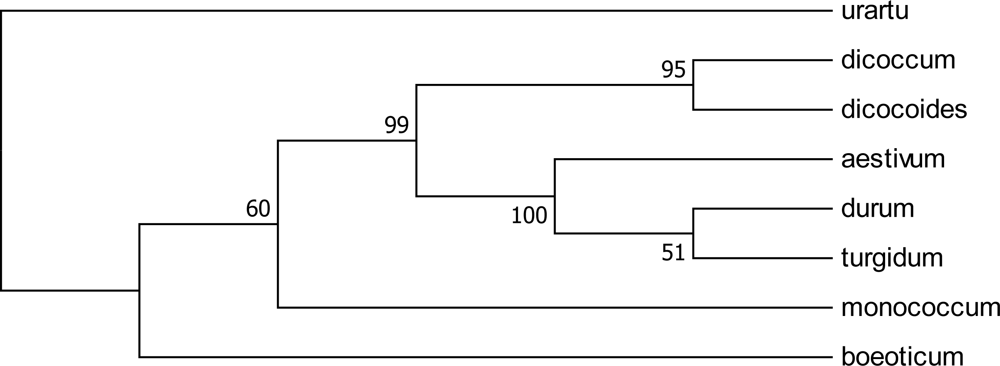
2.1. Genetic Similarity Analysis
2.2. Analysis of Molecular Variance (AMOVA)
3. Experimental Section
Statistical Analyses
4. Conclusion
Acknowledgments
References
- Kihara, H. Cytologische und genetische Studien bei wichtigen Getreidearten mit besonderer Rücksicht auf das Verhalten der Chromosomen und die Sterilität in den Bastarden. Mem. Coll. Sci. Kyoto Imp. Univ 1924, 1, 1–200. [Google Scholar]
- Lilienfeld, F; Kihara, H. Genomanalyse bei Triticum und Aegilops. V. Triticum timopheevi Zhuk. Cytologia 1934, 6, 87–122. [Google Scholar]
- Gill, BS; Appels, R; Botha-Oberholster, A-M; Buell, CR; Bennetzen, JL; Chalhoub, B; Chumley, F; Dvorak, J; Iwanaga, M; Keller, B; et al. A workshop report on whaet genome sequencing: International genome research on wheat consortium. Genetics 2004, 168, 1087–1096. [Google Scholar]
- Harlan, JR. Crops and Man, 2nd ed.; In American Soc. of AgronomyCSSA: Madison, WI, USA, 1992. [Google Scholar]
- Sakamura, T. Kurze Mitteilung über die Chromosomenzahlen und die Verwandtschaftsverhältnisse der Triticum-Arten. Bot. Mag. Tokyo 1918, 32, 151–154. [Google Scholar]
- Zohary, D; Feldman, M. Hybridization between amphiploids and the evolution of polyploids in the wheat (Aegilops-Triticum) group. Evolution 1962, 16, 44–61. [Google Scholar]
- Kimber, G; Feldman, M. Wild Wheat, an Introduction; College of Agriculture, University of Missouri: Columbia, MO, USA, 1987. [Google Scholar]
- Mandy, G. New concept of the origin of Triticum aestivum. Acta Agron. Hung 1970, 19, 413–417. [Google Scholar]
- Allard, RW. Genetic basis of the evolution of adaptedness in plants. Euphytica 1996, 92, 1–11. [Google Scholar]
- Hoisington, D; Khairallah, M; Ribaut, JM; Skovmand, B; Taba, S; Warburton, M. Plant genetic resources: What can they contribute toward increased crop productivity? Proc. Natl. Acad. Sci.USA 1999, 99, 8133–8138. [Google Scholar]
- Rejesus, RM; van Ginkel, M; Smale, M. Wheat breeder’s perspectives of genetic diversity and germplasm use; Wheat special report 40. CYMMIT: Texcoco, Mexico, 1996. [Google Scholar]
- Vavilov, NI. The theory of the origin of cultivated plants after Darwin. Navka (Science) 1940, 2, 55–75. [Google Scholar]
- Tsunewaki, K. Comparative gene analysis of common wheat and its ancestral species. II. Waxiness, growth habit and awnedness. Jpn. J. Bot 1966, 19, 175–229. [Google Scholar]
- Nakai, Y. Isosyme variation in Aegilops and Triticum, IV The origin of the common wheats revealed from the study on esterase isozymes in synthesized wheats. Jpn. J. Genet 1979, 54, 175–189. [Google Scholar]
- Nishikawa, K; Furuta, Y; Wada, T. Genetic studies on alpha-amylase isozymes in wheat. III. Intraspecific variation in Aegilops squarrosa and the birthplace of hexaploid wheat. Jpn. J. Genet 1980, 55, 325–336. [Google Scholar]
- Helguera, M; Khan, IA; Dubcovsky, J. Development of PCR markers for wheat leaf rust resistance gene Lr47. Theor. Appl. Genet 2000, 101, 625–631. [Google Scholar]
- Parker, GD; Chalmers, KJ; Rathjen, AJ; Langridge, P. Mapping loci associated with milling yield in wheat (Triticum aestivum L.). Mol. Breed 1999, 5, 561–568. [Google Scholar]
- Parker, GD; Langridge, P. Development of a STS marker linked to a major locus controlling flour colour in wheat (Triticum aestivum L.). Mol. Breed 2000, 6, 169–174. [Google Scholar]
- Zhao, ZD; Liu, JJ; Dong, JY; Zhang, L; Li, Q. Association between TOM and cooking quality of Chinese noodle. Acta Agronomica Sin 1998, 24, 738–742. [Google Scholar]
- Peng, ZS; Yen, C; Yang, JL. Chromosomal Location of genes for supernumerary spikelet in bread wheat. Euphytica 1998, 103, 109–114. [Google Scholar]
- Zanetti, S; Winzeler, M; Keller, M; Keller, B; Messmer, M. Genetic Analysis of Pre-Harvest Sprouting Resistance in a Wheat 3 Spelt Cross. Crop Sci 2000, 40, 1406–1417. [Google Scholar]
- Ramana, JV; Singh, D. Major genes involved in chlorophyll synthesis in bread wheat (Triticum aestivum L.) Cultivars. Indian J. Genet. Plant Breed 2002, 62, 67–80. [Google Scholar]
- Zheng, YL; Yen, C; Yang, JL. Monosomic analysis for total and sterile floret number per spike in the multispikelet line 10-A of common wheat. Wheat Inf. Service 1993, 77, 25–28. [Google Scholar]
- Vagujfalvi, A; Crosatti, C; Galiba, G; Dubcovsky, J; Cattivelli, L. Two loci on wheat chromosome 5A regulate the differential cold-dependent expression of the cor14b gene in frosttolerant and frost-sensitive genotypes. Mol. Gen. Genet 2000, 263, 194–200. [Google Scholar]
- Davydov, VA. Characterization of the stomatal apparatus in monosomic lines of chinese spring wheat. Genetika (Moskva) 1999, 35, 546–550. [Google Scholar]
- Sutka, J; Galiba, G; Snape, JW. Inheritance of frost resistance in wheat (Triticum aestivum L.). Acta Agron. Hung 1997, 45, 257–263. [Google Scholar]
- Veisz, O; Sutka, J. Role of the chromosome 4B in suppressing frost resistance in winter wheat (Triticum aestivum L.). Cereal Res. Commun 1998, 26, 47–52. [Google Scholar]
- Yuanwu, Z; Yujun, Y. Study on the γ radiosensitivity of monosomic lines of Sumai 3 to radiation from 60Co. Acta Agron. Sin 1995, 21, 429–433. [Google Scholar]
- Hammer, K; Filatenko, AA; Alkhanjari, S; Al-Maskri, A; Buerkert, A. Emmer (Triticum dicoccon Schrank) in Oman. Genet. Resour. Crop Evol 2004, 51, 111–113. [Google Scholar]
- Filatenko, AA; Hammer, K. A new gross morphological variation in the genus Triticum L. Genet Resour. Crop Evol 2007, 54, 231–232. [Google Scholar]
- Tsegaye, S; Tesemma, T. The genetics of some isozyme variants in tetraploid wheats (Triticum turgidum L.). Hereditas 1995, 123, 147–154. [Google Scholar]
- Cheniany, M; Ebrahimzadeh, H; Salimi, A; Niknam, V. Isozyme variation in some populations of wild diploid wheats in Iran. Biochem. Syst. Ecol 2007, 35, 363–371. [Google Scholar]
- Mesfin, A; Frohberg, RC. RFLP Markers Associated with High Grain Protein from Triticum turgidum L. var. dicoccoides Introgressed into Hard Red Spring Wheat. Crop Sci 1999, 39, 508–513. [Google Scholar]
- D’ovidio, R; Tanzarella, OA; Masci, S; Lafiandra, D; Porceddu, E. RFLP and PCR analyses at Gli-1, Gli-2, Glu-1 and Glu-3 loci in cultivated and wild wheats. Hereditas 1992, 116, 79–85. [Google Scholar]
- Figliuolo, G; Perrino, P. Genetic diversity and intra-specific phylogeny of Triticum turgidum L. subsp. dicoccon (Schrank) Thell. revealed by RFLPs and SSRs. Genet. Resour. Crop Evol 2004, 51, 519–527. [Google Scholar]
- Röder, MS; Korzun, V; Wendehake, K; Plashke, J; Tixier, MH; Leory, P; Ganal, MW. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Mohammadi, SA; Khodarahmi, M; Jamalirad, S; Jalal Kamali, MR. Genetic diversity in a collection of old and new bread wheat cultivars from Iran as revealed by simple sequence repeat-based analysis. Ann. Appl. Biol 2009, 154, 67–76. [Google Scholar]
- Gupta, PK; Balyan, HS; Edvards, KJ; Isaac, P; Korzun, V; Roder, M; Gautier, M-F; Joudrier, P; Schlatter, AR; Dubcovskly, J; et al. Genetic mapping of 66 new microsatellite (SSR) loci in bread wheat. Theor. Appl. Genet 2002, 105, 413–422. [Google Scholar]
- Bossolini, E; Krattinger, SG; Keller, B. Development of simple sequence repeat markers specific for the Lr34 resistance region of wheat using sequence information from rice and Aegilops tauschii. Theor. Appl. Genet 2006, 113, 1049–1062. [Google Scholar]
- Bryan, GJ; Collins, AJ; Stephenson, P; Orry, A; Smith, JB; Gale, MD. Isolation and characterization of microsatellites from hexaploid bread wheat. Theor. Appl. Genet 1997, 94, 557–563. [Google Scholar]
- Plaschke, J; Ganal, MW; Röder, MS. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Appl. Genet 1995, 91, 1001–1010. [Google Scholar]
- Stachel, M; Lelley, T; Grausgruber, H; Vollmann, J. Application of microsatellites in wheat (Triticum aestivum L.) for studying genetic differentiation caused by selection for adaptation and use. Theor. Appl. Genet 2000, 100, 242–248. [Google Scholar]
- Huang, SX; Sirikhachornkit, A; Su, XJ; Faris, J; Gill, B; Haselkorn, R; Gornicki, P. Genes encoding plastid acetyl-CoA carboxylase and 3-Phosphoglycerate kinase of the Triticum/Aegilops complex and evolutionary history of polyploid wheat. Proc. Natl. Acad. Sci. USA 2002, 99, 8133–8138. [Google Scholar]
- Eujayl, I; Sorrells, M; Baum, M; Wolters, P; Powell, W. Isolation of EST-derived microsatellite markers for genotyping the A and B genomes of wheat. Theor. Appl. Genet 2002, 104, 339–407. [Google Scholar]
- Buchanan, FC; Adams, LJ; Littlejohn, RP; Maddox, JF; Crawford, AM. Determination of evolutionary relationships among sheep breeds using microsatellites. Genomics 1994, 22, 397–403. [Google Scholar]
- Condit, R; Hubbell, S. Abundance and DNA sequence of two-base repeat regions in tropical tree genomes. Genomics 1991, 34, 66–71. [Google Scholar]
- Kkaya, MS; Bhagwat, AA; Cregan, PB. Length polymorphisms of simple sequences repeat DNA in soybean. Genetics 1992, 132, 1131–1139. [Google Scholar]
- Lagercrantz, U; Ellegren, H; Andersson, L. The abundance of various polymorphic microsatellite motifs differs between plants and vertebrates. Nucleic Acids Res 1993, 21, 1111–1115. [Google Scholar]
- Senior, ML; Heun, M. Mapping maize microsatellites and polymerase chain reaction confirmation of the targeted repeats using a CT primer. Genome 1993, 36, 884–889. [Google Scholar]
- Wu, KS; Tanksley, SD. Abundance, polymorphism and genetic mapping of microsatellites in rice. Mol. Gen. Genet 1993, 241, 225–235. [Google Scholar]
- Bell, CJ; Ecker, JR. Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics 1994, 19, 137–144. [Google Scholar]
- Saghai-Maroof, MA; Soliman, K; Jorgensen, RA; Allard, RW. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosome location and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar]
- Rongwen, J; Akkaya, MS; Bhagwat, AA; Lavi, U; Crecan, PB. The use of microsatellite DNA markers for soybean genotype identification. Theor. Appl. Genet 1995, 90, 43–48. [Google Scholar]
- Liu, ZW; Biyashev, RM; Saghai-Maroof, MA. Development of simple sequence repeat DNA markers and their integration into a barley linkage map. Theor. Appl. Genet 1996, 93, 869–876. [Google Scholar]
- Mörchen, M; Cuguen, J; Michaelis, G; Hänni, C; Saumitou-Laprade, P. Abundance and length polymorphism of microsatellite repeats in Beta vulgaris L. Theor. Appl. Genet 1996, 92, 326–333. [Google Scholar]
- Proven, J; Powell, W; Waugh, R. Microsatellite analysis of relationships within cultivated potato (Solanum tuberosum). Theor. Appl. Genet 1996, 92, 1078–1084. [Google Scholar]
- Szewc-McFadden, AK; Kresovich, S; Bliek, SM; Mrrchell, SE; McFerson, JR. Identification of polymorphic, conserved simple sequence repeats (SSRs) in cultivated Brassica species. Theor. Appl. Genet 1996, 93, 534–538. [Google Scholar]
- Taramino, G; Tingey, S. Simple sequence repeats for germplasm analysis and mapping in maize. Genome 1996, 39, 277–287. [Google Scholar]
- Smulders, MJM; Bredemeijer, G; Rus-Kortekaas, W; Arens, P; Vosman, B. Use of short microsatellites from database sequences to generate polymorphisms among Lycopersicum esculentum cultivars and accessions of other Lycopersicum species. Theor. Appl. Genet 1997, 91, 681–690. [Google Scholar]
- Röder, MS; Plaschke, J; König, SU; Börner, A; Sorrells, ME; Tanksley, SD; Ganal, MW. Abundance, variability and chromosomal location of microsatellites in wheat. Mol. Gen. Genet 1995, 246, 327–333. [Google Scholar]
- Ma, ZQ; Röder, M; Sorrells, ME. Frequencies and sequence characteristics of di-, tetra-nucleotide microsatellites in wheat. Genome 1996, 39, 123–130. [Google Scholar]
- Stephenson, P; Bryan, G; Kirby, J; Collins, A; Devos, K; Busso, C; Gale, M. Fifty new microsatellite loci for the wheat genetic map. Theor. Appl. Genet 1998, 97, 946–949. [Google Scholar]
- Varshney, RK; Prasad, M; Roy, JK; Röder, MS; Balyan, HS; Gupta, PK. Intregated physical maps of 2DL, 6BS and 7DL carrying loci for grain protein content and preharvest sprouling tolerance in bread wheat. Creal Res. Commun 2001, 29, 33–40. [Google Scholar]
- Korzun, V; Röder, MS; Ganal, MW; Worland, AJ; Law, CN. Genetic analysis of the dwarfing gene Rht8 in wheat Part 1. Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.). Theor. Appl. Genet 1998, 96, 1104–1109. [Google Scholar]
- Prasad, M; Varshney, RK; Kumar, A; Balyan, HS; Sharma, PC; Edwards, KJ; Singh, H; Dhaliwal, HS; Roy, JK; Gupta, PK. A microsatellite marker associated with a QTL for grain protein content on chromosome arm 2DL of bread wheat. Theor. Appl. Genet 1999, 99, 341–345. [Google Scholar]
- Roy, JK; Prasad, M; Varshney, RK; Balyan, HS; Blake, TK; Dhaliwal, HS; Singh, H; Edwards, KJ; Gupta, PK. Identification of a microsatellite on chromosomes 6B and a STS on 7D of bread wheat showing an association with preharvest sprouting tolerance. Theor. Appl. Genet 1999, 99, 336–340. [Google Scholar]
- Prasad, M; Varshney, RK; Roy, JK; Balyan, HS; Gupta, PK. The use of microsatellites for detecting DNA polymorphism, genotype identification and genetic diversity in wheat. Theor. Appl. Genet 2000, 100, 584–592. [Google Scholar]
- McLauchlan, A; Henry, RJ; Issac, PG; Edwards, KJ. Microsatellite analysis in cultivated hexaploid wheat and wild wheat relatives. In Plant Genotyping: The DNA Fingerprinting of Plants; Henry, RJ, Ed.; CABI Publishing, CAB International: Wallingford, UK, 2001; pp. 147–159. [Google Scholar]
- Huang, XQ; Börner, A; Röder, MS; Ganal, MW. Assessing genetic diversity of wheat (Triticum aestivum L.) germplasm using microsatellite markers. Theor. Appl. Genet 2002, 105, 699–707. [Google Scholar]
- Zhang, XY; Li, GW; Wang, LF; Wang, HM; You, GX; Dong, YS. An estimation of the minimum number of SSR alleles needed to reveal genetic relationships in wheat varieties. I. Information from large-scale planted varieties and cornerstone breeding parents in Chinese wheat improvement and production. Theor. Appl. Genet 2002, 106, 112–117. [Google Scholar]
- McFadden, ES; Sears, ER. The origin of Triticum spelta and its free-threshing hexaploid relatives. J. Hered 1966, 37, 81–89. [Google Scholar]
- Rahiminegad, MR; Kharazian, N. Illustration of and a key to the genus Triticum L. in Iran. Wheat Inf. Service 2005, 99, 29–34. [Google Scholar]
- Kihara, H. The diversity of DD analyzer, one of the ancestors of common wheat (preliminary reports). Aric. Hort 1944, 19, 889–890. [Google Scholar]
- Johnson, BL. Identification of the apparent B-genome donor of wheat. Can. J. Genet. Cytol 1975, 17, 21–39. [Google Scholar]
- Tsunekawi, K; Ebona, K. Production of near-isogenic lines of common wheat for glaucousness and genetic basis of this trait clarified by their use. Genes Genet. Syst 1999, 74, 33–41. [Google Scholar]
- Rogers, SO; Bendich, AJ. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol 1985, 5, 69–76. [Google Scholar]
- Jackson, JA; Matthews, D. Modified inter-simple sequence repeat PCR protocol for use in conjunction with the Li-Cor gene ImagIR(2) DNA analyzer. Biotechniques 2000, 28, 914–917. [Google Scholar]
- Liu, K; Muse, SV. PowerMarker: Integrated analysis enviornment for genetic marker data. Bioinformatic 2005, 21, 2128–2129. [Google Scholar]
- Jaccard, P. Novelles rescherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat 1908, 44, 223–270. [Google Scholar]
- Rohlf, FJ. NTSYS-PC numerical taxonomy and multivariate analysis system, version 2.02; Exter Publications: Setauket, NY, USA, 1998. [Google Scholar]
- Nei, M. Genetic distance between populations. Am. Nat 1972, 106, 283–293. [Google Scholar]
- Excoffier, L; Smouse, PE; Quattro, JM. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–791. [Google Scholar]
- Gill, BS; Kimber, G. Giemsa C-banding and the evolution of wheat. Proc. Natl. Acad. Sci. USA 1974, 71, 4086–4090. [Google Scholar]
- Dvořák, J. The relationships between the genomes of Triticum urartu and the A and B genomes of Triticum aestivum. Can. J. Genet. Cytol 1976, 18, 371–377. [Google Scholar]
- Wicker, T; Yahiaui, N; Guyot, R; Schlagenhauf, E; Liu, Z; Dubcovsky, J; Keller, B. Rapid genome divergence at orthologous low molecular weight glutenin loci of the A and Am genomes of wheat. Plant Cell 2003, 15, 1186–1197. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Marker | Chr. Loc. | Ann. Temp. | Allele Fr. | Allele No | HE | HO | PIC |
|---|---|---|---|---|---|---|---|
| gwm-601 | 4A | 60 | 0.37 | 15 | 0.7 | 0.85 | 0.66 |
| gwm-135 | 1A | 60 | 0.28 | 14 | 0.84 | 0.66 | 0.83 |
| gwm-71 | 2A | 60 | 0.22 | 18 | 0.86 | 0.86 | 0.85 |
| gwm-666 | 1A, 3A, 5A, 7A | 60 | 0.22 | 22 | 0.86 | 0.98 | 0.84 |
| gwm-311 | 2A, 2B, 6B | 60 | 0.23 | 12 | 0.86 | 0.27 | 0.85 |
| gwm-359 | 2A | 55 | 0.23 | 14 | 0.86 | 0.75 | 0.85 |
| gwm-512 | 2A | 60 | 0.29 | 5 | 0.75 | 0.12 | 0.70 |
| gwm-372 | 2A | 60 | 0.23 | 14 | 0.88 | 0.24 | 0.87 |
| gwm-391 | 3A | 55 | 0.22 | 18 | 0.79 | 0.81 | 0.77 |
| gwm-757 | 3A | 60 | 0.27 | 14 | 0.84 | 0.74 | 0.83 |
| gwm-155 | 3A | 60 | 0.34 | 8 | 0.77 | 0 | 0.74 |
| gwm-291 | 5A | 60 | 0.41 | 16 | 0.77 | 0.59 | 0.74 |
| gwm-494 | 6A, 4A, 3A, 1B | 60 | 0.34 | 12 | 0.77 | 0.87 | 0.74 |
| gwm-427 | 6A | 50 | 0.13 | 20 | 0.92 | 0.24 | 0.92 |
| gwm-635 | 7A, 7B, 7D | 60 | 0.20 | 11 | 0.86 | 0.63 | 0.85 |
| gwm-332 | 7A | 60 | 0.23 | 12 | 0.85 | 0.67 | 0.84 |
| gwm-296 | 2A, 2D, 7D | 55 | 0.14 | 18 | 0.78 | 0.49 | 0.76 |
| gwm-471 | 7A, 7B | 60 | 0.26 | 12 | 0.85 | 0.39 | 0.84 |
| gwm-260 | 7A | 55 | 0.21 | 13 | 0.87 | 0.83 | 0.86 |
| gwm-459 | 6A | 55 | 0.46 | 9 | 0.73 | 0.25 | 0.71 |
| gwm-179 | 5A | 55 | 0.31 | 5 | 0.77 | 0.62 | 0.74 |
| gwm-382 | 2A, 2B, 2D | 60 | 0.27 | 15 | 0.86 | 0.26 | 0.85 |
| gwm-205 | 5A, 5D | 60 | 0.23 | 19 | 0.9 | 0.8 | 0.89 |
| gwm-136 | 1A | 60 | 0.41 | 6 | 0.68 | 0.5 | 0.63 |
| wmc-104 | 1A, 6B | 55 | 0.44 | 10 | 0.74 | 0.22 | 0.72 |
| barc-56 | 5A | 55 | 0.32 | 15 | 0.78 | 0.5 | 0.75 |
| barc-151 | 5A, 7A | 55 | 0.19 | 13 | 0.88 | 0.14 | 0.87 |
| cfa-2086 | 2A | 60 | 0.19 | 17 | 0.86 | 0.46 | 0.85 |
| cfa-2028 | 7A | 55 | 0.33 | 9 | 0.75 | 0.72 | 0.72 |
| cfa-2262 | 3A | 55 | 0.22 | 13 | 0.77 | 0.2 | 0.74 |
| cfa-2263 | 2A | 60 | 0.17 | 11 | 0.88 | 0.25 | 0.87 |
| Mean | 0.29 | 12.8 | 0.79 | 0.49 | 0.77 | ||
| Sum | 410 | ||||||
| Groups | Species | Genetic Similarity |
|---|---|---|
| diplo & diplo | T. monococcum & T.boeoticum | 0.89 |
| T. monococcum & T. urartu | 0.90 | |
| T.boeoticum & T. urartu | 0.90 | |
| tetra & tetra | T. durum & T. turgidum | 0.86 |
| T. durum & T. dicoccum | 0.79 | |
| T. durum & T. dicoccoides | 0.78 | |
| T. turgidum & T. dicoccum | 0.79 | |
| T. turgidum & T. dicoccoides | 0.78 | |
| T. dicoccum & T. dicoccoides | 0.70 | |
| diplo & tetra | T. monococcum & T. durum | 0.85 |
| T. monococcum & T. turgidum | 0.66 | |
| T. monococcum & T. dicoccum | 0.74 | |
| T.monococcum&T.dicoccoides | 0.74 | |
| T. boeoticum & T. durum | 0.82 | |
| T. boeoticum & T. turgidum | 0.65 | |
| T. boeoticum & T. dicoccum | 0.66 | |
| T. boeoticum & T. dicoccoides | 0.69 | |
| T. urartu & T. durum | 0.84 | |
| T. urartu & T. turgidum | 0.64 | |
| T. urartu & T. dicoccum | 0.75 | |
| T. urartu & T. dicoccoides | 0.76 | |
| diplo & hexa | T. monococcum & T. aestivum | 0.77 |
| T. boeoticum & T. aestivum | 0.72 | |
| T. urartu & T. aestivum | 0.72 | |
| tetra & hexa | T. durum & T. aestivum | 0.89 |
| T. turgidum & T. aestivum | 0.86 | |
| T. dicoccum & T. aestivum | 0.64 | |
| T. dicoccoides & T. aestivum | 0.67 |
| Source of variation | d.f | Sum of squares | Mean of squares | Percentage of variation | Variance components | P-value |
|---|---|---|---|---|---|---|
| Among Ploidy levels (groups) | 2 | 235.865 | 117.932 | 7.00 | 2.30879 | <0.001 |
| Among species Within groups | 5 | 286.738 | 57.347 | 17.44 | 5.75630 | <0.001 |
| Among accessions | 47 | 1172.161 | 24.939 | 75.56 | 24.93959 | 0.10948 ± 0.00939 |
| Total | 54 | 1694.764 | 200.218 | 100 | 33.00468 | |
| Species | Collection label | Genome | Locality and altitude (m) |
|---|---|---|---|
| T. monococcum | T. mono-30 | A | Kermanshah, Gardaneh Reno (1480) |
| T. mono -10 | A | Kordestan, 3 km to Saghez (1620) | |
| T. mono -41 | A | Isfahan, Semirom to yasooj (2100) | |
| T. mono -39 | A | Arak to Malayer (2020) | |
| T. mono -40 | A | Tehran, Taleghan valley (1850) | |
| T. mono Syria | A | Provided by Institute of Plant Biology of the University of Zurich | |
| T.boeoticum subsp. thaodar | T.b.t.-37 | A | Kordestan, 5 km after Jenan to Saghez (1770) |
| T.b.t.-8 | A | Chaharmahal Bakhtiari, Shahr-e-Kord, Shapoorabad to Jooneghan (2090) | |
| T.b.t.-34 | A | Arak 15 km to Malayer (1840) | |
| T.boeoticum subsp. boeoticum | T.b.b.-19 | A | Ilam to Kermanshah, Gardaneh Reno (1370) |
| T.b.b.-5 | A | Lorestan, 35 km to Khoramabad from Malavi (1100) | |
| T.b.b.-20 | A | Kermanshah 10 km to Harsin (1330) | |
| T.b.b.-86 | A | Kermanshah to Kamyaran (1340) | |
| T.b.b.-3 | A | Kohkiloye & Boyerahmad,Yasooj, Amirabad (1650) | |
| T.urartu | T.ura-156 | A | West Azarbaijan, Makoo (1580) |
| T.ura-84 | A | Ardabil (1320) | |
| T.ura-2 | A | Kordestan, 10 km Saghez from Asadabad (1440) | |
| T.ura-8 | A | Aradbil, 10 km to Kaghazkanan (1349) | |
| T.ura-59 | A | Chaharmahal Bakhtiari, between Gandoman and Lordegan (2080) | |
| T.ura-Lebanon | A | Provided by Institute of Plant Biology of the University of Zurich | |
| T.durum | T.duru-86 | AB | Kermanshah, Kamyaran (1440) |
| T.duru-24 | AB | Lorestan, Malavi toward Khoram Abad (1200) | |
| T.duru-166 | AB | Chahar mahal Bakhtiari, DoAb Samsami (2000) | |
| T.duru-1 | AB | Kohkiloye & Boyerahmad (990) | |
| T.duru-165 | AB | Chahar mahal Bakhtiari, near Chaghakhor lake (2190) | |
| T.duru-109 | AB | West Azarbaijan, Sardasht to Baneh (1050) | |
| T.duru-15 | AB | Khoosestan, Haftgel to Masjed Soleiman (550) | |
| T.duru-126 | AB | Kordestan, Alamoot 6 Km (1660) | |
| T.duru-7 | AB | Chahar mahal Bakhtiari, Borojen to Izeh (2190) | |
| T.duru-Turky | AB | Provided by Institute of Plant Biology of the University of Zurich | |
| T.duru-Greece | AB | Provided by Institute of Plant Biology of the University of Zurich | |
| T. turgidum | T.turgi-211 | AB | West Azarbaijan, Khoi (1110) |
| T.turgi-45 | AB | Chahar mahal Bakhtiari, Bazoft (2190) | |
| T.turgi-2 | AB | Kohkiloye & Boyerahmad, Yasooj (2880) | |
| T.turgi-43 | AB | Chahar mahal Bakhtiari, Bazoft, Morez valley (2000) | |
| T.turgi-8 | AB | Chahar mahal Bakhtiari, Borojen to Izeh (2190) | |
| T.turgi-10 | AB | Khoosestan, Izeh (900) | |
| T.turgi-194 | AB | Kordestan, between Sanandaj and Saghez (1595) | |
| T.turgi-80 | AB | Kermanshah, Mahi Dasht (1290) | |
| T.turgi-25 | AB | Lorestan, Malavi toward Khoram Abad (1200) | |
| T.turgi-120 | AB | East Azarbaijan, Ahar (1320) | |
| T. dicoccum | T.dicoc-Turkey | AB | Provided by Institute of Plant Biology of the University of Zurich |
| T.dicoc-Iran (Isfahan) | AB | Provided by Institute of Plant Biology of the University of Zurich | |
| T. dicoccoides | T.dicocds-Iran (Kermanshah) | AB | Provided by Institute of Plant Biology of the University of Zurich |
| T.dicocds-Turkey | AB | Provided by Institute of Plant Biology of the University of Zurich | |
| T. aestivum | T.aest-47 | ABD | Chahar mahal Bakhtiari (2000) |
| T.aest-74 | ABD | Ilam, Do Rahe (1410) | |
| T.aest-129 | ABD | Booshehr, Bandargah to Deilam (17) | |
| T.aest-73 | ABD | Khoosestan, Karkheh (13) | |
| T.aest-97 | ABD | Mlayer toward Arak, 50Km (2010) | |
| T.aest-96 | ABD | Tehran, Firooz kouh1 (700) | |
| T.aest-107 | ABD | West Azarbaijan, Boukan to Mahabad (1290) | |
| T.aest-49 | ABD | Isfahan, Daran (2190) | |
| T.aest-82 | ABD | Kermanshah, Mahi Dasht (1290) | |
| Chinese spring | C.S. | ABD | Provided by Institute of Plant Biology of the University of Zurich |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ehtemam, M.H.; Rahiminejad, M.R.; Saeidi, H.; Tabatabaei, B.E.S.; Krattinger, S.G.; Keller, B. Relationships among the A Genomes of Triticum L. Species as Evidenced by SSR Markers, in Iran. Int. J. Mol. Sci. 2010, 11, 4309-4325. https://doi.org/10.3390/ijms11114309
Ehtemam MH, Rahiminejad MR, Saeidi H, Tabatabaei BES, Krattinger SG, Keller B. Relationships among the A Genomes of Triticum L. Species as Evidenced by SSR Markers, in Iran. International Journal of Molecular Sciences. 2010; 11(11):4309-4325. https://doi.org/10.3390/ijms11114309
Chicago/Turabian StyleEhtemam, Mohammad Hosein, Mohammad Reza Rahiminejad, Hojjatollah Saeidi, Badraldin Ebrahim Sayed Tabatabaei, Simon G. Krattinger, and Beat Keller. 2010. "Relationships among the A Genomes of Triticum L. Species as Evidenced by SSR Markers, in Iran" International Journal of Molecular Sciences 11, no. 11: 4309-4325. https://doi.org/10.3390/ijms11114309